Патент на изобретение №2228352
|
||||||||||||||||||||||||||
(54) СПОСОБ ПОЛУЧЕНИЯ БИОМАССЫ И ЦЕЛЕВЫХ ПРОДУКТОВ СИНТЕЗА С ЗАДАННЫМИ ТЕХНОЛОГИЧЕСКИМИ ПАРАМЕТРАМИ
(57) Реферат: Изобретение относится к микробиологии, биохимии и биотехнологии и может быть использовано для получения клеток микроорганизмов с повышенной устойчивостью к неблагоприятным внешним воздействиям и продуктов жизнедеятельности микроорганизмов. Способ предусматривает определение оптимальных профилей изменения технологических параметров и алгоритмов оперативного управления по физиологическим параметрам биомассы. Физиологические параметры определяют исходя из замеров количества биомассы в процессе культивирования. Технологические параметры определяют по возрастной структуре биомассы для фазы замедленного возраста с помощью математических расчетов. Способ позволяет получать биомассу микроорганизмов и продукты ее жизнедеятельности при такой организации контроля процессов, который обеспечивал бы адекватное отражение реальных значений рассчитываемых технологических параметров при минимизации количества значимых показателей процесса культивирования. 15 ил. Изобретение относится к микробиологии, биохимии и биотехнологии и может быть использовано для получения клеток микроорганизмов с повышенной устойчивостью к неблагоприятным внешним воздействиям и продуктов жизнедеятельности микроорганизмов. К настоящему времени известно много способов получения биомассы и/или продуктов жизнедеятельности составляющих ее микроорганизмов с организацией контроля и управления периодических процессов, где контрольными параметрами являются наборы физиологических переменных, задающих параметры биомассы или параметры продуктов ее жизнедеятельности. Например, в обзоре В.В.Бирюкова, Л.Е.Шнайдера. “Управляемые периодические процессы микробиологического синтеза” [Москва ВНИИСЭНТИ, 1986, стр.12-20] описываются способы получения биомассы и продуктов ее жизнедеятельности с организацией контроля концентраций биомассы и продуктов метаболизма, где в качестве контрольных показателей процессов использовались температура и расход аэрирующего воздуха, при этом контрольным показателем управления процессов являлась скорость дозирования субстратов, а управление самой скоростью дозирования осуществлялось, например, с использованием математических моделей процесса или по косвенным параметрам, определяемым в культуральной среде: дыхательному коэффициенту, углеродному дыхательному коэффициенту и другим физиологическим параметрам. В этой работе приведены также примеры управления процессом по непосредственно измеряемым параметрам – рO2, рН. В книге “Основы биохимической инженерии” [Дж.Бейли, Д.Oллис, т.1, стр.484-569] приведены примеры контроля с использованием математических многочленов, например, уравнений Моно, Теиссье, Мозера, Контуа и ряд других для биомассы или Льюдикина-Пайретта – для продукта. При этом контроль процессов, в той или иной степени, связывается или не связывается авторами с физической логикой процесса, а основным условием контроля является минимальное отклонение от экспериментальных данных. Все это требует поиска или расчета многих, индивидуальных для каждого способа контроля процесса, констант, в отдельных случаях труднообъяснимых, а подчас и необъяснимых с точки зрения здравого смысла. Таким образом, в силу сложности и многокомпонетности процессов, проходящих в популяции клеток, в известных способах для контроля выбираются физиологические показатели значимые только для определенного процесса или их группы, а сами способы не могут быть распространены на все виды процессов, а следовательно, не могут служить в качестве универсального способа получения биомассы с заданными технологическими параметрами и способа контроля за технологическими параметрами продуктов ее жизнедеятельности. Поэтому авторами данной заявки решалась задача получать биомассу микроорганизмов и целевых продуктов синтеза как продуктов ее жизнедеятельности при такой организации контроля процессов, который обеспечивал бы адекватное отражение реальных значений рассчитываемых технологических параметров при минимизации количества значимых показателей контроля. Технической задачей, которая решается при использовании заявляемого способа, является оптимизация управляемых процессов биосинтеза, которая позволяет исключить построение индивидуальных математических моделей, построенных на экспериментальных данных, и возможность получения форм клеток микроорганизмов с заданными технологическими параметрами, например, заведомо устойчивых к неблагоприятным внешним воздействиям окружающей среды, что необходимо при производстве вакцин, и/или контролируемое получение продуктов жизнедеятельности микроорганизмов, например, антибиотиков, полисахарида левана, полиоксимасляной кислоты, этанола, уксусной и масляной кислот. В основе предлагаемого способа заложены теоретические положения об энергетических и материальных затратах на рост биомассы и поддержание ее жизнедеятельности для определения внутренней возрастной структуры биосинтезирующей популяции по критерию “содержание клеток, находящихся в нулевом возрасте” [Pirt S.J. “The maintenance energy of bacteria in growing cultures” Proc.Roy.Soc. London,1965, Vol.163B, p.224-231; Derbyshev V.V., Klykov S.P., Glukhov N.N., Scherbakov G.J. “The development of a population under the limitation by the energy supply” Биотехнология, 2001, №2, c.89-96; И.Г.Минкевич “Уравнения динамики микробной культуры как распределенной системы” Микробная конверсия. Фундаментальные и прикладные аспекты Рига, “Зинатне”, 1990, с.22-36], а существенным отличием заявляемого способа, неизвестным из уровня техники, является выбор в качестве контрольного показателя периодических процессов возрастной структуры клеток, например количества клеток в возрасте, равном нулю, определяемой по общей концентрации биомассы при ее культивировании или накоплении целевых продуктов ее жизнедеятельности, при последующем получении интегральных зависимостей в процессе культивирования микроорганизмов или получении зависимостей скорости синтеза каждого из вышеуказанных продуктов от той категории клеток, которая определяет синтез. Использование в качестве единственного контрольного показателя такого параметра как “количество биомассы X” позволяет определять не только внутреннюю структуру биомассы по критериям “количество клеток в нулевом возрасте – Хstab” и “количество клеток в состоянии деления Хdiv”, но и все необходимые зависимости для синтеза какого-либо продукта или утилизации какого-либо субстрата. Это позволяет отказаться от экспериментального определения динамики изменения количества клеток в нулевом возрасте и делящихся клеток для всех процессов. Если необходима динамика изменения концентраций каких-либо продуктов или субстратов, то при известности механизма биохимических превращений нет необходимости экспериментально получать их в каждом конкретном случае, эксперимент можно заменить расчетом и потом лишь периодически сверяться с опытом для их подтверждения. И наоборот, неизвестные механизмы реакций могут быть исследованы с помощью предлагаемого способа путем нахождения констант скоростей исследуемых реакций. Кроме того, это позволяет также отказаться от расчетов констант вышеупомянутых уравнений, применяемых в настоящее время, и вообще исключить стадию математического моделирования конкретного процесса как один из этапов при разработке нового или усовершенствовании старого микробиологического процесса, т.к. предлагаемый способ формально общим образом определяет все микробиологические процессы синтеза биомассы, продуктов ее жизнедеятельности или утилизации субстратов с учетом констант скоростей утилизации или синтеза и набором исходных дифференциальных уравнений для интегрирования в каждом конкретном случае, что обеспечивает универсальность для исследования и проектирования новых микробиологических процессов и модернизации известных. Для этого при получении биомассы и целевых продуктов синтеза с заданными технологическими параметрами путем процесса глубинного культивирования и организации контроля целевых продуктов синтеза, что предполагает определение оптимальных профилей изменения технологических параметров и алгоритмов оперативного управления по физиологическим параметрам, определяемым исходя из замеров количества биомассы при проведении процесса культивирования, физиологические параметры биомассы в фазе замедленного роста определяют по ее возрастной структуре – содержанию делящихся и неделящихся клеток нулевого возраста, рассчитываемых по уравнениям Xdiv=Xp-(Xp-X1)EXP(-A(t-t1))-Xstab1EXP(A(t-t1)) и Хstab=Хstab1·EXP[A·(t-t1)], где Xdiv – количество делящихся клеток на момент времени t; Хstab – количество клеток в нулевом возрасте на момент времени t; Хр – равновесное количество общей биомассы, когда вся энергия, вносимая в сосуд с субстратами, расходуется только на функции поддержания жизнедеятельности клеток; X1 – количество общей биомассы, в момент начала лимитирования по какому-либо субстрату главного фактора, определяющему наступление и развитие фазы замедленного роста; t – время с начала процесса культивирования; t1 – момент времени, соответствующий X1; А=b/а – удельная скорость замедления роста биомассы, где b – удельные затраты на поддержание жизнедеятельности биомассы, единиц субстрата/единиц биомассы в единицу времени, а – трофический коэффициент, единиц субстрата/единиц биомассы, Хstab1-количество клеток в нулевом возрасте в момент начала лимитирования; а для получения целевых продуктов синтеза в качестве технологического параметра синтеза продукта и утилизации субстрата используют текущее количество биомассы делящихся клеток при этом при синтезе продуктов, определяемом только делящимися клетками, используют соотношение Р=Р1+(kdiv/А){ХрLn[(Хр-Х1)/(Хр-X)]-(Х-X1)[1+Xstab1/(Xp(Xp-X1))]}, при синтезе продукта, определяемом как делящимися клетками, так и клетками, находящимися в нулевом возрасте клеточного цикла, используют соотношение Р=Р1+(kdiv/А){ХрLn[(Хр-Х1)/(Хр-Х)]-(Х-Х1)}+(1/A)(kstab-kdiv)Xstab1(X-X1)/(Хр-Х), а при утилизации субстратов, определяемой делящимися клетками и клетками, находящимися в нулевом возрасте клеточного цикла, используют соотношение S=S1-(kdivs/A){XpLn[(Xp-X1)/(Xp-X)]-(X-Х1)}-(1/А)(kstabs-kdivs)Хstab1(Х-Х1)/(Хр-X), где kdiv, kstab, kdivs, kstabs – константы скоростей биохимических реакций; S, Р, Х – количество субстрата, продукта и общей биомассы соответственно, Р1, S1 – количества субстрата и продукта, соответствующие началу фазы замедленного роста. При этом процесс культивирования микроорганизмов при организации контроля интересующего технологического параметра может осуществляться по одной из известных методик, например: отбором проб через различные промежутки времени с их последующим анализом; прямым подсчетом количества клеток под микроскопом в камере Горяева; по определению сухого веса клеток путем использования специальных методик фильтрования, высушивания и последующего взвешивания; по определению, так называемого, “сырого” веса клеток; по определению спектрофотометрических характеристик суспензии клеток; по высеву определенного разведения полученной пробы на плотные питательные среды с последующим подсчетом числа выросших колоний и определению по этому числу количества живых клеток в исходной анализируемой пробе; а также комбинацией вышеперечисленных методов. Данный способ, разработанный для простых процессов, в полной мере применим и для сложных процессов, классифицированных в соответствии с Дейндоерфером: 1) простые процессы, когда превращение питательных веществ в продукты метаболизма происходит без накопления промежуточных веществ со строго определенной стехиометрией; 2) совместные процессы, когда превращение питательных веществ в продукты метаболизма не сопровождается накоплением промежуточных веществ, но стехиометрия процесса может изменяться; 3) последовательные процессы, когда сначала накапливается интермедиат, а затем он расходуется клетками; ступенчатые процессы – питательные вещества селективно превращаются в продукты метаболизма в каком-либо порядке,и рассматриваемых как совокупность (сумма) простых процессов. Доказательством результативности предлагаемого способа, его универсальности и промышленной применимости являются нижеприводимые примеры. Пример 1 (фиг.1) Получение биомассы факультативно анаэробных бактерий рода Salmonella Биомассу выращивали в ферментере при постоянных температуре, перемешивании, аэрации, т.е. при постоянных условиях массопередачи кислорода в культуральную жидкость, и подаче в реакционный объем сбалансированной питательной среды с постоянной скоростью подачи в фазе замедленного роста. Эта скорость дозирования определяла постоянство величины рН в реакционной смеси или ее незначительные колебания в пределах не более ±0,1 единиц от заданного значения 7,1 и, фактически, задавалась самой культурой. Одновременно контролировали значения рO2 для полной уверенности, что предыдущая доза смеси питательных веществ исчерпалась. По ходу процесса отбирались пробы культуральной жидкости, в которых определялась концентрация клеток путем высевов соответствующих разведений культуры на плотную питательную среду и последующим подсчетом числа выросших колоний. Часть проб подвергалась испытанию в тесте “замораживание-оттаивание” –4 мл культуральной жидкости замораживали при -4  С, а через 1 сутки оттаивали в теплой воде и также высевали на плотную питательную среду и считали число выросших колоний. Клетки, не погибшие после такого теста, считаются устойчивыми, высеваются на плотную питательную среду, давая потомство. Принято считать их находящимися в нулевой фазе клеточного цикла роста, когда мембраны и органеллы не затронуты процессами бинарного деления и, следовательно, сама система в целом более устойчива. Здесь клетки имеют наиболее ярко выраженные свойства криорезистентности. Параметры кривой роста общей биомассы определялись по методике, представленной [Derbyshev V.V., Klykov S.P., Glukhov N.N., Scherbakov G.J “The development of a population under the limitation by the energy supply”. – Биотехнология, 2001, №2, с.89-96].
Сначала определяются по динамике биомассы Хр – теоретически максимально возможная для данного массообмена по кислороду концентрация клеток – когда вся вносимая с энергетическим субстратом энергия расходуется только на нужды поддержания жизнедеятельности, X1 – их концентрация при начале кислородного лимитирования, удельная скорость поддержания А=b/а час-1 или удельная скорость замедления роста биомассы, где, в свою очередь, b – удельные затраты на поддержание жизнедеятельности биомассы, грамм субстрата/грамм биомассы в час, а – трофический коэффициент, грамм субстрата/грамм биомассы, далее по расходу энергетического субстрата определяются а и b, скорость массопередачи, кислорода, время окончания процесса.
Концентрация клеток Х в фазе замедленного роста определялась по итоговому уравнению (1.1)
Х=Хр-(Хр-X1)*EXP[-A*(t-t1)],
где Хр=36,5 миллиардов клеток в 1 мл суспензии – равновесная концентрация клеток, когда вся энергия, вносимая в сосуд с субстратами расходуется только на функции поддержания жизнедеятельности клеток;
X1=12 С, а через 1 сутки оттаивали в теплой воде и также высевали на плотную питательную среду и считали число выросших колоний. Клетки, не погибшие после такого теста, считаются устойчивыми, высеваются на плотную питательную среду, давая потомство. Принято считать их находящимися в нулевой фазе клеточного цикла роста, когда мембраны и органеллы не затронуты процессами бинарного деления и, следовательно, сама система в целом более устойчива. Здесь клетки имеют наиболее ярко выраженные свойства криорезистентности. Параметры кривой роста общей биомассы определялись по методике, представленной [Derbyshev V.V., Klykov S.P., Glukhov N.N., Scherbakov G.J “The development of a population under the limitation by the energy supply”. – Биотехнология, 2001, №2, с.89-96].
Сначала определяются по динамике биомассы Хр – теоретически максимально возможная для данного массообмена по кислороду концентрация клеток – когда вся вносимая с энергетическим субстратом энергия расходуется только на нужды поддержания жизнедеятельности, X1 – их концентрация при начале кислородного лимитирования, удельная скорость поддержания А=b/а час-1 или удельная скорость замедления роста биомассы, где, в свою очередь, b – удельные затраты на поддержание жизнедеятельности биомассы, грамм субстрата/грамм биомассы в час, а – трофический коэффициент, грамм субстрата/грамм биомассы, далее по расходу энергетического субстрата определяются а и b, скорость массопередачи, кислорода, время окончания процесса.
Концентрация клеток Х в фазе замедленного роста определялась по итоговому уравнению (1.1)
Х=Хр-(Хр-X1)*EXP[-A*(t-t1)],
где Хр=36,5 миллиардов клеток в 1 мл суспензии – равновесная концентрация клеток, когда вся энергия, вносимая в сосуд с субстратами расходуется только на функции поддержания жизнедеятельности клеток;
X1=12 109 клеток /мл суспензии – концентрация бактерий, когда наступает кислородное лимитирование, главный фактор, определяющий наступление и развитие фазы замедленного роста;
t – время с начала процесса культивирования (момента посева);
t1=4 часа – время, соответствующее X1;
А=b/а=0,23 час-1 – удельная скорость замедления роста биомассы, где b – удельные затраты на поддержание жизнедеятельности биомассы, грамм субстрата/грамм биомассы в час; а – трофический коэффициент, грамм субстрата/грамм биомассы.
Максимальная удельная скорость при проведении этих процессов составила 109 клеток /мл суспензии – концентрация бактерий, когда наступает кислородное лимитирование, главный фактор, определяющий наступление и развитие фазы замедленного роста;
t – время с начала процесса культивирования (момента посева);
t1=4 часа – время, соответствующее X1;
А=b/а=0,23 час-1 – удельная скорость замедления роста биомассы, где b – удельные затраты на поддержание жизнедеятельности биомассы, грамм субстрата/грамм биомассы в час; а – трофический коэффициент, грамм субстрата/грамм биомассы.
Максимальная удельная скорость при проведении этих процессов составила  mах=0,5 час-1.
По ходу процесса из культуральной жидкости (далее КЖ) отбирались пробы для теста “замораживание-оттаивание”. Динамика клеток, выживших после проведения этого теста, находящихся в нулевом возрасте и называемых далее “устойчивыми”-Хstab”, подчинялась следующему уравнению (1.2):
Хstab=Хstab1·EXP[A·(t-t1)] полученному с учетом уравнения (1.1),
где Хstab – концентрация выживших клеток на момент времени t;
Хstab1=4,5109 – то же в момент начала лимитирования кислородом, остальные параметры описаны выше для общей концентрации биомассы.
Максимальная концентрация клеток, которые могут выживать при проведении вышеописанного теста наступает в момент времени tконечное, который определится по уравнению (1.3)
tконечное=t1+(1/A)ln[Xp/(2Xstab1)+(1/(2A Xst1)) mах=0,5 час-1.
По ходу процесса из культуральной жидкости (далее КЖ) отбирались пробы для теста “замораживание-оттаивание”. Динамика клеток, выживших после проведения этого теста, находящихся в нулевом возрасте и называемых далее “устойчивыми”-Хstab”, подчинялась следующему уравнению (1.2):
Хstab=Хstab1·EXP[A·(t-t1)] полученному с учетом уравнения (1.1),
где Хstab – концентрация выживших клеток на момент времени t;
Хstab1=4,5109 – то же в момент начала лимитирования кислородом, остальные параметры описаны выше для общей концентрации биомассы.
Максимальная концентрация клеток, которые могут выживать при проведении вышеописанного теста наступает в момент времени tконечное, который определится по уравнению (1.3)
tконечное=t1+(1/A)ln[Xp/(2Xstab1)+(1/(2A Xst1))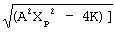 где К=(A2)*(Хst1)*(Хр-Х1) (1.4).
Определенное по этому уравнению время окончания процесса культивирования составило 12,18 часа, что соответствовало действительному времени окончания процесса, когда Xкoнeчнoe=Xstabкoнeчнoe=32,8109 клеток в 1 мл КЖ.
Концентрацию делящихся клеток Хdiv определяли по уравнению
Xdiv=Xp-(Xp-X1)EXP(-A(t-t1))-Хstab 1EXP(A(t-t1)) и Хstab=Хstab1·EXP[A·(t-t1)], (1.5.).
Пример 2 (фиг.2)
Расчет динамики биомассы фотосинтезирующих одноклеточных водорослей Chlorella sp.K
1=5,46 г/л, mах=0,615 сутки-1, А=0,1815 сутки-1, t1=2,5 сутки. 1 г биомассы/литр суспензии=85 миллионов клеток/миллилитр. Расчетная кривая по уравнению X=23,6-18,14·EXP[-0,1815·(t-2,5)] (2.1) для фазы замедленного роста и экспериментальные данные представлены на фиг.2.
Пример 3 (фиг.3 и 4)
Расчет динамик синтеза антибиотиков
а) Пенициллин.
Этот пример одновременно является примером, демонстрирующим возможность определения внутренней структуры биомассы грибов по динамике общего накопления биомассы. Расчет проведен по представленным в Основах биохимической инженерии [Дж.Бейли, Д.Оллис, т.1, стр.547] экспериментальным динамикам накопления продуцента и пенициллина. Расчет накопления продуцента осуществлялся по такой же методике, как и в вышеприведенных примерах. Накопление продукта вторичного метаболизма – пенициллина – по методике авторов, учитывающей динамику делящихся клеток по ходу процесса культивирования, определяемую по уравнению (1.5) и определяющую интегральное накопление продукта с константой скорости синтеза пенициллина kdiv=0,0018 (г/л) пенициллина/(г/л) делящихся клеток в час. Константу определяют по динамике скорости синтеза продукта от концентрации делящихся клеток. Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта, представлены в уравнениях накопления биомассы и пенициллина
X=22,45-14,75·EXP[-0,0286*(t-21)](3.1),
Р=Р1+(kdiv/А)·{ХрLn[(Хр-Х1)/(Хр-Х)]-(Х-Х1)·(1+Xstab1/(Xp-X))}(3.2).
Здесь Хstab1=3,47 г/л может рассчитываться по предлагаемой методике и может быть определено экспериментально.
Р1, определенное экспериментально, составило 0,07 г/л.
б) Грамицидин С (фиг.5 и 6)
Расчет накопления продуцента осуществлялся по такой же методике, как и в вышеприведенных примерах, накопление продукта вторичного метаболизма – грамицидина С проведен согласно предлагаемой методике, учитывающей динамику делящихся клеток по ходу процесса культивирования. Она определяется по уравнению (1.5) и определяет величину константы скорости синтеза грамицидина С kdiv=27,9 (мг грамицидина С/г делящихся клеток в час) и посредством последней – интегральное накопление продукта.
Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта отражены в уравнениях накопления биомассы и грамицидина С, которые, в свою очередь, показаны на соответствующих рисунках.
Здесь Хstab1=0,296 усл.ед./л может рассчитываться по предлагаемой методике и может быть определено экспериментально.
Р1 определено экспериментально и составило 17,7 мг/л.
в) Туримицин (фиг.7 и 8)
Расчет накопления продуцента осуществлялся по такой же методике, как и в вышеприведенных примерах, расчет накопления продукта вторичного метаболизма – туримицина сделан с учетом динамики делящихся клеток по ходу процесса культивирования, определяемой по уравнению (1.5) и определяющей с константой скорости синтеза туримицина С kdiv=0,049 (мг туримицина/г делящихся клеток в час) интегральное накопление продукта.
Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта, отражены в уравнениях накопления биомассы и туримицина, которые, в свою очередь, показаны на соответствующих рисунках.
Здесь Хstab1=1,936 усл.ед./л может быть рассчитан или определен экспериментально. В данном случае наблюдается необходимость введения поправочной константы Р1, которая определена экспериментально для случая туримицина и составила -0,7 мг/л.
Пример 4 (фиг.9 и 10)
Биосинтез высокомолекулярных полимеров оксикислот – полиоксимасляной кислоты (ПОМК)
Накопление продукта вторичного метаболизма ПОМК происходит согласно схеме, учитывающей динамику делящихся клеток по ходу процесса культивирования, определяемую по уравнению (1.5) и динамику (1.2). Эти динамики определяются константой скорости синтеза ПОМК kdiv=88 (%/ед. ОП в сутки) и kstab=45 (%/ед. ОП в сутки).
Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта, отражены в уравнениях накопления биомассы и ПОМК, которые, в свою очередь, показаны на соответствующих рисунках.
Здесь Хstab1=0,126 ед. ОП может рассчитываться по методике и может быть определено экспериментально, В данном случае Р1=15,56% взято из эксперимента.
Пример 5 (фиг.11 и 12)
Синтез микробных полисахаридов и спиртов на примере образования Левана и Этанола бактериями Zymomonas mobilis
Расчет накопления продуцента осуществлялся по такой же методике, как и в вышеприведенных примерах. Накопление продуктов метаболизма – Левана и Этанола происходит по схеме авторов, учитывающей динамику делящихся клеток по ходу процесса культивирования, определяемую по уравнению (1.5) и определяющую с константой скорости синтеза Левана kdiv=1,9455 (г Левана/ед. ОП в час) и этанола kdiv=4,1052 (г Этанола/ед. ОП в час) интегральное накопление продукта.
Х=3,25-1,65ЕХР[-0,063(t-17)], Хр=3,25; Х1=1,6; А=0,065 (1/час).
Максимальная удельная скорость роста=0,063 (1/час), t1=17 час.
Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта отражены в уравнениях накопления биомассы и продуктов, которые, в свою очередь, показаны на соответствующих рисунках.
Здесь Хstab1=0,8 усл.ед ОП может рассчитываться или может быть определено экспериментально. В данном случае Р1=18 г/л для Левана и 42 г/л для спирта. В данном примере были также рассчитаны изменения образования продукта на экспоненциальной фазе роста культуры. Они приведены на фиг.12 пунктирной линией.
Пример 6 (фиг.13)
Здесь Хр=14,3 г/л, Х1=5,33 г/л, Р1=0,02275 г/100 мл, kdivs=0,0014 [(грамм фосфора/100 мл)/(грамм делящихся клеток/литр) в сутки], kstabs=-0,0007 [(грамм фосфора/100 мл)/(грамм клеток в возрасте 0 клеточного цикла/литр)в сутки], А=0,23 (1/сутки), Хstab1=2,49 г/л.
Пример 7(фиг.14 и 15)
Этот пример демонстрирует возможность определения внутренней структуры биомассы Clostridium acetobutilicum по динамике общего накопления биомассы (фиг.14). Расчет проведен по представленным в книге “Основы биохимической инженерии” [Дж.Бейли, Д.Оллис, т.1, стр.551] экспериментальным динамикам накопления продуцента и кислот – уксусной и масляной. Фиг.15 демонстрирует накопление ацетата и бутирата.
Здесь Хр=4,21 г/л, Х1=1,8 г/л, для ацетата Р1=1,4 г/л, для ацетата kdiv=0,4755 [(грамм/литр)/(грамм делящихся клеток/литр) в час], для бутирата kdiv=0,3057 [(грамм/литр)/(грамм делящихся клеток/литр) в час], для бутирата Р1=2 г/л; А=0,173 (1/час), Хstab1=0,88 г/л.
где К=(A2)*(Хst1)*(Хр-Х1) (1.4).
Определенное по этому уравнению время окончания процесса культивирования составило 12,18 часа, что соответствовало действительному времени окончания процесса, когда Xкoнeчнoe=Xstabкoнeчнoe=32,8109 клеток в 1 мл КЖ.
Концентрацию делящихся клеток Хdiv определяли по уравнению
Xdiv=Xp-(Xp-X1)EXP(-A(t-t1))-Хstab 1EXP(A(t-t1)) и Хstab=Хstab1·EXP[A·(t-t1)], (1.5.).
Пример 2 (фиг.2)
Расчет динамики биомассы фотосинтезирующих одноклеточных водорослей Chlorella sp.K
1=5,46 г/л, mах=0,615 сутки-1, А=0,1815 сутки-1, t1=2,5 сутки. 1 г биомассы/литр суспензии=85 миллионов клеток/миллилитр. Расчетная кривая по уравнению X=23,6-18,14·EXP[-0,1815·(t-2,5)] (2.1) для фазы замедленного роста и экспериментальные данные представлены на фиг.2.
Пример 3 (фиг.3 и 4)
Расчет динамик синтеза антибиотиков
а) Пенициллин.
Этот пример одновременно является примером, демонстрирующим возможность определения внутренней структуры биомассы грибов по динамике общего накопления биомассы. Расчет проведен по представленным в Основах биохимической инженерии [Дж.Бейли, Д.Оллис, т.1, стр.547] экспериментальным динамикам накопления продуцента и пенициллина. Расчет накопления продуцента осуществлялся по такой же методике, как и в вышеприведенных примерах. Накопление продукта вторичного метаболизма – пенициллина – по методике авторов, учитывающей динамику делящихся клеток по ходу процесса культивирования, определяемую по уравнению (1.5) и определяющую интегральное накопление продукта с константой скорости синтеза пенициллина kdiv=0,0018 (г/л) пенициллина/(г/л) делящихся клеток в час. Константу определяют по динамике скорости синтеза продукта от концентрации делящихся клеток. Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта, представлены в уравнениях накопления биомассы и пенициллина
X=22,45-14,75·EXP[-0,0286*(t-21)](3.1),
Р=Р1+(kdiv/А)·{ХрLn[(Хр-Х1)/(Хр-Х)]-(Х-Х1)·(1+Xstab1/(Xp-X))}(3.2).
Здесь Хstab1=3,47 г/л может рассчитываться по предлагаемой методике и может быть определено экспериментально.
Р1, определенное экспериментально, составило 0,07 г/л.
б) Грамицидин С (фиг.5 и 6)
Расчет накопления продуцента осуществлялся по такой же методике, как и в вышеприведенных примерах, накопление продукта вторичного метаболизма – грамицидина С проведен согласно предлагаемой методике, учитывающей динамику делящихся клеток по ходу процесса культивирования. Она определяется по уравнению (1.5) и определяет величину константы скорости синтеза грамицидина С kdiv=27,9 (мг грамицидина С/г делящихся клеток в час) и посредством последней – интегральное накопление продукта.
Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта отражены в уравнениях накопления биомассы и грамицидина С, которые, в свою очередь, показаны на соответствующих рисунках.
Здесь Хstab1=0,296 усл.ед./л может рассчитываться по предлагаемой методике и может быть определено экспериментально.
Р1 определено экспериментально и составило 17,7 мг/л.
в) Туримицин (фиг.7 и 8)
Расчет накопления продуцента осуществлялся по такой же методике, как и в вышеприведенных примерах, расчет накопления продукта вторичного метаболизма – туримицина сделан с учетом динамики делящихся клеток по ходу процесса культивирования, определяемой по уравнению (1.5) и определяющей с константой скорости синтеза туримицина С kdiv=0,049 (мг туримицина/г делящихся клеток в час) интегральное накопление продукта.
Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта, отражены в уравнениях накопления биомассы и туримицина, которые, в свою очередь, показаны на соответствующих рисунках.
Здесь Хstab1=1,936 усл.ед./л может быть рассчитан или определен экспериментально. В данном случае наблюдается необходимость введения поправочной константы Р1, которая определена экспериментально для случая туримицина и составила -0,7 мг/л.
Пример 4 (фиг.9 и 10)
Биосинтез высокомолекулярных полимеров оксикислот – полиоксимасляной кислоты (ПОМК)
Накопление продукта вторичного метаболизма ПОМК происходит согласно схеме, учитывающей динамику делящихся клеток по ходу процесса культивирования, определяемую по уравнению (1.5) и динамику (1.2). Эти динамики определяются константой скорости синтеза ПОМК kdiv=88 (%/ед. ОП в сутки) и kstab=45 (%/ед. ОП в сутки).
Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта, отражены в уравнениях накопления биомассы и ПОМК, которые, в свою очередь, показаны на соответствующих рисунках.
Здесь Хstab1=0,126 ед. ОП может рассчитываться по методике и может быть определено экспериментально, В данном случае Р1=15,56% взято из эксперимента.
Пример 5 (фиг.11 и 12)
Синтез микробных полисахаридов и спиртов на примере образования Левана и Этанола бактериями Zymomonas mobilis
Расчет накопления продуцента осуществлялся по такой же методике, как и в вышеприведенных примерах. Накопление продуктов метаболизма – Левана и Этанола происходит по схеме авторов, учитывающей динамику делящихся клеток по ходу процесса культивирования, определяемую по уравнению (1.5) и определяющую с константой скорости синтеза Левана kdiv=1,9455 (г Левана/ед. ОП в час) и этанола kdiv=4,1052 (г Этанола/ед. ОП в час) интегральное накопление продукта.
Х=3,25-1,65ЕХР[-0,063(t-17)], Хр=3,25; Х1=1,6; А=0,065 (1/час).
Максимальная удельная скорость роста=0,063 (1/час), t1=17 час.
Основные параметры, которые были определены по динамике биомассы продуцента и самого продукта отражены в уравнениях накопления биомассы и продуктов, которые, в свою очередь, показаны на соответствующих рисунках.
Здесь Хstab1=0,8 усл.ед ОП может рассчитываться или может быть определено экспериментально. В данном случае Р1=18 г/л для Левана и 42 г/л для спирта. В данном примере были также рассчитаны изменения образования продукта на экспоненциальной фазе роста культуры. Они приведены на фиг.12 пунктирной линией.
Пример 6 (фиг.13)
Здесь Хр=14,3 г/л, Х1=5,33 г/л, Р1=0,02275 г/100 мл, kdivs=0,0014 [(грамм фосфора/100 мл)/(грамм делящихся клеток/литр) в сутки], kstabs=-0,0007 [(грамм фосфора/100 мл)/(грамм клеток в возрасте 0 клеточного цикла/литр)в сутки], А=0,23 (1/сутки), Хstab1=2,49 г/л.
Пример 7(фиг.14 и 15)
Этот пример демонстрирует возможность определения внутренней структуры биомассы Clostridium acetobutilicum по динамике общего накопления биомассы (фиг.14). Расчет проведен по представленным в книге “Основы биохимической инженерии” [Дж.Бейли, Д.Оллис, т.1, стр.551] экспериментальным динамикам накопления продуцента и кислот – уксусной и масляной. Фиг.15 демонстрирует накопление ацетата и бутирата.
Здесь Хр=4,21 г/л, Х1=1,8 г/л, для ацетата Р1=1,4 г/л, для ацетата kdiv=0,4755 [(грамм/литр)/(грамм делящихся клеток/литр) в час], для бутирата kdiv=0,3057 [(грамм/литр)/(грамм делящихся клеток/литр) в час], для бутирата Р1=2 г/л; А=0,173 (1/час), Хstab1=0,88 г/л.
Формула изобретения Способ получения биомассы и целевых продуктов синтеза с заданными технологическими параметрами путем процесса глубинного культивирования и организации контроля целевых продуктов синтеза, включающий определение оптимальных профилей изменения технологических параметров и алгоритмов оперативного управления по физиологическим параметрам, определяемым исходя из замеров количества биомассы при проведении процесса культивирования, отличающийся тем, что технологические параметры биомассы определяют по ее возрастной структуре для фазы замедленного роста, состоящей из делящихся и неделящихся клеток нулевого возраста, рассчитывая по уравнениям: Xdiv=Xp-(Xp-Xl)EXP(-A(t-tl))-ХstablЕХР(A(t-tl)) и Хstab=Хstab l·EXP[A·(t-tl)], где Хdiv -текущее количество биомассы делящихся клеток на момент времени t; Хstab – текущее количество клеток в нулевом возрасте на момент времени t; Хр – равновесное количество общей биомассы, когда вся энергия, вносимая в сосуд с субстратами, расходуется только на функции поддержания жизнедеятельности клеток; Xl – количество общей биомассы в момент начала лимитирования по какому-либо субстрату главного фактора, определяющего наступление и развитие фазы замедленного роста; t – текущее время с начала процесса культивирования; tl – момент времени, соответствующий Xl; А=b/а – удельная скорость замедления роста биомассы, где b – удельные затраты на поддержание жизнедеятельности биомассы, единиц субстрата/единиц биомассы в единицу времени, а – трофический коэффициент единиц субстрата/единиц биомассы, Xstabl – количество клеток в нулевом возрасте в момент начала лимитирования; а для получения целевых продуктов синтеза в качестве технологического параметра синтеза продукта или утилизации субстрата используют текущее количество биомассы делящихся клеток, при этом при синтезе продуктов, определяемом только делящимися клетками, используют соотношение Р=Рl+(kdiv/А){ХрLn[(Хр-Хl)/(Хр-X)]-(Х-Xl)[1+Xstabl/(Xp(Xp-Xl))]}, при синтезе продукта, определяемом как делящимися клетками, так и клетками, находящимися в нулевом возрасте клеточного цикла, используют соотношение P=Pl+(kdiv/A){XpLn[(Xp-Xl)/(Xp-X)]-(X-Xl)}+ (1/A)(kstab-kdiv)Xstabl(Х-Хl)/(Хр-Х), а при утилизации субстратов, определяемой делящимися клетками и клетками, находящимися в нулевом возрасте клеточного цикла, используют соотношение S=Sl(kdivs/A){XpLn[(Xp-Xl)/(Xp-X)]-(X-Xl)}- {1/А)(kstabs-kdivs)Хstabl(Х-Хl)/(Хр-X), где kdiv, kstab, kdivs, kstabs – константы скоростей биохимических реакций; S, Р, Х – количество субстрата, продукта и общей биомассы, соответственно. РИСУНКИ
|
||||||||||||||||||||||||||