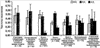Патент на изобретение №2388829
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) СКРИНИНГ ГЕНОТИПА ACTN3 ДЛЯ АНАЛИЗА СПОРТИВНЫХ СПОСОБНОСТЕЙ
(57) Реферат:
Изобретение относится к области генной инженерии, конкретно к генотипическому прогнозированию спортивных способностей, и может быть использовано в спортивной медицине. Способ основан на оценке генотипа ACTN3, где генотип 577RR положительно связан со спринтерскими или силовыми способностями; генотип 577ХХ отрицательно связан со спринтерскими или силовыми способностями; генотип 577ХХ положительно связан с выносливостью; генотип 577RX положительно связан со спринтерскими или силовыми способностями; а генотип 577RX отрицательно связан с выносливостью у женщин. Способ позволяет подобрать вид спорта или вид соревнований для индивидуума, например скоростно-силовой вид спорта или вид спорта, требующий выносливости, а также разработать более оптимальный тренировочный режим для отдельного спортсмена. 2 з.п. ф-лы, 1 ил., 6 табл.
Данная заявка претендует на приоритет Australian Provisional Patent Application ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ Настоящее изобретение относится к способам выбора или подбора вида спорта или вида соревнований для индивидуума (например, скоростно-силовой вид спорта или вид спорта, требующий выносливости) для увеличения его шансов на успех, оптимизации программ тренировок индивидуумов и для предсказания спортивных способностей индивидуума. Определенные осуществления изобретения относятся к выявлению конкретного гена(ов) или изменений гена(ов), коррелирующих с потенциальными спортивными способностями. Более конкретно, изобретение относится к способам генотипирования индивидуума по гену, кодирующему белок скелетных мышц, ОПИСАНИЕ СВЯЗАННОЙ ОБЛАСТИ В возрастающей для спортивной деятельности конкурентной среде все большее значение приобретают программы поиска талантливых индивидуумов для того, чтобы как можно раньше выявлять индивидуума, который может стать элитным атлетом, чтобы обеспечить его преимущества в достижении максимальных успехов. Эти программы поиска талантливых индивидуумов в настоящее время основаны на данных фактических способностей и фенотипических прогностических факторах, определяемых в отношении типа предстоящей тренировки, а также, вероятно, требованиями конкретного вида спорта. Одним из недостатков современных программ тренировок и критериев поиска талантливых индивидуумов является отсутствие возможности определения, достиг ли индивидуум своих потенциальных способностей и, таким образом, способен ли он оптимально реагировать на дальнейшие тренировки. Другим недостатком современных программ поиска талантливых индивидуумов, которые особенно значимы в странах с относительно малой численностью населения на большой географический район, является отсутствие возможности выбора. Индивидуум, выросший в среде с широким доступом к спортивным и тренировочным приспособлениям, с большей вероятностью добьется результата и, следовательно, с большей вероятностью на него обратят внимание тренеры и специалисты по поиску талантливых индивидуумов, чем на молодого человека с возможностями, но который вырос в относительно изолированной области или по другим причинам получивший меньшую подготовку. Подобным образом, легко упустить из вида индивидуумов с возможностями проявить себя в менее распространенных видах спорта, таких как гребля, так как эти спортивные программы в большинстве школ менее доступны. Это снова уменьшает шансы раннего выявления и привлечения, что приводит к дальнейшему упущению тренерами и специалистами по поиску талантливых индивидуумов. Такие недостатки являются дилеммами, стоящими перед такими спортивными организациями как Australian Institute of Sport (AIS), так как потенциальных высококлассных спортсменов предпочтительно отбирать и привлекать в соответствующие в тренировочные программы в молодом возрасте. Существует вероятность того, что сцепления или ассоциации генотипа или генотипических маркеров с определенными физиологическими особенностями могут вносить вклад в способности у высококлассного спортсмена или уменьшать их. С помощью таких способов можно разработать способы скрининга ДНК, которые способствуют отбору индивидуумов с большим спортивным потенциалом. С помощью таких способов скрининга можно преодолеть некоторые ограничения при отборе в современных программах поиска талантливых индивидуумов. Кроме того, с помощью таких способов скрининга можно распознать, кому и когда следует сделать, возможно, небольшие, но важные изменения в индивидуальной программе тренировки.
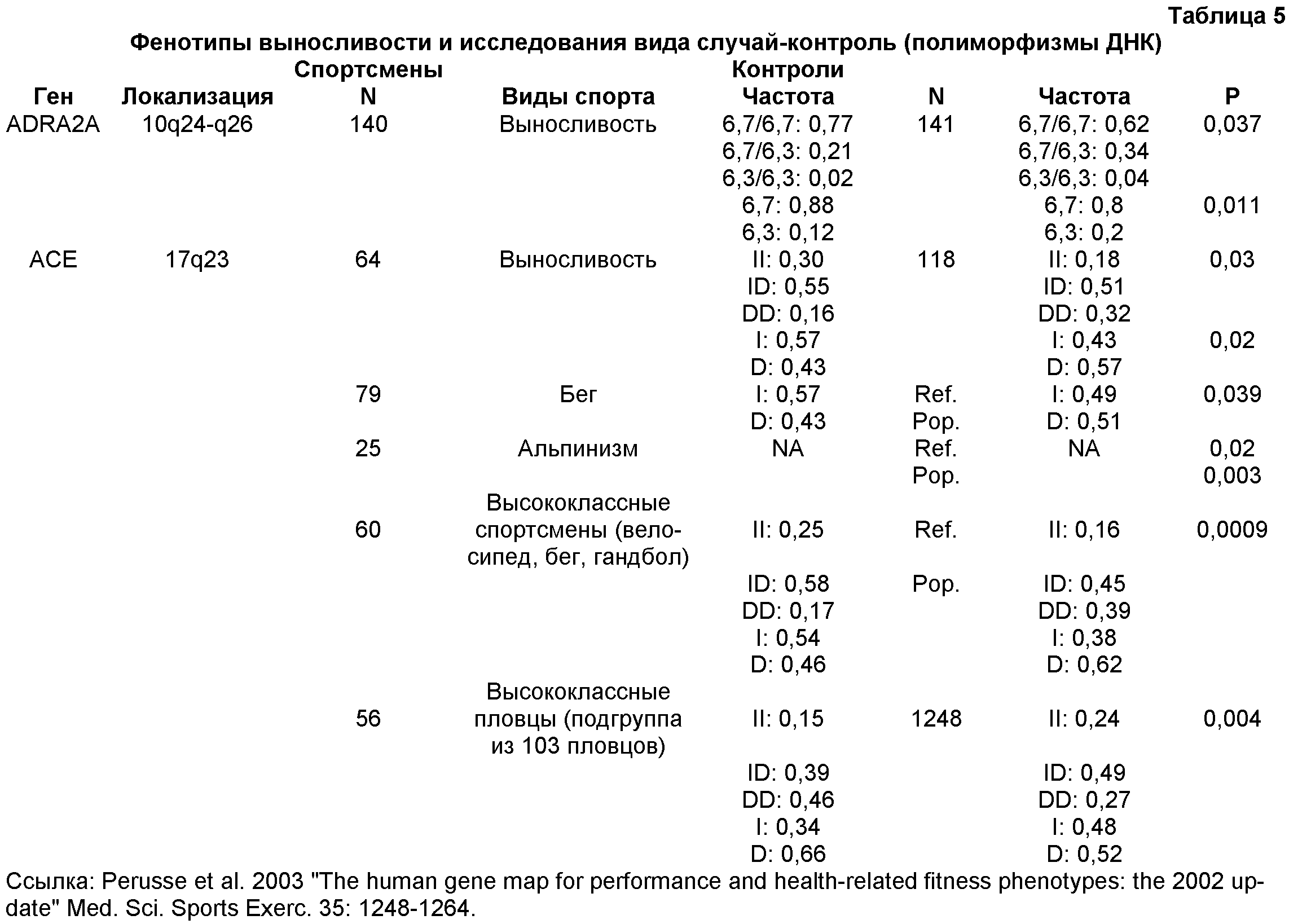
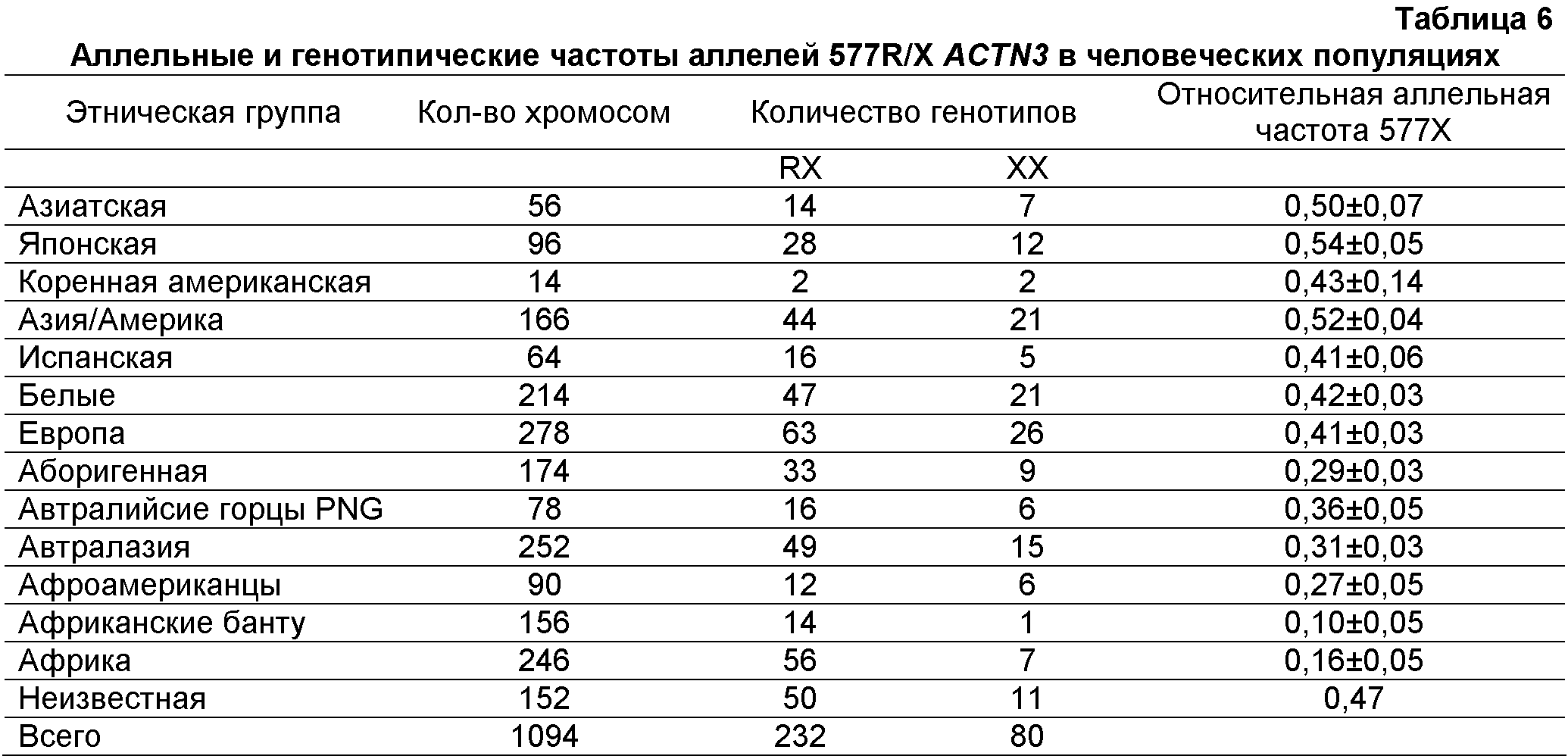
Было обнаружено, что У людей ген Функциональная вырожденность возникает тогда, когда два гена выполняют перекрывающиеся функции так, что инактивация одного из генов оказывает небольшой эффект на фенотип или не оказывает его совсем (рассмотрено в Nowak, M. A. et al., Nature, 388, 167-171, 1997). В человеческой скелетной мышце экспрессия СУЩНОСТЬ ИЗОБРЕТЕНИЯ Несмотря на явную функциональную вырожденность ACTN3 и ACTN2 у людей, исследования генотипа группы высококлассных австралийских спортсменов и известных спринтеров европеоидной расы (в частности, бегуна на короткие дистанции, пловцов и велосипедистов) показали очень низкую частоту гомозиготного состояния приводящей к преждевременному стоп-кодону мутации ACTN3 577X (т.е. нуль-мутации ACTN3, 577XX) относительно не ограниченной популяции австралийских европеоидов. Поэтому полагают, что скрининг генотипа ACTN3, обеспечит значительную помощь при выборе молодых индивидуумов с высоким спортивным потенциалом в области скоростных видов спорта и видов соревнований. Исследования генотипа показали также, что частота генотипа 577XX относительно выше у высококлассных спортсменов видов спорта, требующих выносливости, европеоидной расы. Таким образом, способ скрининга генотипа ACTN3 577XX, может также быть полезным при выявлении молодых индивидуумов с высоким спортивным потенциалом в видах спорта и видах соревнований, требующих выносливости. Настоящее изобретение удовлетворяет потребность в данной области, предоставляя способы скрининга индивидуумов in vitro для анализа спортивного потенциала. В одном из вариантов осуществления можно определить генотип индивидуума для гена ACTN3. В другом варианте осуществления из скелетной мышцы типа 2 выделяют мРНК или белок и анализируют на наличие или отсутствие ACTN3. В другом варианте осуществления индивидуумов идентифицируют, выделяя ДНК из образцов крови или щечного мазка и проводя амплификацию ДНК и ее анализ на присутствие или отсутствие в гене ACTN3 преждевременного стоп-кодона (577X). В других вариантах осуществления предоставлены способы скрининга индивидуумов на анализ спортивного потенциала посредством сочетания скрининга ACTN3 с другими генетическими и физиологическими тестами. Кроме того, описанные способы, посредством генетических оценок, физиологических тестов, физических измерений и/или физиологических оценок обеспечивают разработку тренировочных программ(ы), более подходящих конкретному спортсмену. В другом варианте осуществления изобретение относится к скринингу индивидуумов на выявление высокого спортивного потенциала, способ, например, осуществляют посредством получения от индивидуума образца подходящих мышечных клеток и выявления в образце белка Конкретные варианты осуществления изобретения относятся к способу прогнозирования наличия или отсутствия конкретного фенотипа. Способ включает в себя получение от индивидуума образца нуклеиновой кислоты и определение идентичности одного или нескольких оснований (нуклеотидов) в конкретных (например, полиморфных) участках молекул нуклеиновой кислоты, описанных здесь, где присутствие конкретного основания в данном участке коррелирует с конкретным генотипом, таким образом, прогнозируя наличие, отсутствие или вероятность присутствия или отсутствия у индивидуума данного фенотипа. КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ Приведенный ниже чертеж является частью настоящего описания и включен сюда для дополнительной демонстрации определенных аспектов настоящего изобретения. Изобретение будет легче понять из чертежа в сочетании с подробным описанием представленных здесь конкретных осуществлений. На чертеже представлена частота генотипов ACTN3 в контроле высококлассных спортсменов в области скоростно-силовых видов спорта и высококлассных видов спорта, требующих выносливости спортсменов. В таблице 1 представлены генотипы SNP R577X в ACTN3 у высококлассных спортсменов-европеоидов конкретных дисциплин. В таблице 2 представлены суммарные данные об индивидах, тестируемых на количество и частоту (%) аллелей ACTN3 в контрольной группе и у высококлассных спортсменов в области скоростно-силовых видов спорта и у высококлассных выносливых спортсменов. В таблице 3 представлены SNP, выявленные до настоящего времени в гене ACTN3 и собранные в список с веб-сайта SNP NCBI. В таблице 4 представлены обозначения, полные названия и цитогенетическая локализация ядерных и митохондриальных генов из генетической карты человеческих фенотипов способностей и связанных со здоровьем показателей 2002 года. В таблице 5 представлены фенотипы выносливости и исследования вида случай-контроль (полиморфизмов ДНК). В таблице 6 представлены частоты генотипов и аллелей 577/R/X ACTN3 в различных человеческих популяциях. Определения Термин “высококлассный спортсмен” или его варианты относится к спортсменам, которые демонстрируют самые высокие уровни с точки зрения выносливости, скорости и/или силы (например, такие, которые в своем виде спорта способны состязаться на уровне штата, страны и/или на международном уровне). Как используется здесь, термины “SNP” или “однонуклеотидные полиморфизмы” относятся к заменам одного основания в конкретном положении в геноме организма (например, человека). ПОДРОБНОЕ ОПИСАНИЕ В следующем далее разделе для иллюстрации различных осуществлений изобретения описаны некоторые варианты осуществления, например, способы. Однако, специалисту в данной области понятно, что различные варианты осуществления на практике не требуют применения всех или даже нескольких конкретных деталей, описанных здесь. Хорошо известные способы или компоненты в некоторых случаях не включены в описание. Описаны способы и композиции для скрининга индивидуумов для анализа наличия спортивных возможностей. В одном варианте осуществления изобретения описан способ скрининга индивидуумов для анализа присутствия или отсутствия белка или мРНК ACTN3. В другом варианте осуществления изобретения описан способ скрининга индивидуумов для анализа наличия или отсутствия изменений генотипа ACTN3. В другом варианте осуществления изобретения описан способ скрининга индивидуумов для анализа наличия или отсутствия конкретных генотипов ACTN3, таких как 577RR, 577XR или 577XX. Выявление белка ACTN3 можно проводить прямым или непрямым измерением уровней белка (например, антитела и т.д.). Полиморфизмы ACTN3 и другие генетические изменения В гене, кодирующем белок скелетных мышц На спортивные способности в скорости/силе и/или выносливости могут оказывать положительные воздействия другие гены. Например, сообщалось, что у ангиотензин-превращающего фермента (ACE) присутствуют два аллеля I и D, оказывающие воздействие на спортивные способности. Аллель I ассоциирована с меньшей активностью ACE в сыворотке и ткане (Reider et al., “Sequence variation in the human angiotensin converting enzyme”. Nat Genet, 1999 vol. 22 pp59-62). Сообщалось, что у высококлассных выносливых спортсменов увеличена частота аллеля I (Gayagay et al. 1998 “Elite endurance athletes and the ACE I allele; the role of genes in athletic performance”. Hum Genet 103: 48-50; Montgomery et al. 1998 Human gene for physical performance. Nature 393: 221-222; Myerson et al. 1999 Human angiotensin 1-converting enzyme gene and endurance performance. J Appl Physiol 87: 1313-1316; Nazarov et al. 2001 “The angiotensin converting enzyme I/D polymorphism in Russian athletes” Eur J Hum Genet 9: 797-801). Наоборот, увеличенная частота аллеля ACE D связана с высокими скоростными способностями (Myerson et al. 1999 Human angiotensin 1-converting enzyme gene and endurance performance. J Appl Physiol 87: 1313-1316; Nazarov et al. 2001 “The angiotensin converting enzyme LID polymorphism in Russian athletes” Eur J Hum Genet 9: 797-801; Woods et al. 2001 Elite swimmers and the D allele of the ACEI/D polymorphism. Hum Genet 108: 230-232). Возможно, что между способностью к развитию скорости и показателем выносливости существует соотношение, которое накладывает ограничение на эволюцию физических способностей у людей и других позвоночных (Garland et al. 1990 “Heritability of locomotor performance and its correlates in a natural population” Experientia 46: 530-533). Это подтверждается данными, полученными у десятиборцев мирового класса, демонстрирующими, что способности в спринте на 100 м, толкании ядра, прыжке в длину и беге на 110 м с барьерами (зависящие от взрывной силы и быстрых чувствительных к усталости мышечных волокон) отрицательно коррелируют со способностью в беге на 1500 м (требующими выносливости и активности устойчивых к усталости медленных волокон). (Van Damme et al. 2002 Performance constraints in decathletes. Nature 415: 755-756). Это означает, что индивидуум может быть предрасположен к высококлассным способностям только в одной из двух областей (скорость/сила или выносливость). В конкретных вариантах осуществления изобретения тесты для скрининга ACTN3 можно объединять с одним или несколькими генетическими тестами других ассоциированных со способностями генов. Такие тесты могут включать в себя любой ген, который, как известно в данной области, ассоциирован со способностями в скорости/силе и/или выносливости (например, Rankinen et al 2002, “The human gene map for performance and health-related fitness phenotypes: the 2001 update” Med. Sci. Sports Exerc. 34: 1219-33; Perusse et al. 2003, “The human gene map for performance and health-related fitness phenotypes: the 2002 update” Med. Sci. Sports Exerc. 35: 1248-1264, которые приведены здесь в качестве ссылки в полном объеме). Два обзора (Rankinen et al. 2002; Perusse et al. 2003) подводят итог результатам исследований фенотипов способностей и связанных со здоровьем показателей. Генетическая карта человеческих способностей и связанных со здоровьем показателей представлена в виде фиг.1 в статье 2002 года. Данная карта включает в себя все генетические элементы и QTL (локусы количественных признаков), в которых была показана связь или сцепление с фенотипами, связанными с тренировками. Хромосомы и их области взяты с веб-сайта Gene Map of the Human Genome, the National Center for Biotechnology Information (NCBI), National Institutes of Health, Bethesda, MD. Сокращения локусов и полные названия генов с возможным применением в сочетании со скринингом ACTN3 суммированы в таблице 4. В одном осуществлении, для предсказания наличия у данного индивидуума большого спортивного потенциала, в сочетании с оценкой у индивидуума гена ACTN3, можно применять анализ одного или нескольких из указанных в таблице 4 генов. В таблице 5 подведен итог исследования (Perusse et al., 2003) частот аллелей и генотипов генов ADRA2A (альфа-2A-адренергический рецептор) и ACE (ангиотензин-1-превращающий фермент) у выносливых спортсменов по сравнению с малоподвижными индивидами. В одном варианте осуществления изобретения для более точного предсказания спортивного потенциала индивидуума тестирование генотипа ACTN3 потенциального высококлассного спортсмена можно сочетать с оценкой его генотипа ADRA2A и/или генотипа ACE. В другом варианте осуществления оценку у спортсмена генотипа ACTN3 можно сочетать с оценкой или генотипа ADRA2A и/или генотипа ACE и/или другими физиологическими оценками (например, максимума VO2 и т.д.) для установки спортсмену тренировочного режима. Эволюционное расхождение ACTN3 и ACTN2 Генотипирование приматов, не относящихся к человеку, указывает на то, что нуль-мутация 577X, вероятно, возникла у людей. Мышиный геном содержит четыре ортолога, соответствующих эволюционно консервативным областям четырех человеческих генов. Мышиные ACTN2 и ACTN3 в ходе эмбрионального развития по-разному экспрессируются пространственно и во времени и, в противоположность экспрессии человеческого Для определения происхождения аллеля 577X (и аллеля 523R, находящемуся с 577X в жестком неравновесии по сцеплению), генотипировали 36 неродственных бабуинов (разошедшихся с человеческой линией 25×106 лет назад) и 33 неродственных шимпанзе (разошедшихся с человеческой линией 5×106 лет назад). Все 69 не являющихся людьми приматов в экзонах 15 (523Q) и 16 (577R) являлись гомозиготными по аллелям дикого типа, что означает, что полиморфизмы возникли после разделения линий человека и шимпанзе, или что они у не являющихся людьми приматов обладают очень низкой частотой (Mills et al 2001). В отношении мышей, сходство между мышиными ACTN2 и ACTN3 является таким же, как и между человеческими ACTN2 и ACTN3, т.е. 88% сходства и 79% идентичности. Мышиные белки являются коллинеарными и обладают такими же функциональными доменами, как человеческие белки, – N-концевой связывающий актин домен, четыре центральных повторяющихся домена и C-концевые EF-ручки (Mills et al 2001). У кур присутствует только один ген ACTN скелетных мышц, тогда как мышиный геном содержит четыре ортолога, соответствующие эволюционно консервативным синтенным областям четырех человеческих генов. Сравнение последовательности мышиных и человеческих ACTN2 и ACTN3 позволяет предположить, что эволюция У других млекопитающих, таких как кролики и свиньи, также существуют быстрые и медленные специфичные для мышц изоформы У млекопитающих сохранились обе копии гена, а сравнение последовательностей человеческих и мышиных ACTN2 и ACTN3 демонстрирует, что в ходе эволюции млекопитающих данные гены оставались высококонсервативнымии (Mills et al 2001). Спортивные способности и лошади Лошадь является одним из очень небольшого количества видов животных, кроме некоторых собак и верблюдов, который разводили, содержали и продавали из-за их спортивных способностей и, следовательно, она является еще одной моделью для изучения экспрессии генов, связанных со способностями. Например, ранее было показано сохранение ACTN3, спортивного маркера для спортивных способностей людей, и гена ACTN2 у различных видов. Хотя пока у лошадей не выявили эквивалентного гена, весьма вероятно, что у лошадей существует ген, подобный ACTN3, но он пока не выявлен. В определенных вариантах осуществления изобретения, лошади могут быть подвергнуты скринингу для анализа подобного ACTN3 гена. В других вариантах осуществления для анализа подобного ACTN3 гена скринингу могут быть подвергнуты скаковые лошади, такие как лошади, тренируемые для выступления в дерби. Альтернативно, также могут быть подвергнуты скринингу для анализа дифференциальной экспрессии подобного ACTN3 гена лошади с необходимостью в спринте очень большой интенсивности, такие как пони для поло и лошади для скачек вокруг бочек. Вероятно, что у спринтерских лошадей экспрессируется ген, немного отличающийся от гена выносливых лошадей, и, следовательно, анализ подобного ACTN3 гена может являться индикатором высокого спортивного потенциала у лошадей. Подобно тому, что наблюдается у спортсменов, скрининг гена для анализа небольшого изменения, например, присутствия или отсутствия конкретной нуклеотидной последовательности (например, участка SNP, делеции или вставки) может оказаться у таких животных, как лошади, ценным индикатором высокого спортивного потенциала. Ген, подобный ACTN3, представляет собой ген, выполняющий ту же функцию, что и ACTN3 у других видов, и/или обладает сходствами последовательности с геном ACTN3. Предыдущие исследования указывают на то, что лошадиный ген ангиотензин-превращающего фермента может являться идеальным геном-кандидатом для анализа спортивных способностей лошадей. Человеческий вариант гена содержит полиморфный маркер, ассоциированный с увеличенными спортивными способностями высококлассных выносливых спортсменов, и увеличенным анаболическим ответом к тренировке (Ellis et al, “Characterization of the Equine Angiotensin-converting Enzyme” 7th World Congress on Genetics Applied to Livestock Production, August 19-23, 2002, Montpellier, France Session 05. Horse breeding Abstract of N° 05-07 GENE. N: A.I. Tammen, F.W. Nicholas and H.W. Raadsma. ReproGen, University of Sydney, Camden, Australia). До настоящего времени попытки провести корреляцию между экспрессией ACE и высокими спортивными способностями у лошадей являлись безуспешными. Другие исследования включают в себя изучение гена тяжелых цепей миозина (MyHC) в средней ягодичной мышце лошадей, где была выявлена дифференциальная экспрессия гена у жеребцов, но прямую связь спортивных возможностей и присутствия или отсутствия гена до сих пор не связали со спортивными способностями (Eizema et al “Differential Expression of Equine Myosin heavy-chain mRNA and Protein Isoforms in a Limb muscle” J Histochem Cytochem 2003 Sept; 51 (9): 1207-1216). Полагают, что для более точной оценки высоких спортивных способностей лошади в раннем возрасте у лошадей можно проводить анализ подобного ACTN3 гена и других физиологических и генетических параметров. Полагают, что лошадей перед их скрещиванием можно предварительно подвергать скринингу для выявления более подходящего генетического соответствия. Кроме того, возможно, что скринингу для оценки спортивного потенциала жеребенка до его рождения можно подвергнуть жеребенка в утробе. Полученная из такого скрининга информация могла бы сохранить селекционерам и исследователям лошадей (верблюдов, собак) огромное количество времени и денег, а также выявить потенциальную способность животного на ранней стадии развития. Как и для людей, полученная при генотипическом скрининге лошади информация, а также другие параметры (родословная и т.д.), могут помочь выявить потенциальное высококлассное спортивное животное и/или разработать для конкретного животного (например, пони для поло) наилучший режим тренировки. Однонуклеотидные полиморфизмы (SNP) В различных вариантах осуществления изобретения предоставлены способы определения корреляции между полиморфизмом или генетической вариацией (например, SNP) и фенотипом, который включает в себя: a) получение: образцов от одного или нескольких субъектов; возможно, медицинских карт одного или нескольких субъектов для определения фенотипа субъекта(ов) и аналитических наборов для определения, выявляющих полиморфизм; b) проведение с образцами анализов определения в таких условиях, в которых выявляется присутствие или отсутствие, по меньшей мере, одного полиморфизма, и c) определение корреляции, по меньшей мере, между одним полиморфизмом и фенотипом субъектов. Нуклеиновые кислоты в представляющей интерес области (например, области содержащей в себе представляющую интерес генетическую вариацию) можно анализировать с применением пригодного способа, включающего в себя в качестве неограничивающих примеров ручное определение последовательности с применением радиоактивных маркерных нуклеотидов или автоматическое определение последовательности. Последовательность можно анализировать и выявлять в ней присутствие или отсутствие данного SNP или мутации. Для оценки наличия, отсутствия или изменения конкретного гена для оценки спортивного потенциала индивидуума или модификации тренировочного режима для данного индивидуума можно применять конкретный участок(ки) SNP гена (например 1747 C>T ACTN3). Известные для ACTN3 SNP перечислены в таблице 3. В различных осуществлениях изобретения скрининг SNP 1747 C>T гена ACTN3 можно сочетать со скринингом других известных полиморфных участков в гене ACTN3, включая сюда в качестве неограниченных примеров любой перечисленный в таблице 3 SNP. Другие SNP с потенциальным применением в практическом осуществлении способов по данному изобретению описаны, например, в таблице в опубликованной патентной заявке США с порядковым В альтернативных вариантах осуществления изобретения для скрининга генетических вариантов можно применять другие анализы определения, такие как анализ аллелеспецифической гибридизации. В анализе гибридизации наличие или отсутствие данного SNP или другой генетической вариации определяют на основе способности ДНК из образца гибридизоваться с комплементарной молекулой ДНК (например, с олигонуклеотидным зондом). В данной области известно множество способов анализа гибридизацией, применяющих множество способов для гибридизации и детекции, и любой из таких известных способов можно применять в способах по данному изобретению. Ниже описаны иллюстративные анализы. В некоторых осуществлениях в анализах определения можно применять анализ гибридизацией на чипе ДНК. В таких анализах на твердую подложку прикрепляют ряд олигонуклеотидных зондов. В некоторых осуществлениях сконструированы уникальные для данного SNP или мутации олигонуклеотидные зонды. Представляющий интерес образец ДНК приводят в контакт с “чипом” ДНК и определяют результат гибридизации. Чипы ДНК, включающие в себя изготовленные на заказ чипы ДНК, специфичные для конкретных последовательностей SNP, доступны из коммерческих источников, таких как Affymetrix (Santa Clara, CA). В других иллюстративных осуществлениях полиморфизмы можно определять с применением анализа удлинения праймера SNP-IT (Orchid Biosciences, Princeton, N.J.; например, патенты США Нуклеиновые кислоты Различные варианты осуществления изобретения включают в себя выделение и анализ молекул нуклеиновых кислот, таких как ДНК, мРНК или кДНК. Представляющие интерес нуклеиновые кислоты могут кодировать часть заданного белка или весь заданный белок (например, ACTN3, ACE и т.д.). Как используется здесь, термин “нуклеиновая кислота” включает в себя одноцепочечные и двухцепочечные молекулы, а также ДНК, РНК, химически модифицированные нуклеиновые кислоты и аналоги нуклеиновых кислот. Полагают, что нуклеиновая кислота в объеме данного изобретения может быть длиной в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, приблизительно 110, приблизительно 120, приблизительно 130, приблизительно 140, приблизительно 150, приблизительно 160, приблизительно 170, приблизительно 180, приблизительно 190, приблизительно 200, приблизительно 210, приблизительно 220, приблизительно 230, приблизительно 240, приблизительно 250, приблизительно 275, приблизительно 300, приблизительно 325, приблизительно 350, приблизительно 375, приблизительно 400, приблизительно 425, приблизительно 450, приблизительно 475, приблизительно 500, приблизительно 525, приблизительно 550, приблизительно 575, приблизительно 600, приблизительно 625, приблизительно 650, приблизительно 675, приблизительно 700, приблизительно 725, приблизительно 750, приблизительно 775, приблизительно 800, приблизительно 825, приблизительно 850, приблизительно 875, приблизительно 900, приблизительно 925, приблизительно 950, приблизительно 975, приблизительно 1000, приблизительно 1100, приблизительно 1200, приблизительно 1300, приблизительно 1400, приблизительно 1500, приблизительно 1750, приблизительно 2000, приблизительно 2250, приблизительно 2500 или более нуклеотидных остатков, вплоть до и включая полноразмерную хромосомную ДНК. Способы частичной или полной очистки ДНК и/или РНК из сложных смесей, таких как клеточные гомогенаты и экстракты, хорошо известны в данной области. (См., например, Guide to Molecular Cloning Techniques, eds. Berger and Kimmel, Academic Press, New York, NY, 1987; Molecular Cloning: A Laboratory Manual, 2nd Ed., eds. Sambrook, Fritsch and Maniatis, Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989). Как правило, клетки, ткани и другие источники материала, содержащего нуклеиновые кислоты, сначала гомогенизируют, например, посредством замораживания в жидком азоте с последующим растиранием пестиком в ступке. Некоторые ткани можно гомогенизировать с применением смесителя Waring, гомогенизатора Virtis, гомогенизатора Dounce или другого гомогенизатора. Грубые гомогенизаты можно получать с применением детергентов, таких как додецилсульфат натрия (SDS), Triton X-100, CHAPS (3-[(3-холамидопропил)-диметиламмонио]-1-пропансульфонат), октилглюкозид или других известных в данной области детергентов. Как хорошо известно, для предотвращения деградации представляющих интерес нуклеиновых кислот можно добавлять ингибиторы нуклеазы, такие как ингибиторы РНКазы и ДНКазы. Выделение также можно проводить с применением хаотропных средств, таких как гуанидинизотиоцианат, или органические растворители, такие как фенол. В некоторых осуществлениях для разрушения клеточных белков можно применять обработку протеазой, например, такой как протеиназа K. Корпускулярные примеси можно удалять центрифугированием или ультрацентрифугированием. Для удаления солей или других растворимых примесей можно применять диализ против водного буфера с низкой ионной силой. Нуклеиновые кислоты можно осаждать добавлением этанола при -20°C или добавлением ацетата натрия (pH 6,5, приблизительно 0,3 М) и 0,8 объема 2-пропанола. Осажденные нуклеиновые кислоты можно собирать центрифугированием или, в случае хромосомной ДНК, наматыванием осажденной ДНК на стеклянную пипетку или другой зонд. Специалистам в данной области будет понятно, что перечисленные выше способы являются только иллюстративными и что в зависимости от конкретного типа нуклеиновой кислоты для анализа можно применять множество вариантов. В определенных вариантах осуществления нуклеиновые кислоты для анализа могут представлять собой природные молекулы ДНК или РНК. Практически, описанными способами можно анализировать любую природную нуклеиновую кислоту, включая сюда в качестве неограничивающих примеров хромосомную, митохондриальную или хлоропластную ДНК или рибосомальную, транспортную, гетерогенную ядерную или матричную РНК. Нуклеиновые кислоты можно получать или из прокариотических или из эукариотических источников известными в данной области стандартными способами. Альтернативно, представляющие интерес нуклеиновые кислоты можно получать искусственно, например, посредством ПЦР Амплификация нуклеиновых кислот В конкретных вариантах осуществления, анализируемые для скрининга нуклеиновые кислоты для увеличения силы сигнала вначале можно амплифицировать. Последовательности нуклеиновой кислоты для применения в качестве матрицы для амплификации можно выделять из клеток, содержащихся в биологическом образце (например, ДНК или мРНК из скелетной мышцы), стандартными способами. Нуклеиновая кислота может представлять собой геномную ДНК или фракционированную РНК или РНК всей клетки. Когда применяют РНК, может быть желательным преобразовать РНК в комплементарную ДНК. В одном осуществлении РНК представляет собой РНК всей клетки, и ее непосредственно применяют как матрицу для амплификации. В одном из примеров определение генотипа ACTN3 проводят посредством амплификации (например, посредством ПЦР) полинуклеотидных последовательностей ACTN3 или, более предпочтительно, его фрагмента, включающего в себя SNP 1747 C>T (например, экзон 16), и посредством секвенирования продуктов амплификации или посредством определения присутствия и/или отсутствия SNP 1747 C>T в амплифицированных последовательностях другим способом. В другом примере известно, что аллель 577X содержит участок рестрикции DdeI, который можно легко выявить рестрикцией амплифицированных продуктов посредством DdeI и разделением продуктов рестрикции по размеру (например, посредством электрофореза в геле). Размер продуктов можно использовать для генотипирования локуса ACTN3 у индивидуума. В данной области известны различные формы амплификации, и любой такой известный способ можно применять. Как правило, амплификация включает в себя применение одного или нескольких праймеров, избирательно или специфически гибридизующихся с представляющей интерес последовательностью нуклеиновой кислоты для амплификации. Праймеры Как определено здесь, подразумевают, что термин праймер включает в себя любую нуклеиновую кислоту, способную запускать синтез вновь возникающей нуклеиновой кислоты в зависимом от матрицы процессе. Как правило, праймеры представляют собой олигонуклеотиды длиной от десяти до двенадцати пар оснований, но можно применять более длинные последовательности. Праймеры можно предоставлять в двухцепочечной или одноцепочечной форме, но одноцепочечная форма предпочтительна. Способы конструирования праймеров хорошо известны в данной области и основаны на конструировании комплементарных последовательностей, полученных исходя из стандартного образования пар по Уотсону-Крику (т.е., связывание аденина с тимином или урацилом и связывание гуанина с цитозином). Специалистам в данной области хорошо известны доступные из коммерческих источников и/или публичных источников компьютеризированные программы для выбора и конструирования праймеров для амплификации. Конкретные последовательности праймеров, пригодные для определения прогнозирующих спортивные способности генетических вариантов, таких как SNP 1747 C>T в ACTN3, представлены в следующих ниже примерах. Специалисту в данной области будет понятно, что представленные конкретные последовательности являются только иллюстративными и что в практическом осуществлении способов по изобретению можно применять альтернативные последовательности праймеров и/или зондов. Способы амплификации Для амплификации маркерной последовательности, присутствующей в данном образце, доступен ряд зависимых от матрицы способов. Одним из лучших известных способов амплификации является полимеразная цепная реакция (обозначаемая как ПЦР), подробно описанная в патентах США Один из вариантов осуществления изобретения может включать в себя получение подходящего образца от индивидуума и выявление конкретной матричной РНК, такой как мРНК ACTN3. Иллюстративный пример для применения данного способа представляет собой образец мышечной ткани (например, образец биопсии мышечной ткани, такой как пункционная биопсия). Когда образец ткани получен, его стандартными способами можно подготовить для выделения нуклеиновой кислоты (например, выделение отдельных клеток, расщепление наружных мембран, выделение мРНК с применением Oligo dT и т.д.). Выделение мРНК также можно проводить с применением известных в данной области наборов (Pierce, AP Biotech, etc). Для определения количества амплифицированной мРНК можно провести процедуру амплификации ПЦР с обратной транскрипцией. Способы обратной транскрипции РНК в кДНК хорошо известны и описаны в Sambrook et al., 1989. В альтернативных способах обратной транскрипции применяют термостабильные ДНК-полимеразы. Данные способы описаны в WO 90/07641, поданной 21 декабря 1990 года. Другой способ амплификации нуклеиновых кислот представляет собой лигазную цепную реакцию (“LCR”), описанную в European Application В качестве еще одного способа амплификации по настоящему изобретению также можно использовать репликазу Q Также для амплификации нуклеиновых кислот по настоящему изобретению, может использоваться способ изотермической амплификации, в котором для достижения амплификации молекул-мишеней, содержащих в одной цепи участка рестрикции нуклеотид-5′-[альфа-тио]-трифосфаты, применяют рестрикционные эндонуклеазы и лигазы (Walker et al., Proc. Natl Acad. Sci. USA 89: 392-396, 1992). Амплификация с вытеснением цепи (SDA) является еще одним способом проведения изотермической амплификации нуклеиновых кислот, которая включает в себя циклы вытеснения цепи и синтеза, т.е. ник-трансляцию. Сходный способ, называемый реакция восстановления цепи (RCR), включает в себя отжиг нескольких зондов в области, предназначенной для амплификации с последующей реакцией восстановления, в которой присутствуют только два из четырех основания. Два других основания для удобства выявления можно добавлять в виде биотинилизированных производных. Сходный подход применяют при SDA. Представляющие интерес специфические последовательности также можно выявлять с применением циклической реакции с зондами (CPR). В CPR, зонд с последовательностями неспецифической ДНК на 3′- и 5′-концах и последовательностью специфической РНК между ними гибридизуют с ДНК, находящейся в образце. После гибридизации проводят обработку РНКазой H, а продукты зонда идентифицируют как отдельные продукты, высвободившиеся после расщепления. Исходную матрицу отжигают с зондом в еще одном цикле и реакцию повторяют. По настоящему изобретению можно применять дополнительные способы амплификации, описанные в GB Application Другие способы амплификации нуклеиновых кислот включают в себя системы амплификации, основанные на транскрипции (TAS), включающие в себя амплификацию, основанную на последовательности нуклеиновой кислоты (NASBA) и 3SR Kwohet al., Proc.Natl Acad. Sci. USA 86: 1173 (1989); Gingeras et al., PCT Application WO 88/10315. В NASBA, нуклеиновые кислоты для амплификации можно получать посредством стандартного выделения с фенол/хлороформом, тепловой денатурации клинического образца, обработкой лизирующим буфером и колонками миниспин для выделения ДНК и РНК или выделением РНК с применением гуанидинхлорида. Данные способы амплификации включают в себя отжиг праймера, направленного к специфическим последовательностям. После полимеризации гибриды ДНК/РНК расщепляют с применением РНКазы H, а двухцепочечные молекулы ДНК снова подвергаются тепловой денатурации. В любом случае одноцепочечные ДНК превращают в двухцепочечные посредством добавления второго специфичного к мишени праймера с последующей полимеризацией. Затем двухцепочечные молекулы ДНК многократно транскрибируют такой полимеразой как T7 или SP6. В изотермической циклической реакции РНК обратно транскрибируют в двухцепочечную ДНК и еще раз транскрибируют такой полимеразой как T7 или SP6. Получающиеся укороченные или полные продукты указывают на присутствие представляющих интерес специфических последовательностей. Davey et al., European Application Miller et al., PCT Application WO 89/06700 описали схему амплификации нуклеиновых кислот, основанную на гибридизации промоторной/праймерной последовательности к представляющей интерес одноцепочечной ДНК (“оцДНК”) с последующей транскрипцией с данной последовательности множества копий РНК. Данная схема не является циклической, т.е., с получающихся в результате РНК-транскриптов не образуются новые матрицы. Другие способы амплификации включают в себя “RACE” и “одностороннюю ПЦР” Frohman, M. A., In: PCR PROTOCOLS: A GUIDE TO METHODS AND APPLICATIONS, Academic Press, N. Y. (1990) и Ohara et al., Proc. Natl Acad. Sci USA, 86: 5673-5677 (1989). На стадии амплификации настоящего изобретения можно также применять способы, основанные на лигировании двух (или более) олигонуклеотидов в присутствии нуклеиновой кислоты с последовательностью получающегося “диолигонуклеотида”, что таким образом амплифицирует диолигонуклеотид (например, Wu et al., Genomics 4: 560 1989). Способы разделения После амплификации может быть желательным отделить амплифицированный продукт от матрицы и избытка праймера для цели определения того, произошла ли специфическая амплификация. В одном из вариантов осуществления продукты амплификации разделяют посредством электрофореза в агарозном, агарозно-акриламидном или полиакриламидном геле с применением стандартных способов (например, Sambrook et al., 1989). Альтернативно для проведения разделения можно применять способы хроматографии. Существует множество видов хроматографии, которые можно применять по настоящему изобретению (Freifelder, 1982). Способы идентификации В данной области известно много способов детекции вариантов последовательностей нуклеиновых кислот и можно использовать любой из таких известных способов. В одном из вариантов осуществления детекцию можно проводить посредством саузерн-блоттинга и гибридизации с меченым зондом. Способы, вовлеченные в саузерн-блоттинг, хорошо известны специалистам в данной области (например, Sambrook et al., 1989). В кратком изложении, продукты амплификации разделяют электрофорезом в геле. Затем гель приводят в контакт с мембраной, такой как нитроцеллюлоза, что обеспечивает перенос нуклеиновой кислоты и нековалентное связывание. Затем мембрану инкубируют с конъюгированным с хромофором зондом, способным к гибридизации с представляющим интерес продуктом амплификации. Детекцию проводят посредством экспозиции мембраны на рентгеновской пленке или устройствах детекции испускания ионов. Один из примеров указанного выше описан в патенте США Способы и устройства для детекции вариантов последовательностей нуклеиновых кислот коммерчески доступны из множества источников, таких как Third Wave, Pyrosequencing, Applied Biosystems, Affymetrix, Sequenom, Nanogen и другие, и любые такие коммерческие системы можно использовать для детекции вариантов последовательностей в ACTN3 или других, связанных со способностями, генах. Белки и пептиды В определенных вариантах осуществления описанные способы могут включать в себя детекцию и/или определение количества конкретного белка (например, ACTN3) в образцах для скрининга. Для удобства термины “белок”, “полипептид” и “пептид” здесь применяют взаимозаменяемо. Хотя в данной области известно множество способов количественного определения белка, которые могут использоваться, особенно предпочтительными для количественного определения белков являются анализы, основанные на антителах, такие как ELISA. Специалистам понятно, что описание, приведенное ниже, является только иллюстративным и что для выявления/подсчета количества белков можно применять любые известные способы. В определенных вариантах осуществления белок или пептид может быть выделен или очищен. Способы очистки белков хорошо известны специалистам в данной области. Данные способы на первом уровне включают в себя гомогенизацию и грубое фракционирование клеток тканей и органов на белковую и небелковую фракции. Затем интересующий белок или полипептид можно очищать с применением хроматографических или электрофоретических способов для достижения частичной или полной очистки (или очистки до гомогенности). Аналитическими способами, наиболее подходящими для получения чистого белка, являются ионообменная хроматография, гель-проникающая хроматография, ВЭЖХ (высокоэффективная жидкостная хроматография), FPLC (AP Biotech), электрофорез в полиакриламидном геле, аффинная хроматография, иммуноаффинная хроматография и изоэлектрическое фокусирование. Пример очистки рецепторного белка посредством аффинной хроматографии описан в патенте США Очищенный белок или пептид предназначен для обозначения композиции, отделяемой от других компонентов, где белок или пептид очищены до любой степени, относительно их состояния при получении из природного источника. Следовательно, выделенный или очищенный белок или пептид также относятся к белку или пептиду, лишенному окружения, в котором они находятся в природе. Как правило “очищенный” относится к композиции белка или пептида, подвергнутой фракционированию для удаления других различных компонентов, и где композиция по существу сохраняет свою проявляемую биологическую активность. Когда применяют термин “по существу очищенный”, то данное обозначение относится к композиции, в которой белок или пептид образуют главный компонент композиции, такой как содержание в композиции приблизительно 50%, приблизительно 60%, приблизительно 70%, приблизительно 80%, приблизительно 90%, приблизительно 95% или более белков. В определенных вариантах осуществления описанные способы могут включать в себя очистку одного или нескольких белков или пептидов. Может быть полезным, когда для выделения молекул и последующего выявления или подсчета количества молекул в образце молекул применяют такую очистку образца белка или ДНК как магнитные бусы (Dynal, Dyna beads). Такие способы хорошо известны специалистам в данной области. Антитела В определенных вариантах осуществления может быть желательным получение антитела к конкретным интересующим белкам или пептидам (например, ACTN3). Соответствующий белок или его части можно конъюгировать или химически присоединять к одному или нескольким агентам для усиления их иммуногенности, что хорошо известно в данной области. Предпочтительными агентами являются носители, такие как гемоцианин морского блюдца (KLH) или бычий сывороточный альбумин (BSA). В одном из вариантов осуществления выявление представляющего интерес белка можно проводить вестерн-блоттингом или иммуноцитохимически с применением одного или нескольких специфических антител ко всему интересующему белку (например, ACTN3) или его части со специфическим антителом или его фрагментом (например, Fab-фрагмент или рекомбинантный фрагмент антитела, такой как scFv). Один из примеров антител, которые могут применяться, является антитело против ACTN3 (как описано у North, K.N. et al., Neuromuscular Disorders, 6, 229-235, 1996). В другом варианте осуществления уровень интересующего белка можно определять получением образца от индивидуума (например, мышечная биопсия) и обработкой образца одним или несколькими антителами, направленным на интересующий белок. Термин “антитело” применяют для обозначения любой подобной антителу молекулы, с антигенсвязывающей областью и включает в себя фрагменты антител, такие как Fab’, Fab, F (ab’)2, антитела с одним доменом (DABs), Fv, scFv (одноцепочечные Fv) и тому подобное. Способы получения и применения различных основанных на антителах конструкций и фрагментов хорошо известны в данной области. Средства для получения и характеристики антител также хорошо известны в данной области (см., например, Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988; включенный сюда в качестве ссылки). ELISA В определенных вариантах предпочтительных осуществлений количество интересующего белка, такого как ACTN3, можно определять различными типами твердофазных иммуноферментных анализов (ELISA) или радиоиммуноанализов (RIA), известных в данной области. Также особенно эффективным является использование иммуногистохимического определения с использованием тканевых срезов. Однако понятно, что определение не ограничено такими способами, и также можно применять вестерн-блоттинг, дот-блоттинг, анализы FACS и тому подобное. В одном иллюстративном ELISA, антитела, связывающиеся с интересующими белками (например, ACTN3), иммобилизуют на выбранной поверхности обладающей аффинностью к белкам, такой как лунка в планшете для микротитрования. Исследуемое соединение, в котором предполагают наличие белка или части белка, вносят в лунку. После связывания и отмывки для удаления неспецифически связанных иммунных комплексов можно выявлять связанный антиген (интересующий белок). Выявление, как правило, проводят путем добавления второго специфичного к интересующему белку антитела, которое связано с детектируемой меткой. Данным видом ELISA является “сэндвич ELISA”. Выявление также можно проводить добавлением второго антитела с последующим добавлением третьего антитела, аффинно связывающегося со вторым антителом, где третье антитело связано с детектируемой меткой. В другом иллюстративном примере ELISA образцы, в которых предполагают наличие белка (антигена), иммобилизуют на поверхности лунки, а затем приводят в контакт с антителами по изобретению. После связывания и отмывки для удаления неспецифически связавшихся иммунных комплексов выявляют связавшийся антиген. Когда исходные антитела связаны с детектируемой меткой, иммунные комплексы можно выявлять непосредственно. Альтернативно, иммунные комплексы можно выявлять с применением второго антитела, аффинно связывающегося с первым антителом, где второе антитело связано с детектируемой меткой. Другой вид ELISA, в котором иммобилизуют белки или пептиды, включает в себя применение в выявлении конкуренции антител. В данном виде ELISA в лунки добавляют меченые антитела, оставляют им связываться с интересующим белком и выявляют их посредством их метки. Затем в неизвестном образце определяют количество представляющего интерес антигена посредством смешивания образца с мечеными антителами перед или во время инкубации с покрытыми лунками. Присутствие интересующего антигена в образце уменьшает количество доступного для связывания с лунками антитела и, таким образом, уменьшает окончательный сигнал. Это применимо для выявления антител в неизвестном образце, где с покрытыми антигеном лунками связываются немеченые антитела и также уменьшают количество доступного для связывания с мечеными антителами антигена. При покрытии планшета антигеном или антителом, как правило, лунки планшета инкубируют с раствором антигена или антитела или в течение ночи или в течение указанного количества часов. Затем лунки планшета отмывают для удаления не полностью адсорбированного вещества. Любые оставшиеся доступные поверхности лунок затем “покрывают” неспецифическим белком, т.е. антигенно нейтральным по отношению к тестовой антисыворотке. Данные белки включают в себя бычий сывороточный альбумин (BSA), казеин и растворы молочного порошка. Покрытие позволяет блокировать участки неспецифической адсорбции на иммобилизующей поверхности и, таким образом, уменьшить фон, вызываемый неспецифическим связыванием антисыворотки с поверхностью. При ELISA обычно применяют средства вторичного или третичного выявления, а не прямую процедуру. Таким образом, после связывания белка или антитела с лункой, покрытием неактивным веществом для уменьшения фона и отмывки для удаления несвязавшегося вещества, иммобилизующую поверхность приводят в контакт с контрольным биологическим образцом для тестирования в подходящих условиях, при которых происходит образование иммунных комплексов (антиген/антитело). Затем для выявления иммунного комплекса необходим меченый второй связывающийся лиганд или антитело, или второй связывающийся лиганд или антитело вместе с меченым третьим антителом или третьим связывающимся лигандом. “В подходящих условиях, при которых происходит образование иммунных комплексов (антиген/антитело)” означает, что условия предпочтительно включают в себя разведение антигенов и антител такими растворами как BSA, бычий гамма-глобулин (BGG) и фосфатно-солевой буфер (PBS)/Tween. Данные добавленные средства также способны снижать неспецифический фон. “Подходящие” условия означают, что инкубацию проводят при температуре и в течение периода времени, достаточных для обеспечения эффективного связывания. Стадии инкубации, как правило, проводят в течение приблизительно от 1-2 до 4 часов, при температуре, предпочтительно порядка от 25°C до 27°C, или можно проводить в течение ночи приблизительно при 4°C, или около. После всех стадий инкубации ELISA, контактную поверхность отмывают так, чтобы удалить не участвующий в образовании комплексов материал. Предпочтительный способ отмывки включает в себя отмывку таким раствором как PBS/Tween или боратный буфер. После образования специфических иммунных комплексов между тестируемым образцом и исходно связанным веществом и последующей отмывки можно определить наличие даже небольших количеств иммунных комплексов. Для проведения детекции второе или третье антитело должны иметь связанную метку, обеспечивающую обнаружение. Предпочтительно, им является фермент, вызывающий окрашивание при инкубации с соответствующим хромогенным субстратом. Таким образом, например, желательно осуществлять взаимодействие первого или второго иммунных комплексов или инкубировать их с антителами, конъюгированными с уреазой, глюкозооксидазой, щелочной фосфатазой или пероксидазой водорода, в течение времени и при условиях, благоприятствующих образованию дополнительных иммунных комплексов (например, инкубация в течение 2 часов при комнатной температуре в содержащем PBS растворе, таком как PBS-Tween). После инкубации с меченым антителом и последующей отмывки для удаления несвязавшегося материала, определяют количество метки, например, инкубированием с хромогенным субстратом, таким как мочевина и бромкрезоловый пурпурный или 2,2′-азидоди-(3-этилбензтиазолин-6-сульфоновая кислота [ABTS] и H2O2, в случае пероксидазы в качестве ферментной метки. Затем проводят количественное определение посредством измерения степени образования окраски (например, с применением спектрофотометра в видимом спектре). Наборы В дополнительных вариантах осуществления настоящее изобретение относится к наборам для определения в способах выявления аминокислот или иммунодетекции, описанных выше. В зависимости от типа применяемого анализа набор может содержать одну или несколько пар праймеров для амплификации интересующей последовательности нуклеиновой кислоты, один или несколько зондов, таких как меченые зонды, для определения генетического варианта и один или несколько контрольных последовательностей-мишеней для подтверждения условий амплификации и/или связывания зонда. Контроли могут включать в себя, например, специфические последовательности-мишени для каждого аллеля SNP 1747 C>T ACTN3. Зонды также могут являться специфичными для гибридизации с аллелями SNP 1747 C>T. Также можно включать сюда различные другие пригодные реагенты, такие как буфер, нуклеотиды и полимераза. В наборах для иммуноанализа белка наборы для иммунодетекции могут содержать в пригодных контейнерах интересующий белок или пептид или первое антитело, связывающееся с интересующим белком или пептидом и реактив для иммунодетекции. Наборы могут содержать первое антитело, специфичное к интересующему белку или пептиду, и меченое второе антитело, специфичное к первому антителу. Альтернативно, наборы могут содержать первое и второе антитело, специфичные или селективные для интересующего белка, где меченым является второе антитело. Альтернативно, первое и второе антитело могут являться немечеными, и можно включать сюда третье антитело, специфичное ко второму антителу. Также сюда можно включать другие стандартные реактивы, такие как буфер или различные вещества или реактивы, применяемые для проявления меченого антитела. Контейнеры наборов, как правило, включают в себя, по меньшей мере, одну ампулу, пробирку, флакон, колбу, шприц или другой контейнер, в который можно поместить образец и, предпочтительно, соответствующим образом разделить на аликвоты. Когда предоставлены второй или третий лиганд для связывания или дополнительный компонент, набор также, как правило, содержит второй, третий или другой дополнительный контейнер, в который можно поместить данный лиганд или компонент. Такие наборы могут содержать контейнеры для инъекций или пластиковые изготовленные выдувным формованием контейнеры, в которых хранятся необходимые пробирки Осуществление тестирования В определенных вариантах осуществления используемые способы скрининга могут включать в себя в дополнение к анализу ACTN3 один или несколько тестов для исследования различных показателей. Такие тестирования показателей можно применять в сочетании, например, с тестированием SNP ACTN3 или анализами белка или мРНК ACTN3. Ниже описаны различные иллюстративные тестирования различных показателей. Специалистам в данной области будет понятно, что данные примеры не являются ограничивающими, и можно применять любой тест, известный в данной области. Тестирование VO2 max Тестирование VO2 max обеспечивает прямое измерение физиологического потенциала спортсменов. Скорость максимального потребления кислорода в условиях интенсивной тренировки определяют хорошо известными в данной области способами. Параметры включают в себя пороги аэробного и анаэробного обмена, ритм и частоту сердечных сокращений, дыхательные параметры, максимальную частоту сердечных сокращений и области сердечных сокращений. Тестирование порога анаэробного обмена (лактат крови и легочная вентиляция) Порогом анаэробного обмена является предел тренировки, при котором образование молочной кислоты находится в равновесии с ее устранением. Данная интенсивность эквивалентна 60-120 минутам бега или езды на велосипеде в зависимости от состояния, техники и опыта. Тест проводят одновременным измерением легочной вентиляции, а также уровня лактата в крови. Хотя способы определения легочной вентиляции и лактата крови дают очень сходные результаты, вместе они определяют порог анаэробного обмена более точно. Предоставляемая данным тестом информация включает в себя порог лактата крови и порог легочной вентиляции, частоту сердечных сокращений на пороге анаэробного обмена и скорость (бег) или ватты (езда на велосипеде) на пороге анаэробного обмена. Тестирование анаэробной силы и мощности (тест Wingate) Тест Wingate определяет силу и мощность ног и предназначен для спортсменов силовых видов спорта. Тест представляет собой 30-секундную максимальную нагрузку на велоэргометре, определяющую пиковую мощность и способность противостоять утомлению. Данные, полученные из теста Wingate, включают в себя: (30 с тест) пиковую мощность (ватты), абсолютную, относительную и индекс усталости (как быстро падает сила в течение 30 с теста) и работу (джоули) (расход энергии). Критическая мощность (CP) Целью тестов CP является определение того, каков оптимальный объем работы, который может выдерживать спортсмен для данного временного периода или дистанции. Наиболее общие тесты CP могут включать в себя временной интервал CP (60-180 с), зависящий от вида спорта, и время попытки CP. Основной обмен (RMR) RMR часто также обозначают как расход энергии в покое (REE). Он представляет собой неинвазивный способ определения минимального количества калорий (ккал), потребляемых индивидуумом в день. Чем выше RMR, тем больше калорий сжигает индивидуум. Результаты напрямую измеряют посредством поглощения и выделения O2 и CO2. Один из тестовых протоколов состоит из запрета питания или приема алкоголя в течение 12 часов, запрета таких стимуляторов как кофе в течение 24 часов и запрета нагрузок в течение 24-36 часов. Тест наиболее часто рекомендуют проводить рано утром. Индивидуума присоединяют к устройству измерения метаболизма в течение 30 минут, в то время как он лежит на спине в спокойном состоянии. В течение теста индивидуум выдыхает в устройство измерения метаболизма через мундштук и присоединенный шланг. После окончания данного теста получают следующую информацию: обмен веществ (RMR) – ккал/сутки Тестирование скорости/мощности Тестирование скорости/силы наиболее часто состоит из трех тестов: скорость бега: инфракрасные стробоскопы (5-50 метров); прыжок вверх и сила ног: устройство Vertec и тесты скорости: стандартные и специфичные для вида спорта. Данные тесты помогают при анализе способностей индивидуума, например, в силовых видах спорта. Тестирование силы/гибкости Тестирование силы/эластичности состоит из силы RM (покоящаяся мышца): приседания со штангой, скамья, тяга штанги, жим ногами; мышечной выносливости: частые повторения определенного веса; олимпийские виды тяжелой атлетики: подъем на грудь и толчок, рывок, силовые подъемы на грудь, силовые рывки; гибкость: стандартные и специфичные для вида спорта и сила мышц брюшного пресса и сила нижнего отдела спины. Композиция тела Тестирование композиции тела может состоять из тестирования кожных складок циркулем (сжатие кожи в некоторых участках на теле, таких как подмышка, бедро и т.д.) и расчета процента содержания жира в теле, а также подсчета сухой мышечной массы и массы жира. Другой способ включает в себя погружение в воду в ванну на выдохе. Содержание жира в теле измеряют посредством специального изменяющего устройства, определяющего вытеснение воды. Применимость способов Хотя описанные способы особенно пригодны для прогнозирования спортивных способностей в скоростно-силовых видах спорта и видах соревнований у европеоидов, данные способы также могут являться пригодными для применения в любой другой этнической группе, как правило, демонстрирующей высокую частоту (т.е. предпочтительно, по меньшей мере, 5%, более предпочтительно – по меньшей мере, 10% и наиболее предпочтительно – по меньшей мере, 15%) генотипа 577XX. После анализа большого количества европеоидов и некоторых других этнических групп представляется, что если у конкретного спортсмена, такого как зулус или определенные европеоидные женщины, отсутствует нуль-генотип, то это коррелирует с потенциалом стать высококлассным спортсменом в скоростно-силовых видах спорта в отличие от выносливого спортсмена. Например, нуль-генотип является частым в популяции коренных американцев (29%), азиатской популяции (25%) и у белых европейцев (20%), у горцев PNG (15%), в афроамериканской популяции (13%) и в популяции аборигенов Австралии (10%). Способы по настоящему изобретению можно применять в программах поиска талантливых индивидуумов отдельно или в сочетании с аналогичными способами генотипирования индивидуумов по другим генам, связанным со спортивными способностями. Другие способы, которые можно сочетать с описанными здесь способами, основываются на данных о способностях и фенотипических прогнозах (например, рост и телосложение) и т.д. Таким образом, результаты способов по настоящему изобретению можно применять для выбора или, по меньшей мере, для помощи в выборе, молодых индивидуумов с большим спортивным потенциалом и/или предоставления правил по выбору типа спорта, который молодой индивидуум может выбрать для специализации. В другом осуществлении, основываясь на знании генетических факторов, предсказывающих способности индивидуума к тренировкам (например, уровни белка или мРНК ACTN3 и/или определение SNP), можно продумывать тренировочные программы для потенциального или действующего высококлассного спортсмена, имеющего высокие шансы на успех. Индивидуальные тренировочные программы могут быть направлены на конкретные способности (определенные на основании генетического состава) посредством определения типа тренировок, развитие которых может наиболее вероятно привести к успеху. Это могло бы помочь сократить небольшой интервал между успехом и неудачей у спортсменов высокого уровня, избежать ненужной усталости от избыточных тренировок без ожидаемого успеха (например, их не позволит добиться генетический потенциал); снизить растрачиваемые ресурсы и преждевременные “перегорания”; и может увеличить интервал для достижения долгосрочных целей и повысить самооценку отдельного спортсмена. Ресурсы расходуются каждый раз, когда индивидуума с высоким спортивным потенциалом удаляют из программы по причине того, что он/она не смог/ла достигнуть успеха. На индивидуальном уровне усилия и жертвы, уже предпринятые такими индивидами, могут неблагоприятно влиять на его жизненные цели и самооценку. В этой ситуации, знание генетического состава, одного или в сочетании с другими прогностическими факторами, может объяснить причину отсутствия результата и поможет указать индивиду наиболее реальные жизненные задачи, которые могут включать в себя более подходящий вид спорта. Следовательно, в одном из вариантов осуществления, определение улучшенных тренировочных программ для спортсмена может включать в себя определение конкретного генотипа интересующего гена (например, генотип ACTN3) у спортсмена. Другой пример разработки тренировочной программы для потенциального или настоящего спортсмена может включать в себя сочетание одного или нескольких тестов интересующей молекулы с другими тестами оценки параметров, как указывалось ранее, и анализ результатов двух или более тестов для разработки программы. ПРИМЕРЫ Пример 1: Скрининг на выявление нуль-генотипа (577XX) ACTN3 у высококлассных спортсменов Материалы и методы Человеческую геномную ДНК выделяли из крови группы высококлассных спортсменов (108 выносливых спортсменов и 83 спринтера), 88 индивидуумов африканских зулусов и 152 контрольных индивидуумов австралийских европеоидов, посредством способа экстракции с фенолом:хлороформом после лизиса клеток посредством Triton-X100 и расщепления протеиназой K. С геномной ДНК амплифицировали экзон 16 ACTN3. Праймеры, соответствующие смежным интронным для экзона 16 последовательностям представляли собой: прямой 5’CTGTTGCCTGTGGTAAGTGGG3′ (SEQ ID обратный 5’TGGTCACAGTATGCAGGAGGG3′ (SEQ ID Цикл реакции ПЦР для данных праймеров представлял собой: 35 циклов при 94°C в течение 30 сек, а затем 72°C в течение 1 мин, с заключительным удлинением при 94°C в течение 10 мин. Аллели R577X (кодоны CGA и TGA, соответственно) можно различить по присутствию (577X) или отсутствию (577R) в экзоне 16 участка рестрикции Dde I (C Результаты и обсуждение Результаты анализов генотипирования представлены в таблице 2. Генотипирование ACTN3 проводили у высококлассных спортсменов (т.е. индивидуумов, характеризующихся высшими уровнями в показателях выносливости, скорости и/или силы). В сравнении с контролем, высококлассные спринтеры обладали меньшей частотой нуль-мутации 577XX ACTN3 (6% в сравнении с 18% в контрольной европеоидной популяции; p<0,05), подобно тенденции, наблюдаемой в популяции зулусов. Так как представляется, что способность мышечных волокон типа 2 генерировать усилие с высокой быстротой, скоростью и темпом движений, и способность индивидуума адаптироваться к тренировкам, очень подвержены генетическому влиянию, полагают, что генотип ACTN3, вероятно, является фактором, влияющим на нормальное изменение функции мышц в генеральной совокупности. Основываясь на данных результатах, показано, что генотипирование ACTN3 обладает значительным потенциалом для выбора или, по меньшей мере, помощи в выборе молодых индивидуумов с большим спортивным потенциалом. Пример 2 Методы Посредством применения описанного Mills et al. (2001) способа генотипирования генотипировали 436 неродственных европеоидных контролей из трех различных источников (150 доноров крови, 71 здоровый ребенок, принимающий участие в несвязанном исследовании, и 215 здоровых взрослых, принимающих участие в программе выявления талантов, совместно с австралийским институтом спорта). Пол являлся известным для 292 контролей женского пола и 134 контролей мужского пола. Генотипировали 429 европеоидных спортсменов из 14 различных видов спорта. Для целей примера спортсменов определяли как “высококлассных”, если они представляли Австралию в их видах спорта на международном уровне; 50 из спортсменов принимали участие в Олимпийских играх. При наличии локализации Для тестирования равномерности частот аллелей и генотипов ACTN3 в группе спортсменов и контрольной группе применяли подход логарифмического моделирования, как описано у Huttley и Wilson (2000), применяемый в языке статистического программирования R (версия 1.6.2), посредством применения пакета (предоставлен J. Maindonald; доступен на веб-сайте The R Project for Statistical Computing). Для сравнений спортсменов и контролей с применением количеств генотипов рассчитывали значения “X” 2. Профили генотипирования трех контрольных групп (150 доноров крови, 71 здоровый ребенок и 215 здоровых взрослых) значимо не отличались ни друг от друга (x2=0,19; P=0,996), ни от ранее генотипированной группы из 107 белых европейцев (Mills et al. 2001), что позволяло предположить, что частоты генотипов в контрольной группе являются репрезентативными для широкой популяции европеоидов. Частоты генотипов ACTN3 у контрольных субъектов мужского и женского полов значительно не варьировали, а в полной группе не наблюдали значимого отклонения от равновесия Харди-Вайнберга (H-W). Данные генотипирования ACTN3 контрольной группы, группы “скорость/сила” и группы “выносливость” обобщены в таблице 2 и на чертеже. Существенных отличий в частоте аллелей и генотипов между группой высококлассных спортсменов как целое и контролями не наблюдали. Однако когда спортсменов разделили на группы “спринт/сила” и “выносливость” и сравнили с контролями, выявили веское доказательство изменения аллельной частоты (x2 В общем, также существовало свидетельство вариации генотипов, не объясняемое различиями в аллельной частоте (x2 Данные сведения позволяют предположить, что аллель ACTN3 577R обеспечивает преимущество для силового и спринтерского видов активности. Ни у одной из высококлассных спортсменок-спринтеров в выборке не наблюдали дефицита Белок Если генотип 577XX усиливает способности к выносливости, а аллель 577R, как представляется, усиливает спринтерские способности, тогда аллели 577R и 577X могут сохраняться в популяции, так как они оба несут селективные преимущества в различных условиях окружающей среды и, таким образом, посредством балансирования отбора сохраняют высокие популяционные частоты. Пример 3 На чертеже в виде гистограммы представлены данные о частоте генотипов ACTN3 у контролей, высококлассных спортсменов в области спринта/силы и выносливых спортсменов. В сравнении со здоровыми европеоидными контролями существует заметное уменьшение частоты генотипа ACTN3 577XX (ассоциированного с недостатком Специалистам в данной области понятно, что в изобретении можно сделать многочисленные отклонения и/или модификации от представленного в конкретных осуществлениях без отклонения от сущности или объема изобретения, описанного в общих чертах. Следовательно, настоящие варианты осуществления необходимо во всех отношениях рассматривать как иллюстративные, а не ограничивающие. Все описанные и представленные здесь в формуле изобретения КОМПОЗИЦИИ, СПОСОБЫ И УСТРОЙСТВА в свете настоящего описания получены и осуществлены без чрезмерного экспериментирования. Хотя композиции и способы по настоящему изобретению описаны на основании предпочтительных вариантов осуществлений, специалистам в данной области понятно, что можно применять КОМПОЗИЦИИ, СПОСОБЫ И УСТРОЙСТВА и на стадиях или в последовательностях стадий описанных здесь способов без отклонения от концепции, сущности и объема изобретения. Более конкретно, очевидно, что определенные агентами, которые близки химически и физиологически, можно заменять описанные здесь агенты с получением таких же или аналогичных результатов. Специалистам в данной области понятно, что все такие сходные заместители и модификации полагают входящими в сущность, объем и концепцию изобретения, как определено приложенной формулой изобретения.
Формула изобретения
1. Способ прогнозирования спринтерских, силовых, а также способностей к выносливости у индивидуума, включающий: 2. Способ по п.1, где генетической вариацией является однонуклеотидный полиморфизм (SNP) 1747 С>Т в гене ACTN3. 3. Способ по п.1, подпункт b), iv), где индивидуумом является женщина.
РИСУНКИ
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 517
517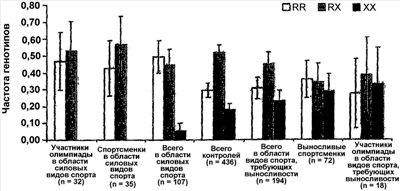
 -актинин-3 (ACTN3). В конкретном осуществлении, генотип ACTN3 определяют для участка однонуклеотидного полиморфизма 1747 C>T.
-актинин-3 (ACTN3). В конкретном осуществлении, генотип ACTN3 определяют для участка однонуклеотидного полиморфизма 1747 C>T. 1-интегрин (Hance, J. E. et al., Archives of Biochemistry & Biophysics, 365, 216-222, 1999 и Otey, C. A. et al., Journal of Biological Chemistry, 268, 21193-21197, 1993). Данные исследования связывания позволяют предположить, что
1-интегрин (Hance, J. E. et al., Archives of Biochemistry & Biophysics, 365, 216-222, 1999 и Otey, C. A. et al., Journal of Biological Chemistry, 268, 21193-21197, 1993). Данные исследования связывания позволяют предположить, что  или другими известными способами амплификации или получением библиотек, таких как BAC, YAC, космидные, плазмидные или фаговые библиотеки, содержащие представляющую интерес нуклеиновую кислоту (См., например, Berger and Kimmel, 1987; Sambrook et al., 1989.). Источник нуклеиновой кислоты для целей анализа не важен и в рамках изобретения полагают, что анализировать можно нуклеиновые кислоты практически из любого источника.
или другими известными способами амплификации или получением библиотек, таких как BAC, YAC, космидные, плазмидные или фаговые библиотеки, содержащие представляющую интерес нуклеиновую кислоту (См., например, Berger and Kimmel, 1987; Sambrook et al., 1989.). Источник нуклеиновой кислоты для целей анализа не важен и в рамках изобретения полагают, что анализировать можно нуклеиновые кислоты практически из любого источника. интенсивность дыхания (RR), интенсивность дыхательного обмена (RER), вентиляция и частота сердечных сокращений в покое, % утилизации углеводов и жиров в покое.
интенсивность дыхания (RR), интенсивность дыхательного обмена (RER), вентиляция и частота сердечных сокращений в покое, % утилизации углеводов и жиров в покое. TNAG). Продукты ПЦР 577R (дикого типа) состоят из фрагментов длиной 205 п.н. и 86 п.н; тогда как продукты ПЦР 577X состоят из фрагментов длиной 108 п.н., 97 п.н. и 86 п.н. Расщепленные фрагменты ПЦР разделяют посредством электрофореза в 10% полиакриламидном геле и визуализируют окрашиванием бромистым этидием.
TNAG). Продукты ПЦР 577R (дикого типа) состоят из фрагментов длиной 205 п.н. и 86 п.н; тогда как продукты ПЦР 577X состоят из фрагментов длиной 108 п.н., 97 п.н. и 86 п.н. Расщепленные фрагменты ПЦР разделяют посредством электрофореза в 10% полиакриламидном геле и визуализируют окрашиванием бромистым этидием. [df=5]=23; P<0,001). Выявили значительные различия в аллельной частоте между спринтерами и контролями и для мужчин (x2
[df=5]=23; P<0,001). Выявили значительные различия в аллельной частоте между спринтерами и контролями и для мужчин (x2