Патент на изобретение №2398874
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) КЛЕТОЧНЫЕ КЛОНЫ КАМБИЯ, СПОСОБ ИХ ВЫДЕЛЕНИЯ И КОНСЕРВАЦИИ
(57) Реферат:
Ткань растения, содержащую камбий, культивируют, индуцируя слой камбия и каллус из других тканей. Из полученной смеси выделяют и пролиферируют клон одной клетки камбия. Создание гомогенной клеточной линии из камбия позволяет минимизировать варьирование скорости роста клеток и образования вторичных метаболитов. Тем самым обеспечивается стабильность производства биологически активных веществ растительного происхождения в ходе долгосрочного культивирования. 4 н. и 11 з.п. ф-лы, 5 ил., 5 табл.
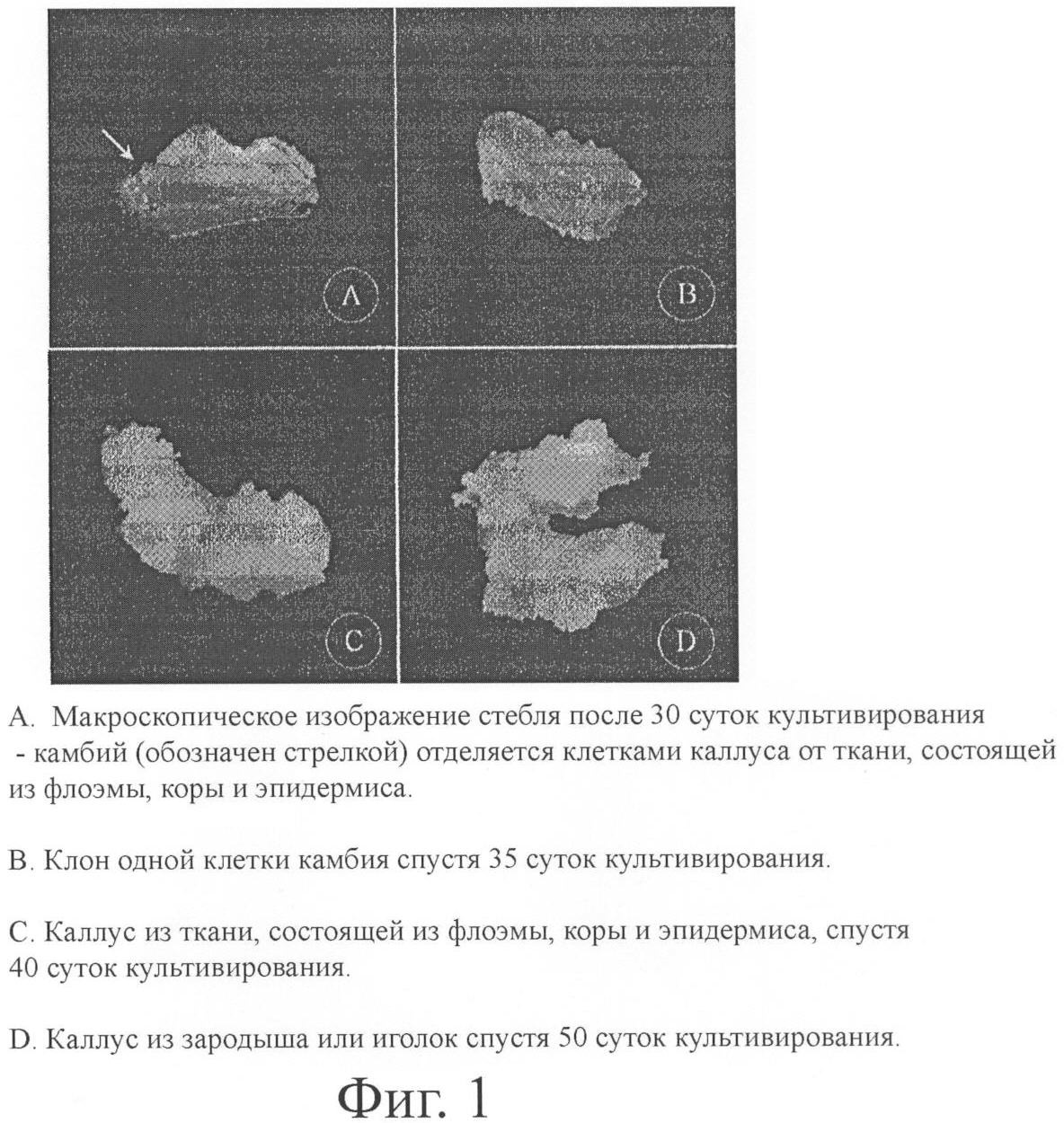
Область техники, к которой относится изобретение Растения используются не только как источник снабжения людей продовольствием, но и как источник экстенсивных химических веществ, включая лекарственные вещества, ароматические вещества, химические вещества для сельского хозяйства, красящие вещества и др. Биологически активные соединения, получаемые из растений, большей частью являются вторичными метаболитами. Отмечается повышенный интерес к вторичным метаболитам, таким как алкалоиды, аллергены, аминокислоты, антрахинон, противолейкемические агенты, противомикробные агенты, противоопухолевые агенты, противовирусные агенты, ферменты, флавоноиды, инсектициды, опиаты, ароматические соединения, пигменты, витамины, полисахариды и др., поскольку большинство из них работают как физиологические активные вещества. По данным Zhong (2002), на сегодняшний день известно около 100000 растительных вторичных метаболитов и свыше 25% применяемых в лечебной практике лекарственных средств представляют собой вещества растительного происхождения. Ежегодно открываются все новые и новые вторичные метаболиты. В способе получения этих метаболитов существует много проблем, таких как трудности с проведением химического синтеза, несмотря на удивительно быстрое развитие в последние годы органической химии; уничтожение природы ее чрезмерной эксплуатацией и загрязнением окружающей среды; колебания в содержании метаболитов и повышение стоимости производства в зависимости от условий культивирования, таких как время года, регион и климат. В связи с этим предпринимались активные попытки по получению вторичных метаболитов способом культуры in vitro, который дает такие преимущества, как возможность регулирования адекватных условий внешней среды и продуцирования в крупном масштабе даже в небольшом пространстве. Предшествующий уровень техники Согласно KR Patent 0130100 производство биологически активных веществ с использованием культуры растительных клеток имеет больше преимуществ перед прямой экстракцией их из растения. Культура растительных клеток рассматривается как оптимальный способ непрерывного производства, на который внешняя среда не оказывает никакого влияния, и как возможность решения вышеуказанных проблем, связанных с нарушениями экологии. Однако Nail & Roberts (2004) указывали на замедленную скорость роста и низкую продуктивность культуры растительных клеток для производства вторичных метаболитов. Для решения этой проблемы проводились исследования по оптимизации питательной среды, условий культивирования, процесса и по элиситации в целях повышения продуктивности и др. (Zhong, 2002). В Международном патенте WO 93/17121 используются разные питательные среды для получения культуры различных видов Taxus (тис, семейство хвойных) с целью увеличения скорости роста клеток и продуцирования paclitaxel (паклитаксел). По результатам экспериментов установлены условия элиситации для массового производства paclitaxel. Несмотря на успехи в производстве ценных вторичных метаболитов, вариабельность все еще остается главной проблемой производства paclitaxel из Taxus и других ценных веществ из многочисленных растительных систем. Производство вторичных метаболитов на промышленной основе с применением крупномасштабной культуры растительных клеток возможно только в случае стабильного поддержания быстрого роста клеток и высокого продуцирования метаболитов в долгоживущей культуре. Способность клеточных линий к продуцированию различных метаболитов не является стабильной, что служит причиной потери клеточными линиями их начальной продуктивности в процессе пересевов клеток в культуре; поэтому нет необходимости много говорить здесь о том, что успех и неудачи зависят от того, как преодолеваются эти проблемы. В культуре растительных клеток, хотя все клетки являются клетками одного растения, продуктивность образования метаболитов каждой клеточной линией различна и нестабильна. Следовательно, создание клеточных линий, обладающих высокой продуктивностью и генетической стабильностью, является важнее всего остального. Клеточные линии из клеток одного вида и клеток многих видов Линии растительных клеток из клеток одного вида показывают более низкую вариабельность, чем клеточные линии из клеток многих видов; это обусловливает их более высокую продуктивность. В изобретениях предшествующего уровня техники в качестве лучших эксплантатов для индукции клеточных линий использовались стебель, корень, семена, иголки (хвоя) и листья. Эти стебель, корень, семена, иголки и листья являются тканями, состоящими из клеток с различными функциями и морфологией. Каллус, клеточные линии, происходящие из этих тканей, не из их числа. Поэтому попытки по снижению изменчивости продуктивности каллуса из тканей, состоящих из клеток многих видов, довольно ограничены. Агрегация клеток Одной из отличительных особенностей культуры растительных клеток является агрегация клеток. Согласно патенту 0364478 диаметр растительной клетки составляет от 30 до 300 мкм, т.е. примерно в 30 раз превышает диаметр животной клетки. Вследствие того, что стенки растительных клеток показывают присущую им от природы тенденцию к слипанию друг с другом, невозможно получить суспензию, которая состояла бы только из диспергированных одиночных клеток. Соотношение и размер агрегатов клеток варьируют в зависимости от вида растения и питательной среды, в которой выращивалась культура. Nail & Roberts указывали на то, что агрегация клеток приводит к различиям в локальном окружении между внутренней и наружной частями клеток, что может послужить причиной гетерогенности культуры и в конечном итоге привести к изменениям в росте и метаболизме. Цель суспензионной культуры состоит в получении чистых одиночных клеток. Для достижения этой цели применялись фильтрация, мацерация и культура протопласта с использованием ферментов. Однако фильтрация и мацерация не обеспечивают полностью чистых одиночных клеток. Способ культуры протопласта, который удаляет клеточные стенки, является самым надежным способом генерирования одиночных клеток, но фермент, используемый для культуры протопласта, вызывает повреждение или разрушение клеточных стенок, что приводит к изменению физиологии клеток. Более того, гидрофобные вторичные метаболиты, такие как paclitaxel, могут накапливаться в клеточной стенке, поэтому изменения в клеточной стенке тесно связаны с продуктивностью. Таким образом, агрегация клеток долгое время служила главным препятствием для получения точных результатов измерения роста клеток по количеству и для проведения биохимических проб на отдельные клетки. Согласно Nail & Roberts (2004), если удастся выделить культуру из одной клетки, то в этом случае можно легко и более быстро получить информацию о поведении клеточных единиц в культуре, например о биосинтезе, накоплении, распаде и др. вторичных метаболитов. Дедифференциация Дедифференцированная клеточная линия, какой является каллус, показывает высокую вариабельность в продуцировании вторичных метаболитов вследствие сомаклональной вариации. Каллус, выделенный из постоянных тканей, таких как листья, стебель, корень и семена, которые состоят из клеток с различными функциями и морфологией, обычно показывает значительные изменения даже в слаборазличающихся микроокружениях, поскольку он является вторичной меристемой, образовавшейся путем дедифференциации. С учетом этой чувствительности Hirasuna et al. (1996) провели исследования с целью идентификации условий культивирования клеток, главным образом, таких, как начальная плотность посева клеток, интервал между пересевами и температура, и более точного, насколько это возможно, поддержания этих условий. Наращивание производства Для того чтобы поставить продуцирование вторичных метаболитов с применением культуры растительных клеток на промышленную основу, необходимо, прежде всего, увеличить масштабы этого продуцирования. После опубликования многих патентов и статей с сообщениями об успешном производстве метаболитов с применением клеточной культуры в лабораторном масштабе для массового производства метаболитов стал использоваться биореактор. Согласно патенту 0290004 применение биореактора для массового производства обеспечивает очень разное окружение для культуры из пробирки в лабораторном масштабе, что приводит к снижению скорости роста и продуктивности и к изменению метаболитов. Когда для массового производства стал применяться биореактор, то изменения скорости роста, продуктивности и метаболитов превратились в проблему на пути коммерциализации производства биологически активных веществ с применением клеточной культуры. С точки зрения эффективности перемешивания и аэрации для наращивания растительных клеточных культур предпочтительнее использовать биореактор, подача воздуха в который осуществляется от размещенного вне реактора источника электроэнергии, или биореактор с импеллером. Однако в биореакторе жизнеспособность клеток резко снижается, поскольку растительные клетки являются слабыми для воздействия на них сдвига. Поэтому необходим способ, позволяющий снизить воздействие сдвига. Причины чувствительности растительной клетки к сдвигу объясняются ее большим размером, жесткой стенкой клетки, агрегацией и экстенсивной вакуолизацией (Yokoi et al., 1993). Для решения этих проблем, возникающих в биореакторе, в предшествующем уровне техники исследовался биореактор, генерирующий низкое усилие сдвига благодаря регулированию скорости перемешивания и модификации импеллера. Однако результаты исследования оказались отрицательными, поскольку клеточные линии не способны преодолеть различия в микроокружении. Криоконсервация Криоконсервация позволяет длительно сохранять (поддерживать) клетки за счет приостановления большей части метаболических процессов в клетках в условиях чрезвычайно низкой температуры. Это позволяет извлекать клетки после криоконсервации без генетических изменений, изменений характеристик и способности к биосинтезу. С помощью криоконсервации можно предупредить потери клеток от заражения и минимизировать генетические изменения в непрерывных клеточных линиях. В GMP (Международные правила организации хорошей практики производства (ВОЗ)) консервация клеточных линий на длительный период является обязательной для стабильного снабжения сырьем. Обычно культивируемые животные клетки могут подвергаться криоконсервации на многие годы, но применимость подобной техники криоконсервации для культивированных растительных клеток вызывает сомнение. Культивированные растительные клетки являются гетерогенными и показывают множество различий в физиологии и морфологии. Поэтому растительные суспензионные клетки требуют проведения многократного процесса криоконсервации, а неадекватная криоконсервация может стать причиной вариабельности. Кондиционирующие факторы Kim et al. (2000) продемонстрировали, что деление клеток можно стимулировать путем добавления некоторого количества среды из активно делящихся культур к культурам, клетки в которых потеряли способность к делению. В производстве антоцианина с использованием суспензионной культуры розы продуктивность повышалась, когда некоторое количество среды из суспензионной культуры земляники добавили к суспензионной культуре розы. Поэтому факторы, продуцируемые и секретируемые из культивированных клеток для стимуляции роста клеток или продуцирования вторичных метаболитов, называются кондиционирующими факторами. Однако эти кондиционирующие факторы конкретно не были идентифицированы, и существует лишь такое понятие – кондиционирующие факторы, которые действуют как химические сигналы для роста клеток и продуцирования метаболитов. Так, имеется несколько сообщений об активных веществах, таких как фосфаты и кальмодий (calmodium), которые могут считаться кондиционирующими факторами. Кондиционирующие факторы могут поставляться через кондиционированные среды или клетки-хелперы. Перфузионное культивирование Среди способов культуры клеток существует способ периодического культивирования, включающий совместную инокуляцию клетки и среды в начале введения питательных веществ без последующего их добавления. Существует также способ непрерывного культивирования, предусматривающий добавление новой питательной среды одновременно с удалением отработанной питательной среды, которая содержит метаболиты, при соответствующей скорости в период культивирования с целью предупреждения истощения питательных веществ. Периодическое культивирование трудновыполнимо на промышленном уровне из-за его низкой продуктивности. Среди способов непрерывного культивирования в последнее время все большее внимание привлекает перфузионное культивирование. В перфузионной культуре клетки остаются в биореакторе, а новая питательная среда добавляется по мере удаления отработанной питательной среды, которая содержит метаболиты. Согласно Zhang et al. (2000) элиситация является одним из наиболее эффективных путей стимулирования продуцирования вторичных метаболитов в клеточной культуре. Элиситация активизирует синтез вторичных метаболитов, но индуцирует ингибирование роста клеток и быстрое снижение их жизнеспособности. Следовательно, синтез вторичных метаболитов элиситацией можно поддерживать только в течение короткого периода времени и он очень ограничен. В интерпретации Wang et al. (2001) перфузионное культивирование – это стратегия минимизации указанных негативных воздействий элиситации и максимизации продуктивности. Wang et al. (2001) и Wu & Lin (2003) сообщили следующее. Вторичные метаболиты, продуцируемые элиситацией, накапливаются внутри клетки (в вакуолях или клеточной стенке) или высвобождаются из клетки (питательной среды). В процессе культивирования высвобождение вторичных метаболитов из клетки и удаление ее из питательной среды может облегчить очистку и уменьшить торможение биосинтеза по типу обратной связи и распад и конверсию продуктов. Таким образом, за счет удаления отработанной питательной среды и добавления новой питательной среды внутренняя и внешняя секреция метаболитов может продлить период жизнеспособности и биосинтез клеток. И это может заметно увеличить продуктивность. Хранение и секреция вторичных метаболитов показывают значительные различия в зависимости от клеточных линий. Линия клеток Taxus media (Wickremesinhe and Arteca 1994) ничего не выделяла. Следовательно, требуется создание такой клеточной линии, которая обладает выдающейся секретирующей способностью. Культура камбия Камбий является боковой меристемой, которая располагается на боковой стороне растения. У голосемянных и древесных двудольных растений наблюдается гипертрофический рост, обусловленный почти не ослабевающей активностью прокамбия или камбия; результатом этого является существование гигантских растений, имеющих более 11000 годичных колец. В генетике меристемы классифицируются как первичная и вторичная меристема. Первичная меристема представляет собой меристему, которая формируется в процессе эмбриогенеза и участвует в росте растения после прорастания семян. Вторичная меристема представляет собой меристему, которая образуется в результате дедифференциации постоянной ткани растения. Камбий является первичной меристемой с меристематической непрерывностью, берущей начало из прокамбия без вмешательства постоянной ткани. Рост этой первичной меристемы не ограничивается только развитием верхушечной почки и может продолжаться, если для этого будут созданы условия. Поэтому культура камбия использовалась для быстрого массового размножения клеток. В ранее проводившихся исследованиях эксплантаты камбия получали следующим образом: после очищения древесного стебля от вторичной коры в нем делали два доходящих до ксилемы продольных разреза на расстоянии 5 мм друг от друга и глубиной примерно 1 мм. Эти эксплантаты были названы исследователями “камбием”, который состоял из части флоэмы, камбия и небольшого кусочка ксилемы (Jouira et al., 1998). Уместно будет сказать, что клетки, индуцированные вышеупомянутым способом, происходят не из одного камбия, а из многих тканей, которые могут иметь серьезные анатомические различия, а именно – из флоэмы, камбия и ксилемы. Поэтому, по мнению авторов настоящего изобретения, вышеупомянутый способ не является идеальной техникой для тщательного выделения только камбия из различных тканей, составляющих стебель. В связи с этим существует потребность в создании способа выделения только камбия из различных тканей стебля. Раскрытие изобретения Техническая проблема Целью настоящего изобретения является создание способа получения клона одной клетки путем выделения и культивирования только камбия из побега или стебля. Если говорить более конкретно, то целью изобретения является решение проблемы вариабельности в растительной клеточной культуре и разработка способа стабильного производства растительных биологически активных веществ путем выделения только камбия с применением комбинированных способов химии клетки и физиологической химии в отличие от используемого в предшествующем уровне техники способа физического разделения с помощью скальпеля. Другой целью настоящего изобретения является выделение и культивирование только камбия из побегов Taxus (тис) и создание способа производства паклитаксела (paclitaxel). Техническое решение Для достижения вышеуказанных целей настоящее изобретение в одном аспекте обеспечивает способ выделения клона одной растительной клетки из камбия, который включает: (а) подготовку и последующую стерилизацию растительной ткани; (б) сбор ткани, содержащей камбий, из указанной стерилизованной растительной ткани; (в) культивирование указанной ткани, содержащей камбий, и индуцирование тем самым слоя камбия, пролиферирующего из камбия, и слоя каллуса, извлеченного из других участков, за исключением камбия, и пролиферирующего в неправильной форме; и (г) сбор клона одной клетки путем отделения указанного слоя камбия от указанного слоя каллуса. Предпочтительно стадия (в) включает культивирование указанной ткани в питательной среде, содержащей ауксин. И в предпочтительном варианте воплощения изобретения питательная среда содержит 1~3 мг/л ауксина. В другом аспекте настоящее изобретение обеспечивает клон одной клетки, индуцированный из растительного камбия, при этом клон одной клетки имеет следующие характеристики: (а) свыше 90% клеток в суспензионной культуре существуют как одиночные клетки; (б) имеет множество вакуолей морфологически; (в) растет быстрее, чем клеточная линия из других участков, за исключением камбия, того же растения, и культивируется стабильно в течение длительного времени; (г) обладает низкой чувствительностью к усилию сдвига в биореакторе; и (д) является по своей природе недифференцированным. Предпочтительно растение является растением рода Taxus (тис). И в предпочтительном варианте воплощения изобретения клон одной клетки, выделенный из камбия растения рода Taxus, обладает способностью к высвобождению paclitaxel (паклитаксел), которая в 270~720 раз превышает такую же способность клеточных линий, выделенных из других участков того же растения, за исключением камбия. В еще одном аспекте настоящее изобретение обеспечивает способ продуцирования растительных биологически активных веществ, включающий стадии: (а) продуцирования активных веществ путем культивирования вышеупомянутого клона одной клетки и (б) сбора указанных активных веществ. Предпочтительно культивирование на стадии (а) включает удаление питательной среды, используемой для культивирования культуры вышеуказанного клона одной клетки, и добавление новой питательной среды. В предпочтительном варианте воплощения изобретения клон одной клетки является клоном одной клетки, выделенным из камбия растения рода Taxus, а соединение является паклитакселом (paclitaxel). В этом случае питательная среда может также содержать один или более материалов, выбранных из группы, состоящей из метилжасмоната, фенилаланина и хитозана. И, наконец, настоящее изобретение обеспечивает способ консервирования растительной клеточной линии, который включает криоконсервацию клона одной клетки, выделенного из растительного камбия, который был изолирован вышеуказанным способом. Преимущества С помощью способов настоящего изобретения возможно получение культуры клона одной клетки, который обладает меристематической непрерывностью первичной меристемы без участия дедифференциации, путем точного выделения только камбия из различных тканей побега или стебля древесного растения. Клеточная линия настоящего изобретения обеспечивает стабильное продуцирование биологически активных веществ, обусловленное снижением изменчивости скорости роста клеток и характера роста в процессе долгосрочного культивирования. Указанная линия клеток является оптимальной и для крупномасштабного производства на промышленном уровне, поскольку она менее чувствительна к усилию сдвига в биореакторе по сравнению с линиями клеток, используемыми в способах предшествующего уровня техники, благодаря ее пониженной способности к агрегации и значительной вакуолизации. Активация метаболитов может стимулироваться добавлением кондиционирующих факторов к этой клеточной линии, а жизнеспособность клеток и биосинтез могут продлеваться во времени по мере высвобождения из клеток значительного количества продукции во внеклеточную среду в ходе перфузионного культивирования. Высокая скорость восстановления после криоконсервации, обусловленная гомогенностью и способностью указанной линии клеток к делению, позволяет создать банк клеток. Настоящим изобретением подтверждается тесная взаимосвязь между гомогенностью культуры и изменчивостью вторичных метаболитов, а способ настоящего изобретения помогает разработать стратегию коммерциализации, поскольку он регулирует и снижает вариабельность в продуцировании различных биологически активных веществ. Описание фигур Фиг.1 показывает часть, выделенную в ходе индукции одноклеточного клона из камбия. Фиг.2 показывает скорость роста, выраженную как общее производство биомассы тремя различными клеточными культурами из камбия, зародыша и иголок. Фиг.3 показывает изображение агрегации клеток в культурах, полученных из двух различных тканей (зародыша или иголок и камбия). Фиг.4 показывает влияние элиситации на продуцирование paclitaxel в процессе суспензионного культивирования Taxus. Фиг.5 показывает влияние кондиционирующих факторов на продуцирование paclitaxel в процессе суспензионного культивирования Taxus. Способы изобретения Примеры изобретения объясняются ниже. Способ индукции и пролиферации клона одной клетки камбия используется не только в системе производства paclitaxel, но и может использоваться также в системе производства всех растительных вторичных метаболитов. Нижеследующие примеры даются только в целях иллюстрации изобретения и не ограничивают его масштаб. Пример 1. Подготовка растительных материалов и выделение камбия Собирали семена, иголки, побеги тиса. Собранные материалы сразу опускали в раствор 100 мг/л антиоксиданта – аскорбиновой кислоты (L-аскорбиновая кислота, DUCHEFA, Нидерланды), пересевали и консервировали. Их стерилизовали с поверхности с учетом морфологии и физиологических характеристик материалов. (1) Семена: после стерилизации семян 70%-м этанолом в течение одной минуты их погружали в 1% раствор Clorox на 48 часов и промывали от 3 до 4 раз стерильной водой. Затем из семян извлекали зародыши в раствор, содержащий 0,5% PVP (поливинилпирролидон, DUCHEFA, Нидерланды), 50 мг/л аскорбиновой кислоты (L-аскорбиновая кислота, DUCHEFA, Нидерланды) и 70 мг/л лимонной кислоты (DUCHEFA, Нидерланды), и проращивали на питательной среде для индукции каллуса. (2) Иголки и побеги: после 24-часовой обработки раствором, содержащим 1% Benomyl (Dongbu Hannong Chemical, Южная Корея)+1% Daconil (Dongbu Hannong Chemical, Южная Корея)+1% сульфата стрептомицина (DUCHEFA, Нидерланды)+0,1% Cefotaxime sodium (DUCHEFA, Нидерланды), иголки и побеги ополаскивали водопроводной водой в течение 30 секунд для удаления остатков химических веществ и фенольных соединений. После стерилизации их последовательно 70% этанолом (DC Chemical, Южная Корея) в течение одной минуты, 30% пероксидом водорода (LG Chemical, Южная Корея) в течение 15 минут, 1% раствором CLOROX в течение 15 минут и 3% раствором CLOROX в течение 5 минут их промывали от 3 до 4 раз дистиллированной водой. Для предупреждения окисления оба конца иголок отрезали в раствор, содержащий 5% PVP, 50 мг/л аскорбиновой кислоты и 70 мг/л лимонной кислоты, и культивировали на питательной среде для индукции каллуса. (3) Выделение камбия из побега или стебля: удерживая ксилему, являющуюся центральной частью побега или стебля, пинцетами, счищали флоэму, кору и ткани эпидермиса, включая камбий. Эту счищенную ткань, содержащую камбий, помещали на питательную среду таким образом, чтобы камбий соприкасался с поверхностью среды. Пример 2. Индукция одноклеточного клона из выделенного камбия После 4-7 суток культивирования наблюдалось деление клеток камбия, а на 15 сутки начал формироваться каллус из слоя, состоявшего из флоэмы, коры и эпидермиса, которые были верхней частью камбия. На 30 сутки культивирования камбий начал отделяться от верхнего слоя ткани, содержавшего флоэму, кору и эпидермис; после того как оба этих слоя полностью разделились естественным путем, их по отдельности культивировали на разных чашках Петри (фиг.1). В целях индукции клеток и каллуса можно использовать известные универсальные питательные среды для культивирования растительных клеток и тканей, например mВ5 (модифицированная среда Gamberg’s B5), MS (среда Murashige & Skoog), WPM (LLoyed & McCown), SM (среда Schenk & Hildebrand), LP (Quoirin & Lepiovre). Возможно применение всех этих сред. При необходимости можно вносить в среды различные добавки либо снижать количество компонентов сред или полностью исключать их. Из перечисленных сред наиболее подходящей является среда mB5. Состав среды тВ5 приводится в табл.1.
Культуры выращивали на питательной среде, в которую был добавлен регулятор роста растений – ауксин (1-3 мг/л), в темноте при 25±1°С. Камбий состоял из гомогенных клеток, поэтому деление клеток было равномерным, а пролиферация происходила в форме пластинки. С другой стороны, ткань, содержавшая флоэму, кору и эпидермис, пролиферировала в неправильной форме из-за различий в составе многих видов клеток. Наблюдалось саморасщепление слоя между камбием и тканью, содержавшей флоэму, кору и эпидермис (фиг.1). Камбий был гомогенным, а ткань, содержавшая флоэму, кору и эпидермис, гетерогенной, поэтому саморасщепление слоя было, по-видимому, результатом различной скорости деления. Спустя 15 суток культивирования на эксплантатах зародыша и игл в результате дифференциации образовались каллусы, состоявшие из гетерогенных клеток, и эти каллусы пролиферировали в неправильных формах из-за разной скорости деления различных клеток, подобно ткани, содержавшей флоэму, кору и эпидермис (фиг.1). Пример 3. Получение долгоживущей культуры Из каллусов белые и ломкие каллусы, показавшие хорошую скорость роста, пересевали на новую питательную среду каждый 21 день. Скорость роста культур зародыша и иголок была очень нестабильной, и эти культуры часто показывали тенденцию к потемнению. В противоположность этому скорость роста культур камбия была высокой, а изменения окраски культур не наблюдалось. Таким образом, появилась возможность выбора стабильных клеток. Спустя шесть месяцев культивирования большая часть культур зародыша и иголок имела желтую или светло-коричневую окраску и образовала агрегаты. Культуры камбия имели бело-желтую окраску и сохранялись как одиночные клетки или небольшие кластеры клеток. Скорость роста культур, которые потемнели и образовали агрегаты, замедлилась, и в конечном итоге, эти культуры погибли из-за присутствия химического вещества – фенола, который они выделяли. Согласно автору настоящего изобретения спустя 6 месяцев начались трудности с поддержанием и массовой пролиферацией культур зародыша и иголок, а культуры камбия стабильно сохранялись в течение более 20 месяцев долгосрочного культивирования без изменения скорости роста клеток, характера роста и уровня агрегации (фиг.2). Другими словами, вариабельность в характере роста зависит, по-видимому, от гомогенности и гетерогенности исходных растительных материалов. Пример 4. Получение суспензионной культуры клеток Культуры зародыша и иголок и камбия культивировались по отдельности в склянке, содержащей жидкую питательную среду (табл.2).
Они культивировались на встряхивающем устройстве, вращающемся со скоростью 100 об/мин, в темноте при 25±1°С. В условиях пересевов с двухнедельным интервалом постоянно поддерживалась высокая жизнеспособность культур как фаза экспоненциального роста. Измерялся уровень агрегации, который служит основной причиной варьирования продуктивности клеток. Проводилась количественная оценка клеточных агрегатов с применением биологического микроскопа (СХ31, Olympus, Япония). Результаты вышеописанного эксперимента приводятся в табл.3.
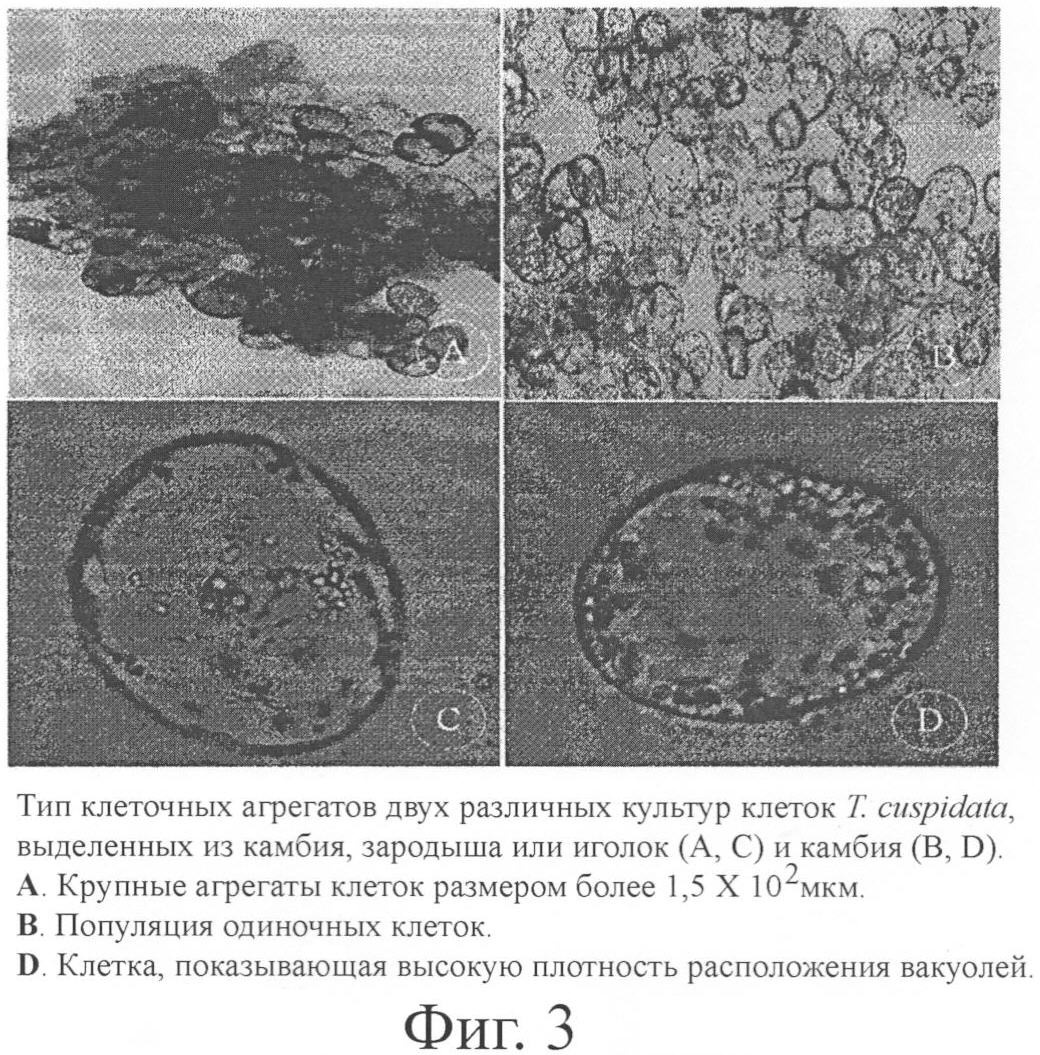
Крупные агрегаты клеток размером более 1,5×103 мкм; Умеренные агрегаты клеток: 1×103 мкм; Мелкие агрегаты клеток: 4×102 мкм < размер < 1×103 мкм. В случае суспензии культур зародыша и иголок около 60% клеток образовали агрегаты размером более 1,5 мм, а в суспензии культур камбия 90% клеток культивировались как одиночные клетки. Пример 5. Наращивание производства Культуры зародыша и иголок и камбия культивировались в биореакторе с эрлифтом 3L (Swig-Won SciTech, Южная Корея) в темноте при 25±1°С. В случае культур зародыша и иголок отмечалась значительная вариабельность в размерах и форме клеток по сравнению с культурой в пробирке. Диаметр клеточных агрегатов увеличился до 2-3 мм, что тормозило поток внутри биореактора и послужило причиной образования в биореакторе неперемешиваемой зоны. Клетки, налипшие на внутренней стенке биореактора, образовали годичное кольцо. Клетки в центре годичного кольца погибли спустя 20 суток из-за недостаточного поступления питательной среды. В конечном итоге, отмершие клетки выделили токсические вещества, и эти вещества снизили жизнеспособность всех клеток в биореакторе. В противоположность этому меньшая степень агрегации культур камбия способствовала равномерной циркуляции воздуха в биореакторе, вследствие чего появилась возможность сократить количество подаваемого воздуха с 200 мл до 150 мл в минуту и значительно уменьшить количество пузырьков, образовавшихся на поверхности питательной среды. Время удвоения культур зародыша и иголок в пробирке составило 12 суток, а в биореакторе оно увеличилось до 21 суток. Это явилось результатом образования годичного кольца и быстрого снижения жизнеспособности клеток вследствие их чувствительности к сдвигу, вызванной агрегацией клеток и жесткой клеточной стенкой. Время удвоения культур камбия составило от 4 до 5 суток, и в этом случае не наблюдалось различий между культурой в пробирке и в биореакторе; в биореакторе оно скорее сократилось (табл.4). Культуры камбия образовали очень маленькое годичное кольцо в биореакторе, и это годичное кольцо легко растворилось при перемешивании среды обыкновенной палочкой. Более того, не было отмечено снижения жизнеспособности клеток, поскольку чувствительность к сдвигу была ниже вследствие меньшей агрегации клеток и множественной вакуолизации.
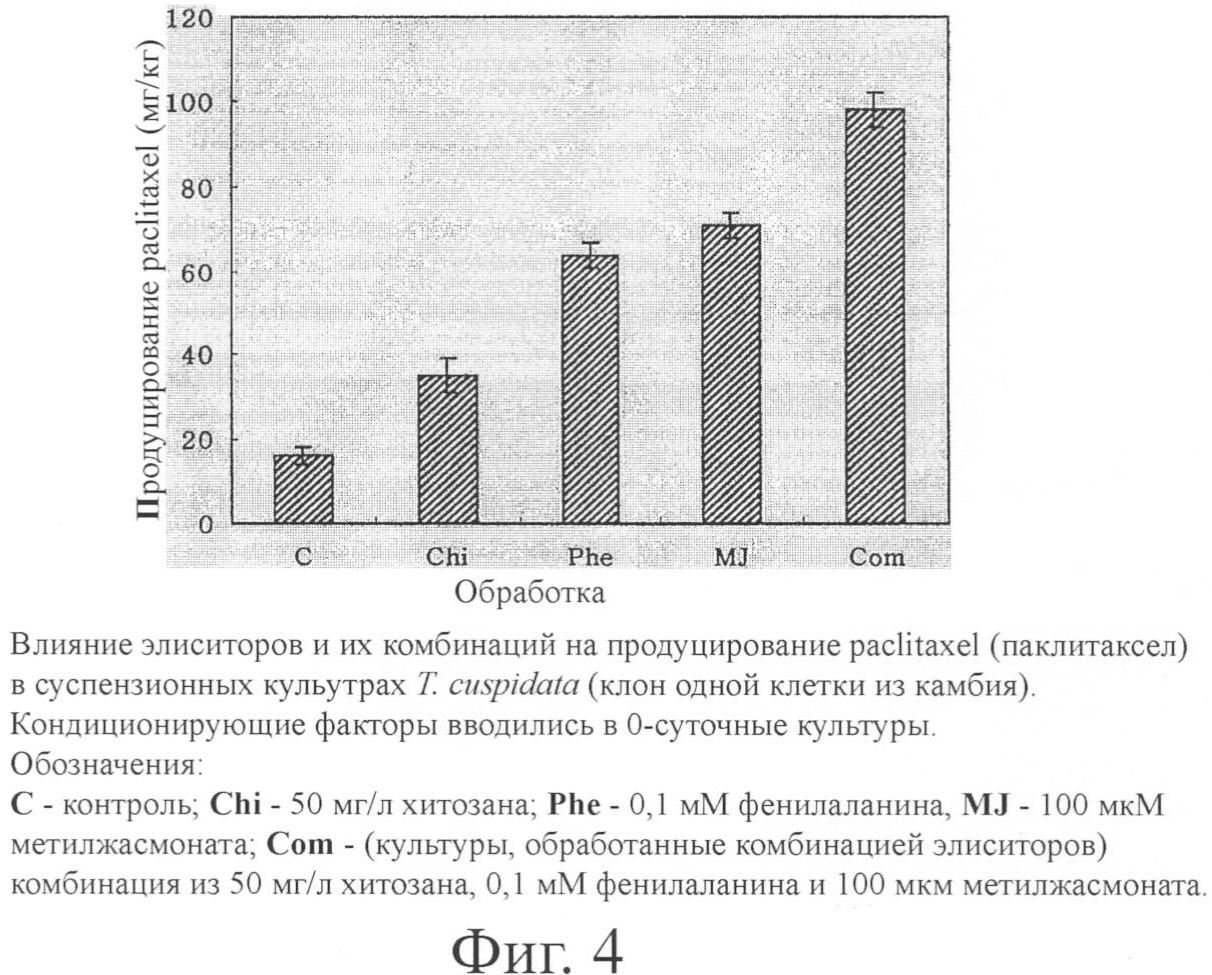
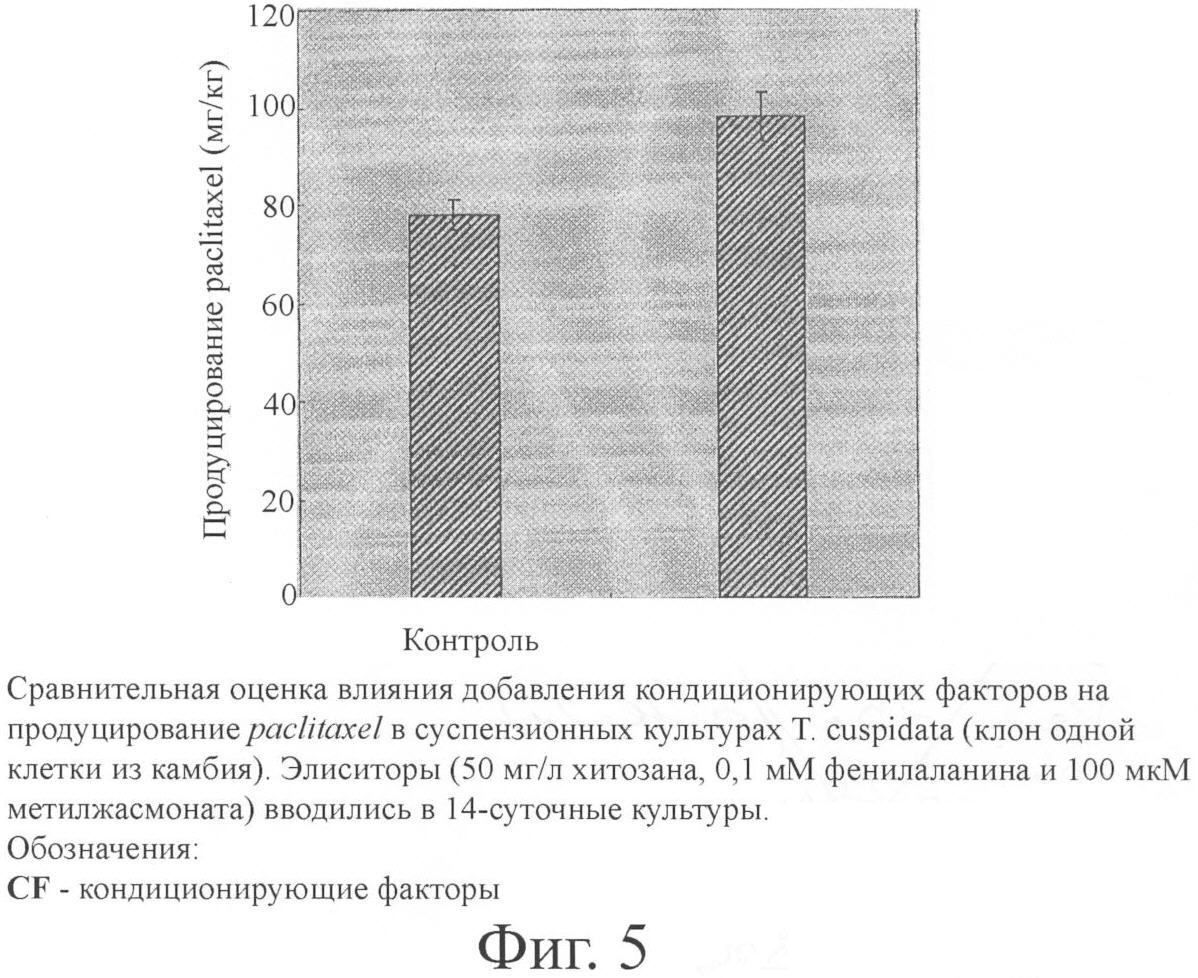
Пример 6. Элиситор Элиситор контролирует молекулярный сигнал в растительных клетках и широко используется для увеличения продуцирования вторичных метаболитов. По результатам обработки метилжасмонатом в качестве элиситора и 10 других видов элиситоров авторами настоящего изобретения был отмечен положительный эффект метилжасмоната на продуцирование paclitaxel (паклитаксела). Появилась возможность достигнуть относительно высокого продуцирования метаболитов с помощью комбинации метилжасмоната с другими элиситорами. Продуцирование paclitaxel было особенно эффективным в случае обработок метилжасмонатом, хитозаном и фенилаланином (фиг.4). Пример 7. Кондиционирующие факторы Вторичные метаболиты растений продуцируются в процессе роста клеток или после прекращения роста клеток. Поэтому для производства таких метаболитов, как paclitaxel, более всего пригодно двухстадийное культивирование с раздельным проведением первой стадии роста клеток и второй стадии образования метаболитов. В настоящем примере на первой стадии пролиферация клеток достигала большого масштаба за счет оптимизации роста клеток, а на второй стадии условия культивирования были изменены с целью оптимизации продуцирования метаболитов. Клеточные линии с высокой способностью к продуцированию вторичных метаболитов росли медленнее и отмирали быстрее, чем клеточные линии с низкой способностью к образованию вторичных метаболитов. Поэтому массовая пролиферация была затруднена, и крупномасштабное производство метаболитов стало невозможным. В настоящем изобретении клеточные линии с низкой способностью к пролиферации и высокой способностью к продуцированию метаболитов не использовались для пролиферации в крупном масштабе; они скорее применялись как клетки-хелперы, имеющие кондиционирующие факторы для продуцирования вторичных метаболитов. Авторы изобретения наблюдали образование paclitaxel после добавления клеток-хелперов. Результаты суммированы на фиг.5. Пример 8. Перфузионное культивирование На 14 сутки культивирования культуры зародыша и иголок и камбия обрабатывали элиситором. Начиная с момента элиситации, отработанная питательная среда отбиралась в асептических условиях с помощью пипетки каждые 5 суток и одновременно заменялась таким же количеством новой питательной среды. Продуцирование paclitaxel в клетках и среде наблюдалось спустя 45 суток долгосрочного культивирования. Результат суммирован в табл.5.
Обновление питательной среды, введенной в клеточные культуры спустя 5 суток после элиситации, которая проводилась путем добавления 50 мг/л хитозана, 0,1 мМ фенилаланина и 100 мкМ метилжасмоната к 14-суточным культурам. Эксперимент с повторным обновлением питательной среды каждые 5 суток. В зависимости от клеточных линий высвобождение paclitaxel из клеток в среду было различным. Высвобождающая способность культур камбия превосходила высвобождающую способность культур в способах предшествующего уровня техники. Более того, применение перфузионного культивирования облегчало высвобождение вторичных метаболитов в среду. Улучшение внеклеточного высвобождения вторичных метаболитов за счет использования клона одной клетки камбия путем периодической замены питательной среды имело важное значение, поскольку это обеспечивало непрерывную рециркуляцию биомассы и упрощало очистку. Другими словами, периодическую замену питательной среды в культуре одноклеточного клона камбия можно рассматривать как стабильный способ продуцирования ценных метаболитов в долгоживущей культуре, так как он предупреждает задержку по типу обратной связи накопленных метаболитов в клетке, распад и конверсию метаболитов в питательной среде. Пример 9. Криоконсервация На 6 или 7 сутки культивирования клетки суспензионной культуры предварительно культивировалась в питательной среде, содержащей 0,16 М маннита, в течение 3 суток при комнатной температуре, а затем поддерживались (сохранялись) при 4°С в продолжение 3 часов. Затем проводился сбор выросших в культуре клеток, и они переносились в криофлакон емкостью 4 мл со средой, содержавшей 40% этиленгликоля (Sigma, США) и 30% сорбита (DUCHEFA, Нидерланды), где культивировались в течение 3 минут при 4°С. Клетки суспензионной культуры, обработанные криоконсервантами, замораживались после погружения в жидкий азот. Размораживание замороженных погружением в жидкий азот на более чем 10 минут культивированных клеток проводилось на водяной бане 40°С в продолжение 1-2 минут. Для повторного выращивания криоконсервированные клетки переносились на полутвердую ростовую среду (табл.1), содержащую 0,5 М сорбита, и размягчались при комнатной температуре в течение 30 минут. Клетки культивировались на полутвердой ростовой среде, содержавшей 0,1 М сорбита, в течение 24 часов, а затем дважды культивировались на полутвердой ростовой среде без сорбита в продолжение 24 часов. После этого оценивалась жизнеспособность клеток. Пример 10. Анализ содержания paclitaxel После отделения клеток от среды отобранных образцов анализировалось содержание paclitaxel. Масса клеток измерялась после полного их высушивания в вакуумном эксикаторе (Sam Shin Glass, Южная Корея). Примерно 100 мг (в пересчете на сухую массу) клеток смешивались с 4 мл раствора (1:1 об/об) метанола (Sigma, США) и метилхлорида (Sigma, США) и трижды экстрагировались ультразвуковым очистителем (Branson, США) с интервалом в один час при комнатной температуре. Клетки полностью высушивались и экстрагировались несколько раз с использованием 4 мл метилхлорида. Отделенный слой органического растворителя подвергался вакуумной сушке, а осадок растворялся в 1 мл метанола. Растворенный экстракт равномерно перемешивался ультразвуковым очистителем. Затем после центрифугирования (8000 g, 5 мин) осадок удалялся. Среда (1-5 мл), которая была отделена от клеток, объединялась с таким же объемом метилхлорида и после полного вымешивания трижды экстрагировалась. После удаления органического растворителя под вакуумом и полного высушивания она вновь растворялась в 0,5 мл метанола. Для анализа содержания стандартных веществ paclitaxel применялся способ HPLC (жидкостная хроматография высокого разрешения, Shiseido, Япония) и реактивы фирмы Sigma. Для поддержания Capcell pak (С 18, MGII, 5 мкм, 3,0 мм × 250 мм, Shiseido, Япония) при 40°С использовалась печь, а вода и ацетонитрил (Burdick & Jackson, США) (50:50 об/об) объединялись в качестве подвижной фазы и равномерно стекали по каплям со скоростью 0,5 мл/мин. Использовался UV-VIS-детектор (в ультрафиолетовой и видимой частях спектра) (Shiseido, Япония). Промышленное применение В настоящем изобретении получение клона одной клетки первичной меристемы, показывающей меристематическую непрерывность без дедифференциации, путем выделения только камбия из побега или стебля приводит к повышению продуктивности вследствие сокращения времени удвоения по сравнению с клеточными линиями в способах предшествующего уровня техники. Это обеспечивает также стабильную продуктивность в результате меньшего варьирования скорости роста клеток и характера роста в ходе долгосрочного культивирования и делает возможным увеличение масштаба производства вследствие меньшей агрегации и множественной вакуолизации клеточных линий. Эти клеточные линии можно выделять после криоконсервации без генетических изменений. Список литературы 1. Gamborg, O.L., Miller, R.A., Ojima, K. 1968. Nutrient requirement of suspension cultures of soybeans root cells (Потребность в питательных веществах суспензионных культур клеток корня соевых бобов). Exp. Cell. Res., 50: 151. 2. Hirasuna T.J., Pestchanker L.J., Srinivasan V., Shuler M.L. (1996). Taxol production in suspension cultures of Taxus baccata (Получение таксола в суспензионных культурах Taxus baccata). Plant Cell Tissue and Organ Culture. 44: 95-102. 3. Jouira H.B., Hassairi A., Bigot C., Dorion N. (1998). Adventitious shoot production from strips of stem in the Dutch elm hybrid ‘Commelin’: plantlet regeneration and neomycin sensitivity (Адвентициальное получение побегов из полосок стебля голландского гибрида ильма ‘Commelin’: регенерация побегов и чувствительность к неомицину). Plant Cell Tissue and Organ Culture. 53: 153-160. 4. Kim M.H., Chun S.H., Kim D.I. (2000). Growth promotion of Taxus brevifolia cell suspension culture using conditioned medium (Стимуляция роста суспензионной культуры клеток Taxus brevifolia за счет использования кондиционированной питательной среды). Biotechnol. Bioprocess. Eng. 5: 350-354. 5. Lloyd G. and McCown В. (1980). Commercially-feasible micropropagation of mountain laurel (Kalmia latifolia) by use of shoot-tip culture (Реализация в промышленных условиях вегетативного размножения кальмии широколистной (Kalmia latifolia) путем использования культуры верхушек побегов). Plant Prop. Proc. 30: 421. 6. Muraghige T. and Skoog F. (1962). A revised medium for rapid growth and bioassays with tobacco culture (Модифицированная питательная среда для быстрого роста и биоанализа культуры табака). Physiol. Plant. 15: 473-497. 7. Nail M.C., Roberts S.C. (2004). Preparation of single cells from aggregated Taxus suspension cultures for population analysis (Выделение одиночных клеток из агрегатированных суспензионных культур Taxus для анализа популяции). Published online 10 may 2004 in wiley interscience (опубл. в режиме он-лайн 10 мая 2004 г. в wiley interscience). 8. Quoirin M., Lepoivre P. (1977). Acta. Hart. 78: 437. 9. Schenk R.U., Hildebrandt A.C. (1972). Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell culture (Питательная среда и способика индуцирования и роста культуры клеток однодольных и двудольных растений). Can. J. Bot. 50: 199. 10. Wang C., Wu J., Mei X. (2001). Enhanced taxol production and release in Taxus chinensis cell suspension cultures with selected organic solvents and sucrose feeding (Ускорение продуцирования и высвобождения таксола (taxol) в суспензионных культурах клеток Taxus chinensis с применением селективных органических растворителей и подпитки сахарозой). Biotechnol. Prog. 17: 89-94. 11. Wang С., Wu J., Mei X. (2001). Enhancement taxol production and excretion in Taxus chinensis cell culture by fungal elicitation and medium renewal (Ускорение продуцирования и выделения таксола в культуре клеток Taxus chininsis элиситацией грибами и обновлением питательной среды). Appl. Microbiol. Biotechnol. 55: 404-410. 12. Wickremesinhe E.R.M., Arteca R.N. (1994). Taxus cell suspension cultures: optimizing growth and production taxol (Суспензионные культуры клеток Taxus: оптимизация роста и производства таксола). J. Plant Physiol. 144: 183-188. 13. Wu J., Lin L. (2003). Enhancement taxol production and release in Taxus chinensis cell cultures by ultrasound, methyl jasmonate and in situ solvent extraction (Ускорение продуцирования и высвобождения таксола в культурах клеток Taxus chinensis ультразвуком, метилжасмонатом и экстракцией растворителем in situ). Appl. Microbiol. Biotechnol. 62: 151-155. 14. Yokoi H., J. K-oga, K. Yamamura and Y. Seike (1993). High density cultivation of plant cells in a new aeration-agitation type fermentor (Культивирование растительных клеток при высокой плотности посева в новом ферментере аэрационно-перемешивающего типа). 75: 48-52. 15. Zhang C.H., Mei X.G., Liu L., Yu L.J. (2000). Enhanced paclitaxel production induced by the combination of elicitors in cell suspension cultures of Taxus chinensis (Ускоренное продуцирование паклитаксела, индуцируемое комбинацией элиситоров в суспензионных культурах клеток Taxus chinensis). Biotechnology Letters. 11: 1561-1564. 16. Zhong J.J. (2002). Plant cell culture for production of paclitaxel and other taxanes (Культура растительных клеток для производства паклитаксела и других таксанов). J. Bioscience and Bioengineering. 94: 591-599.
Формула изобретения
1. Способ выделения клеток, происходящих из камбия растения, включающий: 2. Способ по п.1, где ткань, содержащую камбий растения согласно стадии (а), стерилизуют. 3. Способ по п.1, где стадия (в) включает выделение указанного слоя камбия из слоя каллуса (наплыва), извлеченного из других районов за исключением камбия и пролиферирующего в неправильной форме. 4. Способ по п.1, где стадия (б) включает культивирование указанной ткани в среде, содержащей ауксин. 5. Способ по п.4, где среда содержит 1-3 мг/л ауксина. 6. Способ по п.1, где растение относится к роду Taxus. 7. Выделенные клетки, полученные из камбия растения способом по п.1, имеющие следующие характеристики: 8. Выделенные клетки по п.7, полученные из слоя, индуцированного в ткани растения, включающей камбий растения и слой, пролиферирующий из камбия ткани, и не включают каллус, происходящий из растения. 9. Выделенные клетки по п.7, где растение относится к роду Taxus. 10. Выделенные клетки по п.9, полученные из камбия рода Taxus, способные высвобождать в 270-720 раз больше паклитаксела, чем клетки, полученные из других районов кроме камбия рода Taxus. 11. Способ продуцирования биологически активных веществ растительного происхождения, включающий стадии: (а) продуцирование активных веществ путем культивирования клеток, выделенных согласно способу по п.1, и (б) сбор указанных активных веществ. 12. Способ по п.11, в котором стадия культивирования (а) включает отбор питательной среды, применяемой при культивировании указанной выделенной клеточной культуры, и последующее добавление новой питательной среды. 13. Способ по п.11, в котором выделенные клетки представляют собой выделенные клетки, полученные из камбия рода Taxus, и активное вещество представляет собой паклитаксел. 14. Способ по п.13, в котором выделенные клетки, полученные из камбия рода Taxus, культивируют в среде, дополнительно содержащей один или более материал, выбранный из группы, состоящей из метилжасмоната, фенилаланина и хитозана. 15. Способ консервации растительной клеточной линии, включающий криоконсервацию клеток, полученных из растительного камбия, выделенных по способу по п.1.
РИСУНКИ
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

 -нафталин-уксусная кислота
-нафталин-уксусная кислота