Патент на изобретение №2389026
|
||||||||||||||||||||||||||
(54) СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ АПУРИН/АПИРИМИДИН-ЭНДОНУКЛЕАЗЫ ЧЕЛОВЕКА
(57) Реферат:
Изобретение относится к области молекулярной биологии и биохимии. Для осуществления способа определения активности апурин/апиримидин-эндонуклеазы человека запускают реакцию путем смешивания растворов субстрата и фермента в соотношении 1:1 и регистрируют изменение интенсивности флуоресценции субстрата в интервале времени 1-2000 сек. Для этого в кювете флуориметра смешивают растворы субстрата (1,0×10-6 М), Tris-HCl буфера и образца АП-эндонуклеазы человека (1,0×10-7 М, 1,0×10-8 М, 1,0×10-9 М, 1,0×10-10 М). В качестве субстрата используют двухспиральный комплекс комплементарных олигодезоксирибонуклеотидов длиной 13 или более звеньев, содержащий в центральном положении остаток 2-оксиметил-3-окси-тетрагидрофурана и несущий красители флуоресцеин и дабцил. Изменение интенсивности флуоресценции субстрата в реакционной смеси регистрируют в спектральном диапазоне 500-550 нм, возбуждение флуоресценции проводят на длине волны 494 нм. Кинетические кривые обрабатывают методом нелинейной регрессии с помощью программы DynaFit. Использование способа позволяет повысить точность и достоверность определения активности апурин/апиримидин-эндонуклеазы человека, а также отказаться от использования радиоактивно-меченного субстрата и стандартизовать единицы измерения активности в соответствии с международной системой. 1 ил.
Изобретение относится к области биохимии и касается способа определения активности фермента репарации, апурин/апиримидин-эндонуклеазы человека (АП-эндонуклеаза, фермент Apel). Молекула ДНК, главный носитель генетической информации, может повреждаться под воздействием химических агентов, УФ- или ионизирующего излучения. Повреждения генетического аппарата обладают цитотоксическим и мутагенным эффектом и способны приводить к развитию сердечно-сосудистых, нейродегенеративных и онкологических заболеваний. Показано, что уровень повреждений в ДНК возрастает при таких заболеваниях, как лейкемия, болезнь Паркинсона, болезнь Альцгеймера, рассеянный склероз, гепатит, рак груди, рак легких и многих других (Cooke M.S. et al. FASEB J. 2003, 17, 1195-1214; Evans M.D. et al. Mutat. Res. 2004, 567, 1-61). Все живые организмы способны удалять повреждения и восстанавливать исходную структуру ДНК. Считается, что по пути эксцизионной репарации оснований (BER) удаляется большинство повреждений азотистых оснований ДНК и АР-сайты. Общий механизм BER включает в себя удаление поврежденного азотистого основания, остатка дезоксирибозы и фосфата и последующее восстановление вырезанного нуклеотида репликативным аппаратом клетки. Удаление поврежденного или неправильно спаренного основания осуществляется ДНК-гликозилазами. Большинство из них могут удалять несколько различных модифицированных оснований. Затем апуриновые/апиримидиновые эндонуклеазы удаляют оставшийся дезоксирибофосфатный остаток. Образовавшийся пробел в один нуклеотид содержит на 3′-конце гидроксильную группу, а на 5′-конце остаток фосфорной кислоты. ДНК-полимераза присоединяет нужный нуклеотид, и ДНК-лигаза заканчивает процесс BER, восстанавливая целостность дезоксирибофосфатного остова. Изучение этих ферментов в последнее время вызывает особый интерес. Знания о механизмах и особенностях действия ферментов репарации могут приблизить человечество к решению проблем раннего старения и лечения болезней, связанных с высоким уровнем генерации мутаций в геноме. АП-эндонуклеазы обнаружены в клетках всех живых организмов. Субстратом для них являются АП-сайты, возникающие в ДНК как в результате спонтанных процессов, так и в процессе действия ДНК-гликозилаз. АП-эндонуклеазы вносят разрыв с 5′-стороны от повреждения, оставляя 3′-ОН группу, необходимую для последующей инициации синтеза ДНК-полимеразой. Разработка новых способов определения активности АП-эндонуклеазы человека является актуальной задачей и чрезвычайно важна для определения уровня репарационной защиты людей, подверженных повышенным геномным нагрузкам (летчики и подводники, испытывающие окислительный стресс при дыхании кислородсодержащими смесями; работники и спортсмены, подверженные значительному УФ- и/или радиационному облучению и др.). Активность фермента характеризуют величиной каталитической константы kcat, равной числу оборотов фермента в единицу времени. За удельную активность (А) фермента принимают, как широко распространено в энзимологии (см. Биохимия, 2004, Т.69, Вып.1, С.134-144), количество мкмоль продукта, образующееся в минуту, в расчете на 1 мг фермента (мкмоль × мин-1 × мг-1). Известен способ определения активности фермента Apel путем взаимодействия с субстратом, содержащим остаток 2-оксиметил-3-окси-тетрагидрофурана (F) (Т.Izumi, С.Н.Schein, N.Oezguen, Y.Feng and W.Braun. Biochemistry. 2004, 43, 684-689; A.K.Mantha, N.Oezguen, К.K.Bhakat, T.Izumi, W.Braun and S.Mitra J. Mol. Biol. 2008, 379, 28-37). В качестве субстрата используют 32Р-меченный дуплекс длиной 43 нуклеотида. Фермент Apel (22 пМ) инкубируют с субстратом (1-200 нМ) в течение 4 минут при температуре 37°C. Реакцию останавливают добавлением раствора, содержащего 10 мМ ЭДТА и 88% формамид. Анализ проводят электрофорезом в 8%-ом полиакриламидном геле, содержащем 8 М мочевину. Гель сушат и проводят авторадиографию с помощью прибора фосфоимиджер (Storm, Molecular Dynamics). Строят зависимость скорости расщепления субстрата V0 от концентрации субстрата S0, которую обрабатывают по уравнению Михаэлиса (1) (Michaelis L., Menten M.L. Biochem. Z. 1913. 49. p.333) и определяют значения KM и kcat.
Наиболее ближайшим к заявленному способу прототипом является способ определения активности фермента Apel методом прерывания реакции (R.L.Maher and L.В.Bloom J.Biol. Chem. 2007, 282, 30577-30585). В основе метода лежит быстрое, в течение Проводят накопление 10 точек в интервале времени 0-100 мс. Анализ состава реакционной смеси в отобранных точках проводят методом гель-электорофореза в 20%-ном полиакриламидном геле, содержащем 7 М мочевину. Проводят измерение профиля радиоактивности вдоль дорожки геля и строят кинетические зависимости накопления продукта реакции. Данные обрабатывают с использованием программы SigmaPlot (Systat Software) и получают значения KM и kcat. Недостатками прототипа являются необходимость синтеза и анализа радиоактивного ДНК-субстрата, а также длительность и трудоемкость метода, связанные с использованием прибора KinTek RQF-3. Технической задачей изобретения является упрощение известного способа, повышение точности и достоверности результатов определения активности АП-эндонуклеазы человека и стандартизация единиц измерения активности в соответствии с международной системой. Способ заключается в регистрации во времени изменения интенсивности флуоресценции субстрата в реакционной смеси при взаимодействии с АП-эндонуклеазой человека. Запускают реакцию путем смешивания растворов субстрата и фермента в соотношении 1:1 и регистрируют изменение интенсивности флуоресценции субстрата в интервале времени 1-2000 сек. Для этого в кювете флуориметра смешивают растворы субстрата (1,0×10-6 М), буфера (50 мМ Tris-HCl (pH 7,5), 50 мМ KCl, 5 мМ MgCl2, 1 мМ дитиотреит, 9% глицерин) и образца АП-эндонуклеазы человека (1,0×10-7 М, 1,0×10-8 М, 1,0×10-9 М, 1,0×10-10 М). В качестве субстрата используют двухспиральный комплекс комплементарных олигодезоксирибонуклеотидов длиной 13 или более звеньев, содержащий в центральном положении остаток 2-оксиметил-3-окси-тетрагидрофурана (F), например 5′-FLU-С-С-Т-С-Т-C-F-С-С-Т-Т-С-С-3′ 3′-DABCYL-G-G-A-G-A-G-C-G-G-A-A-G-G-5′, где FLU – остаток флуоресцеина, DABCYL – остаток дабцила. Остаток 2-оксиметил-3-окси-тетрагидрофурана является специфическим сайтом узнавания фермента Apel. Один из олигодезоксирибонуклеотидов несет флуоресцентный краситель на 5′-конце, а комплементарный олигодезоксирибонуклеотид несет краситель на 3′-конце. Флуоресцентные красители подобраны таким образом, что спектр испускания флуоресценции одного красителя (флуоресцеин, FLU) перекрывается со спектром поглощения второго красителя (дабцил, DABCYL). Образование дуплекса приводит к пространственному сближению флуорофора FLU и тушителя флуоресценции DABCYL, в результате чего происходит эффективное тушение интенсивности флуоресценции (резонансный перенос энергии флуоресценции, FRET). Взаимодействие субстрата и фермента Apel приводит к расщеплению рибозо-фосфатного остова ДНК по специфическому сайту F. В результате чего в продукте реакции цепи олигонуклеотидов расходятся, что приводит к пространственному разделению флуорофора и тушителя. Интенсивность флуоресценции увеличивается по мере накопления продукта реакции. Регистрируют изменение интенсивности флуоресценции субстрата в реакционной смеси в спектральном диапазоне 500-550 нм, возбуждение флуоресценции проводят на длине волны 494 нм. Кинетические кривые обрабатывают методом нелинейной регрессии с помощью программы DynaFit (Kuzmic P. Anal. Biochem. 1996. 237. p.260-273). Использование программы позволяет минимизировать математическую обработку данных и получить значения KM и kcat. Отличиями заявленного способа от прототипа являются: – использование нерадиоактивного дуплекса олигонуклеотидов длиной 13 или более звеньев, несущих комбинацию флуоресцентных красителей флуоресцеин и дабцил, причем один из олигодезоксирибонуклеотидов несет флуоресцентный краситель на 5′-конце, а комплементарный олигодезоксирибонуклеотид несет краситель на 3′-конце, что позволяет упростить способ определения активности (отказаться от использования радиоактивно-меченного субстрата, накладывающего жесткие ограничения по технике безопасности работы, и от трудоемкого анализа состава реакционной смеси методом гель-электорофореза); – регистрацию интенсивности флуоресценции субстрата проводят в широком интервале времени 1-2000 секунд, что позволяет повысить точность и достоверность определения активности фермента (в прототипе измерение проводят в коротком интервале времени 0-100 мс); – кинетические кривые обрабатывают методом нелинейной регрессии с помощью программы DynaFit, которая позволяет минимизировать время обработки кинетических кривых и достоверно рассчитать константы KM и kcat. Изобретение иллюстрируется следующим примером конкретного выполнения. Пример Определение активности фермента Apel с использованием заявленного способа. Для определения активности фермента Apel готовят буфер следующего состава: 50 мМ Tris-HCl (pH 7,5), 50 мМ KCl, 5 мМ MgCl2, 1 мМ дитиотреит, 9% глицерин. Приготавливают в буфере 1,0×10-6 М раствор субстрата, представляющий собой дуплекс: 5′-FLU-С-С-Т-С-Т-C-F-С-С-Т-Т-С-С-3′ 3′-DABCYL-G-G-A-G-A-G-C-G-G-A-A-G-G-5′, где FLU – остаток флуоресцеина, DABCYL – остаток дабцила, F – остаток 2-оксиметил-3-окси-тетрагидрофурана. Готовят растворы фермента Apel в буфере 1,0×10-7 М, 1,0×10-8 М, 1,0×10-9 М, 1,0×10-10 М. Запускают реакцию путем смешивания растворов субстрата и фермента в соотношении 1:1 и регистрируют изменение интенсивности флуоресценции субстрата в интервале времени 1-2000 сек. Возбуждение флуоресценции проводят на длине волны 494 нм, регистрацию проводят в спектральном диапазоне 500-550 нм. На чертеже представлена зависимость интенсивности флуоресценции от времени реакции. Кинетические кривые получены при взаимодействии 1,0×10-6 М субстрата и 1,0×10-7 М, 1,0×10-8 М, 1,0×10-9 М, 1,0×10-10 М фермента Apel. Для обработки данных используют схему Михаэлиса-Ментен (Michaelis L., Menten M.L. Biochem. Z. 1913. 49. p.333). Кинетические кривые анализируют с помощью программы DynaFit (Kuzmic P. Anal. Biochem. 1996. 237, p.260-273), которая позволяет проводить численное интегрирование и подгонку параметров методом нелинейной регрессии. В результате обработки получают значения KM и kcat. KM=(1,1±0,4)×10-7 М и kcat=2,4±0,3 с-1. За удельную активность (А) фермента принимают количество мкмоль продукта, образующееся в минуту, в расчете на 1 мг фермента (мкмоль × мин-1 × мг-1), и рассчитывают по формуле:
где 2,8×10-2 – пересчетный коэффициент, равный количеству мкмоль фермента Apel в 1 мг (молекулярная масса фермента Apel равна 35,5 кДа). Получено, что kcat=2,4±0,3 с-1=144±18 мин-1. Активность А=2,8×10-2×144=4,03 (мкмоль × мин-1 × мг-1). Использование способа позволяет повысить точность и достоверность определения активности фермента Apel и наряду с этим отказаться от использования радиоактивно-меченного субстрата и стандартизовать единицы измерения активности в соответствии с международной системой.
Формула изобретения
1. Способ определения активности апурин/апиримидин-эндонуклеазы человека, включающий смешивание растворов фермента с ДНК-субстратом, содержащим остаток 2-оксиметил-3-окси-тетрагидрофурана, и вычисление значения каталитической константы (kcat, мин-1) по заданным уравнениям с использованием компьютерной программы, отличающийся тем, что в качестве ДНК-субстрата используют дуплекс олигонуклеотидов длиной 13 или более звеньев, несущих комбинацию флуоресцентных красителей флуоресцеин и дабцил, при этом один из олигодезоксирибонуклеотидов, содержащий в центральном положении остаток 2-оксиметил-3-окси-тетрагидрофурана (F) несет флуоресцентный краситель на 5′-конце, а комплементарный олигодезоксирибонуклеотид несет краситель на 3′-конце, при этом растворы фермента и субстрата смешивают в соотношении 1:1 с последующим измерением активности фермента путем регистрации изменения интенсивности флуоресценции субстрата в интервале времени 0-2000 с, причем возбуждение флуоресценции проводят на длине волны 494 нм, а изменение интенсивности флуоресценции субстрата регистрируют в спектральном диапазоне 500-550 нм. 2. Способ по п.1, отличающийся тем, что субстрат используют в концентрации, равной 1,0·10-6 М, а фермент в концентрациях 1,0·10-7 М, 1,0·10-8 M, 1,0·10-9 M, 1,0·10-10 M. 3. Способ по п.1, отличающийся тем, что значение каталитической константы получают методом нелинейной регрессии с помощью программы DynaFit.
РИСУНКИ
|
||||||||||||||||||||||||||

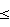 1 мс, смешивание растворов фермента и субстрата с помощью прибора KinTek RQF-3 (KinTek). Фермент инкубируют с субстратом, состоящим из 32Р-меченной цепи 5′-[32P]-d(GCGTCAAAATGTFGGTATTTCCATG)-3′ и комплементарной цепи 5′-d(CATGGAAATACCTACATTTTGACGC)-3′. Реакцию проводят в буферном растворе состава 50 мМ HEPES, pH 8,0, 100 мМ KCl, 5 мМ MgCl2, 5% глицерин, 0,2 мг/мл БСА. Концентрация фермента составляет 0,2 мкМ, концентрация субстрата составляет 0,5, 1,0, 2,0 мкМ. Прерывание реакции происходит при быстром смешивании с 0,2 М раствором NaOH, затем в раствор добавляют 96% формамид, 20 мМ ЭДТА и нагревают до 95°C.
1 мс, смешивание растворов фермента и субстрата с помощью прибора KinTek RQF-3 (KinTek). Фермент инкубируют с субстратом, состоящим из 32Р-меченной цепи 5′-[32P]-d(GCGTCAAAATGTFGGTATTTCCATG)-3′ и комплементарной цепи 5′-d(CATGGAAATACCTACATTTTGACGC)-3′. Реакцию проводят в буферном растворе состава 50 мМ HEPES, pH 8,0, 100 мМ KCl, 5 мМ MgCl2, 5% глицерин, 0,2 мг/мл БСА. Концентрация фермента составляет 0,2 мкМ, концентрация субстрата составляет 0,5, 1,0, 2,0 мкМ. Прерывание реакции происходит при быстром смешивании с 0,2 М раствором NaOH, затем в раствор добавляют 96% формамид, 20 мМ ЭДТА и нагревают до 95°C.
