Патент на изобретение №2169009
|
||||||||||||||||||||||||||
(54) СПОСОБЫ ИНДУЦИРОВАНИЯ Т-КЛЕТОЧНОЙ ТОЛЕРАНТНОСТИ К ТКАНЕВОМУ ИЛИ ОРГАННОМУ ТРАНСПЛАНТАТУ
(57) Реферат: Изобретение относится к медицине и касается способов индуцирования Т-клеточной толерантности к тканевому или органному трансплантату. Сущность изобретения: способы включают введение субъекту 1) аллогенной или ксеногенной клетки, которая экспрессирует донорские антигены и которая имеет лиганд на клеточной поверхности, который взаимодействует с рецептором на поверхности реципиентной клетки, которая опосредует контакт-зависимую хелпер-эффекторную функцию и 2) антагониста рецептора, который ингибирует взаимодействие лиганда с рецептором. Преимущество изобретения состоит в индукции толерантности к донорским антигенам и создает возможность для успешной трансплантации донорской ткани или органа без иммуноопосредованного отторжения донорского трансплантата. 3 с. и 35 з.п. ф-лы, 2 табл., 7 ил. Для индуцирования антиген-специфической T-клеточной активации и клональной экспансии к поверхности покоящихся T-лимфоцитов должны быть доставлены два сигнала, обеспеченных антигенпредставляющими клетками (АПК) (Jenkins M. and Schwartz R. (1987) J.Exp.Med. 165, 302-319; Mueller D.L. et al. (1990) J. Immunol. 144, 3701-3709; Williams I.R. and Unnanue E.R. (1990) J. Immunol. 145, 85-93). Первый сигнал, который представляет специфичность иммунной реакции, передается через T-клеточный рецептор (TCR) после распознавания инородного атигенного пептида, представленного в связи с главным комплексом гистосовместимости (ГКГС). Второй сигнал, называемый костимуляцией, индуцирует T-клетки к пролиферации и появлению функциональности (Schwartz R. H. (1990) Science 248, 1349-1356). Костимуляция не является ни антиген-специфической, ни ограниченной ГКГС и, полагают, обеспечивается одной или несколькими отдельными молекулами клеточной поверхности, экспрессированными АПК (Jenkins M. K. et al. (1988) J. Immunol. 140, 3324-3330; Linsley P. S. et al. (1991) J. Exp. Med. 173, 721-730; Gimmi C.D. et al. (1991) Proc. Natl. Acad. Sci. USA. 88, 6575-6579; Yong J.W. et al. (1992) J. Clin. Invest. 90, 229-237; Koulova L. et al. (1991) J. Exp. Med. 173, 759-762; Reiser H. et al. (1992) Proc. Natl. Acad. Sci. USA. 89, 271-275; van-Seventer G.A. et al. (1990) J. Immunol. 144, 4579-4586; LaSalle J.M. et al. (1991) J. Immunol. 147, 774-780; Dustin M.I. et al. (1989) J. Exp. Med. 169, 503; Armitage R.J. et al. (1992) Nature 357, 80-82; Liu Y. et al. (1992) J. Exp. Med. 175, 437-445). Один костимуляторный каскад, включенный в T-клеточную активацию, включает молекулу CD28 на поверхности T-клеток. Эта молекула может получить костимуляторный сигнал, доставленный лигандом на B-клетках или других АПК. Лиганды для CD28 включают членов семейства B7 B-лимфоцитных активационных антигенов, таких как B7-1 и/или B7-2 (Freedman A.S. et al. (1987) J. Immunol. 137, 3260-3267; Freeman G.J. et al. (1989) J. Immunol. 143, 2714-2722; Freeman G.J. et al. (1991) J. Exp. Med. 174, 625-631; Freeman G. J. et al. (1993) Science 262, 909-911; Azuma M. et al. (1993) Nature 366, 76-79; Freeman G.J. et al. (1993) J. Exp. Med. 178, 2185-2192). B7-1 и B7-2 также являются лигандами для другой молекулы – CTLA4, присутствующей на поверхностях активированных T-клеток, хотя роль CTLA4 при костимуляции неясна. Доставка к T-клетке антиген-специфического сигнала костимуляторным сигналом ведет к активности T-клеток, которая может включать как пролиферацию T-клеток, так и секрецию цитокинов. В противоположность этому, полагают, что доставка к T-клетке антиген-специфичного сигнала в отсутствии костимуляторного сигнала индуцирует в T-клетке состояние иммунологической неотвечаемости или анергии, индуцируя, таким образом, антиген-специфическую толерантность в T-клетке. Взаимодействие между T-клетками и B-клетками играет центральную роль в иммунных реакциях. Индукция гуморального иммунитета к тимус-зависимым антигенам требует “помощи”, обеспечиваемой T-клетками-хелперами (далее – Th-клетками). Хотя некоторая помощь, обеспеченная B-лимфоцитами, передается растворимыми молекулами, выделяемыми Th-клетками (например, лимфокинами, такими как IL-4 и IL-5), активация B-клеток также требует контакт-зависимого взаимодействия между B-клетками и Th-клетками. См. Hirohata et al. J. Immunol. , 140: 3736-3744 (1988); Barlett et al., J. Immunol., 143: 1745-1754 (1989). Это указывает, что активация B-клеток включает обязательное взаимодействие молекул клеточной поверхности B-клеток и Th-клеток. Следовательно, молекула (молекулы) на T-клетке опосредуют контакт-зависимые хелпер-эффекторные функции T-клетки. Контакт-зависимое взаимодействие между молекулами на B-клетках и T-клетках также поддерживает наблюдение, что изолированные плазматические мембраны активированных T-клеток могут обеспечивать хелперные функции, необходимые для активации B-клеток. См. Brian, Proc. Natl. Acad. Sci. USA, 85: 564-568 (1988); Hodgkin et al., J. Immunol., 145: 2025-2034 (1990); Noelle et al., J. Immunol., 146: 1118-1124 (1991). Молекула CD40 идентифицирована на поверхностях незрелых и зрелых B-лимфоцитов и при перекрестном сшивании антителами индуцирует пролиферацию B-клеток. См. Valle et al., Eur. J. Immunol., 19: 1463-1467 (1989); Gordon et al., J. Immunol., 140: 1425-1430 (1988); Gruber et al., J. Immunol., 142: 4144-4152 (1989). Молекулы CD40 клонированы и охарактеризованы. См. Stamenkovic et al. , EMBO J. , 8: 1403-1410 (1989). Лиганд для CD40 – gp39 (называемый также CD40-лигандом или CD4OL) – также клонирован молекулярно и охарактеризован. См. Armitage et al., Nature, 357: 80-82 (1992); Lederman et al. , J. Exp. Med., 175: 1091-1101 (1992); Hollenbaugh et al., EMBO J., 11: 4313-4319 (1992). Белок gp39 экспрессируется на активированных, а не на покоящихся, CD4+ – Th-клетках. См. Spriggs et al., J. Exp. Med., 176: 1543-1550 (1992); Lane et al., Eur. J. Immunol., 22: 2573-2578 (1992); Roy et al., J. Immunol. , 151: 1-14 (1993). Клетки, трансфицированные геном gp39, и экспрессирующие белок gр39 на своей поверхности, могут вызывать B-клеточную пролиферацию и вместе с другими стимуляторными сигналами могут индуцировать продуцирование антител. См. Armitage et al., Nature, 357: 80-82 (1992); Hollenbaugh et al., EMBO J., 11: 4313-4319 (1992). Краткое изложение сущности изобретения Молекулы клеточной поверхности, которые опосредуют контакт-зависимые хелпер-эффекторные функции T-клеток, важны для индуцирования иммунных реакций, которые требуют помощи T-клеток. Например, взаимодействие gp39 на T-клетках с CD40 на B-клетках играет центральную роль при активации реакций B-клеток на антиген. Настоящее изобретение основывается, по крайней мере частично, на открытии, что молекулы клеточной поверхности, которые опосредуют контакт-зависимые хелпер-эффекторные функции T-клеток, также играют критическую роль в реакции T-клеток к аллоантигенам. В частности, обнаружено, что при определенных условиях вмешательство во взаимодействии gр39 с лигандом на аллогенной клетке, которая представляет аллоантигены T-клетке, может индуцировать толерантность в T-клетке. Предпочтительно, аллогенная клетка, которая представляет аллоантигены T-клетке, требует взаимодействия между лигандом gp39 на T-клетке, чтобы иметь возможность обеспечить сигналы, необходимые для активации T-клетки. Ингибирование взаимодействия лиганда gp39 на аллогенной клетке с gp39 на T-клетке предотвращает активацию T-клеток и индуцирует, скорее, аллоантиген-специфическую толерантность T-клеток. Индуцирование T-клеточной толерантности к аллоантигенам, как описывается в настоящем изобретении, может использоваться в качестве препаративной схемы для пересадки ткани или органа. Соответственно, способы настоящего изобретения в особенности пригодны для индуцирования T-клеточной толерантности к донорской ткани или органу у реципиента ткани или органа. Упомянутые способы включают введение реципиенту трансплантата 1) аллогенных или ксеногенных клеток, которые экспрессируют по крайней мере один донорский антиген и которые имеют лиганд на клеточной поверхности, который взаимодействует с рецептором на поверхности реципиентной T-клетки, которая опосредует контакт-зависимые хелпер-эффекторные функции; 2) антагониста молекулы на поверхности реципиентной T-клетки, которая опосредует контакт-зависимые хелпер-эффекторные функции. Антагонист ингибирует взаимодействие между молекулой на T-клетке и ее лигандом на аллогенной или ксеногенной клетке. В предпочтительном варианте осуществления изобретения рецептор на поверхности T-клетки, которая опосредует контакт-зависимые хелпер-эффекторные функции, представляет собой gp39. При таком варианте осуществления изобретения антагонист представляет собой молекулу, которая ингибирует взаимодействие gp39 на T-клетке с лигандом gp39 на аллогенной или ксеногенной клетке. Особенно предпочтительный антагонист gp39 представляет собой антитело против gp39. В другом варианте осуществления настоящего изобретения антагонист gp39 представляет собой растворимую форму лиганда gp39, например растворимый CD40. Аллогенные или ксеногенные клетки, которые вводятся реципиенту, представляют собой предпочтительно лимфоидные клетки, например B-клетки. Альтернативно, аллогенные или ксеногенные клетки представляют собой малые покоящиеся B-клетки. Аллогенные или ксеногенные клетки и антагонист обычно вводят реципиенту перед пересадкой реципиенту ткани или органа. Например, лимфоидные клетки (например, B-клетки) от донора ткани или органа вводят реципиенту вместе с антагонистом до трансплантации реципиенту ткани или органа. Способы настоящего изобретения могут применяться, например, для индуцирования T-клеточной толерантности к пересаженным тканям или органам, таким как печень, почка, сердце, легкое, кожа, мышца, нервная ткань, желудок и кишки. В одном из вариантов осуществления настоящего изобретения трансплантированная ткань содержит панкреатические островки. Соответственно, изобретение относится к способу лечения диабета, включающему введение пациенту, нуждающемуся в таком лечении, 1) аллогенных или ксеногенных клеток, которые экспрессируют донорские антигены; 2) антагониста рецептора на поверхности T-клеток реципиента, которые опосредуют контакт-зависимые хелпер-эффекторные функции, такого как антагонист gp39 (например, антитело против gp39); и 3) донорских панкреатических островков. Краткое описание рисунков На фиг. 1 приводится графическое изображение срока жизнеспособности аллотрансплантатов панкреатических островков, пересаженных мышам с “химическим” диабетом, которым предварительно ввели одно антитело против gp39 или предварительно ввели одни нефракционированные или фракционированные аллогенные клетки селезенки. Фиг. 2А и фиг. 2Б являются графическими изображениями срока жизнеспособности трансплантатов панкреатических островков, пересаженных мышам с “химическим” диабетом, которые предварительно получали однократную дозу фракционированных аллогенных клеток селезенки вместе с лечением антителом против gp39 (MR1) либо в течение 2 недель (фиг. 2А), либо в течение 7 недель (фиг. 2Б), при этом срок жизнеспособности определяют по снижению концентрации глюкозы в плазме. Каждая кривая представляет результаты, полученные для отдельной мыши. Незаштрихованные значки относятся к реципиентам, у которых островковые аллотрансплантаты самопроизвольно переставали действовать. Заштрихованные значки относятся к мышам, у которых трансплантаты функционировали при окончании эксперимента. На фиг. 3А, 3Б и 3В изображаются профили проточной цитометрии при окрашивании активированных в течение 6 часов человеческих лимфоцитов периферической крови с одним из CD40g (фиг. 3А), mAb (монокл. антитело) 4D9-8 (фиг. 3Б) или mAb 4D9-9 (фиг. 3В). На фиг. 4А, 4Б и 4В изображаются профили проточной цитометрии при окрашивании активированных в течение 6 часов человеческих лимфоцитов периферической крови, выращенных в присутствии циклопорина А, окрашенных одним из mAb 4D9-8 (фиг. 4а), mAb 4D9-9 (фиг. 4Б) или CD40Ig (фиг. 4В). На фиг. 5А и 5Б изображаются профили проточной цитометрии при окрашивании активированных в течение 6 часов человеческих лимфоцитов периферической крови CD40Ig в присутствии немеченного mAb 4D9-8 (фиг. 5А) или немеченного mAb 4D9-9 (фиг. 5Б). На фиг. 6 приводятся графики ингибирования пролиферации человеческих B-клеток растворимым gp39 и IL-4, когда клетки выращиваются в присутствии моноклональных антител 4D9-8, 4D9-9, 24-31, 24-43, 89-76 или 89-79 к человеческому gp39. На фиг. 7 графически изображается ингибирование реакции алло-специфической смешанной культуры лимфоцитов, когда клетки выращиваются в присутствии моноклональных антител 24-31 или 89-79 против человеческого gp39. Подробное описание изобретения Настоящее изобретение относится к способам индуцирования in vivo T-клеточной толерантности к донорскому трансплантату ткани или органа у реципиента трансплантата. Упомянутые способы включают введение реципиенту трансплантат 1) аллогенных или ксеногенных клеток, которые экспрессируют донорские антигены и которые имеют лиганд на клеточной поверхности, который взаимодействует с рецептором на поверхности реципиентной T-клетки, которая опосредует контакт-зависимые хелпер-эффекторные функции; 2) антагониста рецептора на поверхности T-клетки, который ингибирует взаимодействие лиганда и рецептора. Как здесь установлено, термин “реципиент” относится к субъекту, которому должны пересаживаться ткань или орган, пересаживаются ткань или орган или пересажены ткань или орган. Как здесь установлено, “аллогенные” клетки получают от различных индивидуумов того же вида, что и реципиент, и они экспрессируют “аллоантигены”, которые отличаются от антигенов, экспрессированных клетками реципиента. “Ксеногенные” клетки получают от видов, иных, чем реципиент, и они экспрессируют “ксеноантигены”, которые отличаются от антигенов, экспрессированных клетками реципиента. Как здесь установлено, термин “донорские антигены” относится к антигенам, экспрессированным донорской тканью или донорским органом, которые трансплантируют реципиенту. Донорские антигены могут быть аллоантигенами или ксеноантигенами, в зависимости от источника трансплантата. Аллогенные или ксеногенные клетки, введенные реципиенту как часть схемы вызывания толерантности, экспрессируют донорские антигены, т. е. экспрессируют некоторые или все те же антигены, присутствующие на донорской ткани или органе, которые трансплантируют. Аллогенные или ксеногенные клетки получают предпочтительно от донора тканевого или органного трансплантата, но их можно получать из одного или нескольких источников, имеющих с донором общие антигенные детерминанты. Кроме аллогенных и ксеногенных клеток, реципиенту в качестве части схемы вызывания толерантности вводят антагонист молекулы на T-клетках, которая опосредует контакт-зависимые хелпер-эффекторные функции. Как установлено здесь, молекула или рецептор, который опосредует контакт-зависимые хелпер-эффекторные функции, представляет собой молекулу или рецептор, который экспрессируется на Th-клетке и взаимодействует с лигандом на эффекторной клетке (например, на B-клетке), при этом взаимодействие молекулы с ее лигандом является необходимым для генерации реакции эффекторной клетки (т.е. активации B-клетки). Теперь обнаружено, что, помимо вовлечения в реакции эффекторных клеток, такая молекула или рецептор вовлекается в реакцию T-клетки к антигену. Предпочтительно молекула на T-клетке, которая опосредует контакт-зависимую хелпер-эффекторную функцию, представляет собой gp39. Соответственно, в предпочтительных вариантах осуществления изобретения способы изобретения включают введение реципиенту трансплантата аллогенных или ксеногенных клеток и антагониста gp39. Активация реципиента T-клеток аллогенными и ксеногенными клетками включает взаимодействие между gp39 на реципиентных T-клетках и gp39 на аллогенных или ксеногенных клетках. При ингибировании этого взаимодействия антагонистом gp39 T-клетки реципиента не активируются донорскими антигенами, экспрессированными аллогенными и ксеногенными клетками, но скорее в них вызывается толерантность к донорским антигенам. Индукция толерантности к донорским антигенам у реципиента создает возможность, таким образом, для успешной трансплантации донорской ткани или органа без иммуноопосредованного отторжения донорского трансплантата. Различные аспекты настоящего изобретения подробнее описываются в следующих разделах. I. Антагонисты gp39 В соответствии со способами изобретения, реципиенту вводят антагонист gp39, чтобы воспрепятствовать взаимодействию gp39 на T-клетках реципиента с лигандом gp39 на аллогенных и ксеногенных клетках, таких как B-клетки, введенных реципиенту. Агонист gp39 определяется как молекула, которая препятствует этому взаимодействию. Агонист gp39 может представлять собой антитело, направленное против gp39 (например, моноклональное антитело против gp39), фрагмент или производное антитела, направленного против gp39 (например, фрагменты Fab или F(ab’)2, химерные антитела или очеловеченные антитела) растворимые формы лиганда gp39 (например, растворимый CD40), растворимые формы слитого белка лиганда gp39 (например, растворимый CD40Ig), или фармацевтические средства, которые разрушают или препятствуют взаимодействию gp39 – CD40. А. Антитела Млекопитающее (например, мышь, хомяк или кролик) может быть иммунизировано иммуногенной формой белка gp39 или фрагментом белка (например, пептидным фрагментом), которая выявляет образование антител у млекопитающего. Клетка, которая экспрессирует gp39 на своей поверхности, может также использоваться в качестве иммуногена. Альтернативные иммуногены включают очищенный белок gp39 или фрагменты белка. Очистку gp39 от экспрессирующей gp39 клетки можно осуществить стандартными методами очистки. Кроме того, в клетке-хозяине может быть экспрессирована кДНК gp39 (Armitage et al., Nature, 357: 80-82 (1992); Lederman et al., J. Exp. Med., 175: 1091-1101 (1992); Hollenbaugh et al., EMBO J., 11: 4313-4319 (1992)), например, в линии бактериальных клеток или клеток млекопитающего, и gp39 очищают от клеточных культур стандартными методами. Альтернативно, с использованием известных методов (например, F-moc или T-boc химического синтеза) на основе аминокислотной последовательности gp39 (описанной в Armitage et al., Nature, 357: 80-82 (1992); Lederman et al., J. Exp. Med., 175: 1091-1101 (1992); Hollenbaugh et al. , EMBO J., 11: 4313-4319 (1992)) могут быть синтезированы пептиды gp39. Методы предоставления иммуногенности белку включают конъюгирование с носителями или другие технические приемы, хорошо известные в технике. Например, белок может быть введен в присутствии адъюванта. Развитие иммунизации может быть проконтролировано путем обнаружения титров антител в плазме или сыворотке. Для оценки уровней антител могут использоваться стандартные ELISA или другие методы иммуноанализа с иммуногеном в качестве антигена. После иммунизации могут быть получены антисыворотки и, если желательно, из сывороток можно выделить поликлональные антитела. Для получения моноклональных антител продуцирующие антитела клетки (лимфоциты) могут быть собраны у иммунизированного животного и слиты с миеломными клетками стандартными методами слияния соматических клеток, при этом эти клетки иммортализируются и образуются клетки гибридомы. Такие методы хорошо известны в технике. Например, гибридомная техника, первоначально разработанная Kohler и Milstein (Nature (1975) 256: 495-497), а также разработаны другие технические приемы, такие как гибридомный метод с человеческими B-клетками (Kozbar et al., Immunol. Today (1983) 4:72), EBV-гибридомный способ для продуцирования человеческих моноклональных антител (Cole et al., Monoclonal Antibodies in Cancer Therapy (1985) (Allen R. Bliss, Inc., pages 77-96), и скрининг библиотек химерных антител (Huse et al., Science (1989) 246: 1275). Клетки гибридом для получения антител, специфически реактивных к белку или пептиду, могут быть скринированы иммунохимическим способом, и моноклональные тела выделены. Используемый здесь термин “антитело” предполагается и для обозначения его фрагментов, которые являются специфически реактивными в отношении белка gp39, или его пептида, или слитного белка gp39. Антитела могут быть фрагментированы с использованием известных методов, и фрагменты скринируют для применения таким же способом, какой описан выше для целых антител. Например, фрагменты F(ab’)2 могут быть генерированы обработкой антител пепсином. Получающийся в результате фрагмент F(ab’)2 может быть обработан для восстановления дисульфидных мостиков с образованием фрагментов Fab’. Также предполагается, что антитела в настоящем изобретении включают биспецифические и химерные молекулы, имеющие часть анти-gp39. Когда антитела, продуцированные в субъектах, не относящихся к человеку, используются для лечения людей, они распознаются с переменным успехом как инородные, и у пациента может быть вызвана иммунная реакция. Один из подходов для снижения до минимума или устранения этой проблемы, которому следует отдать предпочтение при общей иммуносупрессии, состоит в продуцировании производных химерных антител, т.е. молекул антител, которые комбинируют вариабельную область животного, не относящегося к человеку, и человеческую константную область. Молекулы химерных антител могут включать, например, антиген-связывающий домен из антитела мыши, крысы или другого вида, и человеческие константные области. Описано множество подходов к получению химерных антител, и они могут применяться для получения химерных антител, содержащих вариабельную область иммуноглобулина, которая распознает gp39. См., например, Morrison et al., Proc. Natl. Acad. Sci U.S.A. 81: 6851 (1985); Takeda et al., Nature 314: 452 (1985), Cabilly et al., патент США N 4 816 557; Boss et al. , патент США N 4 816 397; Tanaguchi et al., публикация европейского патента EP 1 711 496; публикация европейского патента 0 173 494, патент Соединенного королевства GB 2 177 096B. Ожидается, что такие химерные антитела будут у человека менее иммуногенными, чем соответствующие нехимерные антитела. Для целей лечения людей моноклональные или химерные антитела, специфически реакционноспособные с белком gp39 или пептидом, затем могут быть очеловечены посредством продуцирования химер человеческой вариабельной области, в которых части вариабельных областей, особенно консервативные остовные области антиген-связывающего домена, имеют человеческое происхождение, и только гипервариабельные участки имеют нечеловеческое происхождение. Такие измененные иммуноглобулиновые молекулы могут быть получены любым из нескольких методов, известных в технике (см., например. Teng et al., Proc. Natl. Acad. Sci. U.S.A., 80: 7308-7312 (1983); Kozbor et al., Immunology Today, 4: 7279 (1983); Olsson et al., Meth. Enzymol., 92: 3-16 (1982)), и предпочтительно их получают по рекомендациям публикации PCT ВОИС 92/06193 или EP 0 239 400. Очеловеченные антитела могут быть произведены коммерчески, например Scotgen Limited, 2 Holly Road, Twickencham, Middlesex, Великобритания. Другой способ генерирования специфических антител, или фрагментов антител, реактивных против белка или пептида gp39, состоит в скринировании библиотек экспрессии кодирующих иммуноглобулиновых генов или их частей, экспрессированных в бактериях, белком или пептидом gp39. Например, полные Fab-фрагменты, VH-области и FV-области могут быть экспрессированы в бактериях с использованием фаговых библиотек экспрессии. См., например, Ward et al., Nature, 341: 544-546 (1989); Huse et al., Science, 246: 1275-1281 (1989); и McCafferty et al., Nature, 348: 552-554 (1990). Скринирование таких библиотек, например пептидом gp39, может идентифицировать фрагменты иммуноглобулина, реагирующие с gp39. Альтернативно, для продуцирования антител или их фрагментов может быть использована SCID-hu-мышь (доступны от Genpharm). Методология продуцирования моноклональных антител, направленных против gp39, включая человеческий gp39 и мышиный gp39, и моноклональных антител, подходящих для применения в способах настоящего изобретения, описывается подробнее в примере 2. Моноклональные антитела против человеческого gp39 по настоящему изобретению являются предпочтительными для использования при индуцировании толерантности антигенспецифических T-клеток. Предпочтительными антителами являются моноклональные антитела 3E4, 2H5, 2H8, 4D9-8, 4D9-9, 24-31, 24-43, 89-76 и 89-79, описанные в примере 2. Особенно предпочтительными антителами являются моноклональные антитела 89-76 и 24-31. Гибридомы 89-76 и 24-31, продуцирующие антитела 89-76 и 24-31, соответственно, депонированы по условиям Будапештсткого договора, в Американской коллекции типовых культур, Parklawn Drive, Роквилл, Мэриленд, 2 сентября 1994. Гибридоме 89-76 присвоен инвентарный номер ATCC HB117-13, и гибридоме 24-31 присвоен инвентарный номер ATCC HB11712. Антитела 24-31 и 89-76 являются изотипом IgG1. В другом варианте осуществления изобретения для использования в способах настоящего изобретения mAb против человеческого gp39 связывает эпитоп, распознаваемый моноклональным антителом, выбранным из группы, состоящей из 3E4, 2H5, 2H8, 4D9-8, 4D9-9, 24-31, 89-76 и 89-79. Предпочтительнее mAb против человеческого gp39 связывает эпитоп, распознаваемый моноклональным антителом 24-31 или моноклональным антителом 89-76. Способность mAb связывать эпитоп, указанный любым из вышеупомянутых антител, может быть определена стандартными проверками перекрестной конкуренции. Например, антитело, которое связывает те же эпитопы, которые узнаны mAb 24-31, будет конкурировать со связыванием меченного 24-31 с активированными Т-клетками, в то время как антитело, которое связывает другой, не узнанный mAb 24-31 эпитоп, не будет конкурировать при связывании меченного 24-31 с активированными Т-клетками. Б. Растворимые лиганды для gp39 Другие антагонисты gp39, которые можно вводить для индуцирования толерантности T-клеток, включают растворимые формы лиганда gp39. Одновалентный растворимый лиганд gp39, такой как растворимый CD40, может связываться с gp39, ингибируя, таким образом, взаимодействие gp39 с CD40 на B-клетках. Термин “растворимый” указывает, что лиганд не ассоциируется постоянно с клеточной мембраной. Растворимый лиганд gp39 можт быть получен химическим синтезом или предпочтительно методом рекомбинантных ДНК, например, путем экспрессии только внеклеточного домена лиганда (при отсутствии трансмембранных и цитоплазматических доменов). Предпочтительным растворимым лигандом gp39 является растворимый CD40. Альтернативно, растворимый лиганд gp39 может находиться в форме слитого белка. Такой слитый белок содержит по крайней мере часть лиганда gp39, присоединенную ко второй молекуле. Например, CD40 может быть экспрессирован в виде слитого белка с иммуноглобулином (т.е. слитый белок CD40Ig). В одном из вариантов осуществления изобретения продуцируется слитый белок, содержащий аминокислотные остатки внеклеточного домена части CD40, связанные с аминокислотными остатками последовательности, соответствующей шарнирам CH2 и CH3 тяжелой цепи иммуноглобулина, т.е. Cyl, с образованием слитого белка CD40Ig (см., например, LinsLey et al., (1991) J. Exp. Med. 1783: 721-730; Capon et al., (1989) Nature 337: 525-531; и Capon, патент США 5 116 964). Слитый белок может быть получен путем химического синтеза или предпочтительно методом рекомбинантных ДНК на основе кДНК CD40 (Stamenkovic et al., EMBO J., 8: 1403-1410 (1989)). II. Клетки для индукции антиген-специфической толерантности Настоящее изобретение основано, по крайней мере частично, на открытии, что представление аллоантигенов T-клеткам аллогенными клетками в присутствии антагониста gp39 приводит в результате к T-клеточной толерантности к аллоантигенам. Клетки, которые способны индуцировать толерантность по такому механизму, включают клетки, которые представляют антиген и активируют T-клетки путем взаимодействия с gp39 (т.е. для доставки соответствующих сигналов к T-клетке для активации T-клеток необходимо взаимодействие между gp39 на T-клетке и лигандом gp39 на клетке, представляющей антиген). Ингибирование взаимодействия лиганда на аллогенной или ксеногенной клетке с gp39 на реципиентных T-клетках предотвращает T-клеточную активацию алло- или ксеноантигенами и, скорее, индуцирует T-клеточную толерантность к антигенам. Препятствование активации T-клетки через gp39 может предотвратить индукцию костимуляторных молекул на аллогенной или ксеногенной клетке (например, молекул семейства B7 на B-клетке), и таким образом клетка доставляет к T-клетке только антигенный сигнал в отсутствие костимуляторного сигнала, индуцируя, таким образом, толерантность. Соответственно, по способам настоящего изобретения реципиенту вводят аллогенные или ксеногенные клетки. Аллогенная или ксеногенная клетка способна представить антиген Т-клеткам реципиента и представляет собой, например, B-лимфоцит – клетку, “профессионально” представляющую антиген (например, моноцит, дендритную клетку, клетку Лагерганса), или другую клетку, которая представляет антиген имунным клеткам (например, кератиноцит, эндотелиальную клетку, астроцит, фибробласт, олигодендроцит). Более того, предпочтительно, чтобы аллогенная или ксеногенная клетка имела пониженную способность стимулировать костимуляторный сигнал в реципиентных T-клетках. Например, аллогенная или ксеногенная клетка может утратить экспрессию или экспрессировать только на низком уровне, костимуляторных молекул, таких как белков семейства B7 (например, B7-1 и B7-2). Экспрессия костимуляторных молекул на потенциальных аллогенных или ксеногенных клетках, которые используются в способах настоящего изобретения, может быть оценена стандартными методами, например проточной цитометрией, с использованием антител, направленных против костимуляторных молекул. Предпочтительными аллогенными или ксеногенными клетками для индуцирования T-клеточной толерантности являются лимфоидные клетки, например лимфоциты периферической крови или селезеночные клетки. Предпочтительными лимфоидными клетками для индуцирования T-клеточной толерантности являются B-клетки. B-клетки могут быть выделены в чистом виде из смешанной популяции клеток (например, клеток других типов в периферической крови или селезенке) стандартными способами разделения клеток. Например, прилипающие клетки можно удалить путем культивирования селезеночных клеток в пластиковых чашках и извлечения популяции неприлипающих клеток. T-клетки можно удалить из смешанной популяции клеток путем обработки антителом против T-клеток (например, анти-Thy1.1 и/или анти-Thy1.2) и комплементом. В одном из вариантов осуществления изобретения в качестве антиген-представляющих клеток используются покоящиеся лимфоидные клетки, предпочтительно покоящиеся B-клетки. Покоящиеся лимфоидные клетки, такие как покоящиеся B-клетки, можно выделить методами, известными в технике, например, основываясь на их малом размере и плотности. Покоящиеся лимфоидные клетки можно выделить, например, проточным элютриационным центрифугированием, как описано в примере 1. При использовании проточного элютриационного центрифугирования популяция малых, покоящихся лимфоидных клеток, очищенная от клеток, которые могут активировать T-клеточные реакции, может быть получена путем собирания фракции (фракций) при 14-19 мл/мин, предпочтительно при 19 мл/мин (при 3200 об/мин). С другой стороны, малые покоящиеся лимфоциты (например, B-клетки) могут быть выделены путем центрифугирования в прерывистом градиенте плотности, например при использовании градиента фиколла или перколла, и после центрифугирования может быть получен слой, содержащий малые покоящиеся лимфоциты. Малые покоящиеся B-клетки также можно отличить от активированных B-клеток путем проверки стандартными методами (например, иммунофлюоресценцией), экспрессии костимуляторных молекул, таких как B7-1 и/или В7-2, на поверхности активированных B-клеток. Аллогенные или ксеногенные клетки, введенные реципиенту, функционируют, по крайней мере частично, как представляющие донорские антигены реципиентным Т-клеткам. Таким образом, клетки экcпрессируют антигены, которые также экспрессируются донорской тканью или органом. Обычно это может осуществляться посредством использования аллогенных или ксеногенных клеток, полученных от донора тканевого или органного трансплантата. Например, от донора ткани или органа могут быть выделены периферические лимфоидные клетки, B-клетки или селезеночные клетки, и использованы в способах настоящего изобретения. Альтернативно, аллогенные или ксеногенные клетки могут быть получены из источника иного, чем донор ткани или органа, если такие клетки имеют антигенные детерминанты, общие с тканью или органом донора. Например, могут использоваться аллогенные или ксеногенные клетки, которые экспрессируют (по большей части или все) такие же антигены главного комплекса гистосовместимости, что и донорская ткань или орган. Таким образом, могут использоваться аллогенные или ксенгенные клетки из источника, который является гаплотипом ГКГС, совместимым с донором ткани или органа (Например, ближайший родственник донора трансплантата). III. Введение клеток и антагонистов gp39 T-клеточная толерантность к органному или тканевому трансплантату может быть индуцирована путем введения реципиенту трансплантата антагониста gp39 в сочетании с аллогенной или ксеногенной клеткой, которые экспрессируют донорские антигены и взаимодействуют с реципиентными T-клетками через gp39. В предпочтительном варианте осуществления изобретения аллогенную или ксеногенную клетку и антагонист gp39 вводят реципиенту совместно или одновременно. С другой стороны, антагонист gp39 можно вводить до введения аллогенных или ксеногенных клеток, например, когда антагонист представляет собой антитело с длительным временем полужизни. В предпочтительном варианте осуществления изобретения антагонист и аллогенные или ксеногенные клетки вводят реципиенту перед трансплантацией реципиенту органа или ткани (т.е. реципиент предварительно получает антагонист и клетки). Например, введение аллогенных и ксеногенных клеток и антагониста может быть осуществлено за несколько дней (например, за пять-восемь дней) перед пересадкой ткани или органа. Обнаружено, что введение однократной дозы аллогенных клеток (в сочетании с антагонистом) является достаточным для индукции T-клеточной толерантности к донорской ткани или органу (см. пример 1). Число вводимых клеток может изменяться в зависимости от типа используемых клеток, типа тканевого или органного трансплантата, массы реципиента, общего состояния реципиента и других переменных, известных специалисту. Подходящее число клеток для использования в способе настоящего изобретения может быть определено специалистом в этой области техники обычными способами (например, как описано в примере 1). Клетки вводят в форме и путем, которые подходят для индукции T-клеточной толерантности у реципиента. Клетки могут быть введены в физиологически приемлемом растворе, таком как забуференный фосфатом физиологический раствор, или в подобном носителе. Предпочтительно клетки вводят внутривенно. Антагонист по изобретению вводят субъекту в биологически совместимой форме, пригодной для фармацевтического введения in vivo для индуцирования T-клеточной толерантности. “Биологически совместимая форма, пригодная для введения in vivo” означает форму антагониста, в которой любые токсические эффекты определяются лечебным действием соединения. Термин “субъект” предполагается для обозначения живых организмов, в которых может быть вызвана иммунная реакция, например, млекопитающих. Примерами субъектов являются люди, собаки, кошки, мыши, крысы и их трансгенные виды. Антагонист gp39 может вводиться в любой фармакологической форме, необязательно, с фармацевтически приемлемым носителем. Введение терапевтически активного количества антагониста определяется как введение количества, эффективного при дозах и в течение времени, необходимых для достижения нужного результата (например, T-клеточной толерантности). Например, терапевтически активное количество антагониста gp39 может изменяться в соответствии с такими факторами, как стадия заболевания, возраст, пол и масса особи и способность антагониста вызывать нужную реакцию у особи. Схемы дозировки могут быть скорректированы для обеспечения оптимального терапевтического ответа. Например, могут вводиться несколько небольших доз ежедневно, или доза может быть пропорционально снижена, в соответствии с потребностями терапевтической ситуации. Как описано в примере 1, для лечения антителом против gp39 схема эффективного лечения может включать установление введения антитела перед трансплантацией ткани или органа (например, за пять-восемь дней до трансплантации) с последующим повторяющимся введением антитела (например, каждый второй день) в течение нескольких недель (например, двух-семи недель) после пересадки. Активное соединение (например, антагонист, такой как антитело) может вводиться обычным способом, таким как инъекция (подкожная, внутривенная и т. п.), пероральное введение, ингаляция, трансдермальная аппликация или ректальное введение. В зависимости от способа введения активное соединение может быть покрыто материалом для защиты соединения от действия ферментов, кислот и других естественных условий, которые могут инактивировать соединение. Предпочтительным способом введения является внутривенная инъекция. Чтобы ввести антагонист gp39 способом, иным, чем парентеральное введение, может понадобиться нанести на антагонист покрытие из материала или ввести этот материал вместе с антагонистом, который предупредит его инактивацию. Например, антагонист может вводиться особи в соответствующем носителе или разбавителе, вводиться вместе с ферментными ингибиторами или в соответствующем носителе, таком как липосомы. Фармацевтически приемлемыми разбавителями являются физиологический раствор и водные буферные растворы. Ферментными ингибиторами являются ингибитор панкреатического трипсина, диизопропилфосфат (DEP) и тразилол. Липосомы включают эмульсии “вода в масле в воде”, а также обычные липосомы (Strejan et al. (1984) J. Neuroimmunol. 7:27). Активное соединение также может быть введено парентерально или интраперитонеально. Также могут быть получены дисперсии в глицерине, жидких полиэтиленгликолях, в их смесях и в маслах. При обычных условиях хранения и применения эти препараты могут содержать консервант для предупреждения развития микроорганизмов. Фармацевтические композиции, пригодные для применения в инъекциях, включают стерильные водные растворы (когда все растворено в воде) или дисперсии и стерильные порошки для приготовления стерильных растворов для инъекций или дисперсии для немедленного применения. Во всех случаях композиция должна быть стерильной и должна быть жидкой в такой степени, чтобы можно было легко использовать шприц. Она должна быть устойчивой в условиях изготовления и хранения и должна быть предохранена от загрязнения микроорганизмами, такими как бактерии или грибы. Носитель должен представлять собой растворитель или дисперсионную среду, содержащую, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль, и т.п.), и их подходящие смеси. Подходящая текучесть может поддерживаться, например, с помощью покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии, и с применением поверхностно-активных веществ. Предупреждение действия микроорганизмов может быть достигнуто с помощью различных антибактериальных и противогрибковых средств, например парабенов, хлорбутанола, фенола, аскорбиновой кислоты, тимеросала и подобных средств. Во многих случаях предпочтительно включать в композицию изотонические агенты, например сахара, многоатомные спирты, такие как маннит, сорбит, хлорид натрия. Пролонгированное поглощение инъецируемых композиций может вызываться включением в композиции средства, которое замедляет абсорбцию, например, моностеарата алюминия или желатина. Стерильные инъецируемые композиции можно получить путем включения активного соединения (например, антагониста gp39) в требуемом количестве в соответствующий растворитель, в сочетании с одним или несколькими ингредиентами, перечисленными выше, в соответствии с требованиями, с последующей стерилизацией фильтрацией. Вообще, дисперсии готовят путем введения активного соединения в стерильный носитель, который содержит основную дисперсионную среду и требуемые другие ингредиенты из числа перечисленных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций предпочтительными способами получения являются вакуумная сушка и сушка вымораживанием, которые дают порошок активного ингредиента (например, антагониста) плюс любой дополнительный желательный ингредиент из их раствора, предварительно стерилизованного фильтрацией. Когда активное соединение соответствующим образом защищено, как описано выше, белок можно вводить орально, например, с инертным разбавителем или усвояемым съедобным носителем. Используемый здесь термин “фармацевтически приемлемый носитель” включает любые растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические агенты и замедлители и т.п. Применение таких сред и агентов с фармацевтически активными веществами хорошо известны в технике. Применение любых обычных сред или агентов в лекарственных композициях рассматривается во всех случаях, за исключением их несовместимости с активным соединением. В композиции также могут включаться дополнительные активные соединения. Особенно выгодно составлять парентеральные композиции в виде единиц лекарственной формы для облегчения введения и равномерности дозировки. Под единицей лекарственной формы здесь подразумевается физически разделенные единицы, пригодные для лечения особей млекопитающих в качестве стандартных доз; причем каждая единица содержит заранее установленное количество активного соединения, рассчитанное на получение нужного лечебного действия, в сочетании с требуемым фармацевтическим носителем. Определение единиц лекарственной формы по изобретению диктуются и напрямую зависят от (а) особых свойств активного соединения и конкретного терапевтического эффекта, которого нужно достичь, и (б) ограничений, присущих технике приготовления составов такого активного соединения для лечения чувствительных особей. После или вместе с описанной здесь схемой вызывания толерантности, донорскую ткань или орган пересаживают реципиенту трансплантата обычными способами. IV. Применение способов изобретения Способы настоящего изобретения могут использоваться в самых разных ситуациях при пересадке ткани и органов. Упомянутые способы могут применяться для индуцирования T-клеточной толерантности у реципиента трансплантата ткани или органа, такого как печень, почка, сердце, легкое, кожа, мышца, нервная ткань, желудок и кишки. Таким образом, способы настоящего изобретения могут применяться при лечении заболеваний иди состояний, которые влекут за собой пересадку ткани или органа (например, пересадка печени при лечении гиперхолестеринемии, пересадка мышечных клеток при лечении мышечной дистрофии, пересадка нервной ткани при лечении болезни Юнтингтона или болезни Паркинсона, и т.п.). В предпочтительном варианте осуществления изобретения трансплантированная ткань содержит панкреатические островки. Соответственно, изобретение касается способа лечения диабета посредством трансплантации клеток панкреатических островков. Упомянутый способ включает введение субъекту, нуждающемуся в таком лечении, 1) аллогенных или ксеногенных клеток, которые экспрессируют донорские антигены, 2) антагониста молекулы, экспрессированной на T-клетках реципиента, которая опосредует контакт-зависимую хелпер-эффекторную функцию, такого как антагонист gp39 (например, антитела против gp39), и 3) донорских клеток панкреатических островков. Предпочтительно, аллогенные или ксеногенные клетки и антагонист вводят реципиенту перед введением ему панкреатических островков. Настоящее изобретение также иллюстрируется следующими далее примерами, которые не следует рассматривать, как ограничивающие. Все ссылки, патенты и опубликованные заявки на патенты, цитируемые в настоящей заявке, включаются в нее в качестве ссылок. Пример 1. Индукция толерантности к аллотрансплантатам панкреатических островков посредством введения реципиенту аллогенных клеток и анти-39 В настоящее время установлено, что аллотрансплантация зависит от общей иммуносупресии, которая неспецифически отсекает иммуноэффекторные функции. Однако иммуносупрессивные фармацевтические препараты могут вызвать значительные побочные эффекты. Кроме того, такой подход отражает аллотрансплантация островков Лангерханса для лечения диабета (см., например, Robertson R. P, (1992) N. Engl. J. Med. 327, 1861). Терапия с антителами, направленными против T-клеток, может дать возможность для успешной аллотрансплантации островков грызунам, но такой подход дает слишком одинаковые результаты при общей иммуносупрессии (Carpenter C.B. (1990) N. Engl. Med. 322, 1224; Roark J. H. et al. (1992) Transplantation 54, 1098; Kahan B.D. (1992) Curr. Opin. Immunol. 4, 553). В этом примере толерантность к островковым трансплантатам индуцируется у реципиента трансплантата посредством манипулирования презентацией аллоантигена T-клеткам с целью предотвращения их активации. Срок клеток жизнеспособности аллотрансплантатов у мышей C57BL/6 (H-2b) с “химическим” диабетом проверяют, используя методологию, описанную ниже. Индукция диабета Самцов мышей C57B1/6J (H-2b) приводят в состояние диабета посредством внутривенного введения стрептозотоцина (140 мг/кг). Постоянный диабет подтверждается демонстрацией концентрации глюкозы в плазме, 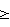 400 мг/мл, в трех случаях в течение одной недели.
Фракционирование аллогенных селезеночных клеток 400 мг/мл, в трех случаях в течение одной недели.
Фракционирование аллогенных селезеночных клетокДонорские аллогенные клетки для предварительного введения реципиентам трансплантатов получают от гибридных животных (C57хBALB/c) (H-2b/d)F1 для предотвращения реакции “трансплантат против хозяина”. Чтобы выделить малые лимфоцитные клетки, суспензии селезеночных клеток от 8-недельных самок мышей (C57хBALB/c)F1 очищают от эритроцитов и затем фракционируют по размеру посредством элютрации, как описано в Tony H.P. et al. (1985) J. Exp. Med. 161, 223; и в Gosselin E.J et al. (1988) J. Immunol. 140, 1408. Вкратце, малые лимфоциты выделяют посредством проточного эдютриацинного центрифугирования, используя, например, центрифугу модели J-6В (Beckman Instruments, Palo Alto CA). Приблизительно 1-5  108 клеток в 8 мл культурной среды или в уравновешенном солевом растворе с 1,5% фетальной бычьей сыворотки обрабатывают дезоксирибонуклеазой, помещают в элютрационную камеру при исходной скорости противотока 13,5 мл/мин, и вращают при 4oC с постоянной скоростью 3200 об/мин. Фракция малых клеток с очень незначительной примесью больших клеток элюируется, как правило, при 14-19 мл/мин хотя точная скорость потока может зависеть от среды, в которой суспендируются клетки. В экспериментах, описанных здесь, фракцию малых клеток собирают при 19 мл/мин (при 3200 об/мин). Эта фракция полностью очищена, по данным о вспомогательной клеточной функции устойчивости к излучению (3000 рад), при анализе с T-клеточными линиями, специфическими либо для кроличьего IgG и H2d (CDC35), либо для аллореактивных к H2b(D10.G4). Малые клетки и нефракционированные клетки дважды промывают в безсывороточной среде перед инъекцией в хвостовую вену реципиентам аллотрансплантатов. Получают приблизительно 40-100 106 нефракционированных селезеночных клеток (C57хBALB/c)F1(H-2b/d) или 40-100 106 элютриированных малых селезеночных клеток (C57хBALB/c)F1(H-2b/d).
Предварительная обработка реципиентов трансплантатов 108 клеток в 8 мл культурной среды или в уравновешенном солевом растворе с 1,5% фетальной бычьей сыворотки обрабатывают дезоксирибонуклеазой, помещают в элютрационную камеру при исходной скорости противотока 13,5 мл/мин, и вращают при 4oC с постоянной скоростью 3200 об/мин. Фракция малых клеток с очень незначительной примесью больших клеток элюируется, как правило, при 14-19 мл/мин хотя точная скорость потока может зависеть от среды, в которой суспендируются клетки. В экспериментах, описанных здесь, фракцию малых клеток собирают при 19 мл/мин (при 3200 об/мин). Эта фракция полностью очищена, по данным о вспомогательной клеточной функции устойчивости к излучению (3000 рад), при анализе с T-клеточными линиями, специфическими либо для кроличьего IgG и H2d (CDC35), либо для аллореактивных к H2b(D10.G4). Малые клетки и нефракционированные клетки дважды промывают в безсывороточной среде перед инъекцией в хвостовую вену реципиентам аллотрансплантатов. Получают приблизительно 40-100 106 нефракционированных селезеночных клеток (C57хBALB/c)F1(H-2b/d) или 40-100 106 элютриированных малых селезеночных клеток (C57хBALB/c)F1(H-2b/d).
Предварительная обработка реципиентов трансплантатовРеципиентам трансплантатов предварительно вводят либо нефракционированные аллогенные селезеночные клетки (C57хBALB/c)F1(H-2b/d), либо элютриированную “фракцию 19” селезеночных клеток малого диаметра, которые очищены от активности АПК (выделены так, как описано выше), либо моноклональное антитело против gp39 (MR1, см. пример 2, опыт 3); либо комбинацию аллогенных клеток и антитела против gp39. Клетки фракции 19 проверяют при двух различных интервалах доз – при малой дозировке в 40-44 106 клеток или при высокой дозе – 77-88 106 клеток. Контрольным животным не вводят ни аллогенные клетки, ни антитело. Аллогенные клетки вводят реципиентам трансплантата посредством инъекции в хвостовую вену за пять-восемь суток до трансплантации островкового трансплантата. Антитело MR1 вводят при дозе 250 мкг/мышь дважды в неделю, начиная за 7 дней до трансплантации островков и продолжая в течение 2-7 недель или до неблагоприятного исхода трансплантации. Первую инъекцию антитела обычно дают в тот же день, что и первую инъекцию аллогенных селезеночных клеток.
Трансплантация островкового трансплантатаАллогенные островки BALB/c(H-2d) выделяют модифицированным способом коллагеназного переваривания (Gottlieb P.A., et al. (1990) Diabets 39, 643). Островки при дозе 30 островков на г массы тела имплантируют в субренальную капсулу реципиентной мыши (C57B1/6J(Н-2b) сразу же после выделения. Срок жизнеспособности трансплантата определяют по сохранению концентрации глюкозы в плазме на уровне  200 мг/мл.
Результаты 200 мг/мл.
РезультатыВ первой серии экспериментов реципиентам островковых трансплантатов предварительно вводили либо одни аллогенные селезеночные клетки, либо одно антитело против gp39. Как видно на фиг. 1, при отсутствии предварительно введенных селезеночных клеток, все островковые трансплантаты отторгаются в пределах 13 суток после пересадки (9  2 сут; интервал 5-13 сут; N=23). Недостаточный срок жизнеспособности островков также наблюдается у животных, которым вводили только нефракционированные селезеночные клетки, содержащие нормальную активность АПК (63 сут; интервал 4-12 сут; N=7), или низкие дозы (40-44 106 клеток) фракции 19, очищенных от АПК, селезеночных клеток (73 сут; интервал 3-14 сут, N=16). Напротив, инъекция более высокой дозы фракции 19, очищенных от АПК, малых спленоцитов (75-88 106 клеток) продлевает срок жизнеспособности аллотрансплантатов (1910 сут; интервал 7-40 сут; N=16). Этот эффект продолжительности срока жизнеспособности трансплантата статистически довольно значителен (F3,58=17,3, p < 0,001, при сравнении с группами, не получавшими ничего, получавшими нефракционированные селезеночные клетки или меньшую дозу селезеночных клеток фракции 19), но не был постоянным. Протяженный, но ограниченный срок жизнеспособности аллогенных островков у реципиентов с диабетом, получивших малые клетки фракции 19, очищенной от АПК, приводит к мысли, что одни эти клетки не могут поддерживать срок жизнеспособности трансплантата. Дополнительным группам реципиентов трансплантатов ввели 77-88 106 клеток фракции 20. Эту фракцию также, в подавляющем большинстве, составляют малые лимфоциты, но их популяция отличается от популяции фракции 19 тем, что содержит доступную измерению функцию АПК. Реципиенты этих клеток (N= 6) быстро отторгают свои трансплантаты (среднее = 8,5 сут; интервал 6-12). Другой группе реципиентов трансплантатов вводили только моноклональное антитело против gp39 – MR1. Фиг. 1 показывает, что островковые аллотрансплантаты выходят из строя в пределах 15 дней у 7/11 мышей, получивших только анти-gp39 mAb. Остальные четыре мыши имели фукционирующие трансплантаты при завершении эксперимента на 48 сутки. Результаты показывают, что введение реципиенту одного антитела против gp39 MR1 может продлить срок жизнеспособности островкового трансплантата (среднее = 20-19 сут; интервал 9-неопредел., N=5). Степень пролонгации статистически подобна степени, достигнутой с применением более высокой дозы одних селезеночных клеток фракции 19, и существенно выше степени, достигнутой в трех других группах (p < 0,05).
Ряд экспериментов, описанных выше, показывает, что одно введение высоких доз фракции 19 селезеночных клеток, очищенных от АПК, или анти-gp39 mAb может увеличить срок жизнеспособности трансплантатов панкреатических островков при сравнении с отсутствием такого введения. Однако, любая обработка только одним препаратом не является эффективной при индуцировании у реципиента длительной толерантности к островковым трансплантатам. Обнаружено, что комбинированное введение аллогенных селезеночных клеток и анти-gp39 является более эффективным, чем введение одного каждого реагента. Результаты приводятся на фиг. 2, при этом каждая кривая представляет данные для отдельной мыши. Незаштрихованные значки относятся к реципиентам, у которых трансплантаты разрушаются самопроизвольно. Заштрихованные значки относятся к мышам, у которых островковые трансплантаты функционировали при окончании эксперимента. Фиг. 2 показывает, что неопределенный срок жизнеспособности достигается у всех животных, получавших в течение 7 недель анти-gp39 mAb и однократную инъекцию фракции 19 – селезеночных клеток, очищенных от АПК (N= 6). Изменение этой схемы путем снижения длительности введения анти-gp39 ослабляет, но не отменяет, благоприятное действие на срок жизнеспособности трансплантата. Неопределенный срок жизнеспособности трансплантата достигается у 6/8 реципиентов, когда анти-gp39 mAb вводят только в течение 2 недель в сочетании с фракцией 19 селезеночных клеток (фиг. 2А). Неопределенный срок жизнеспособности трансплантатов также наблюдается у реципиентов, получавших анти-gp39 в течение 2-7 недель в сочетании с одной инъекцией нефракционированных аллогенных селезеночных клеток.
Для подтверждения функции трансплантата и отсутствия инсулиновой секреции оставшимися нативными островками, неразрушенными при введении стрептозотоцина, удаляют почки, несущие субренальные имплантаты. Во всех случаях односторонняя нефрэктомия дает в результате рецидив гипергликемии (>300 мг/мл) в пределах 3 суток.
Островковые аллотрансплантаты и природная поджелудочная железа исследовались гистологически у всех животных либо когда трансплантат разрушался, либо по окончании эксперимента. Гистологические срезы островковых аллотрансплантатов в почках реципиентов фракционированных аллогенных малых лимфоцитов и продолжительного (7 недель) введения mAb MR1 показывает обильные интактные островки, видимые ниже ренальной капсулы, которые лишены мононуклеарной инфильтрации и содержат хорошо гранулированные инсулин- и глюкагон-положительные клетки. Напротив, гистологические срезы островковых аллотрансплантатов в почках реципиентов, получавших одно анти-gp39 mAb, показывают характерное интенсивное воспаление с мононуклеарными клетками и разрушение сопутствующих островковых клеток. Во всех поджелудочных железах хозяев морфология островков едина со стрептозотоциновым диабетом.
Пример 2. Продуцирование и исследование антител против gp39 2 сут; интервал 5-13 сут; N=23). Недостаточный срок жизнеспособности островков также наблюдается у животных, которым вводили только нефракционированные селезеночные клетки, содержащие нормальную активность АПК (63 сут; интервал 4-12 сут; N=7), или низкие дозы (40-44 106 клеток) фракции 19, очищенных от АПК, селезеночных клеток (73 сут; интервал 3-14 сут, N=16). Напротив, инъекция более высокой дозы фракции 19, очищенных от АПК, малых спленоцитов (75-88 106 клеток) продлевает срок жизнеспособности аллотрансплантатов (1910 сут; интервал 7-40 сут; N=16). Этот эффект продолжительности срока жизнеспособности трансплантата статистически довольно значителен (F3,58=17,3, p < 0,001, при сравнении с группами, не получавшими ничего, получавшими нефракционированные селезеночные клетки или меньшую дозу селезеночных клеток фракции 19), но не был постоянным. Протяженный, но ограниченный срок жизнеспособности аллогенных островков у реципиентов с диабетом, получивших малые клетки фракции 19, очищенной от АПК, приводит к мысли, что одни эти клетки не могут поддерживать срок жизнеспособности трансплантата. Дополнительным группам реципиентов трансплантатов ввели 77-88 106 клеток фракции 20. Эту фракцию также, в подавляющем большинстве, составляют малые лимфоциты, но их популяция отличается от популяции фракции 19 тем, что содержит доступную измерению функцию АПК. Реципиенты этих клеток (N= 6) быстро отторгают свои трансплантаты (среднее = 8,5 сут; интервал 6-12). Другой группе реципиентов трансплантатов вводили только моноклональное антитело против gp39 – MR1. Фиг. 1 показывает, что островковые аллотрансплантаты выходят из строя в пределах 15 дней у 7/11 мышей, получивших только анти-gp39 mAb. Остальные четыре мыши имели фукционирующие трансплантаты при завершении эксперимента на 48 сутки. Результаты показывают, что введение реципиенту одного антитела против gp39 MR1 может продлить срок жизнеспособности островкового трансплантата (среднее = 20-19 сут; интервал 9-неопредел., N=5). Степень пролонгации статистически подобна степени, достигнутой с применением более высокой дозы одних селезеночных клеток фракции 19, и существенно выше степени, достигнутой в трех других группах (p < 0,05).
Ряд экспериментов, описанных выше, показывает, что одно введение высоких доз фракции 19 селезеночных клеток, очищенных от АПК, или анти-gp39 mAb может увеличить срок жизнеспособности трансплантатов панкреатических островков при сравнении с отсутствием такого введения. Однако, любая обработка только одним препаратом не является эффективной при индуцировании у реципиента длительной толерантности к островковым трансплантатам. Обнаружено, что комбинированное введение аллогенных селезеночных клеток и анти-gp39 является более эффективным, чем введение одного каждого реагента. Результаты приводятся на фиг. 2, при этом каждая кривая представляет данные для отдельной мыши. Незаштрихованные значки относятся к реципиентам, у которых трансплантаты разрушаются самопроизвольно. Заштрихованные значки относятся к мышам, у которых островковые трансплантаты функционировали при окончании эксперимента. Фиг. 2 показывает, что неопределенный срок жизнеспособности достигается у всех животных, получавших в течение 7 недель анти-gp39 mAb и однократную инъекцию фракции 19 – селезеночных клеток, очищенных от АПК (N= 6). Изменение этой схемы путем снижения длительности введения анти-gp39 ослабляет, но не отменяет, благоприятное действие на срок жизнеспособности трансплантата. Неопределенный срок жизнеспособности трансплантата достигается у 6/8 реципиентов, когда анти-gp39 mAb вводят только в течение 2 недель в сочетании с фракцией 19 селезеночных клеток (фиг. 2А). Неопределенный срок жизнеспособности трансплантатов также наблюдается у реципиентов, получавших анти-gp39 в течение 2-7 недель в сочетании с одной инъекцией нефракционированных аллогенных селезеночных клеток.
Для подтверждения функции трансплантата и отсутствия инсулиновой секреции оставшимися нативными островками, неразрушенными при введении стрептозотоцина, удаляют почки, несущие субренальные имплантаты. Во всех случаях односторонняя нефрэктомия дает в результате рецидив гипергликемии (>300 мг/мл) в пределах 3 суток.
Островковые аллотрансплантаты и природная поджелудочная железа исследовались гистологически у всех животных либо когда трансплантат разрушался, либо по окончании эксперимента. Гистологические срезы островковых аллотрансплантатов в почках реципиентов фракционированных аллогенных малых лимфоцитов и продолжительного (7 недель) введения mAb MR1 показывает обильные интактные островки, видимые ниже ренальной капсулы, которые лишены мононуклеарной инфильтрации и содержат хорошо гранулированные инсулин- и глюкагон-положительные клетки. Напротив, гистологические срезы островковых аллотрансплантатов в почках реципиентов, получавших одно анти-gp39 mAb, показывают характерное интенсивное воспаление с мононуклеарными клетками и разрушение сопутствующих островковых клеток. Во всех поджелудочных железах хозяев морфология островков едина со стрептозотоциновым диабетом.
Пример 2. Продуцирование и исследование антител против gp39Опыт 1. Антитела, направленные против человеческого gp39 Для индукции антиген-специфической T-клеточной толерантности у человека предпочтительно вводить антитело, направленное против человеческого gp39. Для продуцирования мышиных моноклональных антител против человеческого gp39 используют следующую методологию. Мышей BALB/c иммунизируют растворимым слитым белком gp39 gp39-CD8 в полном адъюванте Фрейнда (CFA). Затем, спустя 6 недель, мышам вводят растворимый gp39-CD8 в неполном адъюванте Фрейнда (IFA). Растворимый gp39-CD8 дают в растворимой форме через 4 недели после второй иммунизации. Затем, 2 неделями позже, мышей ревакцинируют активированными человеческими лимфоцитами периферической крови, и еще через 2 недели их окончательно ревакцинируют растворимым gp39-CD8. Спленоциты сливались с партнерными клетками слияния NS-1 на 4 сутки после последней иммунизации по протоколам иммунизации. Клоны, продуцирующие антитела против человеческого gp39, выделяют на основе процесса многократного скрининга. Сначала клоны скринируют методом фиксации на бактериологических чашках, используя gp39-CD8. Затем положительные клоны скринируют против контрольного слитого белка CD8 – CD72-CD8. Клоны, которые отмечены как положительные при анализе фиксации на бактериологических чашках, удаляются. Остающиеся клоны затем скринируют с покоящимися и активированными в течение 6 часов человеческими лимфоцитами периферической крови (PBL) с помощью проточного цитометрического анализа. Положительными считаются гибридомы, окрашивающие активированные, а не покоящиеся PBL. Наконец, остающиеся клоны проверяют на их способность блокировать связывание CD40Ig с зафиксированным на бактериологических чашках gp39. Приблизительно 300 клонов скринируют сначала против gp39-CD8 и CD72-CD8 при анализе фиксации на бактериологических чашках. Обнаружено, что из этих клонов 30 находят фиксированный на чашках gp39, но не CD8. Эти клоны затем скринируют для обнаружения gp39 на активированных человеческих PBL. Приблизительно 15 клонов находят молекулу на активированных PBL, но не покоящихся клетках. Далее специфичность подтверждается путем определения способности клонов блокировать связывание CD40Ig при таком анализе. Такими клонами стали 3E4, 2H5 и 2H8. Такие клоны являются предпочтительными для применения в описанных здесь способах. Клоны, которые положительны на активированных, а не на покоящихся PBL, также скринируют на реакционноспособность с клоном активированных крысиных T-клеток POMC8. Клон 2H8 выражает перекрестную реактивность с этой линией крысиных T-клеток. Опыт 2. Антитела, направленные против человеческого gp39 Процедуру иммунизации, подобную процедуре, описанной в примере 1, используют для получения дополнительных антител, направленных против человеческого gp39. Одну мышь BALB/c иммунизируют растворимым gp39-CD8 в CFA, и впоследствии, 4 недели спустя, ей вводят активированные в течение 6 часов человеческие лимфоциты периферической крови. Впоследствии мышь ревакцинируют растворимым gp39-CD8 за 4 дня перед слиянием спленоцитов с клетками слияния NS-1 по стандартным протоколам. Скрининг гибридомных клонов осуществляют с помощью проточной цитометрии при окрашивании активированных в течение 6 часов PBL. Отбирают клоны, окрашивающие активированные, но не покоящиеся, человеческие PBL. Для дальнейшего анализа отбирают 6 клонов – 4D9-8, 4D9-9, 24-31, 24-43, 89-76 и 89-79. Специфичность выбранных антител подтверждается несколькими анализами. Сначала анализ методом проточной цитометрии показывает, что все 6 mAb окрашивают активированные, но не покоящиеся, T-клетки периферической крови (см., как характерный пример, фиг. 3Б и 3В, показывающие окрашивание активированных T-клеток 4D9-8 и 4D9-9 соответственно). Экспрессия молекулы, распознаваемой каждым из шести антител, обнаруживается в пределах 4 часов после активации, является максимальной через 6-8 часов после активации и не обнаруживается через 24 часа после активации. Все шесть mAb распознают молекулу, выраженную на активированных CD3+PBL, в основном, фенотипа CD4+, но часть CD8+T-клеток также экспрессируют молекулу. Экспрессия молекулы, распознаваемой этими шестью антителами, ингибируется присутствием в культуральной среде циклоспорина А, как и экспрессия gp39 (см., например, фиг. 4А и 4Б, показывающие окрашивание T-клеток, обработанных циклоспорином, 4D9-8 и 4D9-9 соответственно). Кинетика и распространение экспрессии молекулы, распознаваемой этими mAb, идентичны этим параметрам gp39, который находит слитый белок человеческого CD40Ig. Кроме того, все шесть моноклональных антител блокируют окрашивание gp39 CD40Ig (см., например, фиг. 5А и 5Б, на которых показывается ингибирование окрашивания gp39 CD40Ig в присутствии 4D9-8 и 4D9-9 соответственно). При анализе по методу ELISA все шесть mAb распознают gp39-CD8, растворимую слитую форму молекулы gp39. Кроме того, все шесть mAb образуют иммунопреципитат молекулы приблизительно в 36 кд из меченых 35S-метионином активированных человеческих PBL. Иммунопреципитированная молекула идентична молекуле, преципитированной слитым белком человеческого CD40Ig. Функциональную активность шести выбранных моноклональных антител (4D9-8, 4D9-9, 24-31, 24-43, 89-76 и 89-79) проверяют следующим образом. Сначала определяют способность mAb ингибировать пролиферацию очищенных человеческих B-клеток, выращенных с IL-4 и растворимым gp39. Очищенные человеческие T-клетки выращивают с gp39 и IL-4 в присутствии или в отсутствии очищенных моноклональных антител или CD40Ig при дозах от 0 до 12,5 мкг/мл. Пролиферацию B-клеток определяют через 3 дня после введения в культуру тимидина. Результаты (приводятся на фиг. 6) показывают, что все шесть mAb могут ингибировать пролиферацию B-клеток, индуцированную gp39 и IL-4. Наиболее эффективными при ингибировании индуцированной пролиферации B-клеток, являются mAb 89-76 и 24-31. Затем проверяют способность mAb ингибировать B-клеточную дифференцировку при измерении по продуцированию Ig, индуцированного анти-CD3 активированными Т-клетками и IL-2. Очищенные IgD+ человеческие B-клетки получают позитивной селекцией с клеточным сортером с возбуждением флуоресценции (FACS), и затем культивируют с анти-CD3 активированными человеческими Т-клетками (обработанными митомицином C) и IL-2 в течение 6 суток в присутствии или в отсутствии очищенных моноклональных антител против gp39 при дозировке от 0 до 10 мкг/мл. На 6 сутки методом ELISA проверяют продуцирование IgM, IgG и IgA. Результаты (даются ниже в табл. 1) показывают, что все шесть антител могут ингибировать T-клеточно-зависимую B-клеточную дифференцировку, при измерении по продуцированию IgM, IgG и IgA. Чтобы проверить влияние mAb против gp39 на T-клеточные реакции, моноклональные антитела включают в стандартные реакции смешанной культуры лимфоцитов (СКЛ). Культивируют 300000 человеческих лимфоцитов периферической крови (респондеры = P) со 100000 облученных аллогенных лимфоцитов периферической крови (стимуляторы = С) в присутствии или в отсутствии mAb против gp39 (10 мкг/мл). Культуры метят импульсно 3H-тимидином на 4, 5 и 6 сутки и через 18 часов собирают клетки, выросшие в культуре. Все шесть mAb против человеческого gp39 ингибируют аллоспецифические реакции при измерении по СКЛ (см., как характерный пример, фиг. 7, показывающий ингибирование аллоспецифических реакций, когда Р и С инкубируются в присутствии 24-31 или 89-76; для положительного контроля используют слитый белок CTLA4-иммуноглобулин и mAb против CD28). Чтобы определить, распознают ли все шесть моноклональных антител определенные эпитопы на молекуле человеческого gp39, осуществляют эксперименты с перекрестным блокированием. Сначала активированные человеческие PBL блокируют каждым из шести mAb (25 мкг/мл). Клетки промывают и затем окрашивают 10 мкг/мл биотинилированного антитела, после чего следует реакция с фитоэритринилированным авидином (PE-Av). Окрашивание клеток PE-Av анализируют FACS. Результаты приводятся ниже в табл. 2. Все антитела блокируют связывание CD40Ig с активированными человеческими PBL. Однако данные, приведенные в табл. 2, ясно показывают неэффективность некоторых антител при блокировке связывания других антител с активированными человеческими PBL, что приводит к предположению, что они распознают отдельные эпитопы на молекулах человеческого gp39. Гибридомы 89-76 и 24-31, продуцирующие антитела 89-76 и 24-31 соответственно, депонированы в соответствии с Будапештским договором в Американской коллекции типовых культур, Parklawn Drive, Роквилл, Мэриленд, 2 сентября 1994. Гибридоме 89-76 присвоен инвентарный номер ATCC HB11713 и гибридоме 24-31 присвоен инвентарный номер ATCC HB11712. Опыт 3. Антитела, направленные против мышиного gp39 В одном из вариантов осуществления настоящего изобретения антагонистом gp39 является моноклональное антитело против мышиного gp39 – MR1. Для продуцирования моноклонального антитела MR1 применяется описанный ниже способ, и его можно использовать для генерации других антител, направленных на gp39. Хомяков иммунизируют интраперитонально 5-106 активированных Th1-клеток (d1.6) с недельными интервалами в течение шести недель. Когда сывороточный титр против мышиных Th1 превышает 1:10000, осуществляют слияние клеток с полиэтиленгликолем, используя иммунные хомячьи спленоциты и NS-1. Супернатант от клеток, содержащих растущие гибридомы, скринируют проточной цитометрией на покоящихся и активированных Th1. Одну особую гибридому, которая продуцирует Mab, которое селективно распознает активированные Th, затем тестируют и субклонируют, чтобы получить MR1. MR1 получают в асците и очищают ионообменной ВЭЖХ. Гибридома MR1 депонирована Американской коллекцией типовых культур, и ей присвоен инвентарный номер HB11048. Эквиваленты Специалисты в этой области техники увидят или смогут осуществить, используя самые обычные экспериментальные методы, много эквивалентов конкретных вариантов осуществления настоящего изобретения, описанных здесь. Такие эквиваленты рассматриваются как входящие в объем приведенной ниже формулы изобретения. Все ссылки и опубликованные заявки на патенты, цитированные в настоящей заявке, включены в нее в качестве ссылок. Формула изобретения
РИСУНКИ
MM4A – Досрочное прекращение действия патента СССР или патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе
Дата прекращения действия патента: 26.04.2009
Извещение опубликовано: 20.07.2010 БИ: 20/2010
|
||||||||||||||||||||||||||