Патент на изобретение №2398777
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) ГУМАНИЗИРОВАННЫЕ АНТАГОНИСТЫ, НАПРАВЛЕННЫЕ ПРОТИВ c-met
(57) Реферат:
Настоящее изобретение относится к области биотехнологии и представляет собой гуманизированную форму мышиного антитела против c-met. Кроме того, изобретение относится к композиции, содержащей антитела по изобретению, а также к способам применения таких антител. Антитела по изобретению позволяют в различной степени нарушать пути передачи сигнала HGF/c-met. 8 н. и 21 з.п. ф-лы, 18 ил., 2 табл.
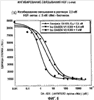
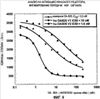
Родственные заявки Настоящая заявка представляет собой действительную заявку, поданную в соответствии со ст. 37 СFR (Свода Федеральных правил) 1.53(b)(1), которая согласно ст. 35 USC (Кодекса законов США) 119(е) испрашивает приоритет предварительной заявки Область, к которой относится изобретение В общих чертах настоящее изобретение относится к области молекулярной биологии и регуляции факторов роста. Более конкретно настоящее изобретение относится к модуляторам пути передачи сигнала HGF/c-met и к применению указанных модуляторов. Предшествующий уровень техники HGF представляет собой мезенхимальный плейотропный фактор, обладающий митогенной, мотогенной и морфогенной активностью в ряде клеток различных типов. Действие HGF опосредуется специфической тирозинкиназой, c-met и аберрантным HGF, причем экспрессия c-met часто наблюдается в различных опухолях (см., например, Maulik et al., Cytokine & Growth Factor Reviews (2002), 13:41-59; Danilkovich-Miagkova & Zbar, J. Clin. Invest. (2002) 109(7):863-867). Регуляция пути передачи сигнала HGF/c-Мet вовлечена в прогрессирование роста опухолей и развитие метастазов (см, например, Trusolino & Comoglio, Nature Rev. (2002), 2:289-300). HGF связывается с внеклеточным доменом тирозинкиназного рецептора Met (RTK) и регулирует различные биологические процессы, такие как рассеяние, пролиферация и выживание клеток. Передача сигнала HGF-Met, в основном, происходит при нормальном развитии эмбриона, а в частности при миграции мышечных клеток-предшественников и развитии печени и нервной системы (Bladt et al., 1995; Hamanoue et al., 1996; Maina et al., 1996; Schmidt et al., 1995; Uehara et al., 1995)). Эволюционные фенотипы Met- и HGF-дефицитных мышей имеют очень большое сходство, что позволяет предположить, что HGF является узнаваемым лигандом для рецептора Met (Schmidt et al., 1995; Uehara et al., 1995). HGF-Met также играет определенную роль в регенерации печени, в ангиогенезе и в заживлении ран (Bussolino et al., 1992; Matsumoto & Nakamura, 1993; Nusrat et al., 1994). Предшественник рецептора Met подвергается протеолитическому расщеплению с образованием внеклеточной субъединицы Было показано, что Met трансформируется в обработанную канцерогеном клеточную линию остеосаркомы (Cooper et al., 1984; Park et al., 1986). Сверхэкспрессия или амплификация гена Met наблюдается при различных раковых заболеваниях человека. Так, например, при раке ободочной кишки наблюдается, по меньшей мере, 5-кратная сверхэкспрессия белка Met, причем, сообщалось, что при метастазах в печени происходит амплификация его гена (Di Renzo et al., 1995; Liu et al., 1992). Также сообщалось, что белок Met сверхэкспрессируется при плоскоклеточной карциноме ротовой полости, гепатоцеллюлярной карциноме, почечно-клеточной карциноме, карциноме молочной железы и карциноме легких (Jin et al., 1997; Morello et al., 2001; Natali et al., 1996; Olivero et al., 1996; Suzuki et al., 1994). Кроме того, сверхэкспрессия мРНК наблюдается в гепатоцеллюлярной карциноме, карциноме желудка и карциноме ободочной кишки (Boix et al., 1994; Kuniyasu et al., 1993; Liu et al., 1992). При почечной папиллярной карциноме в киназном домене Met был обнаружен ряд мутаций, которые вызывают активацию конститутивного рецептора (Olivero et al. 1999; Schmidt et al., 1997; Schmidt et al., 1999). Эти активирующие мутации приводят к конститутивному фосфорилированию тирозина Met, активации МАРК, образованию патологических очагов и к онкогенезу (Jeffers et al., 1997). Кроме того, эти мутации усиливают подвижность и инвазию клеток (Giordano et al., 2000, Lorenzato et al., 2002). HGF-зависимая активация Met в трансформированных клетках опосредует увеличение подвижности, рассеяния и миграции клеток, что, в конечном счете, приводит к инвазивному росту опухоли и к развитию метастазов (Jeffers et al., 1996; Meiners et al., 1998). Было показано, что Met взаимодействует с другими белками, которые стимулируют активацию, трансформацию и инвазию рецептора. Сообщалось, что в неопластических клетках, Met взаимодействует с интегрином Met является членом подсемейства RТК, которое включает Ron и Sea (Maulik et al., 2002). Предсказание структуры внеклеточного домена Met позволяет предположить о наличии общей гомологии Met с семафоринами и плексинами. N-конец Met содержит домен Sema, состоящий приблизительно из 500 аминокислот, которые являются консервативными для всех семафоринов и плексинов. Семафорины и плексины принадлежат к большому семейству секретируемых и мембраносвязанных белков, впервые описанных с точки зрения их роли в развитии нервных тканей (Van Vactor & Lorenz, 1999). Однако совсем недавно было обнаружено, что сверхэкспрессия семафорина коррелирует с инвазией опухоли и с метастазами. Богатый цистеином домен PSI (также называемый доменом Met-родственной последовательности), обнаруженный в плексинах, семафоринах и интегринах, расположен рядом с доменом Sеma, за которым следуют четыре повтора IРТ, которые представляют собой иммуноглобулин-подобные области, присутствующие в плексинах и в факторах транскрипции. Последние исследования позволяют предположить, что домен Sеma Met является достаточным для связывания HGF с гепарином (Gherardi et al., 2003). Кроме того, в публикации Kong-Beltran et al. (Cancer Cell (2004), 6:61-73) сообщалось, что домен Sеma Met является необходимым для димеризации и активации рецептора. Были описаны многочисленные молекулы, нацеленные на HGF/c-met-путь передачи сигнала. Такими молекулами являются антитела, такие как антитела, описанные в патенте США Все цитируемые здесь работы, включая патентные заявки и публикации, во всей своей полноте вводятся в настоящее описание посредством ссылки. Описание изобретения Настоящее изобретение частично основано на идентификации ряда антагонистов биологического пути HGF/c-met, который представляет собой биологический/клеточный процесс, являющийся важной и преимущественной терапевтической задачей. Настоящее изобретение относится к композициям и к способам, основанным на подавлении активации HGF/c-met, включая, но не ограничиваясь ими, подавление связывания HGF с внеклеточной частью c-met и мультимеризацию рецептора. Описанные здесь антагонисты представляют собой важные терапевтические и диагностические средства, которые могут быть использованы для лечения патологических состояний, ассоциированных с аномальной или нежелательной передачей сигнала по пути HGF/c-met. В соответствии с этим настоящее изобретение относится к способам, к композициям, к наборам и к промышленным изделиям, предназначенным для модуляции пути HGF/c-met, включая модуляцию связывания с лигандом c-met, димеризацию c-met, активацию и другие биологические/физиологические активности, ассоциированные с передачей сигнала HGF/c-met. В одном из своих аспектов настоящее изобретение относится к терапевтическим средствам против HGF/c-met, подходящим для терапевтического применения и способным в различной степени нарушать пути передачи сигнала HGF/c-met. Так, например, в одном из своих вариантов настоящее изобретение относится к гуманизированному антителу против c-met, где аффинность указанного моновалентного антитела против человеческого c-met (например, аффинность Fab-фрагмента этого антитела против человеческого c-met), по существу, аналогична аффинности моновалентного мышиного антитела (например, аффинности Fab-фрагмента мышиного антитела против человеческого c-met), включающего последовательность вариабельного домена легкой цепи и тяжелой цепи, представленной на фиг.7 (SEQ ID NO:9 и 10), или состоящего, или по существу состоящего, из указанной последовательности. В другом своем варианте настоящее изобретение относится к гуманизированному антителу против c-met, где аффинность моновалентного антитела против человеческого c-met (например, аффинность Fab-фрагмента антитела против человеческого c-met) ниже, например, по меньшей мере, в 3, 5, 7 или 10 раз, чем аффинность моновалентного мышиного антитела (например, аффинность Fab-фрагмента мышиного антитела против человеческого c-met), включающего последовательность вариабельного домена легкой цепи и тяжелой цепи, представленной на фиг.7 (SEQ ID NO:9 и 10), или состоящего, или по существу, состоящего из указанной последовательности. В другом своем варианте настоящее изобретение относится к гуманизированному антителу против c-met, где аффинность моновалентного антитела против человеческого c-met (например, аффинность Fab-фрагмента антитела против человеческого c-met) является более высокой, например, по меньшей мере, в 3, 5, 7, 10 или 13 раз, чем аффинность моновалентного мышиного антитела (например, аффинность Fab-фрагмента мышиного антитела против человеческого c-met), включающего последовательность вариабельного домена легкой цепи и тяжелой цепи, представленной на фиг.7 (SEQ ID NO:9 и 10), или состоящего, или по существу, состоящего из указанной последовательности. В одном из вариантов изобретения аффинность моновалентного мышиного антитела против человеческого c-met, по существу, аналогична аффинности связывания Fab-фрагмента, содержащего последовательности вариабельного домена антитела, продуцируемого гибридомной клеточной линией, депонированной в Американской коллекции типовых культур рег. В одном из аспектов настоящего изобретения антагонист HGF/c-met согласно изобретению включает антитело против c-met, содержащее: (а) по меньшей мере, одну, две, три, четыре или пять последовательностей гипервариабельной области (HVR), выбранных из группы, состоящей из: (i) HVR-L1, включающей последовательность А1-А17, где А1-А17 представляет собой KSSQSLLYTSSQKNYLA (SEQ ID NO:1) (ii) HVR-L2, включающей последовательность В1-В7, где В1-В7 представляет собой WASTRES (SEQ ID NO:2) (iii) HVR-L3, включающей последовательность С1-С9, где С1-С9 представляет собой QQYYAYPWT (SEQ ID NO:3) (iv) HVR-Н1, включающей последовательность D1-D10, где D1-D10 представляет собой GYTFTSYWLH (SEQ ID NO:4) (v) HVR-H2, включающей последовательность E1-E18, где E1-E18 представляет собой GMIDPSNSDTRFNPNFKD (SEQ ID NO:5) и (vi) HVR-H3, включающей последовательность F1-F11, где F1-F11 представляет собой XYGSYVSPLDY (SEQ ID NO:6), а Х не является R; и (b) по меньшей мере, один вариант HVR, где указанный вариант последовательности HVR имеет модификацию, по меньшей мере, в одном остатке последовательности, представленной в SEQ ID NO:1, 2, 3, 4, 5 или 6. В одном из вариантов изобретения HVR-L1 антитела согласно изобретению содержит последовательность SEQ ID NO:1. В одном из вариантов изобретения HVR-L2 антитела согласно изобретению содержит последовательность SEQ ID NO:2. В одном из вариантов изобретения HVR-L3 антитела согласно изобретению содержит последовательность SEQ ID NO:3. В одном из вариантов изобретения HVR-Н1 антитела согласно изобретению содержит последовательность SEQ ID NO:4. В одном из вариантов изобретения HVR-Н2 антитела согласно изобретению содержит последовательность SEQ ID NO:5. В одном из вариантов изобретения HVR-Н3 антитела согласно изобретению содержит последовательность SEQ ID NO:6. В одном из вариантов изобретения HVR-Н3 содержит последовательность TYGSYVSPLDY (SEQ ID NO:7). В одном из вариантов изобретения HVR-Н3 содержит последовательность SYGSYVSPLDY (SEQ ID NO:8). В одном из вариантов изобретения антитело согласно изобретению, содержащее эти последовательности (в описанной здесь комбинации), является гуманизированным или человеческим. В одном из своих аспектов настоящее изобретение относится к антителу, содержащему одну, две, три, четыре, пять или шесть областей HVR, где каждая область HVR содержит последовательность, или состоит, или, по существу, состоит из последовательности, которая выбрана из группы, состоящей из SEQ ID NO:1, 2, 3, 4, 5, 6, 7 и 8, и где SEQ ID NO:1 соответствует HVR-L1, SEQ ID NO:2 соответствует HVR-L2, SEQ ID NO:3 соответствует HVR-L3, SEQ ID NO:4 соответствует HVR-Н1, SEQ ID NO:5 соответствует HVR-Н2, а SEQ ID NO:6, 7 или 8 соответствуют HVR-Н3. В одном из вариантов изобретения антитело согласно изобретению содержит HVR-L1, HVR-L2, HVR-L3, HVR-Н1, HVR-Н2 и HVR-Н3, где каждая из этих областей содержит SEQ ID NO:1, 2, 3, 4, 5 и 7, в соответствующем порядке. В одном из вариантов изобретения антитело согласно изобретению содержит HVR-L1, HVR-L2, HVR-L3, HVR-Н1, HVR-Н2 и HVR-Н3, где каждая из этих областей содержит SEQ ID NO:1, 2, 3, 4, 5 и 8 в соответствующем порядке. Варианты HVR в антителе согласно изобретению могут иметь модификации в одном или нескольких остатках HVR. В одном из вариантов изобретения вариант HVR-L2 содержит 1-5 (1, 2, 3, 4 или 5) замен в любой комбинации в следующих положениях: В1 (М или L), В2 (P, T, G или S), B3 (N, G, R или Т), В4 (I, N или F), В5 (P, I, L или G), В6 (A, D, T или V) и В7 (R, I, M или G). В одном из вариантов изобретения вариант HVR-Н1 содержит 1-5 (1, 2, 3, 4 или 5) замен в любой комбинации в следующих положениях: D3 (N, P, L, S, A, I), D5 (I, S или Y), D6 (G, D, T, K, R), D7 (F, H, R, S, T или V) и D9 (М или V). В одном из вариантов изобретения вариант HVR-Н2 содержит 1-4 (1, 2, 3 или 4) замены в любой комбинации в следующих положениях: Е7 (Y), Е9 (I), Е10 (I), Е14 (T или Q), E15 (D, K, S, T или V), E16 (L), E17 (E, H, N или D) и Е18 (Y, E или Н). В одном из вариантов изобретения вариант HVR-Н3 содержит 1-5 (1, 2, 3, 4 или 5) замен в любой комбинации в следующих положениях: F1 (T, S), F3 (R, S, H, T, A, K), F4 (G), F6 (R, F, M, T, E, K, A, L, W), F7 (L, I, T, R, K, V), F8 (S, A), F10 (Y, N) и F11 (Q, S, H, F). Буква(ы) в скобках после каждого положения означает(ют) репрезентативную замену (т.е. модификацию) аминокислоты, причем для специалиста в данной области очевидно, что эффективность замен одних аминокислот другими аминокислотами, в соответствии с настоящим изобретением, может быть оценена рутинными методами, известными специалистам и/или описанными в настоящей заявке. В одном из вариантов изобретения HVR-L1 имеет последовательность SEQ ID NO:1. В одном из вариантов изобретения в положении F1 в варианте HVR-Н3 присутствует Т. В одном из вариантов изобретения в положении F1 в варианте HVR-Н3 присутствует S. В одном из вариантов изобретения в положении F3 в варианте HVR-Н3 присутствует R. В одном из вариантов изобретения в положении F3 в варианте HVR-Н3 присутствует S. В одном из вариантов изобретения в положении F7 в варианте HVR-Н3 присутствует Т. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-Н3, где в положении F1 присутствует T или S, в положении F3 присутствует R или S, а в положении F7 присутствует Т. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-Н3, где в положении F1 присутствует Т, в положении F3 присутствует R, а в положении F7 присутствует Т. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-Н3, где в положении F1 присутствует S. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-Н3, где в положении F1 присутствует Т, а в положении F3 присутствует R. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-Н3, где в положении F1 присутствует S, в положении F3 присутствует R, а в положении F7 присутствует Т. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-Н3, где в положении F1 присутствует Т, в положении F3 присутствует S, в положении F7 присутствует Т, а в положении F8 присутствует S. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-Н3, где в положении F1 присутствует Т, в положении F3 присутствует S, в положении F7 присутствует Т, а в положении F8 присутствует А. В некоторых вариантах изобретения указанный вариант антитела HVR-Н3 дополнительно содержит HVR-L1, HVR-L2, HVR-L3, HVR-Н1 и HVR-Н2, где каждая из этих областей содержит, в соответствующем порядке, последовательность, представленную в SEQ ID NO:1, 2, 3, 4 и 5. В некоторых вариантах изобретения такие антитела дополнительно включают консенсусную последовательность человеческой каркасной области тяжелой цепи подгруппы III. В одном из вариантов этих антител указанная консенсусная последовательность каркасной области имеет замены в положениях 71, 73 и/или 78. В некоторых вариантах этих антител в положении 71 присутствует А, в положении 73 присутствует Т и/или в положении 78 присутствует А. В одном из вариантов указанных антител эти антитела дополнительно включают консенсусную последовательность человеческой каркасной области легкой цепи кI. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-H2, где в положении В6 присутствует V. В некоторых вариантах изобретения указанный вариант HVR-L2-антитела дополнительно включает HVR-L1, HVR-L3, HVR-Н1, HVR-Н2 и HVR-Н3, где каждая из этих областей содержит, в соответствующем порядке, последовательность, представленную в SEQ ID NO:1, 3, 4, 5 и 6. В некоторых вариантах изобретения указанный вариант HVR-L2-антитела дополнительно включает HVR-L1, HVR-L3, HVR-Н1, HVR-Н2 и HVR-Н3, где каждая из этих областей содержит, в соответствующем порядке, последовательность, представленную в SEQ ID NO:1, 3, 4, 5 и 7. В некоторых вариантах изобретения указанный вариант HVR-L2-антитела дополнительно включает HVR-L1, HVR-L3, HVR-Н1, HVR-Н2 и HVR-Н3, где каждая из этих областей содержит, в соответствующем порядке, последовательность, представленную в SEQ ID NO:1, 3, 4, 5 и 8. В некоторых вариантах изобретения такие антитела дополнительно включают консенсусную последовательность человеческой каркасной области тяжелой цепи подгруппы III. В одном из вариантов этих антител указанная консенсусная последовательность каркасной области имеет замены в положениях 71, 73 и/или 78. В некоторых вариантах этих антител в положении 71 присутствует А, в положении 73 присутствует Т и/или в положении 78 присутствует А. В одном из вариантов этих антител указанные антитела дополнительно включают консенсусную последовательность человеческой каркасной области легкой цепи В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-L2, где в положении Е14 присутствует Т, в положении Е15 присутствует К, а в положении Е17 присутствует Е. В одном из вариантов изобретения антитело согласно изобретению содержит вариант HVR-Н2, где в положении Е17 присутствует Е. В некоторых вариантах изобретения указанный HVR-Н3-вариант антитела дополнительно включает HVR-L1, HVR-L2, HVR-L3, HVR-Н1 и HVR-Н3, где каждая из этих областей содержит, в соответствующем порядке, последовательность, представленную в SEQ ID NO:1, 2, 3, 4 и 6. В некоторых вариантах изобретения указанный вариант антитела HVR-Н2 дополнительно включает HVR-L1, HVR-L2, HVR-L3, HVR-Н1 и HVR-Н3, где каждая из этих областей содержит, в соответствующем порядке, последовательность, представленную в SEQ ID NO:1, 2, 3, 4 и 7. В некоторых вариантах изобретения указанный HVR-Н2-вариант антитела дополнительно включает HVR-L1, HVR-L2, HVR-L3, HVR-Н1 и HVR-Н3, где каждая из этих областей содержит, в соответствующем порядке, последовательность, представленную в SEQ ID NO:1, 2, 3, 4 и 8. В некоторых вариантах изобретения такие антитела дополнительно включают консенсусную последовательность человеческой каркасной области тяжелой цепи подгруппы III. В одном из вариантов этих антител указанная консенсусная последовательность каркасной области имеет замены в положениях 71, 73 и/или 78. В некоторых вариантах этих антител в положении 71 присутствует А, в положении 73 присутствует Т и/или в положении 78 присутствует А. В одном из вариантов этих антител указанные антитела дополнительно включают консенсусную последовательность человеческой каркасной области легкой цепи В одном из своих аспектов настоящее изобретение относится к антителу, содержащему одну, две, три, четыре, пять или все последовательности HVR, представленные на фиг. 2, 3 и/или 4 (SEQ ID NO:56-163). Терапевтическое средство, используемое для введения индивидууму-хозяину, предпочтительно, вырабатывает незначительный иммунный ответ или вообще не вырабатывает иммунного ответа против указанного агента у данного индивидуума. В одном из своих вариантов настоящее изобретение относится к указанному агенту. Так, например, в одном из своих вариантов настоящее изобретение относится к гуманизированному антителу, которое вырабатывает и/или, предположительно, вырабатывает у индивидуума-хозяина иммунный ответ в виде продуцирования человеческого антимышиного антитела (НАМА), на уровне, который, по существу, значительно ниже уровня продуцирования антитела, содержащего последовательности SEQ ID NO:9 и 10. В другом примере настоящее изобретение относится к гуманизированному антителу, которое вырабатывает и/или, предположительно, вырабатывает минимальный иммунный ответ в виде продуцирования человеческого антимышиного антитела (НАМА), или вообще не вырабатывает такого ответа. В одном из примеров антитело согласно изобретению вырабатывает иммунный ответ в виде продуцирования антимышиного антитела на клинически приемлемом уровне или на более низком уровне. Гуманизированное антитело согласно изобретению может содержать одну или несколько последовательностей человеческой и/или человеческой консенсусной последовательности негипервариабельной области (например, каркасной области) в вариабельном домене тяжелой цепи и/или легкой цепи. В некоторых вариантах изобретения в последовательностях человеческой и/или человеческой консенсусной последовательности негипервариабельной области присутствует одна или несколько дополнительных модификаций. В одном из вариантов изобретения вариабельный домен тяжелой цепи антитела согласно изобретению содержит человеческую консенсусную каркасную последовательность, которая, в одном из вариантов изобретения, представляет собой консенсусную каркасную последовательность подгруппы III. В одном из вариантов изобретения антитело согласно изобретению содержит консенсусную каркасную последовательность подгруппы III, модифицированную, по меньшей мере, в одном положении аминокислоты. Так, например, в одном из вариантов изобретения модифицированная консенсусная каркасная последовательность подгруппы III может содержать замену в одном или нескольких из положений 71, 73 и/или 78. В одном из вариантов изобретения указанными заменами являются R71А, N73Т и/или N78А в любой их комбинации. Как известно специалистам, и как более подробно описано ниже, положение/окружение аминокислот, определяющих гипервариабельную область данного антитела, может варьироваться в зависимости от ситуации и различных определений, известных специалистам (как описано ниже). Некоторые положения в вариабельном домене могут рассматриваться как пограничные положения гипервариабельной области, которые, в соответствии с одним из ряда критериев, могут, предположительно, находиться в гипервариабельной области, а в соответствии с другим из ряда критериев они могут, предположительно, находиться за пределами гипервариабельной области. Одно или несколько из этих положений могут также присутствовать в удлиненных гипервариабельных областях (подробно определенных ниже). Настоящее изобретение относится к антителам, содержащим модификации в этих гибридных гипервариабельных положениях. В одном из вариантов изобретения эти гибридные гипервариабельные положения включают одно или несколько из положений 26-30, 33-35В, 47-49, 57-65, 93, 94 и 102 в вариабельном домене тяжелой цепи. В одном из вариантов изобретения эти гибридные гипервариабельные положения включают одно или несколько из положений 24-29, 35-36, 46-49, 56 и 97 в вариабельном домене легкой цепи. В одном из вариантов изобретения антитело согласно изобретению содержит вариант консенсусной каркасной последовательности человеческой цепи подгруппы III, модифицированный в одном или нескольких гибридных гипервариабельных положениях. В одном из вариантов изобретения антитело согласно изобретению содержит вариабельный домен тяжелой цепи, включающий вариант консенсусной каркасной последовательности человеческой цепи подгруппы III, модифицированный в одном или нескольких из положений 27-28, 30, 33-35, 49, 57-65, 94 и 102. В одном из вариантов изобретения указанное антитело имеет замену F27Y. В одном из вариантов изобретения указанное антитело имеет замену Т28N, P, L, S, A или I. В одном из вариантов изобретения указанное антитело имеет замену S30I, T или Y. В одном из вариантов изобретения указанное антитело имеет замену А33W. В одном из вариантов изобретения указанное антитело имеет замену М34L или M34V. В одном из вариантов изобретения указанное антитело имеет замену S35Н. В одном из вариантов изобретения указанное антитело имеет замену T57I. В одном из вариантов изобретения указанное антитело имеет замену Y58R. В одном из вариантов изобретения указанное антитело имеет замену Y59F. В одном из вариантов изобретения указанное антитело имеет замену A60N. В одном из вариантов изобретения указанное антитело имеет замену D61P, T или Q. В одном из вариантов изобретения указанное антитело имеет замену S62N, D, K, T или V. В одном из вариантов изобретения указанное антитело имеет замену V63F или V63L. В одном из вариантов изобретения указанное антитело имеет замену К64Е, H, N, D или Q. В одном из вариантов изобретения указанное антитело имеет замену G65D, Y, E или Н. В одном из вариантов изобретения указанное антитело имеет замену R94T или R94S. В одном из вариантов изобретения указанное антитело имеет замену Y102Q, S, H или F. В одном из вариантов изобретения указанное антитело согласно изобретению имеет указанную модификацию R94T или R94S, а также имеет одну или несколько модификаций в положении 96 и/или 100. В одном из вариантов изобретения указанными модификациями являются замены G96R и/или S100T (т.е. в HVR-Н3). В одном из вариантов изобретения указанное антитело согласно изобретению содержит вариабельный домен легкой цепи, включающий вариант консенсусной каркасной последовательности человеческой цепи каппа подгруппы I, модифицированный в одном или нескольких из положений 24, 25, 29 и 56. В одном из вариантов изобретения указанное антитело имеет замену R24K. В одном из вариантов изобретения указанное антитело имеет замену А25S. В одном из вариантов изобретения указанное антитело имеет замену I29Q. В одном из вариантов изобретения указанное антитело имеет замены S56R, I, M или G. В одном из вариантов изобретения указанное антитело согласно изобретению содержит вариабельный домен тяжелой цепи, включающий вариант человеческой консенсусной каркасной последовательности цепи подгруппы III, модифицированный в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 или во всех положениях 27-28, 30, 33-35, 49, 57-65, 94 и 102. В одном из вариантов изобретения указанную модификацию выбирают из группы, состоящей из F27Y, T28 (N, P, L, S, A или I), S30 (I, T или Y), A33W, M34 (L,V), S35H, T57I, Y58R, Y59F, A60N, D61 (P,T,Q), S62 (N, D, K, T, V), V63 (F,L), K64(E, H, N, D, Q), G65(D, Y, E, H), R94(T,S) и Y102(Q, S, H, F). В одном из вариантов изобретения антитело согласно изобретению, содержащее указанные модификации R94T или R94S, дополнительно содержит одну или несколько модификаций в положении 96 и/или 100. В одном из вариантов изобретения указанными модификациями являются замены G96R и/или S100T (т.е. в HVR-Н3). В одном из вариантов изобретения указанное антитело согласно изобретению содержит вариабельный домен легкой цепи, включающий вариант консенсусной каркасной последовательности человеческой цепи каппа подгруппы I, модифицированной в 1, 2, 3 или во всех положениях 24, 25, 29 и 56. В одном из вариантов изобретения модификация выбрана из группы, состоящей из R24K, A25S, I29Q и S56(R, I, M, G). Антитело согласно изобретению может содержать любые подходящие человеческие или консенсусные каркасные человеческие последовательности легкой цепи при условии, что такое антитело обладает нужными биологическими свойствами (например, нужной аффинностью связывания). В одном из вариантов изобретения антитело согласно изобретению содержит, по меньшей мере, часть каркасной последовательности (или всю каркасную последовательность) человеческой легкой цепи В одном из вариантов изобретения антитело согласно изобретению содержит вариабельный домен тяжелой и/или легкой цепи, включающий каркасную последовательность, представленную в SEQ ID NO:13 и/или 16 (фиг.1), при условии, что в положении 94 тяжелой цепи отсутствует R (а предпочтительно, но необязательно, присутствует S или Т). В одном из аспектов изобретения антитело согласно изобретению представляет собой гуманизированное антитело против c-met, которое ингибирует связывание человеческого фактора роста гепатоцитов с его рецептором более эффективно, чем эталонное антитело, включая химерное антитело против c-met, содержащее вариабельную последовательность легкой цепи и тяжелой цепи, представленную на фиг.7 (SEQ ID NO:9 и 10). Так, например, в одном из вариантов изобретения антитело согласно изобретению ингибирует связывание HGF при концентрации IC50, которая составляет менее примерно половины значения IC50 химерного антитела. В одном из вариантов изобретения величина IC50 антитела согласно изобретению составляет примерно 0,1; 0,2; 0,3 или 0,4 от величины IC50 химерного антитела. Сравнение эффективностей ингибирования связывания HGF с его рецептором может быть осуществлено различными методами, известными специалистам, включая методы, описанные ниже в примерах. В одном из вариантов изобретения величины IC50 определяют в диапазоне концентраций примерно от 0,01 нМ до 1000 нМ. В одном из аспектов изобретения антитело согласно изобретению представляет собой гуманизированное антитело против c-met, которое ингибирует активацию рецептора человеческого фактора роста гепатоцитов (HGF) более эффективно, чем эталонное антитело, включая химерное антитело против c-met, содержащее вариабельную последовательность легкой цепи и тяжелой цепи, представленную на фиг.7 (SEQ ID NO:9 и 10). Так, например, в одном из вариантов изобретения антитело согласно изобретению ингибирует активацию рецептора при концентрации IC50, которая составляет менее примерно половины значения IC50 химерного антитела. В одном из вариантов изобретения величина IC50 антитела согласно изобретению составляет примерно 0,1; 0,2; 0,3 или 0,4 от величины IC50 химерного антитела. Сравнение эффективностей ингибирования активации рецептора HGF может быть осуществлено различными методами, известными специалистам, включая методы, описанные ниже в примерах. В одном из вариантов изобретения величины IC50 определяют в диапазоне концентраций примерно от 0,1 до 100 нМ. В одном из аспектов изобретения антитело согласно изобретению представляет собой гуманизированное антитело против c-met, которое ингибирует c-met-зависимую пролиферацию клеток более эффективно, чем эталонное антитело, включая химерное антитело против c-met, содержащее вариабельную последовательность легкой цепи и тяжелой цепи, представленную на фиг.7 (SEQ ID NO:9 и 10). Так, например, в одном из вариантов изобретения антитело согласно изобретению ингибирует пролиферацию клеток при концентрации IC50, которая составляет менее примерно половины значения IC50 химерного антитела. В одном из вариантов изобретения величина IC50 антитела согласно изобретению составляет примерно 0,1; 0,2; 0,3 или 0,4 от величины IC50 химерного антитела. Сравнение эффективностей ингибирования пролиферации клеток может быть осуществлено различными методами, известными специалистам, включая методы, описанные ниже в примерах. В одном из вариантов изобретения величины IC50 определяют в диапазоне концентраций примерно от 0,1 до 100 нМ. В одном из вариантов изобретения гуманизированное антитело и химерное антитело являются моновалентными. В одном из вариантов изобретения гуманизированное антитело и химерное антитело содержат одну Fab-область, присоединенную к Fc-области. В одном из вариантов изобретения эталонное химерное антитело содержит последовательности вариабельных доменов, представленные на фиг.7 (SEQ ID NO:9 и 10), которые присоединены к человеческой Fc-области. В одном из вариантов изобретения человеческой Fc-областью является область IgG (например, IgG1, 2, 3 или 4). В одном из своих аспектов настоящее изобретение относится к антителу, содержащему вариабельный домен тяжелой цепи, включающий последовательности HVR1-НС, HVR2-НС и/или HVR3-НС, представленные на фиг.13. В одном из вариантов изобретения вариабельный домен включает последовательности FR1-НС, FR2-НС, FR3-НС и/или FR4-НС, представленные на фиг.13. В одном из вариантов изобретения указанное антитело содержит последовательности СН1 и/или Fc, представленные на фиг.13. В одном из вариантов изобретения антитело согласно изобретению содержит вариабельный домен тяжелой цепи, включающий последовательности HVR1-НС, HVR2-НС и/или HVR3-НС и последовательности FR1-НС, FR2-НС, FR3-НС и/или FR4-НС, представленные на фиг.13. В одном из вариантов изобретения антитело согласно изобретению содержит вариабельный домен тяжелой цепи, включающий последовательности HVR1-НС, HVR2-НС и/или HVR3-НС и последовательности СН1 и/или Fc, представленные на фиг.13. В одном из вариантов изобретения антитело согласно изобретению содержит вариабельный домен тяжелой цепи, включающий последовательности HVR1-HC, HVR2-HC и/или HVR3-HC; последовательности FR1-HC, FR2-HC, FR3-HC и/или FR4-HC, представленные на фиг.13; и последовательности СН1 и/или Fc, представленные на фиг.13. В одном из вариантов изобретения Fc-область антитела согласно изобретению содержит комплекс, состоящий из полипептида, содержащего последовательность Fc, представленную на фиг.13, и полипептида, содержащего последовательность Fc, представленную на фиг.14. В одном из своих аспектов настоящее изобретение относится к антителу, содержащему вариабельный домен легкой цепи, включающий последовательности HVR1-LС, HVR2-LС и/или HVR3-LС, представленные на фиг.13. В одном из вариантов изобретения указанный вариабельный домен включает последовательности FR1-LС, FR2-LС, FR3-LС и/или FR4-LС, представленные на фиг.13. В одном из вариантов изобретения указанное антитело содержит последовательность CL1, представленную на фиг.13. В одном из вариантов изобретения антитело согласно изобретению содержит вариабельные домены легкой и тяжелой цепей, описанные в предыдущих двух параграфах. В одном из вариантов изобретения указанное антитело является моновалентным и содержит Fc-область. В одном из вариантов изобретения Fc-область содержит, по меньшей мере, один Антагонисты согласно одному или нескольким аспектам изобретения могут быть использованы для модуляции HGF/c-met-ассоциированных эффектов, включая, но не ограничиваясь ими, активацию с-met, последующую передачу молекулярного сигнала (например, активированное митогеном фосфорилирование протеинкиназы (МАРК)), пролиферацию клеток, миграцию клеток, выживание клеток, морфогенез клеток и ангиогенез. Эти эффекты могут модулироваться по любому биологически релевантному механизму, включая дизрупцию лиганда (например, HGF), связывающегося с c-met, фосфорилирование c-met и/или мультимеризацию c-met. В соответствии с этим в одном из своих вариантов настоящее изобретение относится к антителу-антагонисту против с-met, которое ингибирует связывание HGF с c-met. В одном из вариантов изобретения антитело-антагонист с-met согласно изобретению ингибирует мультимеризацию с-met (например, димеризацию c-met). В одном из вариантов изобретения антитело-антагонист c-met согласно изобретению ингибирует димеризацию домена Sema c-met. В одном из примеров антитело-антагонист c-met негативно влияет на способность домена Sema c-met осуществлять димеризацию c-met. Такое негативное влияние может быть прямым или опосредованным. Так, например, антитело-антагонист c-met может связываться с последовательностью в домене Sеma c-met и, тем самым, ингибировать взаимодействие указанного связанного домена с его партнером по связыванию (таким как другая молекула c-met). В другом примере антитело-антагонист c-met может связываться с последовательностью, которая не присутствует в домене Sеma c-met, но при этом такое связывание приводит к нарушению способности домена Sеma c-met взаимодействовать с его партнером по связыванию (таким как другая молекула c-met). В одном из вариантов изобретения антитело-антагонист согласно изобретению связывается с c-met (например, с внеклеточным доменом), в результате чего нарушается димеризация c-met. В одном из вариантов изобретения антитело-антагонист согласно изобретению связывается с c-met, в результате чего нарушается способность домена Sеma c-met осуществлять димеризацию c-met. Так, например, в одном из вариантов изобретения настоящее изобретение относится к антителу-антагонисту, которое, после его связывания с молекулой c-met, ингибирует димеризацию указанной молекулы. В одном из вариантов изобретения антитело-антагонист c-met согласно изобретению специфически связывается с последовательностью в домене Sеma c-met. В одном из вариантов изобретения антитело-антагонист согласно изобретению ингибирует димеризацию c-met, включая его гомодимеризацию. В одном из вариантов изобретения антитело-антагонист согласно изобретению ингибирует димеризацию c-met, включая его гетеродимеризацию (то есть димеризацию c-met с молекулой, не являющейся c-met). В некоторых случаях может оказаться предпочтительным получение антитела-антагониста c-met, которое не препятствует связыванию лиганда (такого как HGF) с c-met. В соответствии с этим в одном из своих вариантов настоящее изобретение относится к антителу, которое не связывается с HGF-связывающим сайтом на c-met. В другом варианте изобретения антитело согласно изобретению, по существу, не ингибирует связывание HGF с c-met. В одном из вариантов изобретения антитело согласно изобретению, по существу, не конкурирует с HGF за связывание с c-met. В одном из примеров антитело-антагонист согласно изобретению может быть использовано в комбинации с одним или несколькими другими антагонистами, где указанные антагонисты направлены на подавление различных процессов и/или функций в пути передачи сигнала HGF/c-met. Так, например, в одном из вариантов изобретения антитело-антагонист c-met согласно изобретению связывается с эпитопом на c-met, отличающимся от эпитопа, связывающегося с другим антагонистом c-met (таким как Fab-фрагмент моноклонального антитела, продуцируемого гибридомной клеточной линией, депонированной в Американской коллекции типовых культур рег. В одном из своих вариантов настоящее изобретение относится к антителу-антагонисту c-met, которое ингибирует мультимеризацию c-met и связывание с лигандом. Так, например, антитело-антагонист согласно изобретению, которое ингибирует мультимеризацию c-met (например, димеризацию), может, кроме того, обладать способностью конкурировать с HGF за связывание с c-met. В одном из вариантов антитела-антагониста c-met согласно изобретению связывание указанного антагониста с c-met приводит к ингибированию активации c-met под действием HGF. В другом варианте антитела-антагониста c-met согласно изобретению связывание указанного антагониста с c-met в клетке приводит к ингибированию пролиферации, выживания, рассеяния, морфогенеза и/или подвижности этой клетки. В одном из вариантов изобретения антитело-антагонист c-met согласно изобретению специфически связывается, по меньшей мере, с частью домена Sеma c-met или его варианта. В одном из примеров антитело-антагонист согласно изобретению специфически связывается, по меньшей мере, с одной из последовательностей, выбранной из группы, состоящей из LDAQT (SEQ ID NO:15)(например, остатков 269-273 с-met), LTEKRKKRS (SEQ ID NO:16)(например, остатков 300-308 c-met), KPDSAEPM (SEQ ID NO:17)(например, остатков 350-357 c-met) и NVRCLQHF (SEQ ID NO:18)(например, остатков 381-388 с-met). В одном из вариантов изобретения антитело-антагонист согласно изобретению специфически связывается с конформационным эпитопом, образованным частью или всей, по меньшей мере, из одной последовательностью, выбранной из группы, состоящей из LDAQT (например, остатков 269-273 с-met), LTEKRKKRS (например, остатков 300-308 c-met), KPDSAEPM (например, остатков 350-357 c-met) и NVRCLQHF (например, остатков 381-388 с-met). В одном из вариантов изобретения антитело-антагонист согласно изобретению специфически связывается с аминокислотной последовательностью, которая, по меньшей мере, на 50, 60, 70, 80, 90, 95 или 98% идентична или аналогична последовательностям LDAQT, LTEKRKKRS, KPDSAEPM и/или NVRCLQHF. В одном из вариантов изобретения антитело согласно изобретению специфически связывается с рецептором HGF животного первого вида и не связывается специфически с рецептором HGF животного второго вида. В одном из вариантов изобретения указанным животным первого вида является человек и/или примат (например, собакоподобная обезьяна), а указанным животным второго вида является животное семейства мышиных (например, мышь), и/или животное семейства собачьих. В одном из вариантов изобретения указанным животным первого вида является человек. В одном из вариантов изобретения указанным животным первого вида является примат, например, собакоподобная обезьяна. В одном из вариантов изобретения указанным животным второго вида является животное семейства мышиных, например, мышь. В одном из вариантов изобретения указанным животным второго вида является животное семейства собачьих. В одном из своих аспектов настоящее изобретение относится к композициям, содержащим одно или несколько антител-антагонистов согласно изобретению и носитель. В одном из вариантов изобретения указанный носитель является фармацевтически приемлемым. В одном из своих аспектов настоящее изобретение относится к нуклеиновым кислотам, кодирующим антитело-антагонист против c-met согласно изобретению. В одном из своих аспектов настоящее изобретение относится к векторам, содержащим нуклеиновую кислоту согласно изобретению. В одном из вариантов изобретения настоящее изобретение относится к клеткам-хозяевам, содержащим нуклеиновую кислоту или вектор согласно изобретению. Указанным вектором может быть вектор любого типа, например рекомбинантный вектор, такой как экспрессионный вектор. При этом могут быть использованы любые из различных клеток-хозяев. В одном из вариантов изобретения клеткой-хозяином является прокариотическая клетка, например, E.coli. В одном из вариантов изобретения клеткой-хозяином является эукариотическая клетка, например клетка млекопитающего, такая как клетка яичника китайского хомячка (СНО). В одном из своих аспектов настоящее изобретение относится к способам получения антагониста согласно изобретению. Так, например, настоящее изобретение относится к способу получения антитела-антагониста против c-met (которое, как определено в настоящем изобретении, содержит полноразмерные последовательности и их фрагменты), где указанный способ включает экспрессию в подходящей клетке-хозяине рекомбинантного вектора согласно изобретению, кодирующего указанное антитело (или его фрагмент), и выделение указанного антитела. В одном из своих аспектов настоящее изобретение относится к промышленному изделию, включающему контейнер и композицию, содержащуюся в этом контейнере, где указанная композиция включает одно или несколько антител-антагонистов против с-met согласно изобретению. В одном из вариантов изобретения указанная композиция содержит нуклеиновую кислоту согласно изобретению. В одном из вариантов изобретения указанная композиция содержит антитело-антагонист, а также носитель, который, в некоторых вариантах изобретения, является фармацевтически приемлемым. В одном из вариантов изобретения промышленное изделие согласно изобретению также включает инструкции по введению указанной композиции (например, антитела-антагониста) индивидууму. В одном из своих аспектов настоящее изобретение относится к набору, содержащему первый контейнер, включающий композицию, содержащую одно или несколько антител-антагонистов против c-met согласно изобретению, и второй контейнер, включающий буфер. В одном из вариантов изобретения указанный буфер является фармацевтически приемлемым. В одном из вариантов изобретения композиция, содержащая антитело-антагонист, также содержит носитель, который, в некоторых вариантах изобретения, является фармацевтически приемлемым. В одном из вариантов изобретения указанный набор также включает инструкции по введению указанной композиции (например, антитела-антагониста) индивидууму. В одном из своих аспектов настоящее изобретение относится к применению антитела-антагониста против c-met согласно изобретению при получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное расстройство, иммунное расстройство (такое как аутоиммунное расстройство) и/или расстройство, ассоциированное с нарушением ангиогенеза. В одном из своих аспектов настоящее изобретение относится к применению нуклеиновой кислоты согласно изобретению при получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное расстройство, иммунное расстройство (такое как аутоиммунное расстройство) и/или расстройство, ассоциированное с нарушением ангиогенеза. В одном из своих аспектов настоящее изобретение относится к применению экспрессионного вектора согласно изобретению при получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное расстройство, иммунное расстройство (такое как аутоиммунное расстройство) и/или расстройство, ассоциированное с нарушением ангиогенеза. В одном из своих аспектов настоящее изобретение относится к применению клетки-хозяина согласно изобретению при получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное расстройство, иммунное расстройство (такое как аутоиммунное расстройство) и/или расстройство, ассоциированное с нарушением ангиогенеза. В одном из своих аспектов настоящее изобретение относится к применению промышленного изделия согласно изобретению при получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное расстройство, иммунное расстройство (такое как аутоиммунное расстройство) и/или расстройство, ассоциированное с нарушением ангиогенеза. В одном из своих аспектов настоящее изобретение относится к применению набора согласно изобретению при получении лекарственного средства для терапевтического и/или профилактического лечения заболевания, такого как рак, опухоль, клеточно-пролиферативное расстройство, иммунное расстройство (такое как аутоиммунное расстройство) и/или расстройство, ассоциированное с нарушением ангиогенеза. Настоящее изобретение относится к способам и композициям, которые пригодны для модулирования болезненных состояний, ассоциированных с нарушением регуляции HGF/c-met-путей передачи сигнала. HGF/c-met-путь передачи сигнала участвует во множестве биологических и физиологических функций, включая, например, пролиферацию клеток и ангиогенез. Таким образом, в одном из своих аспектов настоящее изобретение относится к способу, включающему введение индивидууму антитела согласно изобретению. В одном из своих аспектов настоящее изобретение относится к способу ингибирования c-met-активированной пролиферации клеток, где указанный способ включает контактирование клеток или тканей с эффективным количеством антитела согласно изобретению и, тем самым, ингибирование пролиферации клеток, ассоциированной с активацией c-met. В одном из своих аспектов настоящее изобретение относится к способу лечения патологического состояния, ассоциированного с нарушением регулирования активации c-met у индивидуума, где указанный способ включает введение указанному индивидууму эффективного количества антитела согласно изобретению и, тем самым, лечение указанного состояния. В одном из своих аспектов настоящее изобретение относится к способу ингибирования роста клеток, экспрессирующих c-met или фактор роста гепатоцитов или то и другое, где указанный способ включает контактирование указанных клеток с антителом согласно изобретению и, тем самым, ингибирование роста указанных клеток. В одном из вариантов изобретения указанные клетки контактируют с HGF, экспрессируемым другой клеткой (например, посредством паракринного эффекта). В одном из своих аспектов настоящее изобретение относится к способу терапевтического лечения млекопитающего, имеющего раковую опухоль, содержащую клетки, экспрессирующие c-met или фактор роста гепатоцитов или то и другое, где указанный способ включает введение указанному млекопитающему эффективного количества антитела согласно изобретению и, тем самым, эффективное лечение указанного млекопитающего. В одном из вариантов изобретения указанные клетки контактируют с HGF, экспрессируемым другой клеткой (например, посредством паракринного эффекта). В одном из своих аспектов настоящее изобретение относится к способу лечения или предупреждения клеточно-пролиферативного расстройства, ассоциированного с повышенной экспрессией или активностью c-met или фактора роста гепатоцитов, или того и другого, где указанный способ включает введение индивидууму, нуждающемуся в таком лечении, эффективного количества антитела согласно изобретению и, тем самым, эффективное лечение или предупреждение указанного клеточно-пролиферативного расстройства. В одном из вариантов изобретения указанным пролиферативным расстройством является рак. В одном из своих аспектов настоящее изобретение относится к способу ингибирования роста клеток, где рост указанных клеток, по меньшей мере частично, зависит от рост-потенцирующего действия c-met или фактора роста гепатоцитов, или того и другого, и где указанный способ включает контактирование указанных клеток с эффективным количеством антитела согласно изобретению и, тем самым, ингибирование роста указанных клеток. В одном из вариантов изобретения указанные клетки контактируют с HGF, экспрессируемым другой клеткой (например, посредством паракринного эффекта). Настоящее изобретение относится к способу терапевтического лечения опухоли у млекопитающего, где рост указанной опухоли, по меньшей мере частично, зависит от рост-потенцирующего действия c-met или фактора роста гепатоцитов, или от того и другого, и где указанный способ включает контактирование указанных клеток с эффективным количеством антитела согласно изобретению и, тем самым, эффективное лечение указанной опухоли. В одном из вариантов изобретения указанные клетки контактируют с HGF, экспрессируемым другой клеткой (например, посредством паракринного эффекта). Способы согласно изобретению могут быть применены для воздействия на любое релевантное патологическое состояние, например на клетки и/или ткани, ассоциированные с нарушением HGF/c-met-пути передачи сигнала. В одном из вариантов изобретения клетка, которая является мишенью в способе согласно изобретению, представляет собой раковую клетку. Так, например, раковой клеткой может быть клетка, выбранная из группы, состоящей из клеток рака молочной железы, клеток рака ободочной кишки, клеток рака легких, клеток папиллярной карциномы (например, щитовидной железы), клеток рака толстой кишки, клеток рака поджелудочной железы, клеток рака яичника, клеток рака шейки матки, клеток рака центральной нервной системы, клеток остеосаркомы, клеток карциномы почек, клеток гепатоцеллюлярной карциномы, клеток рака мочевого пузыря, клеток карциномы желудка, клеток плоскоклеточной карциномы головы и шеи, клеток меланомы и лейкозных клеток. В одном из вариантов изобретения клетка, которая является мишенью в способе согласно изобретению, представляет собой гиперпролиферирующую и/или гиперпластическую клетку. В одном из вариантов изобретения клетка, которая является мишенью в способе согласно изобретению, представляет собой диспластическую клетку. В другом варианте изобретения клетка, которая является мишенью в способе согласно изобретению, представляет собой метастатическую клетку. Способы согласно изобретению могут также включать дополнительные стадии лечения. Так, например, в одном из вариантов изобретения указанный способ дополнительно включает стадию, в которой клетку- и/или ткань-мишень (например, раковую клетку) подвергают лучевой терапии или химиотерапии. Как описано в настоящей заявке, активация c-met представляет собой важный биологический процесс, нарушение регуляции которого приводит к различным патологическим состояниям. В соответствии с этим в одном из вариантов способов согласно изобретению клеткой-мишенью (например, раковой клеткой) является клетка, в которой, по сравнению с нормальной клеткой той же самой ткани, наблюдается повышенный уровень активации c-met. В одном из вариантов изобретения способ согласно изобретению вызывает гибель клетки-мишени. Так, например, контактирование этой клетки с антагонистом согласно изобретению может приводить к нарушению способности данной клетки передавать сигнал по c-met-пути, и, тем самым, к ее гибели. Нарушение регуляции активации c-met (и тем самым, передачи сигнала) может быть обусловлено различными изменениями в клетках, включая, например, сверхэкспрессию HGF (узнаваемого лиганда c-met) и/или самого c-met. В соответствии с этим в некоторых вариантах изобретения способ согласно изобретению направлен на уничтожение клетки (например, раковой клетки), в которой c-met или фактор роста гепатоцитов, либо то и другое, экспрессируются на более высоком уровне, чем в нормальной клетке той же самой ткани. Клетка, экспрессирующая c-met, может регулироваться посредством HGF, происходящего от различных источников, то есть по аутокринному или паракринному механизму. Так, например, в одном из способов согласно изобретению клетка мишень контактирует/связывается с фактором роста гепатоцитов, экспрессируемым в другой клетке (например, по паракринному механизму). Указанная другая клетка может происходить от той же самой или от другой ткани. В одном из вариантов изобретения клетка-мишень контактирует/связывается с HGF, экспрессируемым самой клеткой-мишенью (например, по аутокринному механизму/петле). Активация и/или передача сигнала c-met может также происходить независимо от лиганда. Следовательно, в одном из вариантов способов согласно изобретению активация c-met в клетке-мишени происходит независимо от лиганда. Краткое описание графического материала На фиг.1 проиллюстрировано выравнивание последовательностей вариабельных областей легкой и тяжелой цепей: консенсусной последовательности легкой цепи человеческого антитела подгруппы I, консенсусной последовательности тяжелой цепи человеческого антитела подгруппы III, мышиного антитела против c-met 5D5 и На фиг.2 представлены различные последовательности HVR аффинно зрелых антител, выбранных из библиотек с отдельно рандомизированной HVR. На фиг.3 представлены последовательности HVR-Н3 аффинно зрелых антител, выбранных из пула библиотек, состоящих из комбинации из шести библиотек, включающих все шесть HVR, где каждая библиотека рандомизирована по одной HVR. На фиг.4 представлены результаты анализа Biacore выбранных антител против c-met. На фиг.5А,В и 6А,В представлены репрезентативные акцепторные консенсусные каркасные последовательности человеческого антитела, которые могут быть использованы в настоящем изобретении вместе со следующими идентификаторами последовательностей: Консенсусные каркасные последовательности вариабельной области тяжелой цепи (VН)(фиг.5А,В) консенсусная каркасная последовательность человеческой VH подгруппы I без CDR Кэбата (SEQ ID NO:19); консенсусная каркасная последовательность человеческой VH подгруппы I без удлиненных гипервариабельных областей (SEQ ID NO:20-22); консенсусная каркасная последовательность человеческой VH подгруппы II без CDR Кэбата (SEQ ID NO:23); консенсусная каркасная последовательность человеческой VH подгруппы II без удлиненных гипервариабельных областей (SEQ ID NO:24-26); консенсусная каркасная последовательность человеческой VH подгруппы III без CDR Кэбата (SEQ ID NO:27); консенсусная каркасная последовательность человеческой VH подгруппы III без удлиненных гипервариабельных областей (SEQ ID NO:28-30); акцепторная каркасная последовательность человеческой VH без CDR Кэбата (SEQ ID NO:31); акцепторная каркасная последовательность человеческой VH без удлиненных гипервариабельных областей (SEQ ID NO:32-33); акцепторная каркасная последовательность 2 человеческой VH без CDR Кэбата (SEQ ID NO:34); акцепторная каркасная последовательность 2 человеческой VH без удлиненных гипервариабельных областей (SEQ ID NO:35-37); Консенсусные каркасные последовательности вариабельной области легкой цепи (VL)(фиг.6А,В) консенсусная каркасная последовательность человеческой VL каппа подгруппы I (SEQ ID NO:38); консенсусная каркасная последовательность человеческой VL каппа подгруппы II (SEQ ID NO:39); консенсусная каркасная последовательность человеческой VL каппа подгруппы III (SEQ ID NO:40); консенсусная каркасная последовательность человеческой VL каппа подгруппы IV (SEQ ID NO:41). На фиг.7 представлены последовательности вариабельного домена легкой цепи (LС) и тяжелой цепи (НС) донорного антитела (мышиного антитела 5D5). На фиг.8 представлены графические данные для блокирования связывания HGF с его рецептором антителом согласно изобретению. На фиг.9 представлены графические данные для ингибирования активации рецептора HGF антителом согласно изобретению. На фиг.10 представлены графические данные для ингибирования пролиферации клеток антителом согласно изобретению: На фиг.11А,В представлены данные ингибирования фосфорилирования рецептора в присутствии антитела согласно изобретению. На фиг.11А проиллюстрировано фосфорилирование рецептора клеток Н358. На фиг.11В проиллюстрировано фосфорилирование рецептора клеток Н358, трансфицированных фактором HGF. На фиг.12 представлены графические данные, иллюстрирующие эффективность in vivo антитела согласно изобретению. На фиг.13 представлены аминокислотные последовательности каркасной области (FR), гипервариабельной области (HVR), первого константного домена (CL или СН1) и Fc-области (Fc) одного из вариантов антитела согласно изобретению (5D5.v2). Представленная последовательность Fc имеет мутации Т366S, L368A и Y407V во На фиг.14 представлена последовательность полипептида Fc, содержащая мутацию T366W в Способы осуществления настоящего изобретения Настоящее изобретение относится к способам, композициям, наборам и к промышленным изделиям для идентификации и/или использования ингибиторов пути передачи сигнала HGF/c-met. Подробное описание этих способов, композиций, наборов и промышленных изделий приводится ниже. Общие методы Настоящее изобретение может быть осуществлено, если это не оговорено особо, с применением стандартных методов, известных специалистам в области молекулярной биологии (включая методы рекомбинантных ДНК), микробиологии, биологии клетки, биохимии и иммунологии. Такие методы подробно описаны в литературе, например, в руководствах: Определения Используемый здесь термин
Используемый здесь термин Используемые здесь термины Термин В одном из вариантов изобретения
В одном из вариантов изобретения Используемые здесь термины
В случае, если для сравнения аминокислотных последовательностей используется программа ALIGN-2, то % идентичности данной аминокислотной последовательности А с данной аминокислотной последовательностью В (или, другими словами, данная аминокислотная последовательность А имеет или составляет определенный % идентичности с данной аминокислотной последовательностью В) вычисляют следующим образом: 100 · Х/Y, где Х представляет собой число аминокислотных остатков, которые были оценены как полностью соответствующие при выравнивании последовательностей с использованием программы ALIGN-2, с помощью которой проводили сравнение последовательностей А и В, и где Y представляет собой полное число аминокислотных последовательностей в последовательности В. При этом следует отметить, что если длина аминокислотной последовательности А не равна длине аминокислотной последовательности В, то % идентичности аминокислотной последовательности А аминокислотной последовательности В не равен % идентичности аминокислотной последовательности В аминокислотной последовательности А. Если это не оговорено особо, то общий % идентичности используемых здесь аминокислотных последовательностей вычисляли, как описано в предыдущем параграфе с использованием компьютерной программы ALIGN-2. Таблица А
Используемый здесь термин Используемые здесь термины Используемый здесь термин Используемый здесь термин Термины
Используемый здесь термин Описанные здесь моноклональные антитела, в частности, включают
Используемый здесь термин Используемый здесь термин Используемые здесь термины
Термин
Используемый здесь термин Термин Термины Используемый здесь термин Используемые здесь термины Нарушения ангиогенеза могут приводить к развитию многих расстройств, которые могут быть подвергнуты лечению композициями и способами согласно изобретению. Такими расстройствами являются ненеопластические и неопластические состояния. Неопластическими состояниями являются, но не ограничиваются ими, состояния, описанные выше. Ненеопластическими состояниями являются, но не ограничиваются ими, нежелательная или аберрантная гипертрофия, артрит, ревматоидный артрит (РА), псориаз, псориазные бляшки, саркоидоз, атеросклероз, атеросклеротические бляшки, диабетическая и другие пролиферативные ретинопатии, включая ретинопатию недоношенных или ретролентальную фиброплазию, неоваскулярная глаукома, старческая дегенерация желтого пятна, диабетический отек желтого пятна, неоваскуляризация роговицы глаза, неоваскуляризация трансплантата роговицы глаза, отторжение трансплантата роговицы глаза, неоваскуляризация сетчатки/сосудистой оболочки глаза, неоваскуляризация угла глазного яблока (покраснение глаза), неоваскулярное заболевание глаз, рестеноз сосудов, артериовенозная мальформация (АВМ), менингиома, гемангиома, ангиофиброма, гиперплазия щитовидной железы (включая болезнь Грейвса), трансплантация роговицы глаза и других тканей, хроническое воспаление, воспаление легких, острое поражение легких/респираторный дистресс-синдром взрослых (РДСВ), сепсис, первичная легочная гипертензия, злокачественные экссудаты в области легких, отек головного мозга (например, ассоциированный с ударом/закрытым повреждением/травмой головы), синовит, образование паннуса при РА, оссифицирующий миозит, гипертрофия костей, остеоартрит (ОА), не поддающийся лечению асцит, поликистоз яичника, эндометриоз, заболевания, ассоциированные с секвестрацией жидкости в третьем пространстве (полости полых органов) (панкреатит, туннельный синдром, ожоги, заболевание кишечника), фиброма матки, преждевременные роды, хроническое воспаление, такое как ВЗК (болезнь Крона и язвенный колит), отторжение почечного аллотрансплантата, воспалительное заболевание кишечника, нефротический синдром, нежелательный или аберрантный рост тканевой массы (не-раковых тканей), гемофилическая артропатия, гипертрофированные рубцы, подавление роста волос, синдром Ослера-Вебера, пиогенная гранулема, ретролентальная фиброплазия, склеродермия, трахома, васулярная адгезия (сращение сосудов), синовит, дерматит, преэклампсия, асциты, выпоты в область перикарда (например, ассоциированные с перикардитом) и плевральные выпоты. Используемый здесь термин Термин
Используемый здесь термин
В это определение также входят противогормональные средства, регулирующие, снижающие, блокирующие или ингибирующие действие гормонов, которые могут стимулировать рост раковой опухоли, и эти средства часто применяются для системного или общего лечения. Такими средствами могут быть сами гормоны. Примерами таких средств являются антиэстрогены и селективные модуляторы рецептора эстрогена (SERM), включая, например, тамоксифен (включая тамоксифен Нолвадекс®), ралоксифен Эвиста®, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY117018, онапристон и торемифен Фарестон®; антипрогестероны; ингибиторы рецептора эстрогена (ERD); средства, подавляющие или блокирующие функции яичников, например, агонисты рилизинг-фактора лютеинизирующего гормона (LHRH), такие как Лупрон® и ацетат лейпролида Элигард®, ацетат гозерелина, ацетат бузерелина и триптерелин; другие антиандрогены, такие как флутамид, нилутамид и бикалутамид; и ингибиторы ароматазы, ингибирующие фермент ароматазу и регулирующие продуцирование эстрогенов в коре надпочечника, такие как, например, 4(5)-имидазолы, аминоглутетимид, ацетат мегестрола Мегаза®, экземестан Аромазин®, форместан, фадрозол, ворозол Ривизор®, летрозол Фемара® и анастрозол Аримидекс®. Кроме того, в такое определение химиотерапевтических средств входят бифосфонаты, такие как клодронат (например, Бонефос® или Остак®), этидронат Дидрокал®, NE-58095, золедроновая кислота/золедронат Зомета®, алендронат Фозамакс®, памидронат Аредия®, тилудронат Скелид® или ризедронат Актонел®, а также троксацитабин (нуклеозидный аналог 1,3-диоксоланцитозина); антисмысловые олигонуклеотиды, в частности, олигонуклеотиды, ингибирующие экспрессию генов пути передачи сигнала, участвующих в нежелательной пролиферации клеток, такие как, например, РКС-альфа, Raf, H-Ras и рецептор эпидермального фактора роста (EGF-R); вакцины, такие как вакцина Тератопа® и вакцины, применяемые в генотерапии, например вакцина Алловектин®, вакцина Лейвектин® и вакцина Ваксид®; ингибитор топоизомеразы 1 Луртотекан®; rmRH Абареликс®; дитозилат лапатиниба (ингибитор тирозинкиназы, состоящий из двух небольших молекул Erb-2 и EGFR, также известный как GW572016) и фармацевтически приемлемые соли, оксиды или производные всех вышеуказанных соединений. Используемый здесь термин
Получение вариантов антител, продуцирующих пониженный НАМА-ответ или вообще не продуцирующих такого ответа Одним из важных аспектов клинической разработки подходящих терапевтических средств является снижение или элиминация НАМА-ответа (см. например, Khaxzaeli et al., J. Natl. Cancer. Inst. (1988), 80:937; Jaffers et al., Transplantation (1986) 41:572; Shawler et al., J. Immunol. (1985) 135:1530; Sears et al., J. Biol. Response Mod. (1984) 3:138; Miller et al., Blood (1983) 62:988; Hakimi et al., J. Immunol. (1991), 147:1352; Reichmann et al., Nature (1988) 332:323; Junghans et al., Cancer Res. (1990) 50:1495). Как описано в настоящей заявке, настоящее изобретение относится к антителам, которые были гуманизированы, так, что такая гуманизация приводила к снижению или элиминации НАМА-ответа. Варианты таких антител могут быть также получены известными рутинными методами, некоторые из которых описаны ниже. Так, например, аминокислотная последовательность описанного здесь антитела может служить в качестве исходной (родительской) последовательности, которая может быть использована для придания вариабельности каркасной(ым) и/или гипервариабельной(ым) последовательности(ям). Выбранная каркасная последовательность, к которой присоединена исходная гипервариабельная последовательность, называется здесь акцепторной человеческой каркасной последовательностью. Хотя акцепторные человеческие каркасные последовательности могут происходить или могут быть получены из человеческого иммуноглобулина (его VL- и/или VH-областей), однако, предпочтительно, чтобы акцепторные человеческие каркасные последовательности происходили или были получены из человеческой консенсусной каркасной последовательности, поскольку такие каркасные последовательности обладают, как было продемонстрировано, минимальной иммуногенностью для человека или вообще не обладают такой иммуногенностью. Если акцептор происходит от человеческого иммуноглобулина, то человеческая каркасная последовательность может быть, но необязательно, выбрана исходя из ее гомологии донорной каркасной последовательности путем выравнивания донорной каркасной последовательности с различными человеческими каркасными последовательностями в наборе человеческих каркасных последовательностей, а наиболее гомологичная каркасная последовательность может быть, но необязательно, выбрана в качестве акцептора. В одном из вариантов изобретения описанные здесь человеческие консенсусные каркасные последовательности происходят или их получают из консенсусных каркасных последовательностей VH подгруппы III и/или VL цепи каппа подгруппы I. Таким образом, акцепторная человеческая каркасная последовательность VH может содержать одну, две, три или все из нижеследующих каркасных последовательностей: FR1, включающей EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:42); FR2, включающей WVRQAPGKGLEWV (SEQ ID NO:43); FR3, включающей RFTISХ1DХ2SKNTХ3YLQMNSLRAEDTAVYYC (SEQ ID NO:50), где Х1 представляет собой А или R, Х2 представляет собой Т или N, а Х3 представляет собой А или L, FR4, включающей WGQGTLVTVSS (SEQ ID NO:45). Примерами консенсусных каркасных последовательностей VH-области являются: консенсусная каркасная последовательность человеческой VH подгруппы I без CDR Кэбата (SEQ ID NO:19); консенсусная каркасная последовательность человеческой VH подгруппы I без удлиненных гипервариабельных областей (SEQ ID NO:20-22); консенсусная каркасная последовательность человеческой VH подгруппы II без CDR Кэбата (SEQ ID NO:23); консенсусная каркасная последовательность человеческой VH подгруппы II без удлиненных гипервариабельных областей (SEQ ID NO:24-26); консенсусная каркасная последовательность человеческой VH подгруппы III без CDR Кэбата (SEQ ID NO:27); консенсусная каркасная последовательность человеческой VH подгруппы III без удлиненных гипервариабельных областей (SEQ ID NO:28-30); акцепторная каркасная последовательность человеческой VH без CDR Кэбата (SEQ ID NO:31); акцепторная каркасная последовательность человеческой VH без удлиненных гипервариабельных областей (SEQ ID NO:32-33); акцепторная каркасная последовательность 2 человеческой VH без CDR Кэбата (SEQ ID NO:34); или акцепторная каркасная последовательность 2 человеческой VH без удлиненных гипервариабельных областей (SEQ ID NO:35-37). В одном из вариантов изобретения акцепторная каркасная человеческая последовательность VH содержит одну, две, три или все из нижеследующих каркасных последовательностей: FR1, включающей EVQLVESGGGLVQPGGSLRLSCAAS (SEQ ID NO:42); FR2, включающей WVRQAPGKGLEWV (SEQ ID NO:43); FR3, включающей RFTISADTSKNTAYLQMNSLRAEDTAVYYC (SEQ ID NO:51); FR3, включающей RFTISADTSKNTAYLQMNSLRAEDTAVYYCA (SEQ ID NO:52); RFTISADTSKNTAYLQMNSLRAEDTAVYYCAR (SEQ ID NO:53); RFTISADTSKNTAYLQMNSLRAEDTAVYYCS (SEQ ID NO:54); RFTISADTSKNTAYLQMNSLRAEDTAVYYCSR (SEQ ID NO:55); и FR4, включающей WGQGTLVTVSS (SEQ ID NO:45). Акцепторная каркасная человеческая последовательность VL может содержать одну, две, три или все из нижеследующих каркасных последовательностей: FR1, включающей DIQMTQSPSSLSASVGDRVTITC (SEQ ID NO:46); FR2, включающей WYQQKPGKAPKLLIY (SEQ ID NO:47); FR3, включающей GVPSRFSGSGSGTDFTLTISSLQPEDFATYYC (SEQ ID NO:48); FR4, включающей FGQGTKVEIK (SEQ ID NO:49). Примерами консенсусных каркасных последовательностей VL являются: консенсусная каркасная последовательность человеческой VL цепи каппа подгруппы I (SEQ ID NO:38); консенсусная каркасная последовательность человеческой VL цепи каппа подгруппы II (SEQ ID NO:39); консенсусная каркасная последовательность человеческой VL цепи каппа подгруппы III (SEQ ID NO:40); или консенсусная каркасная последовательность человеческой VL цепи каппа подгруппы IV (SEQ ID NO:41). Хотя акцепторная последовательность может быть идентична выбранной человеческой каркасной последовательности, независимо от того, происходит ли она от человеческого иммуноглобулина или от человеческой консенсусной каркасной последовательности, однако, в настоящем изобретении также рассматривается акцепторная последовательность, которая, по сравнению с последовательностью человеческого иммуноглобулина или человеческой консенсусной каркасной последовательностью, может содержать уже существующие аминокислотные замены. Эти уже существующие замены являются, предпочтительно, минимальными, а обычно они представляют собой только четыре, три, две модификации или одну аминокислотную модификацию аминокислот по сравнению с последовательностью человеческого иммуноглобулина или консенсусной каркасной последовательностью. В акцепторные человеческие каркасные последовательности VL и/или VH вводят остатки гипервариабельной области не-человеческого антитела. Так, например, могут быть включены остатки, соответствующие остаткам CDR по Кэбату, остаткам гипервариабельной петли по Чотия, остаткам AbМ и/или контактным остаткам. При этом могут быть, но необязательно, включены остатки удлиненной гипервариабельной области, а именно 24-34 (L1), 50-56 (L2) и 89-97 (L3), 26-35 (Н1), 50-65 или 49-65 (Н2) и 93-102, 94-102 или 95-102 (Н3). Что касается обсуждаемого в настоящей заявке В описанных здесь примерах варианты гибридной гипервариабельной области получают путем мутагенеза нуклеиновой кислоты, кодирующей человеческие акцепторные последовательности, методом Кункеля с использованием отдельного олигонуклеотида для каждой гипервариабельной области. Kunkel et al., Methods Enzymol. 154:367-382 (1987). Соответствующие замены могут быть внесены в каркасную и/или в гипервариабельную область рутинными методами для коррекции и повторного подтверждения соответствующих взаимодействий Фаговое (фагмидное) представление (также называемое в некоторых случаях методом фагового представления) может применяться как удобный и быстрый метод продуцирования и скрининга множества различных потенциальных вариантов антител в библиотеке, генерированной путем рандомизации последовательностей. Однако специалистам известны и другие методы получения и скрининга модифицированных антител. Метод фагового (фагмидного) представления является эффективным средством генерирования и отбора новых белков, связывающихся с лигандом, таким как антиген. Методы фагового (фагмидного) представления позволяют генерировать крупные библиотеки вариантов белков, из которых могут быть быстро отобраны белки, имеющие последовательности, связывающиеся с молекулой-мишенью с высокой аффинностью. Нуклеиновые кислоты, кодирующие варианты полипептидов, обычно присоединяют к последовательности нуклеиновой кислоты, кодирующая белок вирусной оболочки, такой как ген белка III или ген белка VIII. Были разработаны моновалентные системы фагмидного представления, в которых последовательность нуклеиновой кислоты, кодирующая белок или полипептид, присоединена к последовательности нуклеиновой кислоты, кодирующей часть гена белка III (Bass S., Proteins 8:309 (1990); Lowman & Wells, Methods: A Companion to Methods in Enzymology, 3:205 (1991)). В моновалентной системе фагмидного представления гибридный ген экспрессируется на низком уровне, а также экспрессируется ген белков III дикого типа, в результате чего сохраняется инфекционность частиц. Методы генерирования пептидных библиотек и скрининга этих библиотек описаны во многих патентах (например, в патенте США Библиотеки антител или антигенсвязывающих полипептидов были получены различными методами, включая модификацию одного гена путем встраивания рандомизированных последовательностей ДНК или клонирования семейства родственных генов. Методы представления антител или антигенсвязывающих фрагментов с применением фагового (фагмидного) представления описаны в патентах США Методы замены выбранных аминокислот, кодируемых матричной нуклеиновой кислотой, хорошо известны специалистам, и некоторые из них описаны в настоящей заявке. Так, например, остатки гипервариабельной области могут быть заменены в соответствии с методом Кункеля (см., например, Kunkel et al., Methods Enzymol. 154:367-382 (1987)). Последовательность олигонуклеотидов включает один или несколько сконструированных наборов кодонов для модифицируемых остатков гипервариабельной области. Набор кодонов представляет собой различные последовательности нуклеотидных триплетов, используемых для кодирования нужных вариантов аминокислот. Наборы кодонов могут быть представлены символами, обозначающими конкретные нуклеотиды или эквимолярные смеси нуклеотидов, указанные ниже в соответствии с кодом, утвержденным Международным биохимическим союзом (IUB). Коды IUB
Так, например, в наборе кодонов DVK D может представлять собой нуклеотиды А или G или Т; V может представлять собой А или G или С; и К может представлять собой G или Т. Этот набор кодонов может включать 18 различных кодонов и может кодировать аминокислоты Ala, Trp, Tyr, Lys, Thr, Asn, Lys, Ser, Arg, Asp, Glu, Gly и Сys. Наборы олигонуклеотидов или праймеров могут быть синтезированы стандартными методами. Набор олигонуклеотидов может быть синтезирован, например, методом твердофазного синтеза и может содержать последовательности, представляющие все возможные комбинации нуклеотидных триплетов, составляющих указанный набор кодонов и кодирующих нужную группу аминокислот. Методы синтезы олигонуклеотидов с В одном из методов последовательности нуклеиновой кислоты, кодирующие варианты аминокислот, могут быть получены путем олигонуклеотид-опосредуемого мутагенеза. Этот метод хорошо известен специалистам и описан Zoller et al., Nucleic. Acids. Res., 10:6487-6504 (1987). Вкратце, последовательности нуклеиновой кислоты, кодирующие варианты аминокислот, получают путем гибридизации последовательностей олигонуклеотидов, имеющих нужные наборы кодонов, с матричной ДНК, где указанная матрица представляет собой одноцепочечную форму плазмиды, содержащую последовательность матричной нуклеиновой кислоты, кодирующей вариабельную область. После гибридизации ДНК-полимеразу используют для синтеза полноразмерной второй комплементарной цепи матрицы, которая, таким образом, будет включать олигонуклеотидный праймер и будет содержать наборы кодонов, имеющиеся в данной олигонуклеотидной последовательности. В общих чертах олигонуклеотиды имеют длину, по меньшей мере, в 25 нуклеотидов. Оптимальный олигонуклеотид имеет 12-15 нуклеотидов, которые полностью комплементарны матрице с обеих сторон от нуклеотида(ов), кодирующего(их) мутацию(и). Это дает гарантию того, что данный олигонуклеотид будет соответствующим образом гибридизоваться с одноцепочечной молекулой матричной ДНК. Олигонуклеотиды могут быть легко синтезированы методами, известными специалистам, например, методами, описанными Crea et al., Proc. Natl. Acad. Sci., USA, 75:5765 (1978). Матричную ДНК генерируют с помощью векторов, которые происходят от бактериофаговых векторов М13 (подходящими векторами являются коммерчески доступные векторы М13mp18 и М13mp19), или с помощью векторов, содержащих одноцепочечный фаговый ориджин репликации, описанный Viera et al., Meth. Enzymol., 153:3 (1987). Таким образом, ДНК, которая должна быть мутирована, может быть встроена в один из этих векторов с получением одноцепочечной матрицы. Получение одноцепочечной матрицы описано в разделах 4.21-4.41 руководства Sambrook et al., см. выше. Для модификации нативной последовательности ДНК олигонуклеотид гибридизуют с одноцепочечной матрицей в подходящих условиях гибридизации. Затем добавляют ДНК-полимеризующий фермент, обычно ДНК-полимеразу Т7 или фрагмент ДНК-полимеразы I Кленова, для синтеза комплементарной цепи матрицы с использованием олигонуклеотида в качестве праймера. Таким образом, получают гетеродуплексную молекулу, в которой одна цепь ДНК кодирует мутированную форму гена 1, а другая цепь (исходная матрица) кодирует нативную не модифицированную последовательность гена 1. Затем эту гетеродуплексную молекулу переносят в подходящую клетку-хозяина, обычно прокариота, такого как JМ101 E.coli. После культивирования клетки высевают на планшеты с агарозой и скринируют с использованием олигонуклеотидного праймера, радиоактивно меченного 32-фосфатом, для идентификации бактериальных колоний, содержащих мутированную ДНК. Вышеописанный метод может быть модифицирован в целях получения гомодуплексной молекулы, в которой обе цепи плазмиды имеют мутацию(и). Для этого осуществляют нижеследующие модификации. Одноцепочечный олигонуклеотид отжигают с одноцепочечной матрицей, описанной выше. Смесь трех дезоксирибонуклеотидов, таких как дезоксирибоаденозин (dАТР), дезоксирибогуанозин (dGTP) и дезоксириботимидин (dTT), объединяют с модифицированным тиодезоксирибоцитозином, обозначенным dСТР-(aS)(который может быть получен у Amersham). Затем эту смесь добавляют в комплекс Как было показано ранее, последовательность набора олигонуклеотидов имеет длину, достаточную для гибридизации с матричной нуклеиновой кислотой, и может также, но необязательно, содержать рестрикционные сайты. Матричная ДНК может быть получена с помощью векторов, происходящих от бактериофаговых векторов М13, или с помощью векторов, содержащих одноцепочечный фаговый ориджин репликации, описанный Viera et al., Meth. Enzymol., 153:3 (1987). Таким образом, ДНК, предназначенная для мутирования, должна быть встроена в один из этих векторов с получением одноцепочечной матрицы. Получение одноцепочечной матрицы описано в разделах 4.21-4.41 руководства Sambrook et al., см. выше. В соответствии с другим методом библиотека может быть сконструирована путем создания выше- и нижерасположенных наборов олигонуклеотидов, каждый из которых имеет множество олигонуклеотидов с различными последовательностями, где указанные различные последовательности определяются наборами кодонов, присутствующих в последовательности указанных олигонуклеотидов. Эти наборы из выше- и нижерасположенных олигонуклеотидов вместе с последовательностью матричной нуклеиновой кислоты, кодирующей вариабельный домен, могут быть использованы в полимеразной цепной реакции для получения Последовательности ПЦР-праймеров включают один или несколько наборов кодонов, сконструированных в целях сообщения белкам доступности для растворителя и высокой степени разнообразия положений в гипервариабельной области. Как описано выше, набором кодонов является набор из различных нуклеотидных триплетных последовательностей, кодирующих нужные варианты аминокислот. Антитела, отобранные в соответствующих стадиях скрининга/отбора и удовлетворяющие нужным критериям, могут быть выделены и клонированы стандартными рекомбинантными методами. Векторы, клетки-хозяева и рекомбинантные методы Для продуцирования антитела согласно изобретению методами рекомбинантных ДНК нуклеиновую кислоту, кодирующую такое антитело, выделяют и встраивают в реплицируемый вектор для последующего клонирования (амплификации ДНК) или экспрессии. ДНК, кодирующая антитело, может быть легко выделена и секвенирована в соответствии со стандартными процедурами (например, с использованием олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелую и легкую цепи антитела). Для этой цели подходящими являются многие векторы. Выбор вектора отчасти зависит от используемой клетки-хозяина. Обычно предпочтительными клетками-хозяевами являются либо прокариотические клетки, либо эукариотические клетки (обычно, клетки млекопитающих). Генерирование антител с использованием прокариотических клеток-хозяев Конструирование вектора Полинуклеотидные последовательности, кодирующие полипептидные компоненты антитела согласно изобретению, могут быть получены стандартными рекомбинантными методами. Нужные полинуклеотидные последовательности могут быть выделены и секвенированы из антитело-продуцирующих клеток, таких как гибридомные клетки. Альтернативно полинуклеотиды могут быть синтезированы с помощью синтезатора нуклеотидов или ПЦР-методами. После получения последовательностей, кодирующих полипептиды, эти последовательности встраивают в рекомбинантный вектор, способный реплицироваться и экспрессировать гетерологичные полинуклеотиды в прокариотических хозяевах. В целях настоящего изобретения могут быть использованы многие доступные и известные векторы. Выбор соответствующего вектора зависит, главным образом, от размера нуклеиновых кислот, встраиваемых в указанный вектор, и от конкретной клетки-хозяина, трансформируемой этим вектором. Каждый вектор содержит различные компоненты, в зависимости от его функции (амплификации или экспрессии гетерологичного полинуклеотида или того и другого) и от его совместимости с конкретной клеткой-хозяином, в которой он находится. Компонентами векторов обычно являются, но не ограничиваются ими, ориджин репликации, селективный маркерный ген, промотор, сайт связывания с рибосомой (RBS), сигнальная последовательность, вставка гетерологичной нуклеиновой кислоты и последовательность терминации транскрипции. В основном плазмидные векторы, содержащие репликон и регуляторные последовательности, происходящие от видов, совместимых с клеткой-хозяином, используются с этими же хозяевами. Такой вектор обычно содержит сайт репликации, а также последовательности-маркеры, способствующие проведению фенотипического отбора в трансформированных клетках. Так, например, E.coli обычно трансформируют плазмидой рВR322, происходящей от E.coli определенного вида. рВR322 содержит гены, кодирующие резистентность к ампициллину (Аmp) и к тетрациклину (Теt), и позволяет легко идентифицировать трансформированные клетки. рВR322 и ее производные или другие микробные плазмиды или бактериофаг могут также содержать промоторы, которые могут быть использованы в этом микробном микроорганизме для экспрессии эндогенных белков, либо они могут быть модифицированы так, чтобы они содержали такие промоторы. Примеры производных рВR322, используемых для экспрессии конкретных антител, подробно описаны в патенте США Кроме того, фаговые векторы, содержащие репликон и регуляторные последовательности, совместимые с микроорганизмом-хозяином, могут быть использованы в качестве векторов, трансформирующих эти клетки-хозяева. Так, например, бактериофаг, такой как Экспрессионный вектор согласно изобретению может содержать две или более пар промотор-цистрон, каждая из которых кодирует полипептидные компоненты. Промотором является не транслируемая регуляторная последовательность, локализованная выше (со стороны 5′-конца) от цистрона, модулирующего ее экспрессию. Прокариотические промоторы обычно подразделяются на два класса, индуцибельные и конститутивные. Индуцибельным промотором является промотор, который инициирует повышение уровней транскрипции цистрона, находящегося под его контролем, в ответ на изменения условий культивирования, например, в присутствии или в отсутствии питательных элементов или при изменении температуры. Большинство промоторов, распознаваемых различными потенциальными клетками-хозяевами, хорошо известны специалистам. Выбранный промотор может быть функционально присоединен к цистронной ДНК, кодирующей легкую или тяжелую цепь, путем выделения промотора из ДНК-источника посредством ее гидролиза рестриктирующими ферментами и встраивания выделенной промоторной последовательности в вектор согласно изобретению. Нативная промоторная последовательность и многие гетерологичные промоторы могут быть использованы для прямой амплификации и/или экспрессии генов-мишеней. В некоторых вариантах изобретения используются гетерологичные промоторы, поскольку они, по сравнению с нативным промотором для нужных полипептидов, позволяют значительно повышать уровень транскрипции и увеличивать выходы экспрессируемого гена-мишени. Промоторами, подходящими для использования в прокариотических хозяевах, являются промотор РhoA, промоторные системы В одном из аспектов изобретения каждый цистрон в рекомбинантном векторе содержит такой компонент, как секреторную сигнальную последовательность, регулирующую транслокацию экспрессируемых полипептидов через мембрану. В общих чертах, указанной сигнальной последовательностью может быть компонент вектора либо такой последовательностью может быть часть нужного ДНК-полипептида, встраиваемого в этот вектор. Сигнальной последовательностью, выбранной в целях осуществления изобретения, должна быть последовательность, распознаваемая и процессируемая (то есть отщепляемая сигнальной пептидазой) клеткой-хозяином. В случае использования прокариотических клеток-хозяев, не распознающих и не процессирующих сигнальные последовательности, которые являются нативными для гетерологичных полипептидов, эту сигнальную последовательность заменяют прокариотической сигнальной последовательностью, выбранной, например, из группы, состоящей из лидерной последовательности щелочной фосфатазы, пенициллиназы, lpp или термостабильного энтеротоксина II (SТII), LamB, PhoE, PelB, OmpA и МВР. В одном из вариантов изобретения сигнальными последовательностями, используемыми в обоих цистронах экспрессионной системы, являются сигнальные последовательности SТII или их варианты. В другом аспекте изобретения продуцирование иммуноглобулинов согласно изобретению может происходить в цитоплазме клетки-хозяина, а поэтому в данном случае не требуется присутствия секретирующих сигнальных последовательностей в каждом цистроне. В соответствии с этим легкие и тяжелые цепи иммуноглобулина экспрессируются и подвергаются укладке и сборке с образованием функциональных иммуноглобулинов в цитоплазме. Некоторые штаммы хозяев (например, trxB–-штаммы E.coli) имеют соответствующие условия в цитоплазме, которые благоприятствуют образованию дисульфидных связей, и тем самым, способствуют правильной укладке и сборке субъединиц экспрессируемого белка. Proba & Pluckthun Gene, 159:203 (1995). Настоящее изобретение относится к экспрессионной системе, в которой количественное отношение экспрессированных полипептидных компонентов может быть модулировано для максимизации выхода секретированных и Один из методов модуляции эффективности трансляции описан в патенте США Набор генерируемых векторов, предпочтительно, имеет различные эффективности TIR для каждого присутствующего в нем цистрона. Такой ограниченный набор позволяет проводить сравнение уровня экспрессии каждой цепи, а также выхода нужных продуктов антитела с использованием различных комбинаций эффективностей TIR. Эффективности TIR могут быть определены путем количественной оценки уровня экспрессии репортерного гена, как подробно описано в патенте США Прокариотическими клетками-хозяевами, подходящими для экспрессии антител согласно изобретению, являются архебактерии (Archaebacteria) и эубактерии (Eubacteria), такие как грам-отрицательные или грам-положительные микроорганизмы. Примерами подходящих бактерий являются Esсherichia (например, E.coli), Bacilli (например, B.subtilis), энтеробактерии, бактерии вида Pseudomonas (например, P.aeruginosa), Salmonella typhimurium, Serratia marcescаns, Klebsiella, Proteus, Shigella,Rhizobia, Vitreoscilla или Paracoccus. В одном из вариантов изобретения используются грам-отрицательные клетки. В одном из вариантов изобретения в качестве хозяев согласно изобретению используются клетки E.coli. Примерами штаммов E.coli являются штамм W3110 (Bachmann, Cellular & Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp.1190-1219; АТСС, номер депозита Продуцирование антител Клетки-хозяева трансформируют вышеуказанными экспрессионными векторами и культивируют в стандартной питательной среде, модифицированной так, чтобы она была подходящей для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих нужные последовательности. Термин Прокариотические клетки, используемые для продуцирования полипептидов согласно изобретению, культивируют в среде, известной специалистам и подходящей для культивирования выбранных клеток-хозяев. Примерами подходящей среды являются бульон Луриа (LВ) с необходимыми питательными добавками. В некоторых вариантах изобретения такая среда также содержит средство для отбора, выбранное исходя из конструирования экспрессионного вектора, которое способствуют селективному росту прокариотических клеток, содержащих экспрессионный вектор. Так, например, в среду для культивирования клеток, экспрессирующих ген резистентности к ампициллину, добавляют ампициллин. Помимо источников углерода, азота и неорганического фосфата в среду могут быть также включены любые необходимые добавки в соответствующих концентрациях, вводимые как отдельно, так и в смеси с другими добавками или средой, такими как комплексный источник азота. Культуральная среда может, но необязательно, содержать один или несколько восстановителей агентов, выбранных из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритритола и дитиотреитола. Прокариотические клетки-хозяева культивируют при соответствующих температурах. Для культивирования E.coli, предпочтительной температурой является, например, температура примерно от 20 до 39°С, более предпочтительно, примерно от 25 до 37°С, а еще более предпочтительно, примерно при 30°С. рН среды может варьироваться в пределах примерно от 5 до 9, в зависимости, главным образом, от организма хозяина. Для E.coli, рН составляет, предпочтительно, примерно от 6,8 до 7,4, а более предпочтительно, примерно 7,0. Если в экспрессионном векторе согласно изобретению используется индуцибельный промотор, то экспрессия белка индуцируется в условиях, подходящих для активации данного промотора. В одном из аспектов изобретения для регуляции транскрипции полипептидов используются промоторы РhoA. В соответствии с этим трансформированные клетки-хозяева культивируют в среде для индуцирования с ограниченным содержанием фосфата. Предпочтительной средой с ограниченным содержанием фосфата является среда С.R.А.Р. (см., например, Simmons et al., J. Immunol. Methods (2002) 263:133-147). В комбинации с используемой векторной конструкцией могут быть использованы и различные другие индукторы, известные специалистам. В одном из вариантов изобретения экспрессируемые полипептиды согласно изобретению секретируются в периплазму клеток-хозяев и могут быть выделены из этой периплазмы. Выделение белка, главным образом, осуществляют путем дизрупции микроорганизма такими методами, как осмотический шок, обработка ультразвуком или лизис. После дизрупции клеток клеточный дебрис или целые клетки могут быть удалены путем центрифугирования или фильтрации. Эти белки могут быть дополнительно очищены, например, с помощью хроматографии на аффинной смоле. Альтернативно белки могут транспортироваться в клеточную среду и отделяться от нее. Клетки могут быть удалены из культуры, а супернатант культуры может быть подвергнут фильтрации и концентированию для дополнительной очистки полученных белков. Экспрессированные полипептиды могут быть затем выделены и идентифицированы хорошо известными методами, такими как электрофорез в полиакриламидном геле (ПААГ) и Вестерн-блот-анализ. В одном из аспектов изобретения антитела продуцируют в большом количестве посредством ферментации. Для получения рекомбинантных белков могут быть проведены различные крупномасштабные процедуры ферментации в культуре с подпиткой. Крупномасштабную ферментацию проводят в ферментере емкостью, по меньшей мере, 1000 литров, а предпочтительно, примерно от 1000 до 100000 литров. Такие ферментеры снабжены лопастной мешалкой для равномерного распределения кислорода и микроэлементов, а в частности глюкозы (предпочтительного источника углерода/энергии). Термин В процессе ферментации индуцирование экспрессии белков обычно инициируют после того как клетки, культивируемые в подходящих условиях, достигнут нужной плотности, например, OD550, составляющей примерно 180-220, то есть ранней стационарной фазы. В комбинации с используемой векторной конструкцией могут быть использованы и различные другие индукторы, известные специалистам и описанные выше. Перед индуцированием клетки могут быть культивированы в течение меньшего периода времени. Клетки обычно индуцируют в течение примерно 12-50 часов, хотя время индуцирования может быть увеличено или уменьшено. Для увеличения выхода продукта и для улучшения качества полипептидов согласно изобретению могут быть изменены различные условия ферментации. Так, например, для обеспечения Для минимизации протеолиза экспрессируемых гетерологичных белков (а в частности, белков, которые являются протеолитически чувствительными), в настоящем изобретении могут быть использованы некоторые штаммы-хозяева, дефицитные по протеолитическим ферментам. Так, например, штаммы клеток-хозяев могут быть модифицированы для введения генетической(их) мутации(й) в гены, кодирующие известные бактериальные протеазы, такие как протеаза III, OmpT, DegP, Tsp, протеаза I, протеаза Mi, протеаза V, протеаза VI и их комбинации. Некоторые дефицитные по протеазе штаммы E.coli являются доступными и описаны, например, в публикации Joly et al. (1998), см. выше; Georgiou et al., патент США В одном из вариантов изобретения штаммы E.coli, дефицитные по протеолитическим ферментам и трансформированные плазмидами, сверхэкспрессирующими один или несколько белков-шаперонов, используются в качестве клеток-хозяев в экспрессионной системе согласно изобретению. Очистка антител В одном из вариантов изобретения продуцируемый здесь белок антитела дополнительно очищают для получения препаратов, которые являются, по существу, гомогенными и которые могут быть использованы для проведения дополнительных анализов и для других целей. Для таких случаев могут быть применены стандартные методы очистки белков, известные специалистам. Примерами подходящих методов очистки являются следующие процедуры очистки: фракционирование на иммуноаффинных или ионообменных колонках, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на двуокиси кремния или на катионообменной смоле, такой как DЕАЕ, хроматофокусирование, электрофорез ДСН-ПААГ, осаждение сульфатом аммония и гель-фильтрация с использованием, например, сефадекса G-75. В одном из аспектов изобретения для иммуноаффинной очистки полноразмерных продуктов антител согласно изобретению используют белок А, иммобилизованный на твердой фазе. Белок А представляет собой 41 кД-белок клеточной стенки, происходящий от Staphylococcus aureus, который с высокой аффинностью связывается с Fc-областью антител (Lindmark et al. (1983), J. Immunol. Meth. 62:1-13). Твердой фазой, на которой иммобилизован белок А, является предпочтительно колонка, имеющая стеклянную поверхность или кремнийсодержащую поверхность, а более предпочтительно стеклянная колонка с регулируемым размером пор или колонка с нанесенной на нее кремниевой кислотой. В некоторых вариантах указанная колонка покрыта реагентом, таким как глицерин, для предотвращения неспецифического присоединения примесей. В первой стадии очистки препарат, полученный из клеточной культуры, описанной выше, наносят на твердую фазу с иммобилизованным на ней белком А для специфического связывания представляющего интерес антитела с белком А. Затем твердую фазу промывают для удаления примесей, неспецифически связанных с твердой фазой. И наконец, представляющее интерес антитело отделяют от твердой фазы путем элюирования. Продуцирование антител с использованием эукариотических клеток-хозяев Компонентами векторов обычно являются, но не ограничиваются ими, один или несколько из следующих компонентов: сигнальная последовательность, ориджин репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции. (i) Компонент сигнальная последовательность Вектор, используемый в эукариотических клетках-хозяевах, может также содержать сигнальную последовательность или другой полипептид, имеющий специфический сайт расщепления у N-конца зрелого белка, или представляющий интерес полипептид. Выбранной гетерологичной сигнальной последовательностью, предпочтительно, является последовательность, которая распознается и процессируется (то есть отщепляется сигнальной пептидазой) клеткой-хозяином. Для экспрессии в клетках млекопитающих используются сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например сигнальная последовательность gD вируса простого герпеса. ДНК для такой области предшественника лигируют с антителокодирующей ДНК с сохранением рамки считывания. (ii) Ориджин репликации Обычно такой компонент как ориджин репликации не является необходимым для экспрессионных векторов млекопитающих. Так, например, ориджин репликации SV40 обычно используется только потому, что он содержит ранний промотор. (iii) Компонент селективный ген Экспрессионный и клонирующий векторы могут содержать селективный ген, также называемый селективным маркером. Обычно селективные гены кодируют белки, которые: (а) сообщают резистентность к антибиотиками или к другим токсинам, например к ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементируют дефицит, обусловленный ауксотрофией, если это необходимо, или (с) обеспечивают доставку важных питательных веществ, которые не поступают из комплексных сред. Одним из примеров схемы отбора является использование лекарственного средства, прекращающего рост клетки-хозяина. Клетки, которые были успешно трансформированы гетерологичным геном, продуцируют белок, сообщающий резистентность к лекарственному средству, и, тем самым, способствующий выживанию этих клеток в селективной среде. Для такого доминантного отбора могут быть использованы лекарственные средства, такие как неомицин, микофеноловая кислота и гигромицин. Другими примерами подходящих селективных маркеров для клеток млекопитающих являются маркеры, которые позволяют идентифицировать клетки, способные встраивать в свой геном антителокодирующую нуклеиновую кислоту, например, такие маркеры, как ген DHFR, тимидинкиназы, металлотионеина-I и II, а предпочтительно гены, кодирующие металлотионеин приматов, ген аденозин-дезаминазы, орнитин-декарбоксилазы и т.п. Так, например, клетки, трансформированные селективным геном DHFR, сначала идентифицируют путем культивирования всех трансформантов в культуральной среде, содержащей метотрексат (Mtx), конкурентный антагонист DHFR. Если используется DHFR дикого типа, то подходящей клеткой-хозяином является клеточная линия яичника китайского хомячка (СНО), дефицитная по DHFR-активности (например, АТСС СRL-9069). Альтернативно клетки-хозяева (в частности, хозяева дикого типа, содержащие эндогенный DHFR), трансформированные или ко-трансформированные ДНК-последовательностями, кодирующими антитело, белок DHFR дикого типа и другой селективный маркер, такой как аминогликозид-3′-фосфотрансфераза (АРН), могут быть отобраны путем культивирования клеток в среде, содержащей агент для отбора по селективному маркеру, такой как аминогликозидный антибиотик, например канамицин, неомицин или G418 (см. патент США (iv) Компонент промотор Экспрессионные и клонирующие векторы обычно содержат промотор, распознаваемый организмом-хозяином и функционально присоединенный к нуклеиновой кислоте, кодирующей полипептид антитела. Промоторные последовательности, подходящие для эукариотов, известны специалистам. Фактически все гены эукариотов имеют АТ-богатую область, локализованную приблизительно в области, расположенной на 25-30 нуклеотидов выше от сайта инициации транскрипции. Другой такой последовательностью, локализованной приблизительно в области, расположенной на 70-80 нуклеотидов выше от сайта инициации транскрипции многих генов, является область CNCAAT, где N может означать любой нуклеотид. У 3′-конца большинства эукариотических генов расположена последовательность ААТААА, которая может служить сигналом для присоединения poly A-хвоста к 3′-концу кодирующей последовательности. Все указанные последовательности могут быть соответствующим образом встроены в эукариотические экспрессионные векторы. Транскрипция полипептидов антител из векторов, присутствующих в клетках-хозяевах млекопитающих, регулируется, например, промоторами, полученными из геномов таких вирусов, как полиомавирус, вирус оспы домашней птицы, аденовирус (такой как аденовирус 2), вирус коровьей папиломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и обезьяний вирус 40 (SV40); гетерологичными промоторами млекопитающих, например, промотором актина или промотором иммуноглобулина; и промоторами белков теплового шока, при условии, что такие промоторы являются совместимыми с системами клеток-хозяев. Ранние и поздние промоторы вируса SV40 обычно получают в виде рестрикционного фрагмента SV40, который также содержит ориджин репликации вируса SV40. Предранний промотор человеческого цитомегаловируса обычно получают в виде рестрикционного HindIII-фрагмента Е. Система экспрессии ДНК в клетках-хозяевах млекопитающих, полученная на основе коровьего папиломавируса, используемого в качестве вектора, описана в патенте США (v) Компонент энхансерный элемент Транскрипция ДНК, кодирующей полипептид антитела согласно изобретению в высших эукариотах, часто усиливается при встраивании в вектор энхансерной последовательности. В настоящее время известно много энхансерных последовательностей, происходящих от генов млекопитающих (генов глобина, эластазы, альбумина, (vi) Компонент сайт терминации транскрипции Экспрессионные векторы, используемые в эукариотических клетках-хозяевах, обычно также содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно расположены со стороны 5′-конца, а иногда, со стороны 3′-конца от нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Эти области содержат нуклеотидные сегменты, транскрибированные в виде полиаденилированных фрагментов в нетранслированной части мРНК, кодирующей антитело. В качестве такого компонента, как сайт терминации транскрипции, может быть использована область полиаденилирования коровьего гормона роста (см. заявку WO 94/11026 и имеющееся там описание экспрессионных векторов). (vii) Отбор и трансформация клеток-хозяев Клетками-хозяевами, подходящими для клонирования или экспрессии ДНК в вышеописанных векторах, являются описанные здесь клетки высших эукариотов, включая клетки-хозяева позвоночных. Размножение клеток позвоночных в культуре (в тканевой культуре) уже стало рутинной процедурой. Примерами подходящих линий клеток-хозяев млекопитающих являются клеточная линия CV1 почек обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); клеточная линия почек человеческого эмбриона (клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol. 36:59 (1977)); клетки почек детеныша хомячка (ВНК, АТСС CCL 10); клетки яичника китайского хомячка/DHFR (CHO, Urlaub et al., Proc. Natl. Acad. Sci. USA 77:4216 (1980)); мышиные клетки Сертоли (ТM4, Mather, Biol. Reprod. 23:243-251 (1980)); клетки почек обезьяны (CV1 ATCC CCL 70); клетки почек африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почек собак (MDCK, ATCC CCL 34); клетки печени лабораторной крысы Buffalo (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (HepG2, HB 8065); опухолевые клетки мышиной молочной железы (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., Annal N.Y. Acad. Sci. 383:44-68 (1982)); клетки MRC5; клетки FS4 и клеточная линия человеческой гепатомы (HepG2). Клетки-хозяева трансформируют вышеописанными экспрессионными или клонирующими векторами для продуцирования антитела и культивируют в подходящих питательных средах, модифицированных, если это необходимо, для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих нужные последовательности. (viii) Культивирование клеток-хозяев Клетки-хозяева, используемые для продуцирования антитела согласно изобретению, могут быть культивированы в различных средах. Средами, подходящими для культивирования клеток-хозяев, являются коммерчески доступные среды, такие как среда Хэмса F10 (Sigma), минимальная поддерживающая среда ((МЕМ)(Sigma), RPMI-1640 (Sigma) и модифицированная по способу Дульбекко среда Игла (DMEM), Sigma). Кроме того, в качестве культуральной среды для культивирования клеток-хозяев может быть использована любая среда, описанная в публикации Ham et al., Meth. Enz. 58:44 (1979), Barnes et al., Anal. Biochem. 102:255 (1980), в патентах США (ix) Очистка антитела C применением техники рекомбинантных ДНК антитело может быть продуцировано внутри клеток либо оно может быть непосредственно секретировано в среду. Если в первой стадии антитело продуцируется внутри клеток, то затем клеточный дебрис, или клетки-хозяева, или их лизированные фрагменты удаляют, например, путем центрифугирования или ультрафильтрации. Если антитело секретируется в среду, то обычно сначала концентрируют супернатанты, полученные из таких экспрессионных систем, с использованием коммерчески доступного фильтра для концентрирования белков, например устройства для ультрафильтрации Amicon или Millipore Pellicon. Для ингибирования протеолиза в любой из предшествующих стадий может быть использован ингибитор протеазы, такой как PMSF, а для предупреждения размножения случайно вносимых примесных микроорганизмов могут быть использованы антибиотики. Композиция антитела, полученная из клеток, может быть очищена, например, с помощью хроматографии на гидроксиапатитах, гель-электрофореза, диализа и аффинной хроматографии, при этом предпочтительным методом очистки является аффинная хроматография. Пригодность белка А в качестве аффинного лиганда зависит от вида и изотипа любого Fc-домена иммуноглобулина, присутствующего в данном антителе. Белок А может быть использован для очистки антител, содержащих тяжелые цепи После проведения любой(ых) предварительной(ых) стадии(й) очистки смесь, содержащая представляющее интерес антитело и примеси, может быть подвергнута гидрофобной хроматографии при низком рН с использованием элюирующего буфера при рН примерно 2,5-4,5, предпочтительно, при низкой концентрации соли (например, примерно 0-0,25М). Анализы на активность Антитела согласно изобретению могут быть охарактеризованы по их физическим/химическим свойствам и биологическим функциям с помощью различных анализов, известных специалистам. Очищенные иммуноглобулины могут быть дополнительно охарактеризованы с помощью ряда анализов, включая, но не ограничиваясь ими, N-концевое секвенирование, аминокислотный анализ, эксклюзионная жидкостная хроматография высокого давления в неденатурирующих условиях (ЖХВД), масс-спектрометрия, ионообменная хроматография и гидролиз папаином. В некоторых вариантах изобретения полученные здесь иммуноглобулины анализируют на их биологическую активность. В некоторых вариантах изобретения иммуноглобулины согласно изобретению тестируют на их активность связывания с антигеном. Анализами на связывание с антигеном, которые известны специалистам и могут быть использованы в настоящем изобретении, являются, но не ограничиваются ими, любые анализы на прямое или конкурентное связывание с применением методов, таких как Вестерн-блот-анализ, радиоиммуноанализы, ELISA (твердофазный иммуноферментный анализ), В одном из своих вариантов настоящее изобретение относится к модифицированному антителу, обладающему несколькими, но не всеми эффекторными функциями, что делает его предпочтительным кандидатом для многих применений, в которых время полужизни антител in vivo является важным фактором, особенно в тех случаях, когда некоторые эффекторные функции (такие как, комплемент-зависимая и антитело-зависимая клеточная цитотоксичность (ADCC)) не имеют важного значения или даже являются нежелательными. В некоторых вариантах изобретения для гарантии того, что будут сохраняться только предпочтительные свойства иммуноглобулина, оценивают Fc-активность продуцируемого иммуноглобулина. Анализы на in vitro и/или in vivo цитотоксичность могут быть проведены для подтверждения уменьшения/элиминации CDС- и/или ADCC-активности. Так, например, для гарантии того, что антитело не будет связываться с Fc Гуманизированные антитела Настоящее изобретение относится к гуманизированным антителам. Различные методы гуманизации не-человеческих антител известны специалистам. Так, например, гуманизированное антитело может иметь один или несколько введенных в него аминокислотных остатков, происходящих от источника, не являющегося человеком. Эти не-человеческие аминокислотные остатки часто называют Выбор вариабельных доменов как легкой, так и тяжелой цепи человеческого антитела, используемых при создании гуманизированных антител, является очень важным фактором для снижения антигенности. В соответствии с так называемым методом Важно также, чтобы антитела были гуманизированы с сохранением высокой аффинности к данному антигену, а также других желательных биологических свойств. Для достижения этой цели в соответствии с одним из методов гуманизированные антитела получают путем анализа исходных последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей родительских и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов являются общедоступными и хорошо известны специалистам. Существуют компьютерные программы, которые иллюстрируют и представляют вероятные трехмерные конформационные структуры отобранных последовательностей-кандидатов иммуноглобулинов. Исследование этих представлений позволяет проанализировать вероятную роль данных остатков в функционировании последовательностей иммуноглобулина-кандидата, то есть остатков, которые влияют на способность иммуноглобулина-кандидата связываться с его антигеном. Таким образом, остатки FR могут быть выбраны из реципиентных и Варианты антител В одном из своих аспектов настоящее изобретение относится к фрагменту антитела, содержащему модификации в пограничной области полипептидов Fc, составляющих Fc-область, где указанные модификации облегчают и/или стимулируют гетеродимеризацию. Такие модификации включают введение В некоторых вариантах изобретения рассматриваются описанные здесь модификации аминокислотной последовательности указанных антител. Так, например, может оказаться желательным повысить аффинность связывания и/или другие биологические свойства антитела. Варианты аминокислотной последовательности антитела получают путем введения соответствующих нуклеотидных замен в нуклеиновую кислоту, кодирующую антитело, или путем пептидного синтеза. Такими модификациями, являются, например, делеции, и/или инсерции, и/или замены остатков в аминокислотных последовательностях антитела. При этом в конечную конструкцию могут быть введены любые комбинации делеций, инсерций и замен, при условии, что такая конечная конструкция будет обладать нужными свойствами. Аминокислотные модификации могут быть введены в рассматриваемую аминокислотную последовательность антитела в процессе получения такой последовательности. Метод, применяемый для идентификации некоторых остатков или областей антитела, которые имеют предпочтительную локализацию для мутагенеза, называется Вставками в аминокислотные последовательности являются вставки у амино- и/или карбокси-концов, образующие гибриды, имеющие длину от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки внутри последовательности, состоящие из одного или множества аминокислотных остатков. Примерами концевых инсерций является антитело с N-концевым метионильным остатком или антитело, присоединенное к цитотоксическому полипептиду. Другими инсерционными вариантами молекулы антитела являются гибриды, например, N- или С-концов антитела с ферментом (например, для ADEPT) или полипептидом, который увеличивает время полужизни антитела в сыворотке. Вариантом другого типа является вариант с заменой аминокислоты. Эти варианты имеют, по меньшей мере, один аминокислотный остаток в молекуле антитела, замененный другим остатком. При этом считается, что сайтами, представляющими наибольший интерес для мутагенеза с заменами, являются гипервариабельные области, однако, рассматриваются также и модификации в FR. Консервативные замены представлены в таблице 2 под заголовком
Значительные изменения биологических свойств антитела могут быть достигнуты путем выбора замен, которые значительно отличаются по своему влиянию (а) на структуру полипептидного остова в области замены, например, складчатую или спиральную конформацию, (b) на заряд или гидрофобность молекулы в нужном сайте, или (с) на объем боковой цепи. По общим свойствам своих боковых цепей аминокислоты подразделяются на нижеследующие группы (как описано в публикации A.L. Lehninger, Biochemistry, second ed., pp.73-75, Worth Publishers, New York (1975)): (1) неполярные: Ala (А), Val (V), Leu (L), Ile (I), Pro (Р), Phe (F), Trp (W), Met (М); (2) незаряженные полярные: Gly (G), Ser (S), Thr (Т), Сys (С), Tyr (Y), Asn (N), Gln (Q); (3) кислотные: Asp (D), Glu (Е); (4) основные: Lys (К), Arg (R), His (Н). Альтернативно по общим свойствам боковых цепей природные остатки могут быть подразделены на следующие группы: (1) гидрофобные остатки: норлейцин, Met, Ala, Val, Leu, Ile; (2) нейтральные гидрофильные остатки: Cys, Ser, Thr, Asn, Gln; (3) кислотные остатки: Asp, Glu; (4) основные остатки: His, Lys, Arg; (5) остатки, которые влияют на ориентацию цепи: Gly, Pro; (6) ароматические остатки: Trp, Tyr, Phe. Не-консервативные замены предусматривают замены аминокислоты одного класса аминокислотой другого класса. Такие замены остатков могут быть также введены в консервативные сайты, или, более предпочтительно, в другие остальные (не-консервативные) сайты. Одним из типов вариантов замен являются замены одного или нескольких остатков гипервариабельной области родительского антитела (например, гуманизированного или человеческого антитела). В общих чертах полученный(е) вариант(ы), выбранный(е) для дальнейшего исследования, должен(ны) иметь улучшенные биологические свойства по сравнению со свойствами родительского антитела, от которого он(они) происходит(ят). Стандартным методом генерирования таких вариантов замен является аффинное созревание с применением фагового представления. Вкратце, в несколько сайтов гипервариабельных областей (например, 6-7 сайтов) вносят мутации с получением всех возможных аминокислотных замен в каждом сайте. Полученные таким образом антитела представляют на частицах нитчатых фагов в виде гибридов с продуктом гена III М13, упакованным в каждую частицу. Затем представленные на фаге варианты скринируют на их биологическую активность (например, аффинность связывания), как описано в настоящей заявке. Для идентификации сайтов гипервариабельных областей, которые являются кандидатами на модификацию, может быть проведен аланин-сканирующий мутагенез, позволяющий идентифицировать остатки гипервариабельной области, которые играют значительную роль в связывании с антигеном. Альтернативно или дополнительно может оказаться полезным проанализировать кристаллическую структуру комплекса Молекулы нуклеиновой кислоты, кодирующие варианты аминокислотной последовательности антитела, получают различными методами, известными специалистам. Такими методами являются, но не ограничиваются ими, выделение из природного источника (в случае природных вариантов аминокислотной последовательности) или олигонуклеотид-опосредуемый (или сайт-направленный) мутагенез, ПЦР-мутагенез и кластерный мутагенез ранее полученного варианта или немодифицированного варианта указанного антитела. Может оказаться желательным введение одной или нескольких аминокислотных модификаций в Fc-область полипептидов иммуноглобулина согласно изобретению, и тем самым, создание варианта Fc-области. Такой вариант Fc-области может содержать человеческую последовательность Fc-области (например, Fc-области человеческого IgG1, IgG2, IgG3 или IgG4), имеющий аминокислотную модификацию (например, замену) в одном или в нескольких положениях аминокислот, включая положения цистеина шарнирной области. В соответствии с настоящим описанием и известным уровнем техники в некоторых вариантах изобретения предусматривается, что антитело, используемое в способах согласно изобретению, может содержать одну или несколько модификаций по сравнению с аналогом этого антитела дикого типа, например, в Fc-области. Эти антитела, тем не менее, сохраняют почти все свойства, необходимые для достижения терапевтической эффективности по сравнению со свойствами аналога дикого типа. Так, например, считается, что в Fc-области могут быть сделаны некоторые модификации, которые будут изменять (то есть, либо увеличивать, либо уменьшать) уровень связывания с С1q и/или комплемент-зависимую цитотоксичность (CDС), например, как описано в WO 99/51642 (см. также публикацию Duncan & Winter, Nature 322:738-40 (1988); патент США Иммуноконъюгаты Настоящее изобретение также относится к иммуноконъюгатам или конъюгатам Использование конъюгатов Зевалин® (ибритумомаб тиуксетан, Biogen/Idec) представляет собой конъюгат Химиотерапевтические средства, используемые для получения таких иммуноконъюгатов, описаны выше. Ферментативно активными токсинами и их фрагментами, которые могут быть использованы для этих целей, являются А-цепь дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, А-цепь экзотоксина (от Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модецина, альфа-сарцин, белки Aleurites fordii, белка диантина, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены. Для продуцирования радиоактивно конъюгированных антител могут быть использованы различные радионуклиды. Примерами таких радионуклидов являются 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела и цитотоксического лекарственного средства получают с использованием различных бифункциональных белок-связывающих агентов, таких как N-сукцинимидил-3-(2-пиридилтиол)пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат-HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Так, например, иммунотоксин рицин может быть получен как описано в публикации Vitetta et al. Science, 238:1098 (1987). 14С-меченная 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (МХ-DTPA) является репрезентативным хелатообразующим агентом для конъюгирования радионуклида с антителом (см. WO 94/11026). В настоящем изобретении также рассматриваются конъюгаты антитела и одного или нескольких низкомолекулярных токсинов, таких как калихеамицин, майтанзиноиды, трихотецен и СС1065, и производные этих токсинов, обладающие токсической активностью. Майтанзин и майтанзиноиды В одном из вариантов изобретения антитело (полноразмерное антитело или его фрагменты) согласно изобретению конъюгировано с одной или несколькими молекулами майтанзиноида. Майтанзиноиды представляют собой митотические ингибиторы, которые действуют посредством ингибирования полимеризации тубулина. Майтанзин был впервые выделен у восточно-африканской землеройки Maytenus serrata (патент США Конъюгаты В целях увеличения терапевтического индекса майтанзина и майтанзиноидов эти соединения были конъюгированы с антителами, специфически связывающимися с антигенами опухолевых клеток. Иммуноконъюгаты, содержащие майтанзиноиды, и их терапевтическое применение описаны, например, в патентах США Конъюгаты Конъюгаты Для создания конъюгатов Конъюгаты Линкер может быть присоединен к молекуле майтанзиноида в различных положениях в зависимости от типа связи. Так, например, сложноэфирная связь может быть образована посредством реакции с гидроксильной группой стандартными методами связывания. Такая реакция может осуществляться в положении С-3, имеющем гидроксильную группу, в положении С-14, модифицированном гидроксиметилом, в положении С-15, модифицированном гидроксильной группой, и в положении С-20, имеющем гидроксильную группу. В предпочтительном варианте изобретения указанная связь образуется в положении С-3 майтанзинола или его аналога. Калихеамицин Другой представляющий интерес иммуноконъюгат содержит антитело, конъюгированное с одной или несколькими молекулами калихеамицина. Антибиотики семейства калихеамицинов способны продуцировать двухцепочечные ДНК-разрывы в субпикомолярных концентрациях. Описание получения конъюгатов семейства калихеамицинов можно найти в патентах США Другие цитотоксические средства Другими противоопухолевыми средствами, которые могут быть конъюгированы с антителами согласно изобретению, являются ВСNU, стрептозоцин, винкристин и 5-фторурацил, семейство агентов, известных под общим названием комплекс LL-Е33288, описанный в патентах США Ферментативно активными токсинами и их фрагментами, которые могут быть использованы в этих целях, являются А-цепь дифтерийного токсина, несвязывающиеся активные фрагменты дифтерийного токсина, А-цепь экзотоксина (от Pseudomonas aeruginosa), А-цепь рицина, А-цепь абрина, А-цепь модецина, альфа-сарцин, белки Aleurites fordii, белки диантина, белки Phytolaca americana (PAPI, PAPII и PAP-S), ингибитор Momordica charantia, курцин, кротин, ингибитор Sapaonaria officinalis, гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотецены (см., например, заявку WO 93/21232, опубл.28 октября 1993). В настоящем изобретении также рассматривается иммуноконъюгат, образующийся между антителом и соединением, имеющим нуклеолитическую активность (например, рибонуклеазой или ДНК-эндонуклеазой, такой как дезоксирибонуклеаза; ДНКазой). Для селективной деструкции опухоли может быть использовано антитело, которое может содержать высокорадиоактивный атом. Для продуцирования радиоактивно конъюгированных антител могут быть использованы различные радиоактивные изотопы. Примерами таких радионуклидов являются 211At, 131I, 125I, 90Y, 186Re, 188Re, 153Sm, 212Bi, 32Р, 212Рb и радиоактивные изотопы Lu. Если указанный конъюгат используется для детекции, то он может содержать радиоактивный атом для сцинтиграфических исследований, например 99mТс или 123I, или спин-метку для визуализации методом ядерного магнитного резонанса (ЯМР) (известной также как МР-визуализация, МРВ), такую как иод-123, иод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо. Радиоактивные или другие метки могут быть включены в конъюгат известными методами. Так, например, пептид может быть синтезирован биологическими методами либо он может быть синтезирован методом химического синтеза аминокислот с использованием подходящих аминокислотных предшественников, включающих, например, фтор-19 вместо водорода. Такие метки, как 99mТс или 123I, 186Re, 188Re и 111In могут быть присоединены посредством цистеинового остатка пептида. Иттрий-90 может быть присоединен посредством лизинового остатка. Для введения иода-123 может быть применен метод IODOGEN (Fraker et al. (1978) Biochem. Biophys. Res. Commun. 80:49-57). Другие методы подробно описаны в публикации Конъюгаты антитела и цитотоксического лекарственного средства могут быть получены с использованием различных бифункциональных белок-связывающих агентов, таких как N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP), сукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат, иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметиладипимидат-HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис(п-азидобензоил)гександиамин), производные бис-диазония (такие как бис-(п-диазонийбензоил)этилендиамин), диизоцианаты (такие как толуол-2,6-диизоцианат) и бис-активные соединения фтора (такие как 1,5-дифтор-2,4-динитробензол). Так, например, иммунотоксин рицин может быть получен, как описано в публикации Vitetta et al. Science, 238:1098 (1987). 14С-меченная 1-изотиоцианатобензил-3-метилдиэтилентриаминпентауксусная кислота (МХ-DTPA) является репрезентативным хелатообразующим агентом для конъюгирования радионуклида с антителом (см. WO 94/11026). Линкером может быть Соединениями, специально рассматриваемыми в настоящем изобретении, являются, но не ограничиваются ими, ADC, полученные с использованием перекрестно-связывающих реагентов, таких как BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, MPBH, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо-SIAB, сульфо-SMCC, сульфо-SMPB и SVSB (сукцинимидил-(4-винилсульфон)бензоат), которые являются коммерчески доступными (например, поставляются фирмой Pierce Biotechnology, Inc., Rockford, IL., USA). См. руководство по применению и каталог (Application Handbook and Catalog), 2003-2004, стр. 467-498. Получение конъюгатов В конъюгатах Ab – (L – D)р. (I) Нуклеофильными группами, присутствующими на антителах, являются, но не ограничиваются ими: (i) N-концевые аминогруппы, (ii) аминогруппы боковой цепи, например лизина, (iii) тиоловые группы боковой цепи, например цистеина, и (iv) гидроксильные или аминогруппы сахаров, где указанное антитело является гликозилированным. Аминогруппы, тиольные группы и гидроксильные группы являются нуклеофильными и могут подвергаться реакции взаимодействия с образованием ковалентных связей с нуклеофильными группами на линкерных молекулах или линкерных реагентах, включая: (i) активные сложные эфиры, такие как NНS-эфиры, HOBt-эфиры, галогенформиаты и галегенангидриды кислот; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы. Некоторые антитела имеют восстанавливаемые межцепьевые дисульфиды, то есть цистеиновые мостики. Для конъюгирования с линкерными реагентами антитела могут быть сделаны реакционноспособными путем их обработки восстановителем, таким как DТТ (дитиотреитол). Каждый цистеиновый мостик, теоретически, может образовывать два реакционноспособных тиоловых нуклеофила. В антитела могут быть введены дополнительные нуклеофильные группы посредством реакции взаимодействия лизинов с 2-иминотиоланом (реагентом Траута), что будет приводить к превращению амина в тиол. Конъюгаты Аналогичным образом нуклеофильными группами на молекуле лекарственного средства являются, но не ограничиваются ими, аминогруппы, тиольные, гидроксильные, гидразидные, оксимовые, гидразиновые, тиосемикарбазоновые, гидразинкарбоксилатные и арилгидразидные группы, способные реагировать с образованием ковалентных связей с электрофильными группами на линкерных молекулах и линкерных реагентах, включая: (i) активные сложные эфиры, такие как NНS-эфиры, HOBt-эфиры, галогенформиаты и галегенангидриды кислот; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидные группы. Альтернативно гибридный белок, содержащий антитело и цитотоксическое средство, может быть получен, например, рекомбинантными методами или методом пептидного синтеза. Длина ДНК может составлять соответствующие области, кодирующие две части конъюгата, либо смежные друг с другом, либо разделенные областью, кодирующей линкерный пептид, который не оказывает негативного влияния на желаемые свойства данного конъюгата. В еще одном варианте изобретения указанное антитело может быть конъюгировано с Производные антител Антитела согласно изобретению могут быть дополнительно модифицированы так, чтобы они содержали другие не-белковые молекулы, которые известны специалистам и являются легко доступными. Предпочтительными молекулами, подходящими для дериватизации антител, являются водорастворимые полимеры. Неограничивающими примерами водорастворимых полимеров являются полиэтиленгликоль (ПЭГ), сополимеры этиленгликоля/пропиленгликоля, карбоксиметилцеллюлоза, декстран, поливиниловый спирт, поливинилпирролидон, поли-1,3-диоксолан, поли-1,3,6-триоксан, сополимер этилена/малеинового ангидрида, полиаминокислоты (гомополимеры или статистические сополимеры) и декстран или поли(н-винилпирролидон)полиэтиленгликоль, гомополимеры пропиленгликоля, сополимеры полипропиленоксида/этиленоксида, полиоксиэтилированные полиолы (например, глицерин), поливиниловый спирт и их смеси. При промышленном изготовлении следует отдать предпочтение пропиональдегиду полиэтиленгликоля благодаря его стабильности в воде. Этот полимер может иметь любую молекулярную массу и может быть разветвленным или неразветвленным. Число полимеров, присоединяемых к антителу, может варьироваться, и если присоединяют более чем один полимер, то эти полимеры могут иметь одинаковые или различные молекулы. В общих чертах число и/или тип полимеров, используемых для дериватизации, могут быть выбраны исходя из нескольких факторов, включая, но не ограничиваясь ими, конкретные свойства или функции дериватизируемого антитела, независимо от конкретных условий терапии, при которых будет использоваться указанное производное антитела и т.п. Фармацевтические композиции Терапевтические композиции, содержащие антитело согласно изобретению, получают в виде препаратов, удобных для хранения, путем смешивания антитела, имеющего нужную степень чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington’s Pharmaceutical Sciences, 16th edition, Osol, A. Ed. (1980)), в форме водных растворов, лиофилизованных препаратов или других сухих препаратов. Приемлемые носители, эксципиенты или стабилизаторы в используемых дозах и концентрациях должны быть нетоксичными для реципиентов, и ими являются буферы, такие как фосфат, цитрат, гистидин и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензэтония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные полипептиды (имеющие примерно менее чем 10 остатков); белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатообразующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; металлокомплексы (например, комплексы Zn – белок); и/или неионогенные поверхностно-активные вещества, такие как твинТМ, плюроникТМ или полиэтиленгликоль (ПЭГ). Описанная здесь композиция может также содержать несколько активных соединений, необходимых для конкретного показания, а предпочтительно, имеющих дополнительные активности, не оказывающие негативного влияния друг на друга. Такие молекулы могут присутствовать в комбинации в количествах, эффективных для применения в нужных целях. Активные ингредиенты могут быть также заключены в микрокапсулу, полученную, например, методами коацервации или путем межфазной полимеризации, например, в гидроксиметилцеллюлозную или желатиновую микрокапсулу и полиметилметакрилатную микрокапсулу, соответственно, в системы для доставки коллоидальных лекарственных средств (например, в липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такая методика описана в руководстве Remington’s Pharmaceutical Sciences, 16th edition, Osol, A. Ed. (1980). Композиции, используемые для введения in vivo, должны быть стерильными. Это может быть легко достигнуто путем фильтрации через стерильные фильтрующие мембраны. Могут быть получены препараты замедленного высвобождения. Подходящими примерами препаратов замедленного высвобождения являются полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих иммуноглобулин согласно изобретению, где указанные матрицы имеют форму готовых изделий, например пленок или микрокапсул. Примерами матриц замедленного высвобождения являются полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поливиниловый спирт)), полилактиды (патент США Применения Антитело согласно изобретению может быть использовано, например, в терапевтических методах in vitro, ex vivo и in vivo. Антитела согласно изобретению могут быть использованы в качестве антагониста для частичного или полного блокирования специфической антигенной активности in vitro, ex vivo и/или in vivo. Кроме того, по меньшей мере, некоторые антитела согласно изобретению могут нейтрализовать активность антигена, происходящего от других видов. В соответствии с этим антитела согласно изобретению могут быть использованы для ингибирования специфической антигенной активности, например, в клеточной культуре, содержащей указанный антиген, в организме человека или других млекопитающих, имеющих антиген, с которым перекрестно реагирует антитело согласно изобретению (например, шимпанзе, павианов, игрунок, собакоподобных обезьян и макак-резусов, а также свиней или мышей). В одном из вариантов изобретения антитело согласно изобретению может быть использовано для ингибирования антигенной активности путем контактирования указанного антитела с антигеном так, чтобы осуществлялось ингибирование активности антигена. Предпочтительным антигеном является человеческая белковая молекула. В одном из вариантов изобретения антитело согласно изобретению может быть использовано в способе ингибирования антигена у индивидуума, страдающего расстройством, при котором антигенная активность оказывает негативное воздействие, где указанный способ включает введение данному индивидууму антитела согласно изобретению для ингибирования указанной антигенной активности у данного индивидуума. Предпочтительным антигеном является человеческая белковая молекула, а предпочтительным индивидуумом является человек. Альтернативно указанным индивидуумом может быть млекопитающее, экспрессирующее антиген, с которым связывается антитело согласно изобретению. Кроме того, указанным индивидуумом может быть млекопитающее, которому был введен указанный антиген (например, путем прямого введения антигена или экспрессии антигенного трансгена). Антитело согласно изобретению может быть введено человеку в терапевтических целях. Кроме того, антитело согласно изобретению может быть введено млекопитающему, не являющемуся человеком (например, примату, свинье или мыши) и экспрессирующему антиген, с которым перекрестно реагирует иммуноглобулин, в целях лечения этого млекопитающего либо оно может быть введено животному с моделью человеческого заболевания. Что касается животного-модели, то такие животные могут быть использованы для оценки терапевтической эффективности антител согласно изобретению (например, для определения доз и времени введения). Блокирующими терапевтически эффективными антителами согласно изобретению являются, например, но не ограничиваются ими, анти-HER2 антитело, анти-VEGF антитело, анти-IgЕ антитело, анти-CD11 антитело, антитело против интерферона и антитело против тканевого фактора. Антитела согласно изобретению могут быть использованы для лечения, ингибирования, замедления прогрессирования, предупреждения/замедления возникновения рецидивов, улучшения динамики или предупреждения заболеваний, расстройств или состояний, ассоциированных с аномальной экспрессией и/или активностью одной или нескольких молекул антигена, включая, но не ограничиваясь ими, злокачественные и доброкачественные опухоли; злокачественные заболевания, не относящиеся к лейкозу и лимфолейкозу; патологии нервных клеток, глиальных клеток, астроцитов, клеток гипоталамуса и другие гландулярные, макрофагальные, эпителиальные, стромальные и бластоцелиальные расстройства; а также воспалительные, ангиогенные и иммунологические расстройства. В одном из аспектов изобретения блокирующее антитело согласно изобретению является специфичным к антигену-лиганду и ингибирует антигенную активность путем блокирования или предотвращения взаимодействия лиганда-рецептора с антигеном-лигандом, и тем самым, ингибирует соответствующий путь передачи сигнала и других молекулярных или клеточных процессов. Настоящее изобретение также относится к рецептор-специфическим антителам, которые должны, но необязательно, предотвращать связывание с лигандом и подавлять активацию рецептора, что приводит к ингибированию любых ответов, которые в нормальных условиях должны инициироваться посредством связывания с лигандом. Настоящее изобретение также относится к антителам, которые, предпочтительно, или исключительно, связываются с комплексами В некоторых вариантах изобретения иммуноконъюгат, содержащий антитело, конъюгированное с цитотоксическим агентом, вводят указанному пациенту. В некоторых вариантах изобретения иммуноконъюгат и/или антиген, с которым он связывается, интернализуется в клетке, что приводит к повышению терапевтической эффективности иммуноконъюгата, направленной на уничтожение клетки-мишени, с которой он связывается. В одном из вариантов изобретения цитотоксический агент подавляет нуклеиновую кислоту или взаимодействует с нуклеиновой кислотой в клетке-мишени. Примерами таких цитотоксических агентов являются любые описанные здесь химиотерапевтические средства (такие как майтанзиноид или калихеамицин), радиоактивный изотоп или рибонуклеаза, или ДНК-эндонуклеаза. Антитела согласно изобретению, взятые отдельно или в комбинации с другими композициями, могут быть использованы в терапевтических целях. Так, например, антитело согласно изобретению может быть введено вместе с другим антителом, химиотерапевтическим(и) средством(ами) (включая смеси химиотерапевтических средств), с другим(и) цитотоксическим(и) средством(ами), антиангиогенным(и) средством(ами), цитокинами и/или с рост-ингибирующим(и) средством(ами). Если антитело согласно изобретению ингибирует рост опухоли, то может оказаться особенно желательным его объединение с одним или несколькими другими терапевтическими средствами, также ингибирующими рост опухоли. Так, например, антитело согласно изобретению может быть объединено с анти-VEGF антителом (например, авастином) и/или с анти-ЕrbB антителами (например, с анти-HER2 антителом, герцептином®) для его применения в схеме лечения, например, любых описанных здесь заболеваний, включая рак ободочной кишки, рак молочной железы с метастазами и рак почек. Альтернативно или дополнительно пациент может быть подвергнут комбинированной лучевой терапии (например, облучением внешним пучком или терапии с использованием радиоактивно меченного агента, такого как антитело). Такой указанной выше комбинированной терапией является комбинированное введение (где два или более агентов включены в одну или ту же или в отдельные композиции) и отдельное введение, при котором антитело согласно изобретению может быть введено до и/или после проведения вспомогательной терапии. Антитело согласно изобретению (и вспомогательное терапевтическое средство) вводят различными способами, включая парентеральное, подкожное, внутрибрюшинное, внутрилегочное и интраназальное введение, а если необходимо местное лечение, то антитело вводят в пораженный участок. Парентеральной инфузией является внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное или подкожное введение. Кроме того, подходящим способом введения является периодическое вливание, а в частности вливание антитела в уменьшающихся дозах. Такие дозы могут быть введены любым подходящим способом, например, путем инъекций, таких как внутривенные или подкожные инъекции, и эти дозы отчасти зависят от того, является ли такое введение периодическим или постоянным. Антитело-содержащую композицию согласно изобретению приготавливают, разделяют на дозы и вводят в соответствии с хорошо известной медицинской практикой. Факторами, учитываемыми для приготовления таких композиций, являются конкретное расстройство, подвергаемое лечению, конкретное млекопитающее, подвергаемое лечению, клиническое состояние отдельного пациента, этиологический фактор, вызывающий данное расстройство, участок, на который направлено данное средство, способ введения, схема введения и другие факторы, известные практическим врачам. Указанное антитело должно быть, но необязательно, приготовлено в виде композиции с одним или несколькими средствами, используемыми для предупреждения или лечения рассматриваемого расстройства. Эффективное количество таких других средств зависит от количества антител согласно изобретению, присутствующих в данной композиции, типа расстройства или способа лечения и от других факторов, обсуждаемых выше. Указанные другие средства обычно вводятся в тех же самых дозах и теми же способами, которые обычно использовались ранее, либо эти дозы составляют примерно 1-99% от используемых ранее доз. Для предупреждения или лечения заболеваний подходящие дозы антитела согласно изобретению (используемого отдельно или в комбинации с другими средствами, такими как химиотерапевтические средства) зависят от типа заболевания, подвергаемого лечению, типа антитела, тяжести и курса лечения заболевания, независимо от того вводят ли указанное антитело в профилактических или терапевтических целях, от проводимого ранее лечения, от истории болезни пациента и его восприимчивости к антителу и от назначения лечащего врача. Такое антитело может быть введено пациенту один раз или более во время проведения курса лечения. В зависимости от типа и тяжести заболевания начальная предварительно установленная доза антитела, вводимого пациенту, составляет примерно от 1 мкг/кг до 15 мг/кг (например, 0,1 мг/кг – 10 мг/кг), независимо от того, осуществляют ли одноразовое или многократное введение или непрерывную инфузию данного антитела. Типичная суточная доза может составлять примерно от 1 мкг/кг до 100 мг/кг или более в зависимости от вышеупомянутых факторов. Для повторного введения в течение нескольких дней или более, в зависимости от состояния, лечение может проводиться до тех пор, пока не будет достигнуто желаемое подавление симптомов заболевания. Репрезентативная доза антитела может составлять примерно от 0,05 до 10 мг/кг. Так, например, пациенту могут быть введены одна или несколько доз, составляющих примерно 0,5 мг/кг, 2,0 мг/кг, 4,0 мг/кг или 10 мг/кг (или любая их комбинация). Такие дозы могут быть введены периодически, например, еженедельно или раз в три недели (например, пациенту может быть введено от двух до двадцати, например, примерно шесть доз антитела). При этом сначала могут быть введены более высокие дозы, а затем может быть введена одна или несколько более низких доз. Репрезентативная схема введения доз включает введение начальной дозы, составляющей примерно 4 мг/кг, с последующим еженедельным введением поддерживающей дозы, составляющей примерно 2 мг/кг антитела. Однако могут быть применены и другие схемы введения. Мониторинг эффекта такой терапии может быть легко проведен стандартными методами и с применением стандартных анализов. Промышленные изделия В другом своем аспекте настоящее изобретение относится к промышленному изделию, содержащему материалы, используемые для лечения, предупреждения и/или диагностики описанных выше расстройств. Промышленное изделие содержит контейнер и этикетку или вкладыш, вложенный в упаковку или наклеенный на контейнер. Подходящими контейнерами являются, например, бутыли, флаконы, шприцы и т.п. Контейнеры могут быть изготовлены из различных материалов, таких как стекло или пластик. Этот контейнер содержит композицию, которая сама по себе или в комбинации с другой композицией является эффективной для лечения, предупреждения и/или диагностики данного состояния и может иметь стерильное входное отверстие (например, таким контейнером может быть пакет для внутривенного введения раствора или сосуд, имеющий пробку, протыкаемую иглой для подкожных инъекций). В данной композиции, по меньшей мере, одним активным агентом является антитело согласно изобретению. На этикетке или во вкладыше, вложенном в упаковку, указано, что такая композиция используется для лечения данного конкретного заболевания, такого как рак. Кроме того, указанное промышленное изделие может включать (а) первый контейнер с содержащейся в нем композицией, включающей антитело согласно изобретению, и (b) второй контейнер с содержащейся в нем композицией, включающей дополнительное цитотоксическое средство. В этом варианте изобретения промышленное изделие согласно изобретению может дополнительно содержать вложенный в упаковку вкладыш, в котором указано, что композиции, содержащие первое и второе антитело, могут быть использованы для лечения конкретного состояния, например рака. Альтернативно или дополнительно указанное промышленное изделие может дополнительно содержать второй (или третий) контейнер, включающий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), забуференный фосфатом физиологический раствор, раствор Рингера и раствор декстрозы. Кроме того, оно может включать и другие материалы, необходимые для коммерческого производства и для потребителя, включая другие буферы, разбавители, фильтры, иглы и шприцы. Ниже приводятся примеры способов и композиций согласно изобретению. При этом следует отметить, что исходя из приведенного выше общего описания изобретения могут быть осуществлены и другие различные варианты изобретения. ПРИМЕРЫ Материалы и методы Нумерацию остатков проводили по Кэбату (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed., Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). При этом были использованы однобуквенные обозначения аминокислот. Вырожденность ДНК представлена кодом IUB (N = A/C/G/T, D = A/G/T, V = A/C/G, B = C/G/T, H = A/C/T, K = G/T, M = A/C, R = A/G, S = G/C, W = A/T, Y = C/T). Прямое присоединение гипервариабельной области к акцепторной человеческой консенсусной каркасной области. Фагмидой, используемой в данной работе, является вектор для представления моновалентного Fab-g3 (pV0350-2В), имеющий две открытые рамки считывания под контролем промотора phoA, и описанный, по существу, в работе Lee et al., J. Mol. Biol. (2004) 340(5):1073-93. Первая открытая рамка считывания состоит из сигнальной последовательности stII, присоединенной к доменам VL и СН1 акцепторной легкой цепи, а вторая открытая рамка считывания состоит из сигнальной последовательности stII, присоединенной к доменам VН и СН1 акцепторной тяжелой цепи, за которой следует усеченная последовательность оболочечного белка малого фага Р3. Lee et al., см. выше. VL- и VH-домены мышиного антитела 5D5 (гибридома 5D5.11.6, АТСС, рег. Варианты, полученные путем прямого присоединения, были генерированы с помощью мутагенеза по Кункелю с использованием отдельных олигонуклеотидов для каждой гипервариабельной области. Нужные клоны оценивали путем секвенирования ДНК.
Частично рандомизированные олигонуклеотиды были расположены после последовательностей мышиной гипервариабельной области и охватывали те же самые области, которые были идентифицированы в результате прямого присоединения гипервариабельной области. Набор аминокислот, присутствующих в начале Н2 (в положении 49) в VH-домене, был ограничен аминокислотами А, G, S или Т, что достигалось с использованием кодона RGC. Создание фаговых библиотек. Пулы рандомизированных олигонуклеотидов, сконструированных для каждой гипервариабельной области, были отдельно фосфорилированы в шести реакционных смесях по 20 мкл, содержащих 660 нг олигонуклеотида, 50 мМ триса, рН 7,5, 10 мМ MgCl2, 1 мМ АТФ, 20 мМ ДТТ и 5 ед. полинуклеотид-киназы в течение 1 часа при 37оС. Затем эти шесть фосфорилированных олигонуклеотидных пула объединяли с 20 мкг матрицы Кункеля в 50 мМ триса, рН 7,5, 10 мМ MgCl2, в конечном объеме 500 мкл с получением отношения Достроенный продукт анализировали на смеси трис-ацетат-EDTA/агарозных гелях (Sidhu et al., Methods in Enzymology 328:333-363 (2000)). В основном наблюдались три полосы: нижняя полоса соответствовала Затем достроенный продукт очищали и подвергали электропорации в клетках SS320, после чего их размножали в присутствии хелперного фага М13/КО7, как описано в публикации Sidhu et al., Methods in Enzymology 328:333-363 (2000). Размеры библиотек составляли 1-2 х 109 независимых клонов. Произвольно выбранные клоны, полученные из исходных библиотек, секвенировали для оценки качества библиотеки. Фаговый отбор. Был получен человеческий рецептор HGF, который использовали в форме Fc-гибрида (HGFR-Fc) (Mark et al., J. Biol. Chem. (1992) 267:26166-26171). HGFR-Fc наносили на микротитрационные планшеты MaxiSorp (Nunc) при 5 мкг/мл в PBS. Для первого раунда отбора использовали восемь лунок с мишенями, а одну лунку с мишенью использовали для последующих раундов отбора. Лунки блокировали в течение 1 часа с использованием казеинового ингибитора (Pierce). Фаг выделяли из супернатанта культуры и суспендировали в PBS, содержащем 1% ВSA и 0,05% Твина-20 (PBSВТ). После связывания с лунками в течение 2 часов несвязанный фаг удаляли путем интенсивного промывания буфером PBS, содержащим 0,05% Твина-20 (PBSТ). Связанный фаг элюировали путем инкубирования лунок с 50 мМ HCl, 0,5 М КСl в течение 30 минут. Фаг амплифицировали с использованием клеток Тор10 и хелперного фага М13/КО7, а затем культивировали в течение ночи при 37оС в среде 2YT, содержащей 50 мкг/мл карбанециллина. Для оценки степени обогащения титры фага, элюированного из лунок, покрытых мишенями, сравнивали с титрами фага, выделенного из лунки, не покрытой мишенями. Для созревания аффинности фаговые библиотеки сортировали методом сортировки в растворе. HGFR-Fc биотинилировали путем смешивания 500 мкл 3,6 мг/мл HGFR-Fc в PBS и 10 мкл 1 М фосфата калия, рН 8, с 20 мкл 4 мМ сульфо-NHS-LС-биотина (Pierce). Биотинилированный HGFR-Fc (b-HGFR-Fc) очищали на колонке с NАР5 (Amersham Biosciences) в PBS. Лунки микротитрационного планшета сенсибилизировали 10 мкг/мл нейтравидина в PBS в течение ночи при 4°С, а затем блокировали в течение 1 часа казеиновым ингибитором (Pierce). В первом раунде пэннинга 200 мкл фага, суспендированного в PBS, содержащем 0,05% Твина-20 (PBSТ) и 1% ВSA, смешивали с 10 нМ b-HGFR-Fc в течение 1 часа. Фаг, связанный с b-HGFR-Fc, иммобилизовали на лунках, покрытых нейтравидином, в течение 10 минут, а несвязанный фаг удаляли путем промывки буфером PBSТ. Фаг элюировали с использованием 20 мМ HCl, 500 мМ КСl в течение 45 минут, затем нейтрализовали и размножали в клетках ХL1blue (Stratagene) в присутствии хелперного фага КО7 (New England Biolabs). Последующие раунды сортировки проводили аналогичным образом, за некоторыми исключениями, а именно: в раунде 2 конечная концентрация b-HGFR-Fc составляла 5,6 нМ; в раунде 3 конечная концентрация b-HGFR-Fc составляла 0,1 нМ; в раунде 4 конечная концентрация b-HGFR-Fc составляла 0,5 нМ; а для иммобилизации на нейтравидине к смеси в течение 1 часа добавляли 780 нМ небиотинилированного HGFR-Fc. ELISA фага. Микротитрационные планшеты MaxiSorp сенсибилизировали человеческим HGFR-Fc при 5 мкг/мл в PBS в течение ночи, а затем блокировали казеиновым ингибитором. Фаг, полученный из супернатантов культуры, инкубировали с серийно разведенным HGFR-Fc в PBSТ, содержащем 1% ВSA, в микротитрационном планшете для культивирования тканей в течение 1 часа, а затем 80 мкл смеси переносили в лунки, покрытые мишенью, и оставляли на 15 минут для иммобилизации несвязанного фага. Планшет промывали PBSТ и добавляли ПХ-конъюгированное антитело против М13 (Amersham Pharmacia Biotech)(1:5000 в PBSТ, содержащем 1% ВSA) в течение 40 минут. Планшет промывали PBSТ и проявляли добавлением тетраметилбензидинового субстрата (Kirkegaard & Perry Laboratories, Gaithersburg, MD). Для определения IC50 строили кривую зависимости оптической плотности при 405 нм от концентрации мишени в растворе. Полученную величину использовали для оценки аффинности Fab-клона, представленного на поверхности фага. Продуцирование Fab и определение аффинности. Для экспрессии Fab-белка в целях определения аффинности между тяжелой цепью и g3 в векторе фагового представления встраивали стоп-кодон. Клоны трансформировали в клетки 34В8 E.coli и культивировали в среде АР5 при 30°С (Presta et al. Cancer Res. 57:4593-4599 (1997)). Клетки собирали центрифугированием, суспендировали в 10 мМ триса, 1 мМ EDTA, рН 8 и разрушали с помощью микрофлюидизатора. Fab очищали с помощью аффинной хроматографии на G-белке. Определение аффинности осуществляли с помощью поверхностного плазмонного резонанса с использованием BIAcoreTM-2000. HGFR-Fc иммобилизовали (~1000 единиц отклика (RU)) на чипе СМ5 и вводили различные концентрации Fab (4-500 нМ) в PBSТ. После каждой инжекции чип регенерировали с использованием 100 мМ HCl. Реакцию на связывание корректировали путем вычитания RU из данных, полученных для контрольной проточной кюветы. Для анализа кинетики реакции использовали лангмуровскую модель связывания 1:1 при одновременном подборе kon и koff. Электрохемилюминесцентный анализ на блокирование связывания HGF/c-met антителом ОА5D5 Очищенный белок сМеt-Ig, полученный от Genentech (South San Francisco, CA), биотинилировали путем инкубирования с 20-кратным молярным избытком NHS-Х-биотина в 0,1 М NaНСО3, рН 8,5, с использованием биотин-Х-NHS (Research Organics, Cleveland, OH). Очищенный человеческий 2-цепочечный HGF, полученный от Genentech, метили BV-TAG (cat # 110034) химическим методом с использованием NHS-эфира в соответствии с инструкциями производителей (BioVeris International, Gaithersburg, MD). сМеt-Ig-биотин (500 нг/мл), HGF-рутениевую метку (250 нг/мл) и титры антитела ОА5D5 в концентрации от 3333 до 0,21 нМ инкубировали в аналитическом разбавителе в объеме 100 мкл: PBS + 0,5% ВSA/0,5% Твина-20/0,033% проклина (Supelco Inc. Bellefonte PA). Смеси инкубировали в герметично закрытых полипропиленовых круглодонных 96-луночных планшетах (Corning) в течение 2-4 часов при комнатной температуре и при встряхивании. Затем добавляли магнитные сферы, покрытые стрептавидином (Dynabeads, BioVeris). После окончательного 45-минутного инкубирования при интенсивном перемешивании планшеты считывали на устройстве BioVeris серии М (BioVeris International, Gaithersburg, MD). KIRA (HGF-зависимое фосфорилирование Met) Клетки А549 (АТСС, Manassas, VA) поддерживали в культуральной среде (среде F12 Хэмса/DMEM 50:50 [Gibco, Grand Island, NY], содержащей 10% фетальную телячью сыворотку (FBS, Sigma, St. Louis, MO). Для получения клеток в целях проведения анализа клетки выделяли из конфлюэнтных культур с использованием аккутазы (ICN, Aurora, OH) и высевали в 96-луночные планшеты при плотности 50000 клеток на лунку. После инкубирования в течение ночи при 37оС культуральную среду удаляли и клетки помещали на 30-60 минут в бессывороточную среду, содержащую 0,1% FBS. Для определения способности ОА-5D5 ингибировать фосфорилирование с-Met эту молекулу серийно разводили в концентрации от 200 до 0,19 нМ в среде с 0,1% FBS и добавляли в аналитические планшеты. После 15-минутного инкубирования при 37оС добавляли HGF (50 нг/мл). Затем планшеты инкубировали еще 10 минут при 37оС, среду удаляли и добавляли буфер для лизиса клеток (Cell Signaling Technologies, Cat.# 9803, Beverly, MA, в который была добавлена смесь ингибиторов протеазы, закупленная у фирмы Calbiochem, cat # 539131, San Diego, CA). Лизаты анализировали на фосфорилирование с-Met с помощью электрохемилюминесцентного анализа на аппарате BioVeris серии М (BioVeris International, Gaithersburg, MD). mAb против фосфотирозина (клон 4G10, Upstate, Lake Placid, MY) метили BV-TAG химическим методом с использованием NHS-эфира в соответствии с инструкциями производителей (BioVeris). Антитела против внеклеточного домена с-Met подвергали биотинилированию с использованием биотина-Х-NHS (Research Organics, Cleveland, OH). BV-TAG-меченное 4G10 и биотинилированое mAb против с-Met разводили в аналитическом буфере (PBS/0,5% Твина-20/0,5% ВSA) и добавляли в виде смеси в клеточные лизаты. После инкубирования в течение 1,5-2 часов при комнатной температуре и при интенсивном встряхивании добавляли магнитные сферы, покрытые стрептавидином (Dynabeads, BioVeris). После окончательного 45-минутного инкубирования планшеты считывали на аппарате BioVeris. Культивирование клеток и анализ на пролиферацию ВаF3 представляет собой мышиные IL-3-зависимые лимфоидные клетки, которые обычно не экспрессируют с-Met и не отвечают на HGF. Однако HGF стимулирует пролиферацию и выживание клеток ВаF3-hMet в отсутствии IL-3, полученных путем их трансфекции нормальной полноразмерной кДНК человеческого с-Met (Schwall et al., J. Cell. Biol. (1996) 133:709-718). Клетки ВаF3-hMet и ВаF3-neo пересевали рутинным методом в среду RPMI 1640, содержащую 5% фетальную телячью сыворотку, 4 мкл/л Иммунопреципитация и Вестерн-блот-анализ Клетки Н358 представляют собой клеточную линию, происходящую от не-мелкоклеточной карциномы легких человека (NSCLC). Клетки Н358-PSF2 и H358-PSF8 представляют собой клетки Н358, стабильно трансфицированные человеческим HGF (Tsao et al., Neoplasia, Vol.2, Исследование опухолевого ксенотрансплантата Бестимусных самок мышей подкожно инокулировали 5 миллионами клеток карциномы поджелудочной железы КР4. Когда размер опухоли достигал 150-200 мм3, мышей разделяли на 2 группы по 10 мышей в каждой. Мышам группы 1 два раза в неделю, i.p., инъецировали носитель. Мышам группы 2 два раза в неделю, i.p., инъецировали 30 мг/кг ОА5D5.v2. Размер опухоли измеряли два раза в неделю. Когда объем опухоли в два раза превышал объем исходной опухоли или когда опухоль изъязвлялась, мышей умерщвляли. Результаты и обсуждение Гуманизация 5D5. Человеческая акцепторная каркасная область, используемая для гуманизации 5D5, содержит домен консенсусной последовательности VL человеческой цепи каппа I и вариант домена консенсусной последовательности VH человеческой цепи подгруппы III. Вариант домена VH в человеческой консенсусной последовательности имеет 3 замены: R71А, N73Т и L78А. Домены VL и VH мышиного 5D5 выравнивали с доменами человеческой цепи каппа I и цепи подгруппы III, после чего каждую HVR идентифицировали и присоединяли к человеческой акцепторной каркасной последовательности с получением 5D5-гибрида, который может быть представлен на фаге в виде Fab. При фаговом представлении тестирование 5D5-гибрида на связывание с иммобилизованным HGFR-Fc не обнаруживало какого-либо связывания. Было получена библиотека, в которой каждую из областей HVR 5D5-гибрида подвергали Созревание аффинности hu5D5.v1. Для повышения аффинности hu5D5.v1 в остове hu5D5.v1 генерировали шесть библиотек фагового представления, каждая из которых была мишенью для одной HVR, в целях проведения Таким образом, в результате присоединения 6 HVR мышиного 5D5 к человеческой акцепторной каркасной последовательности HVR-Н3 удлинялась на один остаток в положении 94 (треонин), а внесение двух замен в HVR-Н3 приводило к получению полноразмерного человеческого антитела 5D5, обладающего в 13 раз более высокой аффинностью связывания с HGFR. Кроме того, в анализах на фосфорилирование рецептора и т.п. (данные не приводятся) было определено, что отобранные гуманизированные антитела, описанные в настоящей заявке, обладают биологической активностью, по меньшей мере, сравнимой с активностью родительского антитела 5D5. Характеризация антитела согласно изобретению. Были охарактеризованы антитела против Met в форме одного фрагмента (также называемые (1) В анализе на способность ОА-5D5.v2 блокировать связывание НGF с его рецептором было обнаружено, что ОА-5D5.v2 способно блокировать связывание HGF с его рецептором, по меньшей мере, с такой же эффективностью, как и два конкурентных антитела, а именно химерное однофрагментное антитело (содержащее Fab-фрагмент, происходящий от мышиного родительского антитела 5D5 (вариабельные домены представлены на фиг.7), присоединенного к человеческой Fc-области) и другое антитело согласно изобретению (OA-5D5.v1). При тестировании с использованием всех концентраций антитела в диапазоне примерно от 3333 до 0,21 нМ в условиях, описанных выше в разделе “Материалы и методы”, было обнаружено, что это антитело OA-5D5.v2 имеет величину IC50, которая составляет менее примерно половины величины IC50 эталонного антитела, такого как химерное однофрагментное антитело и OA-5D5.v1. Следует отметить, что OA-5D5.v1 также обладает блокирующей активностью при лучшей концентрации IC50, чем IС50 эталонного химерного антитела. См. фиг.8. (2) В анализе на способность OA-5D5.v2 ингибировать активацию рецептора HGF было обнаружено, что оно способно ингибировать активацию киназного рецептора, по меньшей мере, с такой же эффективностью, как и два конкурентных антитела, описанных в приведенном выше п.(1). При тестировании с использованием всех концентраций антитела в диапазоне примерно от 200 до 0,19 нМ в условиях, описанных выше в разделе “Материалы и методы”, было обнаружено, что это антитело OA-5D5.v2 имеет величину IC50, которая составляет менее примерно половины величины IC50 эталонного антитела, такого как химерное однофрагментное антитело и OA-5D5.v1. См. фиг.9. (3) OA-5D5.v2 также тестировали на перекрестное связывание с рецепторами разных видов, а именно человека, примата (собакоподобной обезьяны), животных семейства собачьих и мышиных (мыши). Было обнаружено, что OA-5D5.v2 специфически связывается с рецептором HGF человека и примата (собакоподобной обезьяны), но не связывается с рецептором HGF животных семейства собачьих и мышиных (мыши) (данные не приводятся). (4) OA-5D5.v2 тестировали на его способность ингибировать пролиферацию клеток в присутствии HGF. Как показано на фиг.10, OA-5D5.v2 ингибировало пролиферацию клеток, по меньшей мере, с такой же эффективностью, как и его аналоги, то есть химерное антитело и OA-5D5.v1 (как описано выше в п.(1)). При тестировании с использованием всех концентраций антитела в диапазоне примерно от 0,01 до 100 нМ в условиях, описанных выше в разделе “Материалы и методы”, было обнаружено, что это антитело OA-5D5.v2 имеет величину IC50, которая составляет менее примерно половины величины IС50 эталонного антитела, такого как химерное однофрагментное антитело и OA-5D5.v1. См. фиг.10. Специфическое связывание OA-5D5.v2 с Met-трансфицированными клетками подтверждали с помощью FACS-анализа (данные не приводятся). (5) OA-5D5.v2 тестировали на его способность ингибировать фосфорилирование тирозина рецептора в присутствии HGF. Как показано на фиг.11А и В, OA-5D5.v2 ингибировало фосфорилирование тирозина рецептора при концентрациях антитела примерно от 10 до 1000 нМ. См. фиг.11А и В. (6) OA-5D5.v2 тестировали на in vivo эффективность при его использовании в модели опухолевого ксенотрансплантата, полученной на основе клеточной линии опухоли поджелудочной железы (КР4). Результаты исследования такой эффективности показали, что антитело OA-5D5.v2 способно ингибировать рост опухоли и способствовать регрессии опухоли in vivo. Как показано на фиг.12, у большинства животных, обработанных указанным антителом, наблюдалось полное исчезновение опухоли. Неполный список публикаций
Формула изобретения
1. Гуманизированная форма мышиного антитела против c-met, включающая 2. Антитело по п.1, где, по меньшей мере, частью каркасной последовательности является человеческая консенсусная каркасная последовательность. 3. Антитело по п.1, содержащее вариант HVR-L2, имеющий последовательность SEQ ID NO:72. 4. Антитело по п.1, содержащее вариант HVR-L2, имеющий последовательность SEQ ID NO:98. 5. Антитело по п.1, содержащее HVR-L1, имеющую последовательность SEQ ID NO:1. 6. Антитело по п.1, содержащее HVR-L3, имеющую последовательность SEQ ID NO:3. 7. Антитело по п.1, содержащее HVR-H3, имеющую последовательность SEQ ID NO:193. 8. Антитело по п.1, содержащее консенсусную каркасную последовательность человеческой цепи подгруппы I. 9. Антитело по п.1, содержащее консенсусную каркасную последовательность человеческой тяжелой цепи подгруппы III. 10. Антитело по п.9, где указанная каркасная последовательность имеет замену в положении 71, 73 и/или 78. 11. Антитело по п.10, где указанной заменой является R71A, N73 Т и/или N78A. 12. Антитело по п.7, где указанный вариабельный домен содержит последовательности FR1-HC, FR2-HC, FR3-HC и/или FR4-HC, представленные на фигуре 13 (SEQ ID NO:187-190). 13. Антитело по п.7 или 12, где указанное антитело содержит последовательность СН1 и/или Fc, представленные на фигуре 13 (SEQ ID NO:194 и/или 195). 14. Антитело по п.7 или 12, где указанный вариабельный домен содержит последовательности FR1-LC, FR2-LC, FR3-LC и/или FR4-LC, представленные на фигуре 13 (SEQ ID NO:179-182). 15. Антитело по п.7 или 12, где указанное антитело содержит последовательность CL1, представленную на фигуре 13 (SEQ ID NO:186). 16. Антитело по п.1, содержащее вариабельный домен тяжелой цепи, включающий 17. Антитело по п.7 или 16, где указанное антитело является моновалентным и содержит Fc-область. 18. Антитело по п.17, где указанная Fc-область содержит первый и второй полипептид, где каждый из указанных первого и второго полипептидов имеет одну или более мутаций по сравнению с человеческой Fc-областью дикого типа. 19. Антитело по п.18, где указанный первый полипептид включает последовательность Fc, представленную на фигуре 13 (SEQ ID NO:195), а указанный второй полипептид включает последовательность, представленную на фигуре 14 (SEQ ID NO:196). 20. Способ ингибирования c-met-активируемой пролиферации клеток, включающий приведение клетки или ткани в контакт с эффективным количеством антитела по любому из пп.1, 16-18. 21. Способ лечения заболевания, ассоциированного с нарушением регуляции HGF/c-met-пути передачи сигнала, включающий введение индивидууму эффективного количества антитела по любому из пп.1, 16-18. 22. Способ лечения индивидуума, страдающего раком, включающий введение указанному индивидууму эффективного количества антитела по любому из пп.1, 16-18. 23. Способ по п.22, где указанным раком является рак легких, рак головного мозга, рак почек, рак желудка, рак ободочной кишки и/или рак поджелудочной железы. 24. Способ лечения продиферативного расстройства у индивидуума, включающий введение указанному индивидууму эффективного количества антитела по любому из пп.1, 16-18. 25. Способ по п.24, где указанным пролиферативным расстройством является рак. 26. Нуклеиновая кислота, кодирующая антитело по любому из пп.1, 16-18. 27. Клетка-хозяин, экспрессирующая антитело по любому из пп.1, 16-18, содержащая нуклеиновую кислоту по п.26. 28. Композиция для подавления пути HGF/c-met, содержащая эффективное количество антитела по любому из пп.1, 16-18. 29. Композиция по п.28, содержащая носитель.
РИСУНКИ
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 517
517 и мембранной остовной субъединицы
и мембранной остовной субъединицы  , связанных дисульфидными связями (Tempest et al., 1988). Субъединица
, связанных дисульфидными связями (Tempest et al., 1988). Субъединица  в 3 раза более низкую
в 3 раза более низкую аффинность по сравнению с аффинностью эталонного антитела (М), то величина Кd для А составляет 3х, величина Кd для М составляет 1х, а отношение Кd для А к Кd для М составляет 3:1. И наоборот, в одном из вариантов изобретения, если антитело согласно изобретению (С) имеет
аффинность по сравнению с аффинностью эталонного антитела (М), то величина Кd для А составляет 3х, величина Кd для М составляет 1х, а отношение Кd для А к Кd для М составляет 3:1. И наоборот, в одном из вариантов изобретения, если антитело согласно изобретению (С) имеет  I.
I.














 GEM.TM-11, может быть использован для получения рекомбинантного вектора, который может быть применен для трансформации восприимчивых клеток-хозяев, таких как E.coli LЕ392.
GEM.TM-11, может быть использован для получения рекомбинантного вектора, который может быть применен для трансформации восприимчивых клеток-хозяев, таких как E.coli LЕ392. fhuA (
fhuA ( 1,
1,  I,
I,  1
1