Патент на изобретение №2181772
|
||||||||||||||||||||||||||
(54) СИНТЕЗ ГАММА-ЛИНОЛЕНОВОЙ КИСЛОТЫ ПОД ДЕЙСТВИЕМ ДЕЛЬТА-6-ДЕСАТУРАЗЫ
(57) Реферат: Изобретение предназначено для использования в селекции растений. С помощью нуклеиновой кислоты, кодирующей фермент  6-десатуразу, трансформируют растения. Указанная трансформация способствует увеличенному превращению линолевой кислоты в 6-десатуразу, трансформируют растения. Указанная трансформация способствует увеличенному превращению линолевой кислоты в  -линоленовую и октадекатетраеновую кислоты. Это позволяет обогатить пищевые растения ненасыщенными жирными кислотами, в том числе арахидоновой кислотой. Одновременно изменение метаболизма в растении за счет перераспределения продуцирования различных жирных кислот придает такому растению устойчивость к холоду. 12 с. и 13 з.п. ф-лы, 10 ил., 4 табл. -линоленовую и октадекатетраеновую кислоты. Это позволяет обогатить пищевые растения ненасыщенными жирными кислотами, в том числе арахидоновой кислотой. Одновременно изменение метаболизма в растении за счет перераспределения продуцирования различных жирных кислот придает такому растению устойчивость к холоду. 12 с. и 13 з.п. ф-лы, 10 ил., 4 табл.
Линолевая кислота (18: 2)(LA) превращается в гамма-линоленовую кислоту (18:3)(GLA) под действием такого фермента, как 6-десатураза. Когда этот фермент или кодирующую его нуклеиновую кислоту вводят в клетки, синтезирующие линолевую кислоту, образуется гамма-линоленовая кислота. Настоящее изобретение относится к нуклеиновым кислотам, содержащим ген 6-десатуразы. Более конкретно, в объем этого изобретения входят нуклеиновые кислоты, содержащие промоторы, кодирующие области и терминальные области генов 6-десатуразы. Настоящее изобретение далее относится к рекомбинантным конструкциям, содержащим кодирующую область 6-десатуразы в функциональном сочетании с разнородными регуляторными последовательностями. Нуклеиновые кислоты и рекомбинантные конструкции по данному изобретению полезны для получения гамма-линоленовой кислоты в трансгенных организмах.
Ненасыщенные жирные кислоты, такие как линолевая кислота (C189,12) и  -линоленовая кислота (C189,12,15), являются важными пищевыми компонентами, которые не могут быть синтезированы в организме позвоночных животных, так как клетки позвоночных животных могут создавать двойные связи в 9-положении жирных кислот, но не могут создавать дополнительные двойные связи между двойной связью в 9-положении и метильным концом цепи жирной кислоты. Поскольку они являются предшественниками других продуктов, линолевая и -линоленовая кислоты являются важными жирными кислотами, источником которых обычно являются растения. В организме млекопитающих линолевая кислота превращается в -линоленовую кислоту (GLA, C186,9,12), которая в свою очередь превращается в арахидоновую кислоту (20:4), особенно важную жирную кислоту, так как она является основным предшественником большинства простагландинов.
Линолевая кислота вследствие конечного превращения в гамма-линоленовую кислоту и арахидоновую кислоту удовлетворяет потребности организма в гамма-линоленовой кислоте и арахидоновой кислоте. Однако существует определенная зависимость между потреблением насыщенных жиров и вероятностью возникновения гиперхолестеринемии, атеросклероза и других клинических нарушений, которые влекут за собой ишемическую болезнь сердца, в то время как потребление ненасыщенных жиров способствует снижению содержания холестерина в крови и уменьшению вероятности заболевания атеросклерозом. Терапевтические преимущества пищевой гамма-линоленовой кислоты, вероятно, связаны с тем, что эта кислота является предшественником арахидоновой кислоты и, таким образом, способствует синтезу простагландина. Поэтому гораздо полезнее включать в рацион питания ненасыщенную гамма-линоленовую кислоту, чем линолевую кислоту. Однако гамма-линоленовая кислота отсутствует практически во всех возделываемых сельскохозяйственных культурах.
Линолевая кислота превращается в гамма-линоленовую кислоту под действием такого фермента, как 6-десатураза. 6-Десатураза, фермент из более чем 350 аминокислот, имеет мембраносвязывающую область и активный центр для десатурации жирных кислот. Если этот фермент ввести в клетки, которые эндогенно синтезируют линолевую кислоту, а не гамма-линоленовую кислоту, то происходит образование гамма-линоленовой кислоты. Настоящее изобретение, предусматривая ген, кодирующий 6-десатуразу, позволяет получить трансгенные организмы, которые содержат функциональную 6-десатуразу и синтезируют гамма-линоленовую кислоту. Помимо возможности синтеза гамма-линоленовой кислоты в больших количествах настоящим изобретением предусматриваются новые пищевые источники гамма-линоленовой кислоты.
Настоящее изобретение включает выделенные гены 6-десатуразы. В частности, выделенные гены содержат промоторы 6-десатуразы, кодирующие области и терминальные области.
Настоящее изобретение далее включает экспрессирующие векторы, содержащие промотор 6-десатуразы, кодирующую область и терминальные области.
Следующим аспектом данного изобретения являются экспрессирующие векторы, содержащие кодирующую область 6-десатуразы в функциональном сочетании с разнородными регуляторными областями, то есть с элементами, не выделяемыми из гена 6-десатуразы.
Настоящим изобретением предусматриваются также клетки и организмы, содержащие векторы по данному изобретению, и потомство таких организмов.
Очередным аспектом настоящего изобретения предусматривается выделенная бактериальная 6-десатураза. Предусматривается также выделенная растительная 6-десатураза.
Еще одним аспектом данного изобретения является способ получения растений с более высоким содержанием гамма-линоленовой кислоты.
Настоящим изобретением предусматривается также способ получения холодоустойчивых растений.
На фиг. 1 изображены профили водорастворимости для выведенных аминокислотных последовательностей 6-десатуразы Synechocystis (часть А) и 12-десатуразы (часть В). Предполагаемые области, пронизывающие мембрану, обозначены сплошными линиями. Гидрофобный индекс высчитан для окна, вмещающего 19 аминокислотных остатков [Kyte, et al. (1982) J. Molec. Biol. 157].
На фиг. 2 изображены профили газожидкостной хроматографии для Anabaena дикого типа (часть А) и трансгенного типа (часть В).
На фиг.3 дано схематическое изображение карт космид cSy75, cSyl3 и cSy7 с перекрывающимися областями и субклонами. Исходные области субклонов cSy75. cSy75-3,5 и cSy7 обозначены пунктирными диагональными линиями. Инактивированные сайты рестрикции указаны в скобках.
На фиг. 4 изображены профили газожидкостной хроматографии для табака дикого типа (часть А) и трансгенного типа (часть В).
На фиг.5А изображена ДНК-последовательность кДНК 6-десатуразы, выделенной из бурачника.
На фиг. 5В изображена белковая последовательность открытой рамки считывания в кДНК 6-десатуразы, выделенная из бурачника. Показаны три аминокислотных элемента, характерных для десатураз, которые в порядке следования представляют собой липидный блок, металлический блок 1 (metal box I) и металлический блок 2 (metal box 2).
На фиг. 6 изображена дендрограмма, показывающая сходство 6-десатуразы бурачника с другими мембраносвязанными десатуразами. Аминокислотная последовательность 6-десатуразы бурачника сравнивается с десатуразами других известных типов на основании данных, приведенных в Gene Works (IntelliGenetics). Числовые данные соответствуют относительным филогенетическим расстояниям между сравниваемыми подгруппами.
На фиг.7 представлена карта рестрикции 221.6.NOS и 121.6.NOS. На карте рестрикции 221. 6.NOS остальной части плазмиды является рВI221, а на карте рестрикции 121.6.NOS остальной частью плазмиды является рВI121.
На фиг.8 изображены профили газожидкостной хроматографии для мнимотрансфецированных (часть А) и трасфецированных 221.6.NOS (часть В) клеток моркови. Показаны положения, соответствующие 18:2, 18:3 и 18:3 (GLA).
На фиг.9 изображены профили газожидкостной хроматографии для нетрансформированного табачного листа (часть А) и табачного листа, трансформированного 121.6.NOS. Показаны положения 18:2, 18:3, 18:3 (GLA) и 18:4.
На фиг. 10 изображены профили газожидкостной хроматографии для нетрансформированных семян табака (часть А) и семян табака, трансформированных 121. 6.NOS. Показаны положения 18:2, 18:3, 18:3 (GLA).
В объем настоящего изобретения входят выделенные нуклеиновые кислоты, кодирующие 6-десатуразу. Для идентификации нуклеиновой кислоты, кодирующей 6-десатуразу, из организма, синтезирующего гамма-линоленовую кислоту, выделяют ДНК. Указанным организмом может быть, например, животная клетка, некоторые грибы (например, Mortierella), некоторые бактерии (например, Synechocystis) и некоторые растения (бурачник, Oenotherа, смородина). Геномную ДНК можно выделить с помощью различных методов, хорошо известных специалистам в этой области, например, с помощью метода, описанного Самбруком и др. (Sambrook et al.) (1989) в справочнике Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY. Выделенную ДНК делят на фрагменты с помощью физических методов или ферментативного расщепления и клонируют в соответствующем векторе, например бактериофаге или космидном векторе, в соответствии с любым хорошо известным методом из целого ряда методов, представленных в справочных материалах, таких как справочник Самбрука и др. (1989). Особое внимание в этом изобретении уделено экспрессирующим векторам, содержащим ДНК по настоящему изобретению. ДНК, кодирующую 6-десатуразу, можно идентифицировать посредством анализа приобретения функции. Вектор, содержащий фрагментарную ДНК, переносят, например, путем инфицирования, трансконъюгирования, трансфекции в организм-хозяин, который синтезирует линолевую кислоту, а не гамма-линоленовую кислоту. Используемый в этом описании изобретения термин “трансформация” означает включение посторонней ДНК в клетку-хозяин. Методы введения рекомбинантной ДНК в организм-хозяин известны специалистам в этой области, и с ними можно ознакомиться, например, в справочнике Самбрука и др.(1989). Синтез гамма-линоленовой кислоты этими организмами (то есть приобретение функции) определяют, например, газовой хроматографией или другими методами, известными специалистам в этой области. Организмы, которые стали синтезировать гамма-линоленовую кислоту, то есть приобрели новую функцию в результате введения вектора, определяются как экспрессирующие ДНК, кодирующую 6-десатуразу, причем указанную ДНК выделяют из организмов. Выделенную ДНК можно снова разделить на фрагменты, клонировать в экспрессирующих векторах и функционально исследовать с помощью вышеуказанных методов с целью более точного определения ДНК, кодирующей 6-десатуразу.
В качестве примера, иллюстрирующего настоящее изобретение, случайную ДНК выделяют из цианобактерии Synechocystis. входящей в Коллекцию культур Пастера (РСС) 6803 и Американскую коллекцию типовых культур (АТСС) 27184, клонируют в космидном векторе и вводят путем трансконъюгирования в штамм цианобактерий Anabaena РСС 7120, АТСС 27893, не синтезирующих гамма-линоленовой кислоты. Синтез гамма-линоленовой кислоты из линолевой кислоты Anabaena контролируют посредством газовой хроматографии, после чего выделяют соответствующий фрагмент ДНК.
Выделенную ДНК секвенируют с помощью методов, хорошо известных специалистам в этой области, которые описаны, например, в справочнике Самбрука и др. (1989).
В соответствии с настоящим изобретением выделяют молекулы ДНК, содержащие гены 6-десатуразы. В частности, из цианобактерий Synechocystis была выделена ДНК длиной 3,588 тысяч пар нуклеотидов (т.п.н.), содержащая ген 6-десатуразы. Была определена нуклеотидная последовательность ДНК длиной 3,588 т.п.н., которая представлена в последовательности с индификационным No 1. Открытые рамки считывания, определяющие потенциальные кодирующие области, находятся в нуклеотидах с 317 по 1507 и в нуклеотидах с 2002 по 3081. Чтобы установить нуклеотиды, имеющие непосредственное отношение к кодированию 6-десатуразы, фрагмент ДНК длиной 3,588 т.п.н., который определяет активность 6-десатуразы, расщепляют на два подфрагмента, каждый из которых содержит только одну открытую рамку считывания. Фрагмент ORF1 содержит нуклеотиды с 1 по 1704, а фрагмент ORF2 содержит нуклеотиды с 1705 по 3588. Каждый фрагмент субклонируют в прямой и обратной ориентации в конъюгированном экспрессирующем векторе (АМ542, Wolk et al.[1984] Proc. Natl. Acad. Sci. USA 81, 1561), который содержит промотор карбоксилазы цианобактерий. Полученные конструкции (то есть ORF1(F), ORF1(R), ORF2(F) и ORF2(R) конъюгируют со штаммом Anabaena дикого типа РСС 7120 с помощью стандартных методов (см. , например, Wolk et al. (1984) Proc. Natl. Acad. Sci. USA 81. 1561). Коньюгированные клетки штамма Anabaena идентифицированы в виде зеленых колоний NeoR на коричневом фоне погибающих неконъюгированных клеток после выращивания в течение двух недель на селективной среде (стандартные минеральные среды BG11N, содержащие 30 мкг/мл неомицина в соответствии с описанием Rippka et al., (1979) J.Gen Microbiol. 111, 1). Зеленые колонии выделяют и выращивают в селективных жидких средах (BG11N+15 мкг/мл неомицина). Липиды экстрагируют стандартными методами (например, по методу Dahmar et al. (1989) Journal of American Oil Chemical Society 66, 543) из полученных трансконъюгантов, содержащих конструкции ORF1 и ORF2 с прямой и обратной ориентацией. Для сравнения липиды экстрагируют также из других культур Anabaena и Synechocystis. Метиловые эфиры жирных кислот анализируют посредством газожидкостной хроматографии, например, с использованием газожидкостного хроматографа Тrасоr-560, оборудованного ионизационным детектором с водородным пламенем и капиллярной колонкой. Результаты газожидкостной хроматографии приведены в таблице 1.
Как показывает газожидкостная хроматография, штамм Anabaеna, не синтезирующий гамма-линоленовую кислоту, приобретает функцию синтеза гамма-линоленовой кислоты, когда путем трансконъюгирования в него вводят конструкцию, содержащую ORF2 с прямой ориентацией. Трансконъюгаты, содержащие конструкции с ORF2 с обратной ориентацией относительно промотора карбоксилазы, или ORF1 с любой ориентацией, не синтезируют гамма-линоленовой кислоты. Этот анализ показывает, что только одна открытая рамка считывания (ORF2) во фрагменте 1884 комплементарной пары гетероциклических оснований ДНК кодирует -линоленовая кислота (C189,12,15), являются важными пищевыми компонентами, которые не могут быть синтезированы в организме позвоночных животных, так как клетки позвоночных животных могут создавать двойные связи в 9-положении жирных кислот, но не могут создавать дополнительные двойные связи между двойной связью в 9-положении и метильным концом цепи жирной кислоты. Поскольку они являются предшественниками других продуктов, линолевая и -линоленовая кислоты являются важными жирными кислотами, источником которых обычно являются растения. В организме млекопитающих линолевая кислота превращается в -линоленовую кислоту (GLA, C186,9,12), которая в свою очередь превращается в арахидоновую кислоту (20:4), особенно важную жирную кислоту, так как она является основным предшественником большинства простагландинов.
Линолевая кислота вследствие конечного превращения в гамма-линоленовую кислоту и арахидоновую кислоту удовлетворяет потребности организма в гамма-линоленовой кислоте и арахидоновой кислоте. Однако существует определенная зависимость между потреблением насыщенных жиров и вероятностью возникновения гиперхолестеринемии, атеросклероза и других клинических нарушений, которые влекут за собой ишемическую болезнь сердца, в то время как потребление ненасыщенных жиров способствует снижению содержания холестерина в крови и уменьшению вероятности заболевания атеросклерозом. Терапевтические преимущества пищевой гамма-линоленовой кислоты, вероятно, связаны с тем, что эта кислота является предшественником арахидоновой кислоты и, таким образом, способствует синтезу простагландина. Поэтому гораздо полезнее включать в рацион питания ненасыщенную гамма-линоленовую кислоту, чем линолевую кислоту. Однако гамма-линоленовая кислота отсутствует практически во всех возделываемых сельскохозяйственных культурах.
Линолевая кислота превращается в гамма-линоленовую кислоту под действием такого фермента, как 6-десатураза. 6-Десатураза, фермент из более чем 350 аминокислот, имеет мембраносвязывающую область и активный центр для десатурации жирных кислот. Если этот фермент ввести в клетки, которые эндогенно синтезируют линолевую кислоту, а не гамма-линоленовую кислоту, то происходит образование гамма-линоленовой кислоты. Настоящее изобретение, предусматривая ген, кодирующий 6-десатуразу, позволяет получить трансгенные организмы, которые содержат функциональную 6-десатуразу и синтезируют гамма-линоленовую кислоту. Помимо возможности синтеза гамма-линоленовой кислоты в больших количествах настоящим изобретением предусматриваются новые пищевые источники гамма-линоленовой кислоты.
Настоящее изобретение включает выделенные гены 6-десатуразы. В частности, выделенные гены содержат промоторы 6-десатуразы, кодирующие области и терминальные области.
Настоящее изобретение далее включает экспрессирующие векторы, содержащие промотор 6-десатуразы, кодирующую область и терминальные области.
Следующим аспектом данного изобретения являются экспрессирующие векторы, содержащие кодирующую область 6-десатуразы в функциональном сочетании с разнородными регуляторными областями, то есть с элементами, не выделяемыми из гена 6-десатуразы.
Настоящим изобретением предусматриваются также клетки и организмы, содержащие векторы по данному изобретению, и потомство таких организмов.
Очередным аспектом настоящего изобретения предусматривается выделенная бактериальная 6-десатураза. Предусматривается также выделенная растительная 6-десатураза.
Еще одним аспектом данного изобретения является способ получения растений с более высоким содержанием гамма-линоленовой кислоты.
Настоящим изобретением предусматривается также способ получения холодоустойчивых растений.
На фиг. 1 изображены профили водорастворимости для выведенных аминокислотных последовательностей 6-десатуразы Synechocystis (часть А) и 12-десатуразы (часть В). Предполагаемые области, пронизывающие мембрану, обозначены сплошными линиями. Гидрофобный индекс высчитан для окна, вмещающего 19 аминокислотных остатков [Kyte, et al. (1982) J. Molec. Biol. 157].
На фиг. 2 изображены профили газожидкостной хроматографии для Anabaena дикого типа (часть А) и трансгенного типа (часть В).
На фиг.3 дано схематическое изображение карт космид cSy75, cSyl3 и cSy7 с перекрывающимися областями и субклонами. Исходные области субклонов cSy75. cSy75-3,5 и cSy7 обозначены пунктирными диагональными линиями. Инактивированные сайты рестрикции указаны в скобках.
На фиг. 4 изображены профили газожидкостной хроматографии для табака дикого типа (часть А) и трансгенного типа (часть В).
На фиг.5А изображена ДНК-последовательность кДНК 6-десатуразы, выделенной из бурачника.
На фиг. 5В изображена белковая последовательность открытой рамки считывания в кДНК 6-десатуразы, выделенная из бурачника. Показаны три аминокислотных элемента, характерных для десатураз, которые в порядке следования представляют собой липидный блок, металлический блок 1 (metal box I) и металлический блок 2 (metal box 2).
На фиг. 6 изображена дендрограмма, показывающая сходство 6-десатуразы бурачника с другими мембраносвязанными десатуразами. Аминокислотная последовательность 6-десатуразы бурачника сравнивается с десатуразами других известных типов на основании данных, приведенных в Gene Works (IntelliGenetics). Числовые данные соответствуют относительным филогенетическим расстояниям между сравниваемыми подгруппами.
На фиг.7 представлена карта рестрикции 221.6.NOS и 121.6.NOS. На карте рестрикции 221. 6.NOS остальной части плазмиды является рВI221, а на карте рестрикции 121.6.NOS остальной частью плазмиды является рВI121.
На фиг.8 изображены профили газожидкостной хроматографии для мнимотрансфецированных (часть А) и трасфецированных 221.6.NOS (часть В) клеток моркови. Показаны положения, соответствующие 18:2, 18:3 и 18:3 (GLA).
На фиг.9 изображены профили газожидкостной хроматографии для нетрансформированного табачного листа (часть А) и табачного листа, трансформированного 121.6.NOS. Показаны положения 18:2, 18:3, 18:3 (GLA) и 18:4.
На фиг. 10 изображены профили газожидкостной хроматографии для нетрансформированных семян табака (часть А) и семян табака, трансформированных 121. 6.NOS. Показаны положения 18:2, 18:3, 18:3 (GLA).
В объем настоящего изобретения входят выделенные нуклеиновые кислоты, кодирующие 6-десатуразу. Для идентификации нуклеиновой кислоты, кодирующей 6-десатуразу, из организма, синтезирующего гамма-линоленовую кислоту, выделяют ДНК. Указанным организмом может быть, например, животная клетка, некоторые грибы (например, Mortierella), некоторые бактерии (например, Synechocystis) и некоторые растения (бурачник, Oenotherа, смородина). Геномную ДНК можно выделить с помощью различных методов, хорошо известных специалистам в этой области, например, с помощью метода, описанного Самбруком и др. (Sambrook et al.) (1989) в справочнике Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY. Выделенную ДНК делят на фрагменты с помощью физических методов или ферментативного расщепления и клонируют в соответствующем векторе, например бактериофаге или космидном векторе, в соответствии с любым хорошо известным методом из целого ряда методов, представленных в справочных материалах, таких как справочник Самбрука и др. (1989). Особое внимание в этом изобретении уделено экспрессирующим векторам, содержащим ДНК по настоящему изобретению. ДНК, кодирующую 6-десатуразу, можно идентифицировать посредством анализа приобретения функции. Вектор, содержащий фрагментарную ДНК, переносят, например, путем инфицирования, трансконъюгирования, трансфекции в организм-хозяин, который синтезирует линолевую кислоту, а не гамма-линоленовую кислоту. Используемый в этом описании изобретения термин “трансформация” означает включение посторонней ДНК в клетку-хозяин. Методы введения рекомбинантной ДНК в организм-хозяин известны специалистам в этой области, и с ними можно ознакомиться, например, в справочнике Самбрука и др.(1989). Синтез гамма-линоленовой кислоты этими организмами (то есть приобретение функции) определяют, например, газовой хроматографией или другими методами, известными специалистам в этой области. Организмы, которые стали синтезировать гамма-линоленовую кислоту, то есть приобрели новую функцию в результате введения вектора, определяются как экспрессирующие ДНК, кодирующую 6-десатуразу, причем указанную ДНК выделяют из организмов. Выделенную ДНК можно снова разделить на фрагменты, клонировать в экспрессирующих векторах и функционально исследовать с помощью вышеуказанных методов с целью более точного определения ДНК, кодирующей 6-десатуразу.
В качестве примера, иллюстрирующего настоящее изобретение, случайную ДНК выделяют из цианобактерии Synechocystis. входящей в Коллекцию культур Пастера (РСС) 6803 и Американскую коллекцию типовых культур (АТСС) 27184, клонируют в космидном векторе и вводят путем трансконъюгирования в штамм цианобактерий Anabaena РСС 7120, АТСС 27893, не синтезирующих гамма-линоленовой кислоты. Синтез гамма-линоленовой кислоты из линолевой кислоты Anabaena контролируют посредством газовой хроматографии, после чего выделяют соответствующий фрагмент ДНК.
Выделенную ДНК секвенируют с помощью методов, хорошо известных специалистам в этой области, которые описаны, например, в справочнике Самбрука и др. (1989).
В соответствии с настоящим изобретением выделяют молекулы ДНК, содержащие гены 6-десатуразы. В частности, из цианобактерий Synechocystis была выделена ДНК длиной 3,588 тысяч пар нуклеотидов (т.п.н.), содержащая ген 6-десатуразы. Была определена нуклеотидная последовательность ДНК длиной 3,588 т.п.н., которая представлена в последовательности с индификационным No 1. Открытые рамки считывания, определяющие потенциальные кодирующие области, находятся в нуклеотидах с 317 по 1507 и в нуклеотидах с 2002 по 3081. Чтобы установить нуклеотиды, имеющие непосредственное отношение к кодированию 6-десатуразы, фрагмент ДНК длиной 3,588 т.п.н., который определяет активность 6-десатуразы, расщепляют на два подфрагмента, каждый из которых содержит только одну открытую рамку считывания. Фрагмент ORF1 содержит нуклеотиды с 1 по 1704, а фрагмент ORF2 содержит нуклеотиды с 1705 по 3588. Каждый фрагмент субклонируют в прямой и обратной ориентации в конъюгированном экспрессирующем векторе (АМ542, Wolk et al.[1984] Proc. Natl. Acad. Sci. USA 81, 1561), который содержит промотор карбоксилазы цианобактерий. Полученные конструкции (то есть ORF1(F), ORF1(R), ORF2(F) и ORF2(R) конъюгируют со штаммом Anabaena дикого типа РСС 7120 с помощью стандартных методов (см. , например, Wolk et al. (1984) Proc. Natl. Acad. Sci. USA 81. 1561). Коньюгированные клетки штамма Anabaena идентифицированы в виде зеленых колоний NeoR на коричневом фоне погибающих неконъюгированных клеток после выращивания в течение двух недель на селективной среде (стандартные минеральные среды BG11N, содержащие 30 мкг/мл неомицина в соответствии с описанием Rippka et al., (1979) J.Gen Microbiol. 111, 1). Зеленые колонии выделяют и выращивают в селективных жидких средах (BG11N+15 мкг/мл неомицина). Липиды экстрагируют стандартными методами (например, по методу Dahmar et al. (1989) Journal of American Oil Chemical Society 66, 543) из полученных трансконъюгантов, содержащих конструкции ORF1 и ORF2 с прямой и обратной ориентацией. Для сравнения липиды экстрагируют также из других культур Anabaena и Synechocystis. Метиловые эфиры жирных кислот анализируют посредством газожидкостной хроматографии, например, с использованием газожидкостного хроматографа Тrасоr-560, оборудованного ионизационным детектором с водородным пламенем и капиллярной колонкой. Результаты газожидкостной хроматографии приведены в таблице 1.
Как показывает газожидкостная хроматография, штамм Anabaеna, не синтезирующий гамма-линоленовую кислоту, приобретает функцию синтеза гамма-линоленовой кислоты, когда путем трансконъюгирования в него вводят конструкцию, содержащую ORF2 с прямой ориентацией. Трансконъюгаты, содержащие конструкции с ORF2 с обратной ориентацией относительно промотора карбоксилазы, или ORF1 с любой ориентацией, не синтезируют гамма-линоленовой кислоты. Этот анализ показывает, что только одна открытая рамка считывания (ORF2) во фрагменте 1884 комплементарной пары гетероциклических оснований ДНК кодирует 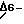 десатуразу. Фрагмент 1884 комплементарной пары гетероциклических оснований ДНК представлен в виде последовательностей с идентификационным No.3. Это подтверждается полным сходством профилей водорастворимости у 6-десатуразы и 12-десатуразы [Wada et al. (1990) Nature 347], как показано на фиг.1, (А) и (В).
Кроме того, в соответствии с настоящим изобретением из бурачника (Borago officinalis) можно выделить кДНК, содержащую ген 6-десатуразы. Была определена нуклеотидная последовательность кДНК длиной 1,685 тысячи пар нуклеотидов (т.п.н.), которая представлена на фиг.5А (последовательность с индентификационным No.4). Инициирующий кодон ATG и терминирующий кодон подчеркнуты. Аминокислотная последовательность, соответствующая открытой рамке считывания в 6-десатуразе бурачника, представлена на фиг.5В (последовательность с идентификационным No. 5).
Выделенные нуклеиновые кислоты, кодирующие 6-десатуразу, можно идентифицировать в других организмах, синтезирующих гамма-линоленовую кислоту, с помощью анализа приобретения функции, описанного выше, или с помощью методов гибридизации нуклеиновых кислот с использованием выделенной нуклеиновой кислоты, которая кодирует Synechocystis или 6-десатуразу бурачника в виде зонда гибридизации. Методы клонирования генома и кДНК хорошо известны специалистам в этой области, и настоящим изобретением предполагается их применение. Зонд гибридизации может содержать всю последовательность ДНК, описываемую как последовательность с идентификационным No.1 или последовательность с идентификационным No.4, либо рестрикционный фрагмент или другой фрагмент ДНК, включающий олигонуклеотидный зонд. Методы клонирования гомологичных генов путем перекрестной гибридизации известны специалистам в этой области, и с ними можно ознакомиться, например, в работе Самбрука (1989) и Белца и др. (Beltz et al.) (1983)Methods in Enzymology 100, 266.
В соответствии с другим методом идентификации гена 6-десатуразы в организме, синтезирующем гамма-линоленовую кислоту, создают библиотеку кДНК из поли-A+PHK, выделенной из полисомной РНК. Чтобы удалить многочисленные экспрессируемые гены из популяции кДНК, кДНК или их фрагменты, соответствующие многочисленным генам кДНК, используют в виде зондов гибридизации для библиотеки кДНК. Негибридизирующиеся зоны удаляют, полученные бактериальные колонии используют для инокуляции жидких культур и секвенируют. Например, чтобы удалить другие кДНК запасного белка семян из библиотеки кДНК, полученной из полисомной РНК бурачника, типы кДНК, соответствующие сильно экспрессированным запасным белкам семян, сначала гибридизируют с библиотекой кДНК. “Выделенную” библиотеку ДНК затем используют для получения меток экспрессированных последовательностей (ETSs), которые применяют для сканирования базы данных, такой как GenBank, с целью идентификации потенциальных десатураторов.
Трансгенные организмы, которые приобретают функцию синтеза гамма-линоленовой кислоты в результате введения ДНК, кодирующей -десатуразу, также приобретают функцию синтеза октадекатетраеновой кислоты (18:46,9,12,15). Октадекатетраеновая кислота обычно присутствует в рыбьем жире и в некоторых видах растений семейства Boraginaceae (Craig et al.[1964] J.Amer. Oil Chem. Soc.41,209-211; Gross et al. [1976] Can.J.Plant Sci.56,659-664). В трансгенных организмах по настоящему изобретению октадекатетраеновая кислота образуется в результате дальнейшей десатурации -линоленовой кислоты под действием 6-десатуразы или десатурации гамма-линоленовой кислоты под действием 12-десатуразы.
359 аминокислот, кодируемых ORF2, то есть открытой рамкой считывания, кодирующей 6-десатуразу Synechocystis, представлены в виде последовательности с индентификационным No. 2. Открытая рамка считывания, кодирующая 6-десатуразу бурачника, представлена в последовательности с идентификационным No. 5. Настоящее изобретение далее включает другие нуклеотидные последовательности, которые кодируют аминокислоты последовательности с идентификационным No.2 и в последовательности с идентификационным No.5. Специалисты в этой области могут легко идентифицировать такие последовательности, которые появляются, например, в результате вырожденности генетического кода. Кроме того, специалисты в этой области с помощью описанного выше анализа приобретения функции могут определить более мелкие субфрагменты фрагментов, содержащих открытые рамки считывания, которые кодируют 6-десатуразу.
Настоящее изобретение включает любой такой полипептидный фрагмент 6-десатуразы и нуклеиновых кислот, которые сохраняют активности в отношении превращения линолевой кислоты в гамма-линоленовую кислоту.
В соответствии с другим аспектом настоящего изобретения вектор, содержащий нуклеиновую кислоту по данному изобретению, или более мелкий фрагмент, содержащий промотор, кодирующую последовательность и терминальную область гена 6-десатуразы переносят в организм, например, цианобактерий, у которых промотор 6-десатуразы и терминальные области являются функциональными. Поэтому данным изобретением предусматриваются организмы, синтезирующие рекомбинантную 6-десатуразу. Еще одним аспектом данного изобретения является выделенная 6-десатураза, которую можно выделить в чистом виде из рекомбинантных организмов с помощью стандартных методов очистки белков (Например, см. Ausubel et al. [1987] Current Protocols in Molecular Biology, Green Publishing Associates, New York).
Векторы, содержащие ДНК, кодирующую 6-десатуразу, также входят в объем настоящего изобретения. Специалистам в этой области должно быть понятно, что можно сконструировать соответствующие векторы, вызывающие экспрессию кодирующей последовательности 6-десатуразы в ряде организмов. Наиболее предпочтительными являются реплицируемые экспрессирующие векторы. Описываемые здесь реплицируемыми экспрессирующими векторами являются молекулы ДНК или РНК, созданные для контролируемой экспрессии требуемого гена, то есть гена 6-десатуразы. Этими десатуразу. Фрагмент 1884 комплементарной пары гетероциклических оснований ДНК представлен в виде последовательностей с идентификационным No.3. Это подтверждается полным сходством профилей водорастворимости у 6-десатуразы и 12-десатуразы [Wada et al. (1990) Nature 347], как показано на фиг.1, (А) и (В).
Кроме того, в соответствии с настоящим изобретением из бурачника (Borago officinalis) можно выделить кДНК, содержащую ген 6-десатуразы. Была определена нуклеотидная последовательность кДНК длиной 1,685 тысячи пар нуклеотидов (т.п.н.), которая представлена на фиг.5А (последовательность с индентификационным No.4). Инициирующий кодон ATG и терминирующий кодон подчеркнуты. Аминокислотная последовательность, соответствующая открытой рамке считывания в 6-десатуразе бурачника, представлена на фиг.5В (последовательность с идентификационным No. 5).
Выделенные нуклеиновые кислоты, кодирующие 6-десатуразу, можно идентифицировать в других организмах, синтезирующих гамма-линоленовую кислоту, с помощью анализа приобретения функции, описанного выше, или с помощью методов гибридизации нуклеиновых кислот с использованием выделенной нуклеиновой кислоты, которая кодирует Synechocystis или 6-десатуразу бурачника в виде зонда гибридизации. Методы клонирования генома и кДНК хорошо известны специалистам в этой области, и настоящим изобретением предполагается их применение. Зонд гибридизации может содержать всю последовательность ДНК, описываемую как последовательность с идентификационным No.1 или последовательность с идентификационным No.4, либо рестрикционный фрагмент или другой фрагмент ДНК, включающий олигонуклеотидный зонд. Методы клонирования гомологичных генов путем перекрестной гибридизации известны специалистам в этой области, и с ними можно ознакомиться, например, в работе Самбрука (1989) и Белца и др. (Beltz et al.) (1983)Methods in Enzymology 100, 266.
В соответствии с другим методом идентификации гена 6-десатуразы в организме, синтезирующем гамма-линоленовую кислоту, создают библиотеку кДНК из поли-A+PHK, выделенной из полисомной РНК. Чтобы удалить многочисленные экспрессируемые гены из популяции кДНК, кДНК или их фрагменты, соответствующие многочисленным генам кДНК, используют в виде зондов гибридизации для библиотеки кДНК. Негибридизирующиеся зоны удаляют, полученные бактериальные колонии используют для инокуляции жидких культур и секвенируют. Например, чтобы удалить другие кДНК запасного белка семян из библиотеки кДНК, полученной из полисомной РНК бурачника, типы кДНК, соответствующие сильно экспрессированным запасным белкам семян, сначала гибридизируют с библиотекой кДНК. “Выделенную” библиотеку ДНК затем используют для получения меток экспрессированных последовательностей (ETSs), которые применяют для сканирования базы данных, такой как GenBank, с целью идентификации потенциальных десатураторов.
Трансгенные организмы, которые приобретают функцию синтеза гамма-линоленовой кислоты в результате введения ДНК, кодирующей -десатуразу, также приобретают функцию синтеза октадекатетраеновой кислоты (18:46,9,12,15). Октадекатетраеновая кислота обычно присутствует в рыбьем жире и в некоторых видах растений семейства Boraginaceae (Craig et al.[1964] J.Amer. Oil Chem. Soc.41,209-211; Gross et al. [1976] Can.J.Plant Sci.56,659-664). В трансгенных организмах по настоящему изобретению октадекатетраеновая кислота образуется в результате дальнейшей десатурации -линоленовой кислоты под действием 6-десатуразы или десатурации гамма-линоленовой кислоты под действием 12-десатуразы.
359 аминокислот, кодируемых ORF2, то есть открытой рамкой считывания, кодирующей 6-десатуразу Synechocystis, представлены в виде последовательности с индентификационным No. 2. Открытая рамка считывания, кодирующая 6-десатуразу бурачника, представлена в последовательности с идентификационным No. 5. Настоящее изобретение далее включает другие нуклеотидные последовательности, которые кодируют аминокислоты последовательности с идентификационным No.2 и в последовательности с идентификационным No.5. Специалисты в этой области могут легко идентифицировать такие последовательности, которые появляются, например, в результате вырожденности генетического кода. Кроме того, специалисты в этой области с помощью описанного выше анализа приобретения функции могут определить более мелкие субфрагменты фрагментов, содержащих открытые рамки считывания, которые кодируют 6-десатуразу.
Настоящее изобретение включает любой такой полипептидный фрагмент 6-десатуразы и нуклеиновых кислот, которые сохраняют активности в отношении превращения линолевой кислоты в гамма-линоленовую кислоту.
В соответствии с другим аспектом настоящего изобретения вектор, содержащий нуклеиновую кислоту по данному изобретению, или более мелкий фрагмент, содержащий промотор, кодирующую последовательность и терминальную область гена 6-десатуразы переносят в организм, например, цианобактерий, у которых промотор 6-десатуразы и терминальные области являются функциональными. Поэтому данным изобретением предусматриваются организмы, синтезирующие рекомбинантную 6-десатуразу. Еще одним аспектом данного изобретения является выделенная 6-десатураза, которую можно выделить в чистом виде из рекомбинантных организмов с помощью стандартных методов очистки белков (Например, см. Ausubel et al. [1987] Current Protocols in Molecular Biology, Green Publishing Associates, New York).
Векторы, содержащие ДНК, кодирующую 6-десатуразу, также входят в объем настоящего изобретения. Специалистам в этой области должно быть понятно, что можно сконструировать соответствующие векторы, вызывающие экспрессию кодирующей последовательности 6-десатуразы в ряде организмов. Наиболее предпочтительными являются реплицируемые экспрессирующие векторы. Описываемые здесь реплицируемыми экспрессирующими векторами являются молекулы ДНК или РНК, созданные для контролируемой экспрессии требуемого гена, то есть гена 6-десатуразы. Этимивекторами предпочтительно являются плазмиды, бактериофаги, космиды или вирусы. Настоящим изобретением предполагается применение шаттл-векторов, например, описанных Волком и др. (Wolk et al.) (1984) Proc. Natl. Acad. Sci. USA. 1561-1565 и Бастосом и др. (Bustos et al.) (1991) J. Bacteriol. 174, 7525-7533. Самбрук и др. (1989), Goeddel, ed (1990) Methods in Enzymology 185 Academic Press, и Perbal (1988) A Practical Guide to Molecular Cloning, John Wiley and Sons, Inc., дают подробный обзор векторов, в которые можно вводить и экспрессировать нуклеиновую кислоту, кодирующую данную 6-десатуразу. Такие векторы содержат также последовательности нуклеиновых кислот, которые могут влиять на экспрессию нуклеиновых кислот, кодирующих 6-десатуразу. Элементы последовательности, способные вызывать экспрессию генного продукта, включают промоторы, энхансеры, предшествующие активирующие последовательности, сигналы терминации транскрипции и сайты полиаденилирования. Настоящим изобретением предусматриваются как конститутивные, так и тканеспецифичные промоторы. Для трансформации растительных клеток особый интерес представляет промотор 35S вируса мозаичной болезни цветной капусты (CaMV) и промоторы, активизируемые во время созревания семян растений. Все эти промоторы и элементы, регулирующие транскрипцию, в отдельности или в сочетании рассматриваются с целью их использования в данных реплицирующих векторах и известны специалистам в этой области. Промотор CaMV 355 описывается, например, Restrepo et al. (1990) Plant Cell 2, 987. В объем настоящего изобретения входят также генетически сконструированные и мутированные регуляторные последовательности.
Специалисты в этой области способны определить векторы и регуляторные элементы, пригодные для экспрессии в определенной клетке-хозяине. Например, для экспрессии 6-десатуразы в цианобактериях пригоден вектор, содержащий промотор из гена, кодирующего карбоксилазу вида Anabaena, функционально связанную с кодирующей областью 6-десатуразы и далее функционально связанную с сигналом терминации вида Synechocystis. Термин “функционально связанный” в этом контексте означает, что промоторные и терминаторные последовательности обеспечивают эффективную регуляцию транскрипции. В качестве другого примера можно привести вектор, пригодный для экспрессии 6-десатуразы в трансгенных растениях, который может содержать характерную для семян промоторную последовательность, выделяемую из гелиантинина, напина или глицинина, функционально связанных с кодирующей областью 6-десатуразы и далее функционально связанных с сигналом терминации семени или сигналом терминации нопалинсинтазы. Еще одним примером может служить вектор, служащий для экспрессии 6-десатуразы в растениях, который может содержать конститутивный промотор или тканеспецифичный промотор, функционально связанный с кодирующей областью 6-десатуразы и далее функционально связанный с конститутивным или тканеспецифичным терминатором или сигналом терминации нопалинсинтазы.
В частности, такие регуляторные элементы, как гелиантинин, описываемые в одновременно рассматриваемой заявке на патент США No.682354, поданной 8 апреля 1991 г., которая включена в это описание изобретения в качестве ссылки, являются промоторными элементами, служащими для экспрессии 6-десатуразы по настоящему изобретению.
Модификации рассматриваемых нуклеотидных последовательностей или регуляторных элементов, выполняющих здесь функции, входят в объем данного изобретения. Такие модификации включают вставки, замены и делеции, в частности замены, которые отражают вырожденность генетического кода.
Стандартные методы конструирования гибридных векторов хорошо известны специалистам в этой области и описываются в приводимых ссылочных материалах, таких как справочник Самбрука и др. (1989), или в любом из множества руководств по лабораторным исследованиям, которые относятся к технологии получения рекомбинантной ДНК и имеют широкое применение. Существует целый ряд способов, предназначенных для лигирования фрагментов ДНК, выбор которых зависит от характера концов фрагментов ДНК. Кроме того, в объем настоящего изобретения входят гибридные векторы, в которые включены элементы нуклеотидной последовательности, облегчающие клонирование, экспрессию или процессинг, например последовательности, кодирующие сигнальные пептиды, последовательности, кодирующие KDEL, которая необходима для удерживания белков в эндоплазматическом ретикулуме, или последовательности, кодирующие транзитные пептиды, которые направляют 6-десатуразу в хлоропласт. Такие последовательности известны специалистам в этой области. Оптимизированный транзитный пептид описывается, например, Van len Broeck et al. (1985) Nature 313, 358. Прокариотические и эукариотические сигнальные последовательности описываются, например, Michelis et al. (1982) Ann. Rev. Microbiol. 36, 425.
Дальнейшим аспектом настоящего изобретения предусматриваются организмы, неявляющиеся цианобактериями или растениями, которые содержат ДНК, кодирующую 6-десатуразу по данному изобретению. Трансгенными организмами, входящими в обьем настоящего изобретения, являются бактерии, цианобактерии, грибы, растения и животные. Выделенную ДНК по настоящему изобретению можно ввести в клетку-хозяин с помощью известных методов, например инфицирования, трансфекции, трансформации и трансконъюгирования. Методы переноса ДНК по настоящему изобретению в такие организмы широко известны и описываются в справочных материалах, таких как работа Самбрука и др. (1989).
Известны разнообразные методы трансформации растений. Ген 6-десатуразы можно ввести в растения с помощью процедуры-регенерации листового диска, описанной Horsch et al. (1985) Science 227, 1229. Можно использовать другие методы трансформации, такие как культивирование протопласта (Horsch et al. (1984) Science 223, 496; DeBlock et al. (1984) EMBO J. 2. 2143; Barton et al. (1983) Cell 32, 1033), которые также входят в объем данного изобретения. В соответствии с предпочтительным вариантом осуществления настоящего изобретения растения трансформируют с помощью векторов, выделенных из Agrobacterium. Однако существуют другие методы, позволяющие ввести гены 6-десатуразы по настоящему изобретению в растительные клетки. Такие альтернативные методы включают биобаллистические способы (Klein et al. (1987) Nature 327, 70), электропорацию, опосредованный химическими веществами ввод ДНК и использование вирусов или пыльцы в качестве векторов.
При необходимости в случае применения метода трансформации гены 6-десатуразы по настоящему изобретению можно ввести в растительный трансформирующий вектор, например бинарный вектор, описанный Bevan (1984) Nucleic Acids Res,12, 8111.
Растительные трансформирующие векторы можно получить путем изменения натуральной системы переноса гена Agrobacterium tumefaciens. Природная система включает плазмиды, индуцирующие образование опухолей (Ti) и содержащие большой сегмент, известный как Т-ДНК, который переносится в трансформированные растения. Другой сегмент Ti-плазмиды, vir область, имеет непосредственное отношение к переносу Т-ДНК. Область Т-ДНК ограничена концевыми повторами. В модифицированных бинарных векторах удалены гены, индуцирующие образование опухолей, и функции vir области используются для переноса чужеродной ДНК, ограниченной граничными последовательностями Т-ДНК. Т-область содержит также селективный маркер для устойчивости к антибиотикам и множественный сайт клонирования, предназначенный для ввода последовательностей для переноса. Такие сконструированные штаммы известны как “обезвреженные” (“disarmed”) штаммы A. tumefaciens и обеспечивают эффективную трансформацию последовательностей, ограниченных Т-областью, в ядерные геномы растений.
Листовые диски со стерилизованной поверхностью инокулируют “обезвреженным” видом A. tumefaciens, содержащим чужеродную ДНК, культивируют в течение двух дней, а затем переносят на среду, содержащую антибиотик. Трансформированные проростки выбирают после укоренения в среде, содержащей соответствующий антибиотик, переносят в почву и регенерируют.
Другим аспектом настоящего изобретения предусматриваются трансгенные растения или потомство этих растений, содержащее ДНК по данному изобретению. В объем настоящего изобретения входят как однодольные, так и двудольные растения. Растительные клетки трансформируют выделенной ДНК, кодирующей 6-десатуразу, с помощью любого из описанных выше методов трансформации растений. Трансформированную растительную клетку, обычно в культуре каллусных тканей или на листовом диске, регенерируют в полностью трансгенном растении с помощь методов, хорошо известных специалистам в этой области (например, Horsch et al. (1985) Science 227, 1129). В соответствии с предпочтительным вариантом осуществления изобретения трансгенным растением является подсолнечник, paпс (oie seed rape), кукуруза, табак, арахис или соя. Поскольку потомство трансформированных растений наследует ДНК, кодирующую 6-десатуразу, семена или черенки трансформированных растений используют для сохранения линии трансгенных растений.
Настоящим изобретением далее предусматривается способ получения трансгенных растений с повышенным содержанием гамма-линоленовой кислоты. Этот способ включает введение ДНК, кодирующей 6-десатуразу, в растительные клетки с низким содержанием или полным отсутствием гамма-линоленовой кислоты, которые содержат линолевую кислоту, и регенерацию растений с повышенным содержанием гамма-линоленовой кислоты из трансгенных клеток. В частности, такие широко распространенные сельскохозяйственные культуры, как соя, рапс, кукуруза, арахис и табак, рассматриваются в качестве трансгенных организмов, но не ограничиваются ими.
Настоящим изобретением далее предусматривается способ получения трансгенных организмов, содержащих гамма-линоленовую кислоту. Этот способ включает введение ДНК, кодирующей 6-десатуразу, в организмы с низким содержанием или полным отсутствием гамма-линоленовой кислоты, но содержащие линолевую кислоту. В соответствии с другим вариантом осуществления настоящего изобретения предусматривается способ, который включает введение одного или нескольких экспрессирующих векторов, содержащих ДНК, кодирующую 12-десатуразу и 6-десатуразу, в организмы, не содержащие ни гамма-линоленовой кислоты, ни линолевой кислоты. В соответствии с этим способом у организмов, не содержащих ни линолевой кислоты, ни гамма-линоленовой кислоты, вызывают синтез линолевой кислоты путем экспрессии 12-десатуразы, после чего синтезируют гамма-линоленовую кислоту в результате экспрессии 6-десатуразы. Экспрессирующие векторы, содержащие ДНК, кодирующую 12-десатуразу или 12-десатуразу и 6-десатуразу, можно получить с помощью методов рекомбинантных ДНК, известных специалистам в этой области (Sambrook et al. 1989), и с помощью последовательности 12-десатуразы, опубликованной в научной литературе (Wada et al. [1990] Nature (London) 347, 200-203). Кроме того, настоящее изобретение позволило установить, что нуклеотиды 2002-3081 последовательности с идентификационным No.1 кодируют 12-десатуразу цианобактерий. Поэтому эту последовательность можно использовать для получения целевых экспрессирующих векторов. В частности, такие широко распространенные сельскохозяйственные культуры, как подсолнечник, соя, рапс, кукуруза, арахис и табак, рассматриваются в качестве трансгенных организмов, но не ограничиваются ими.
Настоящее изобретение далее относится к способу повышения холодоустойчивости растений. Чувствительность к охлаждению может быть связана с фазовым переносом липидов в клеточных мембранах. Температура фазового переноса зависит от степени ненасыщенности жирных кислот в липидах мембраны, поэтому увеличение степени ненасыщенности, например, путем введения 6-десатуразы с целью превращения линолевой кислоты в гамма-линоленовую кислоту, может сделать растение холодоустойчивым или понизить чувствительность к охлаждению. Поэтому настоящим изобретением предусматривается введение ДНК, кодирующей 6-десатуразу, в растительные клетки и регенерация растений с повышенной холодоустойчивостью из указанных трансформированных растительных клеток. В соответствии с предпочтительным вариантом осуществления настоящего изобретения такими растениями являются подсолнечник, соя, рапс, кукуруза, арахис или табак.
Настоящее изобретение далее проиллюстрировано следующими примерами.
ПРИМЕР 1Штаммы и условия культивирования Synechocystis (РСС 6803, АТСС 27184), Anabaena (PCC 7120, АТСС 27893 и Synechoccus (PCC 7942, АТСС 33912) выращивают фотоавтотрофическим путем при 30oС в среде BG11N+(Rippka et al.[1979] J. Gen. Microbiol. 111. 1-61) под светом ламп накаливания (60  E.m-2.S-1). Отбирают космиды и плазмиды и культивируют их в штамме Escherichia соli DH5 в среде Луриа, дополненной антибиотиками в стандартных концентрациях, как описано в работе Maniatis et al. (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring, New York.
ПРИМЕР 2 E.m-2.S-1). Отбирают космиды и плазмиды и культивируют их в штамме Escherichia соli DH5 в среде Луриа, дополненной антибиотиками в стандартных концентрациях, как описано в работе Maniatis et al. (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring, New York.
ПРИМЕР 2Создание геномной библиотеки космид штамма Synechocystis Всю геномную ДНК, полученную из штамма Synechocystis (PCC 6803) частично расщепляют с помощью Sau3A и фракцинируют в градиенте концентрации сахарозы (Ausubel et al. [1987] Current Protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, New York). Фракции, содержащие фрагменты ДНК длиной от 30 до 40 т.п.н. отбирают и лигируют в дефосфорилированный сайт BamHI космидного вектора, pDUCA7 (Bui-kema et al. [1991] J. Bacteriol. 173. 1879-1885). Лигированную ДНК упаковывают in vitro, как описано Ausubel et al. (1987), и упакованный фаг культивируют в культуре E. соli DH5 , содержащей плазмиду pRL528 клетки-помощника синтеза метилазы AvaI и Есо4711, как это описано Buikema et al. (1991). Произвольно выделяют в общей сложности 1152 колонии и помещают отдельно на двенадцать 96-луночных титрационных микропланшетов.
ПРИМЕР 3Экспрессия гамма-линоленовой кислоты в Anabaena с приобретением функции Anabaena (PCC 7120), нитевидные цианобактерии, не имеют гамма-линоленовой кислоты, но содержат значительные количества линолевой кислоты, предшественника гамма-линоленовой кислоты (фигура 2; таблица 2). Библиотеку космид Synechocystis, описанную в примере 2, конъюгируют в Anabaena (PCC 7120) с целью идентификации трансконъюгантов, синтезирующих гамма-линоленовую кислоту. Клетки Anabaena выращивают до полулогарифмической фазы в жидкой среде BG11N+ и повторно суспендируют в такой же среде до конечной концентрации, равной примерно 1х109 клеткам на мл. Культуру Е. coli RP4 в полулогарифмической фазе (Burkard et al. [1979] J. Gen. Microbiol.114, 341-348), выращенную в среде Луриа, содержащей ампициллин, промывают и вновь суспендируют в свежей среде Луриа. После этого Anabaena и RP4 смешивают и равномерно распределяют по планшетам со средой BG11N+, содержащей 5% среды Луриа. Геномную библиотеку космид помещают на планшеты со средой Луриа, содержащие 50 мкг/мл канамицина и 17,5 мкг/мл хлорамфеникола, повторяя эту процедуру дважды, а затем помещают на планшеты со средой BG11N+, содержащей Anabaena и RР4. После инкубации в течение 24 ч при 30oС вводят подслой из 30 мкг/мл неомицина; и продолжают инкубацию при 30oС до появления трансконъюгатов. Отдельные трансконъюгаты отделяют после конъюгирования и выращивают в 2 мл жидкой среды BG11N+ с 15 мкг/мл неомицина. Метиловые эфиры жирных кислот получают из культур дикого типа и культур, содержащих пулы из десяти трансконъюгатов, выполняя следующую процедуру. Дикие и трансгенные цианобактериальные культуры отделяют центрифугированием и дважды промывают дистиллированной водой. Метиловые эфиры жирных кислот экстрагируют из этих культур, как это описано в работе Dahmer et al. (989) J.Amer. Oil. Chem. Soc.66, 543-548, и анализируют посредством газожидкостной хроматографии с помощью хроматографа Тrасоr-560, оборудованного ионизационным детектором с водородным пламенем и капиллярной колонкой (30мх0,25 мм связанного FSOT Superox II, Alltech Associates Inc. , IL). Для идентификации жирных кислот служат значения времени удержания и кохроматографии эталонов (предоставленных компанией Sigma Chemical Co. ). Средний состав жирных кислот определяют в виде отношений пиковой области каждой С 18 жирной кислоты, нормализованной в отношении внутреннего эталона. Типичные профили газожидкостной хроматографии показаны на фиг.2. На этой фигуре изображены метиловые эфиры С18 жирных кислот. Пики идентифицируют путем сравнения значений времени элюирования с известными эталонами метиловых эфиров жирных кислот и подтверждают путем газовой хроматографии и масс-спектрометрии. На фиг.2А показаны результаты газожидкостной хроматографии жирных кислот Anabaena дикого типа. Стрелкой показано время миграции гамма-линоленовой кислоты. На фиг. 2В изображен профиль хроматографии жирных кислот трансконъюгатов Anabaena с pAM542+1.8F. Выявлено два пула, синтезирующих гамма-линоленовую кислоту (из 25 пулов, представляющих 250 трансконъюгатов). Отдельные трансконъюгаты каждого пула, давшего положительные результаты в отношении гамма-линоленовой кислоты, анализируют на синтез гамма-линоленовой кислоты; определяют два независимых трансконъюгата, AS13 и AS75, по одному из каждого пула, которые синтезируют значительные количества гамма-линоленовой кислоты и содержат космиды, cSyl3 и cSy75 (фигура 3). Перекрывающиеся космиды образуют область длиной примерно 7,5 т.п.н. Фрагмент NheI длиной 3,5 т.п.н. космиды cSy75 вновь клонируют в векторе pDUCA7 и переносят в Anabaena, в результате чего имеет место экспрессия гамма-линоленовой кислоты с приобретением функции (таблица 2). Два субфрагмента NheI/Hind III (1,8 и 1,7 т.п.н.) из фрагмента NheI длиной 3,5 т.п.н. космиды cSy75-3,5 субклонируют в “pBLUESCRIPT” (Stratagene) (фигура 3) с целью секвенирования. Используют стандартные методы молекулярной биологии, описанные Maniatis et al. (1982) и Ausubel et а1. (1987). Дидезоксисиквенирование (Sanger et al. [1977] Proc. Natl. Acad. Sci. USA 74, 5463-5467) плазмиды pBSl, 8 выполняют с помощью “секвеназы” (United States Biochemical) в отношении обеих цепей, используя специальные олигонуклеотидные затравки, синтезированные лабораторией прогрессивных технологий синтеза ДНК (Биологический факультет. Техасский университет). Анализ последовательности ДНК производят с помощью программного обеспечения GCG (Мадисон, шт. Винсконсин) в соответствии с описанием Devereux et al. (1984) Nucleic Acids Res.12, 387-395. Субфрагменты Nhe I/HindIII переносят в конъюгальный экспрессирующий вектор АМ542 с прямой и обратной ориентацией в отношении промотора, представляющего собой карбоксилазу цианобактерий, и вводят в Anabaena путем конъюгирования. Трансконъюгаты, содержащие фрагмент длиной 1,8 т.п.н. с прямой ориентацией (АМ542-1,8F), позволяют получить значительные количества гамма-линоленовой кислоты и октадекатетраеновой кислоты (фигура 2; таблица 2). Трансконъюгаты, содержащие другие конструкции, а также обратно ориентированный фрагмент длиной 1,8 т.п.н. или прямо и обратно ориентированный фрагмент длиной 1,7 т.п.н., не синтезируют гамма-линоленовую кислоту на детектируемых уровнях (таблица 2). На фигуре 2 профиль С18 жирной кислоты экстракта из Anabaena дикого типа (фигура 2А) сравнивается с профилем Anabaena трансгенного типа, содержащим фрагмент космиды cSy75-3,5 длиной 1,8 т.п.н. с прямой ориентацией (фигура 2В). Газожидкостная хроматография метиловых эфиров жирных кислот, полученных из фрагмента АМ542-1,8А, позволила выявить пик со временем удержания, идентичным такому же показателю у подлинного эталона гамма-линоленовой кислоты. Анализ этого пика посредством газовой хроматографии и масс-спектрометрии (GC-MS) подтвердил наличие аналогичной картины фрагментации по массе, что и в контрольной пробе гамма-линоленовой кислоты. Аnаbаеnа трансгенного типа с измененными уровнями полиненасыщенных жирных кислот аналогичен дикому типу по скорости роста и морфологии. ПРИМЕР 4 Трансформация Synechococcus генами 6 и 12 десатуразыТретью космиду, cSy7, содержащую ген 12-десатуразы, выделяют путем скрининга геномной библиотеки Synechocystis олигонуклеотидом, синтезированным из опубликованной в литературе последовательности гена 12-десатуразы Synechocystis (Wada et al. [1990] Nature (London) 347, 200-203). В этой космиде идентифицируют фрагмент AvaI длиной 1,7 т. п. н. , содержащий ген 12-десатуразы и используют его в качестве зонда для демонстрации того, что космида cSyl3 помимо гена 6-десатуразы содержит также ген 12-десатуразы (фигура 3). Геномный анализ по методу саузерн-блоттинга далее показывает, что гены как 6-, так и 12-десатуразы уникальны в геноме Synechocystis в том отношении, что оба функциональных гена, имеющих отношение к десатурации С18 жирных кислот, близко связаны в геноме Synechocystis.
Одноклеточные цианобактерии Synechococcus (PCC 7942) не имеют ни ленолевой кислоты, ни гамма-линоленовой кислоты (3). Гены 12- и 6-десатуразы клонируют отдельно и вместе в шаттл-векторе рАМ854 (Bustos et al. [1991] J. Bacteriol. 174, 7525-7533), который содержит последовательности, необходимые для интеграции чужеродной ДНК в геном Synechococcus (Golden et al. [1987] Methods in Enzvmol. 153, 215-231). Synechococcus трансформируют этими генными конструкциями и отбирают колонии, трансформируют этими генными конструкциями и отбирают колонии. Метиловые эфиры жирных кислот экстрагируют из Synechococcus трансгенного типа и анализируют посредством газожидкостной хроматографии.
В таблице 2 показано, что основными жирными кислотами Synechococcus дикого типа являются стеариновая кислота (18:0) и олеиновая кислота (18:1). Цианобактерии Synechococcus, трансформированные рАМ854-12, помимо основных жирных кислот ситезируют линолевую кислоту (18:2). Трансформанты, полученные с помощью рАМ854- 6 и 12, синтезируют как линолеат, так и гамма-линоленовую кислоту (таблица 1). Эти результаты свидетельствуют о том, что цианобактерии Synechococcus, содержащие гены 12- и 6-десатуразы, способны создавать вторую двойную связь в 12-положении и третью двойную связь в 6-положении С18 жирных кислот. Однако у трансформанта, содержащего рАМ854- 6, не обнаружено изменений в составе жирных кислот, что указывает на то, что при отсутствии субстрата, синтезируемого 12-десатуразой, 6-десатураза не проявляет активности. Этот эксперемент далее подтверждает, что фрагмент NheI/Hind III длиной 1,8 т.п.н. (фигура 3) содержит как кодирующую, так и промоторную области гена 6-десатуразы Synechocystis. Цианобактерии Synechococcus трансгенного типа с измененными уровнями полиненасыщенных жирных кислот аналогичны дикому типу по скорости роста и морфологии.
ПРИМЕР 5Нуклеотидная последовательность 6-десатуразыОпределяют нуклеотидную последовательность фрагмента cSy75-3,5 длиной 1,8 т. п.н. Идентифицируют открытую рамку считывания, кодирующую полипептид 359 аминокислот (фигура 4). С помощью анализа на водорастворимость Кайта-Дулитла (Kyte et al. [1982] J.Mol. Biol. 157, 105-132) идентифицируют две области гидрофобных аминокислот, которые могут представлять трансмембранные домены (фигура 1А): кроме того, профиль водорастворимости 6-десатуразы аналогичен 12-десатуразы (1В; Wada et al.) и 9-десатураз (Thiede et al. [1986] J. Biol. Chem. 261, 13230-13235). Однако сходство последовательностей между 6- и 12-десатуразой Synechocystis составляет менее 40% на уровне нуклеотидов и примерно 18% на уровне аминокислот.
ПРИМЕР 6Перенос 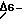 десатуразы цианобактерий в табак десатуразы цианобактерий в табакГен 6-десатуразы цианобактерий активируют в растительном экспрессирующем векторе и переносят в табак с помощью методов переноса генов, опосредственного Agrobacterium. Для гарантии того, что перенесенная десатураза должным образом экспрессирована в листьях и развивающихся семенах и что продукт гена десатуразы введен в эндоплазматический ретикулум или хлоропласт, конструируют различные полигенные экспрессирующие кластеры с открытой рамкой считывания -десатуразы Synechocystis. Компоненты этих кластеров включают: (i) промотор 35S или семяспеци-фичный промотор, полученный из гена гелиантинина подсолнечника, с целью экспрессии гена 6-десатуразы во всех тканях растения или только в развивающихся семенах, (ii) предполагаемый сигнальный пептид из гена экстенсина моркови или гена гелиантинина подсолнечника для введения вновь синтезированной 6-десатуразы в эндоплазматический ретикулум (ЕВ), (iii) сигнальную последовательность сохранения полости эндоплазматического ретикулума (KDEL) у СООН-конца открытой рамки считывания 6-десатуразы и (iv) оптимизированный транзитный пептид, служащий для ввода 6-десатуразы в хлоропласт. Промотор 35S является производным pRTL2, описанным Restrepo et al. Последовательность оптимизированного транзитного пептида описана Van de Broeck et al. (1985). Сигнальный пептид экстенсина моркови описан Chen et al. (1985) EMBO J.9, 2145.
Получают трансгенные растения табака, содержащие рекомбинантный ген десатуразы цианобактерий, представляющий собой ген 6-десатуразы Synechocystis, связанный с последовательностью сохранения эндоплазматического ретикулума (KDEL) и сигнальным пептидом экстенсина, стимулируемого промотором CaMV 35S. Амплификации геномной ДНК трансгенного табака, образуемые в результате полимеразной цепной реакции, показывают, что ген 6-десатуразы включен в геном табака. Метиловые эфиры жирных кислот, полученные из листьев этих трансгенных растений табака, экстрагируют и анализируют посредством газожидкостной хроматографии. Эти трансгенные растения табака накапливают значительные количества гамма-линоленовой кислоты (фигура 4). На фигуре 4 показаны метиловые эфиры жирных кислот, выявленные газожидкостной хроматографией. Пики идентифицированы путем сравнения времени элюирования с известными эталонами метиловых эфиров жирных кислот. Таким образом, цианобактериальные гены, участвующие в обмене жирных кислот, можно использовать для генерации трансгенных растений с измененными составами жирных кислот.
ПРИМЕР 7. Создание библиотеки кДНК бурачника.
Мембраносвязанные полирибосомы выделяют из семян бурачника через 12 дней после опыления в соответствии с протоколом, разработанным для гороха Латкинсом (Latkins) и Девисом (Davies) (1975 Plant Phys. 55:749-756). РНК экстрагируют из полирибосом по методу, описанному Мехлером (Mechler) (1987 Methods in Enzymology 152:241-248. Academic Press).
Поли-А+ РНК выделяют из РНК мембраносвязанных полирибосом, используя гранулы Oligotex-dT (Qiagen). Соответствующую кДНК получают, используя набор для синтеза кДНК ZAP фирмы Stratagene. Библиотеку кДНК создают в векторе ламбда ZAP II (Stratagene), используя набор для синтеза вектора ламбда ZAP II. Первичную библиотеку упаковывают в упаковочный экстракт Gigapack II Gold (Stratagene). Эту библиотеку используют для синтеза меток экспрессированных последовательностей (EST) и последовательности, соответствующие этим меткам, используют для сканирования базы данных GenBank.
ПРИМЕР 8. Протокол гибридизации.
Зонды гибридизации для скрининга библиотеки кДНК бурачника получают путем синтеза случайной праймированной ДНК, описанного Ausubel et al. (1994 Current Protocols in Molecular Biology, Wiley Interscience, N.Y.), при этом они соответствуют ранее идентифицированной кДНК экспрессированного запасного белка семян. Невключенные нуклеотиды удаляют в осадительной колонке G-50 (Boehringer Manheim). Зонд денатурируют для гибридизации путем кипячения на водяной бане в течение 5 минут с последующим быстрым охлаждением на льду. Фильтры для гибридизации предварительно гибридизируют при 60oС в течение 2-4 ч в растворе для предгибридизации (6-кратный раствор хлорида и цитрата натрия (SSC) [Maniatis et al. 1984 Molecular Cloning A Laboratory Manual, Cold Spring Harbor Laboratory] , содержащий 1-кратный раствор Денхартса, 0,05% пиро-фосфата натрия, 100 мкг/мл денатурированной ДНК спермы лосося). Денатурированный зонд добавляют в раствор гибридизации (6-кратный раствор хлорида и цитрата натрия, 1-кратный раствор Денхарта, 0,05% пирофосфата натрия, 100 мкг/мл денатурированной ДНК спермы лосося) и инкубируют, перемешивая при 60oС в течение ночи. Фильтры промывают в 4-, 2-и 1-кратных растворах SET по 15 мин в каждом при 60oС. 20-кратный исходный раствор SET состоит из 3 моль NaCl, 0,4 моль трис-основания, 20 ммоль Nа2ЭДТК-2Н2O. 4-кратный промывочный раствор SET состоит из 4-кратного раствора SET, 12,5 ммоль РO4, рН 6,8 и 0,2% додецилсульфата натрия. 2-кратный промывочный раствор SET состоит из 2-кратного раствора SET, 12,5 ммоль РO4, рН 6,8 и 0,2% додецилсульфата натрия. 1-кратный промывочный раствор SET состоит из 1-кратного раствора SET, 12,5 ммоль НO4, рН 6,8 и 0,2% додецилсульфата натрия. Фильтры сушат на воздухе, а затем подвергают воздействию рентгеновской пленки в течение 24 ч с использованием усиливающих экранов при температуре -80oС.
ПРИМЕР 9. Произвольное секвенирование кДНК, выделенных из библиотеки мембраносвязанных полисом семян бурачника (через 12 дней после опыления)Библиотеку кДНК бурачника пассируют на чашках с низкой плотностью (500 бпяшкообразующих единиц на 150 мм чашках Петри). Наиболее распространенные кДНК запасного белка семян вычленяют путем скрининга с использованием ранее идентифицированных соответствующих кДНК. Негибридизирующие бляшки вырезают, используя процедуру вырезания и реагенты фирмы Stratagene. Полученные бактериальные колонии используют для инокуляции жидких культур и секвенируют либо вручную, либо с помощью автоматического секвенатора ABI. Каждую кДНК секвенируют один раз и метят последовательности, состоящие из 200-300 пар оснований. Все секвенирование выполняют по методу циклического секвенирования (Epicentre). Получено более 300 меток экспрессированных последовательностей. Каждую метку последовательности сравнивают с базой данных GenBank с помощью компьютерной программы BLASTX и идентифицируют ряд генов липидного обмена, включая 6-десатуразу.
Поиски в базе данных с использованием клона кДНК, обозначенного mbp-65, с помощью программы BLASTX и базы данных Gem-Bank привели к выявлению значительной совместимости с 6-десатуразой Synechocystis. Однако установлено, что этот клон не является полной кДНК. Полную кДНК выделяют, используя mbp-65 для скрининга библиотеки мембраносвязанных полисом бурачника. Определяют последовательность выделенной кДНК (фиг. 5A, последовательность с идентификационным No.4) и белковую последовательность открытой рамки считывания (фиг. 5В, последовательность с идентификационным No.5) сравнивают с другими известными десатуразами, используя программу сравнительного анализа первичной структуры белков Geneworks ((IntelligGenetics) фиг.2). Этот анализ показывает, что данная кДНК является геном 6-десатуразы бурачника.
Будучи схожей с известными десатуразами других растений, 6-десатураза бурачника имеет определенные отличия, показанные на дендрограмме, изображенной на фиг. 6. Кроме того, сравнение аминокислотных последовательностей, характерных для десатураз, в частности тех, которые участвуют в связывании металлов (металлический блок 1 и металлический блок 2), позволяет выявить различия, существующие между 6-десатуразой бурачника и десатуразами других растений (таблица 3).
6-Десатураза бурачника отличается от цианобактериальной формы не только всей последовательностью (фиг.6), но также и липидным блоком, металлическим блоком 1 и металлическим блоком 2 аминокислотных частей (таблица 3). Как показано в таблице 3, все три части являются новыми в последовательности. Только металлический блок 2 6-десатуразы бурачника имеет некоторое сходство с металлическим блоком 2 6-десатуразы Synechocystis.
Помимо этого, 6-десатураза бурачника отличается также от другого гена десатуразы бурачника, 12-десатуразы. Р1-81 является полной кДНК, которая идентифицирована на основе анализа меток экспрессированных последовательностей, и демонстрирует большое сходство с 12-десатуразой Arabidopsos (Fad 2).
Сравнение липидного блока, металлического блока 1 и металлического блока 2 аминокислотных частей (таблица 3) у 6- и 12-десатураз бурачника показывает, что в этих областях имеет место незначительная степень подобия. Расположение этих двух последовательностей на дендрограмме, изображенной на фиг.6, показывает, насколько далеки эти два гена друг от друга.
ПРИМЕР 10. Конструкция 222.16NOS для переходной экспрессии.
Вектор рВ1221 (Jefferson et al. 1987 EMBO J. 6:3901-3907) получают для лигирования расщеплением с помощью BamHI EcoICR I (Promega), что позволяет вырезать кодирующую область GUS, не повреждая промотор 35S и терминатор NOS. кДНК 6-десатуразы бурачника вырезают из плазмиды Блюскрипта (Stratagene) путем расщепления с помощью BamHI XhoI. Конец XhoI дефосфолируют, используя фрагмент Кленова. Этот фрагмент затем клонируют в сайтах BamHI/EcoICR I вектора рВI221, получая 221.6NOS (фиг. 7). В 221.6NOS стальной частью (скелетом) рестрикционной карты, изображенной на фиг.7, является вектор рВ1221.
ПРИМЕР 11. Конструкция 121.6.NOS для стабильной трансформации.
Вектор pBI121 (Jefferson et al. 1987 EMBO J.6:3901-3907) получают для лигирования расщеплением с помощью BamHI и EcoICR I (Promega), что позволяет вырезать кодирующую область GUS, не повреждая промотор 35S и терминатор NOS. кДНК 6-десатуразы бурачника вырезают из плазмиды Блюскрипта (Stratagene) путем расщепления с помощью BamHI и XhoI. Конец XhoI дефосфолируют, используя фрагмент Кленова. Этот фрагмент затем клонируют в сайтах BamHI/EcoICR I вектора pBI121, получая 121.6NOS (фиг.7). В 121.6NOS остальной частью (скелетом) рестрикционной карты, изображенной на фиг.7, является вектор pBI121.
ПРИМЕР 12. Переходная экспрессия.
Всю работу, связанную с протопластами, выполняют в стерильном вытяжном шкафу. Один мл упакованных клеток суспензии моркови расщепляют в 30 мл плазмолизующего раствора (25 г/л КС1, 3,5 г/л CaCl2-H2O, 10 ммоль MES, рН 5,6 и 0,2 моль маннитола) с 1% целлюлозы, 0,1% пектолиазы и 0,1% дрейсалазы в течение ночи в темноте при комнатной температуре. Высвобожденные протопласты фильтруют через сито с отверстиями 150 m и гранулируют центрифугированием (100хг, 5 мин), после чего дважды промывают в плазмирующем растворе. Протопласты подсчитывают в двухкамерном гемоцитомере. ДНК трансфецируют в протопласты путем обработки полиэтиленгликолем в соответствии с описанием Нунберга и Томаса (Nunberg and Tomas, 1993, Methods in Plant Molecular Biology and Biotechnology. B.R. Glick and J.E. Thompson, eds., pp.241-248), используя 106 протопластов и 50-70 мкг плазмидной ДНК (221.6.NOS). Протопласты культивируют, встряхивая, в 5 мл MS-среды, дополненной 0,2 моль маннитола и 3 мкл 2,4-дихлорфеноксиуксусной кислоты, в темноте в течение 48 чПРИМЕР 13. Стабильная трансформация табака. Конструкцию на основе плазмидного вектора 121. 6.NOS используют для трансформации табака (Nicotiana tabacum cv. xanthi) посредством Agrobacterium в соответствии со стандартными процедурами (Horsh et al., 1985, Science 227: 1229-1231; Bogue et al., 1990 Mol. Genet. 221: 49-57) за исключением того, что исходные трансформанты выбирают на 100 мкг/мл канамицина.
ПРИМЕР 14. Получение и анализ метиловых эфиров жирных кислот (FAME).
Ткани, взятые из трансфецированных протопластов и трансформированных растений табака, замораживают в жидком азоте и лиофилизуют в течение ночи. Метиловые эфиры жирных кислот получают в соответствии со способом, описанным Дагмером и др. (Dahmer et al., 1989, J. Amer. Oil Chem. Soc. 66: 543-548). В некоторых случаях растворитель снова выпаривают и метиловые эфиры жирных кислот еще раз суспендируют в этилацетате и один раз экстрагируют деионизированной водой для удаления водорастворимых загрязняющих примесей. Метиловые эфиры жирных кислот анализируют посредством газовой хромотографии на колонке J & W Scientific DB-wax (длина 30 м, внутренний диаметр 0,25 мм, пленка толщиной 0,25 мкм).
Пример анализа переходного состояния приведен на фиг.8, который позволяет выявить совокупность из трех независимых трансфекций. Добавление кДНК 6-десатуразы бурачника соответствует появлению гамма-линоленовой кислоты, которая является одним из возможных продуктов 6-десатуразы.
На фигурах 9 и 10 изображены профили газовой хроматографии эфиров жирных кислот, выделенных соответственно из тканей листа и семени контрольных и трансформированных растений табака. На фигуре 9А изображен профиль ткани листа табака дикого типа (xanthi); на фигуре 9В изображен профиль ткани листа табака, трансформированного с помощью 6-десатуразы бурачника при транскрипционной регуляции, осуществляемой промотором 35S CaMV (pBI 1216NOS). Пики соответствуют 18:2, 18:3 (гамма-линоленовая кислота), 18:3 и 18: 4 (октадеканоновая кислота). На фигуре 10А показан профиль газовой хроматографии семян табака дикого типа; на фигуре 10В изображен профиль ткани семени растения табака, трансформированного с помощью pBI 1216NOS. Пики соответствуют 18:2, 18:3 (гамма-линоленовая кислота) и 18:3.Относительное распределение C18 жирных кислот в контрольных и трансгенных семенах табака показано в таблице 4. Приведенные результаты показывают, что гамма-линоленовая кислота включена в триацилглицериды из семян трансгенного табака, содержащего 6-десатуразу бурачника.
Формула изобретения
РИСУНКИ
PC4A Государственная регистрация перехода исключительного права без заключения договора
Дата и номер государственной регистрации перехода исключительного права: 16.11.2010 № РП0001115
Лицо(а), исключительное право от которого(ых) переходит без заключения договора:
(73) Патентообладатель(и):
Адрес для переписки:
Дата публикации: 10.01.2011
|
||||||||||||||||||||||||||