Патент на изобретение №2388825
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) ФЕРМЕНТ КАРБОКСИПЕПТИДАЗА КПSВ, ШТАММ Streptomyces bikiniensis – ПРОДУЦЕНТ КАРБОКСИПЕПТИДАЗЫ КПSВ, ФРАГМЕНТ ДНК SB27-995, КОДИРУЮЩИЙ СИНТЕЗ ЗРЕЛОЙ ФОРМЫ ЭТОГО ФЕРМЕНТА, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА КАРБОКСИПЕПТИДАЗЫ КПSВ
(57) Реферат:
Изобретение относится к биотехнологии и представляет собой фермент карбоксипептидазу KПSB. Также изобретение относится к штамму Streptomyces bikiniensis ВКПМ Ас-1783-продуценту карбоксипептидазы KПSB и способу микробиологического синтеза карбоксипептидазы KПSB. Изобретение позволяет расширить арсенал карбоксипептидаз с широкой специфичностью и способных эффективно отщеплять аминокислотные остатки различной природы от С-конца белков и пептидов. 4 н.п. ф-лы, 4 ил., 2 табл.
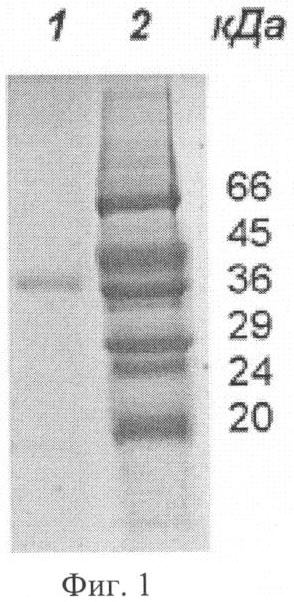
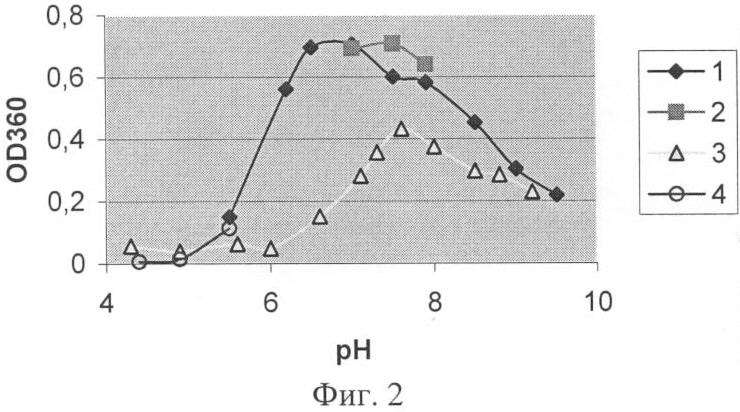
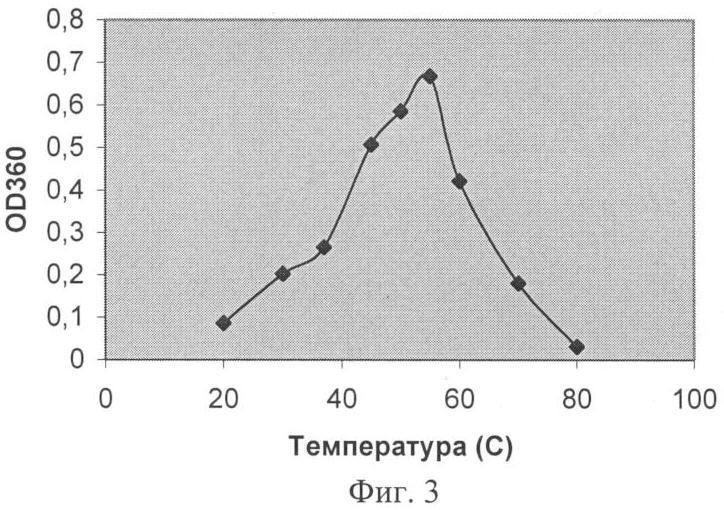
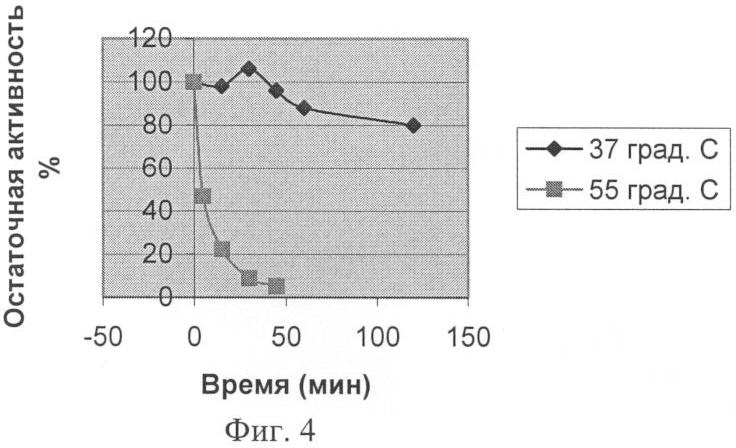
Заявляемая группа изобретений относится к биохимии микроорганизмов и биоорганической химии и касается получения фермента карбоксипептидазы, которая обладает широкой специфичностью в отношении отщепляемого остатка и может быть использована для получения лекарственных средств с терапевтической активностью. При различных заболеваниях желудочно-кишечного тракта человека существенно ухудшается гидролиз белков пищи. В этом случае широко применяют заместительную терапию – использование ферментных препаратов из различных источников для компенсации собственной недостаточности пищеварительных ферментов (Rudmann, 1994, Kreon. Pancreatic enzymes for replacement therapy. Hannover, 1994). Препараты животного происхождения на основе панкреатина перестали отвечать требованию безопасности в связи с появлением новых заболеваний, передающихся от сельскохозяйственных животных человеку. На первый план стали выходить растительные и микробные ферментные препараты (Rudmann, 1994, Kreon. Pancreatic enzymes for replacement therapy. Hannover, 1994). Усвоение белков в кишечнике человека и животных связано с протеолитическим расщеплением этих веществ сначала эндопептидазами с образованием пептидов, а затем экзопептидазами до отдельных аминокислот. При этом аминопептидазы отщепляют аминокислотные остатки с N-, а карбоксипептидазы с С-конца пептидов. Известно большое количество ферментов с карбоксипептидазной активностью. Основываясь на природе каталитического центра, различают металло- и сериновые карбоксипептидазы. Поджелудочная железа человека синтезирует две металлокарбоксипептидазы – карбоксипептидазу А (КПА) и карбоксипептидазу В (КПВ). Согласно современной классификации эти ферменты входят в подсемейство А/В из семейства пептидаз М14 (Handbook of proteolytic enzymes, 1318-1351 (1997)). В это подсемейство входят также многие другие карбоксипептидазы из различных тканей млекопитающих (1989, J.Biol. Chem. 264:20094-9; 1991, J.Biol. Chem. 266:21833-8; 1999, Cancer Res., 59:2981-8; 2002, J.Biol. Chem. 277:14954-64; 2005, Proc. Natl. Acad. Sci. USA 102:3978-83), насекомых (2003, Eur. J. Biochem., 270, 3026-35, 2005, Proc. Natl. Acad. Sci. US A102:16602-7); а также из микроорганизмов (1976, J. Biol. Chem. 251(1): 204-208; 1984, Biokhimiya, 49:292-301). Более половины ферментов, входящих в подсемейство А/В, участвуют в связанном с пищеварением расщеплении белков и пептидов. Отсюда второе название подсемейства А/В – «пищеварительные металлокарбоксипептидазы». Помимо функциональной и структурной классификации, карбоксипептидазы принято разделять по природе отщепляемого ими C-концевого аминокислотного остатка, так называемой первичной субстратной специфичности. КПА из различных источников отщепляет преимущественно гидрофобные (ароматические, или разветвленные алифатические) C-концевые остатки (Handbook of proteolytic enzymes, 1321-1326) (1997). КПВ – только C-концевые положительно заряженные Arg и Lys (Handbook of proteolytic enzymes, 1333-1335 (1997). Среди карбоксипептидаз млекопитающих отсутствуют ферменты с широкой специфичностью по отщепляемому остатку. Ферментные препараты, основанные на панкреатине, содержат смесь двух ферментов КПА и КПВ (Европейская фармакопея, 2001). Карбоксипептидазы микроорганизмов обладают более широкой специфичностью. Так, сериновые карбоксипептидазы, продуцируемые представителями рода Aspergillus, например, та, которая обнаружена в культуральных фильтратах Aspergillus saitoi (EC 3.4.16.1) (1969, Agr. Chem. Soc. Jap. 44:5; 1972, J. Biochem. Tokyo 72:1045-1048; 1973, Biochim. Biophy Acta 293:444-450), освобождает от С-конца субстратов основные, нейтральные и кислые аминокислоты, и даже пролин. Однако использование этих ферментов для улучшения пищеварения малоперспективно в связи с тем, что они функционируют при кислых значениях pH, тогда как содержимое кишечника имеет нейтральную или слабощелочную реакцию. Широкая специфичность показана для карбоксипептидазы Т (КПТ) из Thermoactynomyces vulgaris (1991, FEBS Lett. 291:75-8; 2002, Вопр. Мед. Хим., 48:577-85). Она способна отщеплять гидрофобные, положительно и отрицательно заряженные C-концевые остатки. Вместе с тем, специфичность этого фермента имеет крен в сторону активности КПА. Его способность отщеплять гидрофобные остатки лишь на порядок хуже, чем у КПА. Зато КПТ отщепляет положительные остатки на два порядка медленнее, чем гидрофобные. Скорость отщепления отрицательных остатков этим ферментом также невелика. В качестве ближайшего аналога заявляемого штамма и фермента рассмотрим штамм Streptomyces griseus (S.griseus) К-1 и продуцируемую им карбоксипептидазу КПSG, способную расщеплять субстраты как КПА, так и КПВ (1976, Биохимия 41: 328-334; 1976, J.Biol. Chem. 251:204-208; 1979, J Biochem. 82:615-8). Нуклеотидная последовательность карбоксипептидазы S.griseus K-1 (ACCESSION X65719). В источниках информации отсутствуют сведения о наличии или отсутствии у этого фермента способности отщеплять глутаминовую кислоту. Описание способа культивирования штамма-продуцента S.griseus K-1 в источниках также отсутствует. Имеющиеся в источниках информации данные не дают достаточного материала для того, чтобы составить точное представление о специфичности карбоксипептидазы KПSG. Задачи заявляемой группы изобретений состоят – в расширении арсенала карбоксипептидаз, полученных из микроорганизмов; – в разработке способа получения заявляемой карбоксипептидазы. Задачи решены путем создания заявляемой группы изобретений, которая представляет собой 1. Штамм Streptomyces bikiniensis (S. bikiniensis) ВКПМ Ас-1783-продуцент фермента карбоксипептидазы KПSB. 2. Фермент карбоксипептидаза KПSB, обладающий следующими свойствами: а) способен отщеплять аргинин от субстрата Dnp-Ala-Ala-Arg со следующими кинетическими характеристиками: Км=1,6×10-4 М, kсat=48,8 с-1 и kcat/Км=3,05×105 М-1 с-1; б) способен отщеплять аргинин от субстрата Z-Ala-Ala-Arg со следующими кинетическими характеристиками: Км=5×10-5 М, kcat=52,9 с-1 и kcat/Км=10,6×105 М-1 с-1; в) способен отщеплять лейцин от субстрата Z-Ala-Ala-Leu со следующими кинетическими характеристиками: Км=3×10-5 М, kcat=57,2 с-1 и kcat/Км=19,1×105 М-1 с-1; г) способен отщеплять глутаминовую кислоту от субстрата Z-Ala-Ala-Glu со следующими кинетическими характеристиками: Км=7,6×10-4 М, kcat=5,3 с-1 и kcat/Км=6,97×103 M-1 c-1; д) способен проявлять карбоксипептидазную активность в диапазоне рН от 6 до 9 с рН оптимумом 7,0-7,5; е) способен проявлять карбоксипептидазную активность в диапазоне температур от 30 до 70°C с температурным оптимумом при 55°С; ж) активность фермента КПSВ подавляется ингибиторами металлопротеиназ: ортофенантролином, ЭДТА и ЭГТА, а также реагентами, связывающимися с сульфгидрильной группой: йодацетамидом, двувалентными ионами ртути и меди; з) молекулярная масса фермента КПSВ составляет 36 кДа при определении методом SDS-гельэлектрофореза, и молекулярную массу 36, 699 кДа при определении методом масс-спектроскопии. 3. Фрагмент ДНК SB27-995, кодирующий синтез зрелой формы карбоксипептидазы KПSB и имеющий нуклеотидную последовательность SEQ ID NO 1, либо соответствующую ей нуклеотидную последовательность в силу вырожденности генетического кода. 4. Способ микробиологического синтеза карбоксипептидазы КПSВ, включающий культивирование в подходящих условиях штамма-продуцента этого фермента Streptomyces bikiniensis ВКПМ Ac-1783, либо других штаммов, относящихся к роду Streptomyces, имеющих идентичность не менее 90% с нуклеотидной последовательностью фрагмента ДНК SB27-995, кодирующего синтез зрелой формы карбоксипептидазы КПSВ. Заявляемый штамм S.bikiniensis 27 выделен из почвенных образцов и депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) как Streptomyces bikiniensis ВКПМ Ac-1783. Штамм ВКПМ Ас-1783 идентифицировали как S.bikiniensis методом частичного сиквенса фрагмента гена 16S РНК. Характеристики заявляемого штамма Штамм S.bikiniensis ВКПМ Ас-1783 способен продуцировать карбоксипептидазу КПSВ, которая относится к карбоксипептидазам, обладающим широкой специфичностью в отношении отщепляемого остатка, в количестве до 5 мг/л. Морфологические Относятся к грамположительным бактериям. На среде Гаузе-1 (С.М.Семенов. Лабораторные среды для актиномицетов и грибов. Справочник. Москва, ВО «Агропромиздат», 1990 г., стр.148) штамм образует слегка выпуклые колонии диаметром 3-5 мм. Поверхность колоний гладкая, споры серо-белого цвета, могут быть с экссудатом, по краям возможен узкий (0,5 мм) ободок белого цвета. Края колонии ровные или слегка волнистые. Воздушный мицелий серый, бело-серый. Субстратный мицелий кремовый. Вегетативные гифы (диаметром 0,5-2,0 мкм) образуют разветвленный мицелий. Клетки воздушного мицелия нитевидной формы, спороносцы – прямые, споры – овальные, гладкие. На среде Гаузе-2 (С.М.Семенов. Лабораторные среды для актиномицетов и грибов. Справочник. Москва, ВО «Агропромиздат», 1990 г., стр.152) воздушный мицелий отсутствует. Субстратный мицелий коричнево-бурый. Штамм образует меланоидные пигменты. На среде YSP следующего состава (мас.%): мальт-экстракт 1,5; дрожжевой экстракт 0,5; крахмал растворимый 0,5; СаСО3 0,3; агар 1,8-2,0; вода – остальное, рН 7,0-7,2, штамм образует серо-коричневые колонии диаметром 5-7 мм с гладкой поверхностью и ровными краями. Центр колонии слегка приподнят. Споры серого цвета, субстратный мицелий красно-коричневый. Условия культивирования Оптимальная температура культивирования 28-30°C, оптимальный для роста диапазон рН 6,8-7,2, оптимальное время ферментации составляет 96-120 часов. Посевной материал выращивают на среде YSP, а процесс ферментации осуществляют на среде ПКА-2 следующего состава (мас.%): пептон 0,5; кукурузный экстракт 0,5; глюкоза 1,0; СаСО3 0,05; NaCl 0,5; вода – остальное, рН 7,0-7,2. Хранение штамма Культуру штамма S. bikiniensis ВКПМ Ас-1783 хранят на среде YSP на косяках при температуре 2-4°С или в лиофильно-высушенном состоянии в стеклянных запаянных ампулах при температуре 2-4°С. Способ в общем виде Микробиологический синтез карбоксипептидазы КПSB Способ микробиологического синтеза заявляемой карбоксипептидазы КПSB осуществляют следующим образом. В качестве продуцентов используют штаммы микроорганизмов, относящихся к роду Sreptomyces, способные продуцировать карбоксипептидазу КПSB. Подготовку посевного материала ведут двумя способами. Способ 1. Подготовку спорового посевного материала ведут на агаризованной среде YSP в течение 10-12 суток при температуре 28-30°С. После наступления стадии споруляции культуры производят смыв спор стерильной водой. Способ 2. Подготовку вегетативного посевного материала ведут на жидкой посевной среде (ПС) следующего состава (мас.%): кукурузная мука 2,0 кукурузный экстракт 0,5; СаСО3 0,1; NaCl 0,5; (NH4)2HPO4 0,2; вода – остальное, рН 7,0-7,2. Посевной вегетативный материал выращивают в течение 18-22 часов при температуре 28-30°С в условиях аэрации на качалке при 220-250 об/мин. После наступления стадии споруляции культуры (способ 1) или получения вегетативного мицелия (способ 2) подготовленную культуру высевают в колбы с ферментационной средой, содержащей в качестве источника органического азота пептон либо триптон, либо соевую (кукурузную) муку, либо дрожжевой экстракт. В качестве источника углерода используют глюкозу, либо крахмал, либо мальт-экстракт, либо мелассу. Кроме того, среда содержит неорганические соли Са, Mg, Na, P. Ферментацию проводят в течение 96-120 ч при 28-30°С в условиях аэрации на качалке (220 об/мин). К концу ферментации карбоксипептидаза КПSВ секретируется в культуральную жидкость в количестве от 1 до 5 мг/л. Очистка заявляемого фермента КПSB Очистку заявляемого фермента КПSB из штамма S. bikiniensis ВКПМ Ас-1783 проводят по следующей схеме: (а) с помощью центрифугирования (15000 об/мин) культуральную жидкость отделяют от биомассы; (б) заявляемый фермент КПSB вместе с другими белками, содержащимися в культуральной жидкости, переводят в осадок добавлением сульфата аммония до 80% насыщения; (в) осадок отделяют центрифугированием и растворяют в 10 мМ Трис-НСl буфере (рН 8), содержащем 1 мМ СаСl2 (буфер А); (г) фермент КПSB выделяют из полученного раствора, называемого «раствором секретируемых белков», комбинацией катионообменной хроматографии и различных видов аффинной хроматографии. Все процедуры проводят при 4°С. Карбоксипептидазную активность различных фракций, полученных в ходе выделения фермента, определяют по гидролизу субстрата Dnp-Ala-Ala-Arg. Бацитрацин-силохром, используемый в качестве носителя для аффинной хроматографии, синтезирован ранее (Stepanov, et al., (1981), J. Biochem. Biophys. Methods, 5:177-186). После прохождения раствора секретируемых белков через колонку, заполненную бацитрацин-силохромом, и последующей промывки колонки выделяемый фермент КПSB элюируют 1,0 М раствором хлорида натрия в буфере А. Для очистки КПSB от трипсина, секретируемого штаммом S. bikiniensis ВКПМ Ас-1783, используют аффинную хроматографию на бензамидин-сефарозе (Benzamidine Sepharose 4 FaS Flow, GE Healthcare). Элюат, полученный на предыдущей стадии, пропускают через колонку с бензамидин-сефарозой, уравновешенной буфером А, содержащим 1 М NaCl. Фермент КПSB присутствует в неадсорбируемой фракции. Для катионообменной хроматографии используют SP Sepharose FaS Flow (GE Healthcare). Фракцию, полученную на стадии хроматографии на бензамидин-сефарозе, диализуют против 10 мМ Трис-НСl буфера, рН 7, содержащего 1 мМ СаСl2 (буфер Б), и хроматографируют на колонке, заполненной SP Sepharose. Фермент КПSB элюируют с колонки с помощью градиента концентрации NaCl. Для аффинной хроматографии на аргинин-сефарозе используют Arginine Sepharose 4В (производство GE Healthcare). Элюат, полученный на стадии катионообменной хроматографии, пропускают через колонку с аргинин-сефарозой. Выделяемый фермент элюируют буфером Б, содержащим 0,5 М NaCl. Получение заявляемого фрагмента ДНК SB27-995 Эксперименты по клонированию гена, кодирующего карбоксипептидазу из генома S.bikiniensis ВКПМ Ас-1783, проведены в два этапа. На первом этапе в ходе амплификации вырожденными праймерами получены фрагменты консервативных областей гена; а на втором – клонированы фланговые последовательности гена при помощи ПЦР, осуществляемого со специфичных праймеров, спланированных на основе прочитанной последовательности консервативных участков гена. Заявляемая группа изобретений проиллюстрирована следующими фигурами графического изображения: Фиг.1 – электрофореграмма чистого препарата карбоксипептидазы КПSB. На дорожку 1 нанесено 0,2 мкг чистого препарата фермента КПSB. Дорожка 2 содержит стандарты молекулярной массы. Фиг.2 – диаграмма зависимости активности фермента КПSB от рН. Активность КПSB измеряли при 37°С в течение 10 мин по синтетическому пептидному субстрату Dnp-Ala-Ala-Arg в следующих буферных системах: (1) 50 мМ 1,3 – Бис (трис-(гидроксиметил)-метил-амино) пропан – НСl; (2) 50 мМ Трис-HCl буфер; (3) 30 мМ универсальная буферная система, содержащая лимонную кислоту, KH2РO4; Н3ВО3 и диэтилбарбитуровую кислоту; (4) 50 мМ Na-ацетатный буфер. Фиг.3 – диаграмма зависимости активности фермента КПSB от температуры. Активность КПSB определяли в 50 мМ Трис-НСl буфере (рН 7,6), содержащем 1 мМ CaCl2, с использованием синтетического пептидного субстрата Dnp-Ala-Ala-Arg. Фиг.4 – диаграмма термостабильности фермента КПSB. Активность KHSB после инкубации в течение различных интервалов времени при одной из двух указанных температур (1) – 37°С и (2) – 55°С, определяли при 37°С в 50 мМ Трис-НСl буфере (рН 7,6), содержащем 1 мМ СаСl2, по гидролизу Dnp-Ala-Ala-Arg. Пример 1. Получение карбоскипептидазы КПSB путем культивирования заявляемого штамма Посевной материал, представляющий собой водную суспензию спор штамма S. bikiniensis Ac-1783, выращенного при 30°С в течение 12 суток на среде YSP, засевают в объеме 5% в колбы емкостью 750 мл, содержащие 30 мл ферментационной среды ПКА-2. Колбы помещают на круговую качалку (220 об/мин) и культивируют в течение 96 ч при 30°С. Затем проводят осаждение мицелия путем центрифугирования в течение 20 мин при 6000 об/мин. К 20 мкл центрифугата добавляют к 180 мкл 50 мМ Трис-НСl-буфера, рН 7,6, содержащего 1 мМ СаСl2 (В). (Stepanov V.M. (1995) Methods in enzymology. 248:675-683.) Далее проводят определение ферментативной активности. Для этого к полученной смеси прибавляют 200 мкл водного раствора субстрата Dnp-Ala-Ala-Arg с концентрацией 0,5 мг/мл. Инкубацию проводят 20 мин при 37°С. Реакцию останавливают добавлением 40 мкл 50% уксусной кислоты. В качестве контроля готовят смесь из тех же компонентов с той разницей, что уксусную кислоту добавляют раньше субстрата. Для отделения продукта реакции Dnp-Ala-Ala от оставшегося негидролизованным субстрата Dnp-Ala-Ala-Arg, несущего положительно заряженную группу, реакционную смесь наносят на микроколонку с катионообменником SP-Сефадексом С-25, уравновешенную 1 М уксусной кислотой. Продукт реакции элюируют 1,2 мл 1 М уксусной кислоты. Оптическую плотность элюата измеряют при длине волны 360 нм против контроля и рассчитывают активность (А) карбоксипептидазы КПSB по формуле: А (ед. акт./ мл)=O.D.360×v1 (мл)/ За единицу активности принимают количество фермента, которое в течение 1 минуты катализирует превращение 1 мкМ субстрата в продукт. Уровень карбоксипептидазной активности в культуральной жидкости заявляемого штамма соответствует 0,126 ед. акт./мл. Исходя из удельной активности чистого фермента (см. ниже), это соответствует 1,0 мг/л культуральной жидкости. Пример 2. Очистка заявляемого фермента К 450 мл центрифугата, полученного как в примере 1, прибавляют сульфат аммония до 80% насыщения. Полученную суспензию оставляют на ночь при 4°С, после чего осадок, содержащий фермент, отделяют центрифугированием (15000 об/мин, 40 мин). Полученный осадок растворяют в 40 мл буфера А, перемешивают на магнитной мешалке в течение 1 часа, центрифугируют (15000 об/мин, 40 мин) и диализуют в течение ночи против буфера А. Полученную в результате растворимую фракцию наносят на колонку с бацитрацин-силохромом, уравновешенным буфером А, промывают колонку уравновешивающим буфером и элюируют фракции, содержащие фермент, 1М хлоридом натрия в уравновешивающем буфере. Полученную активную фракцию (60 мл) наносят на колонку с бензамидин-сефарозой, уравновешенную буфером А, содержащим 1М NaCl. Ферментативная активность обнаруживается в составе неадсорбируемой фракции. Фракцию, не адсорбировавшуюся на бензамидин-сефарозе (120 мл), диализуют в течение ночи против 6 л буфера Б. Диализат, содержащий ферментативную активность, наносят на колонку HiTrap SP, уравновешенную буфером Б. Элюцию осуществляют градиентом концентрации NaCl в уравновешивающем буфере. Фракцию, обладающую ферментативной активностью (14 мл), наносят на колонку с Arg-сефарозой, уравновешенную буфером Б, содержащим 100 мМ NaCl. Элюцию проводят тем же буфером, концентрация соли в котором увеличена до 0,5 М. Активность фермента КПSB, полученного при выращивании заявляемого штамма, представлена в Таблице 1.
Из таблицы видно, что в ходе очистки карбоксипептидазы КПSB, синтезированной заявляемым штаммом, удельная активность фермента увеличилась в 180 раз. При этом выход фермента в результате очистки составляет 3,4% от его исходного содержания в культуральной жидкости. Наличие одной полосы при SDS-гельэлектрофорезе (Фиг.1) и единственной аминокислотной последовательности, установленной при N-концевом секвенировании с помощью автоматического метода Эдмана (Asp-Phe-Pro-Ser-Ala-Asp-Ser-Arg-Tyr-His-Asn-Tyr-Ala-Glu-Thr), подтверждает чистоту выделенного фермента Пример 3. Определение молекулярной массы заявляемого фермента SDS-гельэлектрофорез 0,2 мкг препарата чистого фермента (полученного как в примере 2) помещают в лунку 12,5% полиакриламидного геля. Электрофорез проводят при напряжении тока 150 V. В качестве стандартов молекулярной массы используют следующий набор белков: бычий сывороточный альбумин (66 кДа); яичный альбумин (45 кДа); дегидрогеназу фосфоглицеринового альдегида (36 кДа), карбоангидразу (29 кДа); соевый ингибитор трипсина (20,1 кДа). Фиксацию и окрашивание гелей осуществляют кипячением их в 0,25% растворе кумасси R в 7,5% уксусной кислоте, которую используют для отмывки геля от избытка красителя. Как видно из результатов электрофореза (Фиг.1), полученный препарат КПSB не содержит примеси других белков. Молекулярная масса KHSB, определенная этим способом, составляет 36 кДа. Масс-пектроскопия 3 мл образца фермента (0,0016 мг/мл) подвергают высокоэффективной жидкостной хроматографии (ВЭЖХ) на колонке Vydac C4 (4,6×250) (США) в градиенте ацетонитрила 25-60% в течение 40 мин. В результате получают только один белковый пик, что является дополнительным подтверждением чистоты препарата (см. пример 2). Масс-спектр снимают на масс-спектрометре Maldi Masspectrometer Ultimate (Brucer, Germany). Молекулярная масса фермента, определенная этим способом, составляет 36,699 кДа. Пример 4. Определение рН оптимума заявляемого фермента Оптимум рН карбоксипептидазы КПSB определяют с использованием в качестве субстрата Dnp-Ala-Ala-Arg. В качестве буферов используют следующие 50 мМ буфера: Na-ацетатный буфер (рН 4,4-5,5); Бис (трис-(гидроксиметил)-метил-амино) пропан – НСl (5,5-9,5); Трис-НСl буфер (рН 7-7,9), а также 30 мМ универсальный буфер (борная кислота, лимонная кислота, однозамещенный фосфат калия, диэтилбарбитуровая кислота) (рН 4,5-9,2). Все буфера содержат 1 мМ CaCl2. К 20 мкл препарата заявляемого фермента, полученного как в примере 2 (в случае универсального буфера – 50 мкл фермента), добавляют 180 (в случае универсального буфера к 150) мкл буфера с соответствующим значением рН. Затем к полученной смеси прибавляют 200 мкл водного раствора субстрата Dnp-Ala-Ala-Arg с концентрацией 0,5 мг/мл и проводят реакцию в течение 10 мин при температуре 37°С. Данные представлены на Фиг.2. Из полученных данных следует, что фермент обладает активностью в интервале рН 6-9,5 с максимумом в районе 7-7,6 (в зависимости от природы буфера). Несмотря на то что в эксперимент, проводившийся с универсальным буфером, взято в 2,5 раза больше фермента, чем в случае других буферных систем, величины OD360 в интервале рН от 6 до 8,5 в этом случае существенно ниже (линия 3 на Фиг.2). По всей видимости поливалентные анионы, присутствующие в универсальном буфере, ингибируют КПSB, например, за счет связывания ионов кальция. Пример 5. Определение температурного оптимума заявляемого фермента Температурный оптимум определяют, используя в качестве субстрата Dnp-Ala-Ala-Arg. К 190 мкл буфера В добавляют 200 мкл водного раствора субстрата Dnp-Ala-Ala-Arg с концентрацией 0,5 мг/мл и нагревают 2 мин на водяной бане до требуемой температуры. Затем к этой смеси добавляют 10 мкл препарата фермента (полученного как в примере 2) и осуществляют реакцию в течение 10 мин при той же температуре. Данные представлены на Фиг.3. Из них следует, что фермент обладает активностью в ряду температур 30-70°C с максимумом при 55°C. Пример 6. Изучение термостабильности заявляемого фермента Термостабильность КПSB исследуют при температурах 37°С и 55°С с использованием в качестве субстрата Dnp-Ala-Ala-Arg. Для определения термостабильности при 37°С к 150 мкл буфера В прибавляют 100 мкл препарата фермента, полученного как в примере 2, и выдерживают при температуре 37°С. Через интервалы 15, 30, 45, 60, 120 мин отбирают аликвоты по 50 мкл и добавляют к смеси 150 мкл буфера и 200 мкл субстрата, предварительно выдержанных при 37°С в течение 2 мин. Ферментативную реакцию проводят в течение 10 мин при 37°С. Для получения «нулевой точки» 20 мкл раствора фермента прибавляют к смеси 180 мкл буфера и 200 мкл субстрата, предварительно прогретой в течение 2 мин при 37°С. Реакцию проводят, как указано выше. Остаточную активность выражают в виде процента от активности в «нулевой точке». Для определения термостабильности КПSB при 55°С смесь фермента с буфером, приготовленную как описано выше, инкубируют при температуре 55°С. Аликвоты по 50 мкл отбирали через 5, 15, 30 и 45 мин и добавляют к смеси 150 мкл буфера и 200 мкл субстрата, предварительно выдержанных при 37°С в течение 2 мин. Ферментативную реакцию проводят в течение 10 мин при 37°С. Активность фермента в «нулевой точке» определяют так же, как в эксперименте, проводившемся при 37°С. Остаточные активности выражают в виде процента от активности в «нулевой точке». Данные представлены на Фиг.4. Из него следует, что фермент достаточно быстро инактивируется при 55°С. Остаточная активность после 15 и 45 мин инкубации составляет 22,4 и 5,3% соответственно. В то же время фермент стабилен при 37°С. Остаточная активность после 120 минут инкубации составляет 80%. Пример 7. Ингибиторный анализ заявляемого фермента Влияние ингибиторов исследуют, используя в качестве субстрата Dnp-Ala-Ala-Arg. Аликвоту 20 мкл препарата фермента (полученного как в примере 2) прибавляют к 180 мкл буфера В, содержащего один из ингибиторов, представленных в Таблице 2. Инкубацию проводят при комнатной температуре в течение 60 мин. Затем добавляют 200 мкл водного раствора субстрата Dnp-Ala-Ala-Arg с концентрацией 0,5 мг/мл и инкубируют 10 мин при 37°С. Активность фермента в каждом случае выражена в процентах от значения, полученного в отсутствие ингибитора. Из данных, представленных в таблице, можно видеть, что вещества – хелаторы ионов металлов (ортофенантролин полностью, а ЭДТА и ЭГТА в значительной степени) – ингибируют активность фермента. Из этого следует, что карбоксипептидаза КПSB является металлоферментом. Йодацетамид в значительной степени подавляет карбоксипептидазную активность, что говорит о наличие у фермента сульфгидрильной группы, влияющей на его активность. Правда такой реагент на сульфгидрильные группы, как nХМБ, не оказывает ингибирующего действия, но, возможно, это связано со стерическими затруднениями. Из двухвалентных ионов тяжелых металлов только Cu+2 и Hg+2 ингибируют карбоксипептидазу, тогда как ионы марганца, магния и кобальта практически не влияют на ее активность.
Пояснения к примерам 8-11 о субстратной специфичности заявляемого фермента Изучение специфичности фермента КПSB производят на следующих синтетических пептидах с защищенным N-концом: Z-Ala-Ala-Arg, Dnp-Ala-Ala-Arg (субстраты КПВ), Z-Ala-Ala-Leu (субстрат КПА) и Z-Ala-Ala-Glu. Условия реакции отщепления С-концевой аминокислоты от субстратов Z-Ala-Ala-Arg, Dnp-Ala-Ala-Arg, Z-Ala-Ala-Leu и Z-Ala-Ala-Glu следующие: (I) Значение рН равно 7,6; (II) Значение температуры равно 37°С; (III) Количество добавляемого, гомогенно-очищенного фермента составляет 0,06-0,4 мкг/мл (1,7-11,4×10-9 М) в расчете на количество белка. Количество белка определяют колориметрическим способом с применением реактива Бредфорд (производство фирмы «BioRad») и бычьего сывороточного альбумина в качестве стандартного вещества. Для оценки эффективности заявляемого фермента используют методику определения кинетической величины kcat/Km, которая наиболее полным образом характеризует эффективность фермента по отношению к тем или иным субстратам (Fersht A. (1998) Structure and Mechanism in Protein Science: A Guide to Enzyme Catalysis and Protein Folding (Hardcover)). Для нескольких концентраций каждого субстрата изучают ход накопления продукта реакции во времени. Это позволяет вычислить начальную скорость реакции ( Пример 8. Определение каталитических свойств заявляемого фермента в отношении субстрата Dnp-Ala-Ala-Arg Кинетику гидролиза N-замещенного пептида, содержащего на С-конце остаток основной аминокислоты, исследуют на примере субстрата Dnp-Ala-Ala-Arg. 10 мкл фермента с концентрацией 6,81×10-8 М (полученного как в примере 2), смешивают с буфером В и варьирующими концентрациями субстрата (общий объем смеси составляет 400 мкл). В эксперименте используют следующие концентрации субстрата: 0,052; 0,13; 0,26; 0,52, 1,3; 2,6; 5,2 мМ. Реакцию проводят при 37°С в течение следующих интервалов времени: 5, 10, 15, 20, 30 мин. Количество образовавшегося продукта реакции определяют методом, описанным в примере 1. Для каждой концентрации субстрата строят график зависимости O.D.360 от времени реакции, на основании которого определяют значение начальной скорости реакции (
где А – tg угла наклона прямолинейного участка полученной кинетической кривой к оси абсцисс,
«3,0» – фактор, учитывающий изменение объема реакционной среды в ходе измерения активности (см. пример 1). В свою очередь характер зависимости При гидролизе субстрата Dnp-Ala-Ala-Arg заявляемым ферментом определены следующие кинетические константы: Km=1,6×10-4 М, kcat=48,8 с-1, a kcat/Km=3,05×105 М-1 с-1. Эти значения говорят о высокой эффективности выделенного фермента в отношении гидролиза данного субстрата, которая сравнима с эффективностью КПВ. Пример 9. Определение каталитических свойств заявляемого фермента в отношении субстрата Z-Ala-Ala-Arg Кинетику гидролиза N-замещенного пептида, содержащего на С-конце остаток основной аминокислоты, исследуют также с использованием субстрата Z-Ala-Ala-Arg. В необходимый объем буфера В (рассчитанный исходя из того, что суммарный объем реакционной смеси должен составлять 3 мл) вносят варьирующее количество предварительно растворенного в метаноле субстрата. Получают следующие концентрации субстрата: 0,025; 0,035; 0,05; 0,1; 0,2; 0,4 мМ. В каждом случае раствор тщательно перемешивают и помещают в термостатируемую (37°С) кювету спектрофотометра. Через 5 минут в кювету добавляют 200 мкл раствора фермента с концентрацией 2,7×10-8 М, полученного как в примере 2, и регистрируют уменьшение поглощения реакционной смеси при 225 нм во времени. Для каждой концентрации субстрата строят график зависимости изменения поглощения при 225 нм (
где А – тангенс угла наклона прямолинейного участка кинетической кривой к оси абсцисс,
Значения кинетических констант определяют, как в примере 8. В результате были определены следующие кинетические параметры: Km=5×10-5 М, kcat=52,9 с-1, a kcat/Km=10,6×105 М-1 с-1. Они сравнимы с теми, которые получены в предыдущем примере и подтверждают высокую эффективность выделенного фермента в отношении гидролиза субстратов КПВ. Пример 10. Определение каталитических свойств заявляемого фермента в отношении субстрата Z-Ala-Ala-Leu Кинетику гидролиза N-замещенного пептида, содержащего на С-конце остаток гидрофобной аминокислоты, исследуют на примере Z-Ala-Ala-Leu. Реакцию проводят и результаты обсчитывают так, как указано в Примере 9. Используемые концентрации субстрата составляют 0,025; 0,035; 0,05, 0,1 и 0,2 мМ. В результате получены следующие кинетические константы: Km=3×10-5 М, kcat=57,2 с-1, a kcat/Km=19,1×105 М-1 с-1. Полученные параметры говорят о том, что КПSB гидролизует субстрат Z-Ala-Ala-Leu с высокой эффективностью и сравнима по этой характеристике с КПА Пример 11. Определение каталитических свойств заявляемого фермента в отношении субстрата Z-Ala-Ala-Glu Кинетику гидролиза N-замещенного пептида, содержащего на С-конце остаток кислой аминокислоты, исследуют на примере Z-Ala-Ala-Glu. Реакцию проводят и результаты обсчитывают так, как описано в примере 11. Используют следующие концентрации субстрата: 0,2; 1; 2; 4 мМ. Большие концентрации субстрата не были использованы из-за высокого фона при определении оптической плотности. После прединкубации буфера и субстрата в термостатированную кювету добавляют 500 мкл раствора фермента с концентрацией 6,8×10-8, так что концентрация фермента в реакционной смеси составляет 1,14×10-8 М. В результате были получены следующие кинетические параметры: Km=7,6×10-4 М, kcat=5,3 с-1, а kcat/Km=6,97×103 М-1 с-1, которые говорят о том, что карбоксипептидаза КПSB способна гидролизовать субстраты, имеющие на С-конце остаток кислой аминокислоты, хотя и с меньшей эффективностью, чем субстраты, рассмотренные выше. Пример 12. Определение N-концевой последовательности аминокислот заявляемого фермента 2,2 мл образца очищенной карбоксипептидазы КПSB с концентрацией 0,0016 мг/мл хроматографировали на колонке Vydac C4 (4,6×250). Секвенирование образца очищенной карбоксипептидазы производили на автоматическом секвенаторе Procise Sequencing System, model 492 (Applied Biosystems). Была установлена последовательность, состоящая из 15 аминокислотных остатков: Asp-Phe-Pro-Ser-Ala-Asp-Ser-Arg-Tyr-His-Asn-Tyr-Ala-Glu-Thr. Сравнение полученной последовательности с первичной структурой карбоксипептидаз из S. griseus K-1 и S. avermitilis MA-4680 показывает гомологию с участками этих последовательностей, расположенными между остатками 124 и 138. Пример 13. Клонирование фрагмента ДНК SB27-995, кодирующего синтез зрелой формы карбоксипептидазы КПSB Для клонирования гена карбоксипептидазы синтезирована серия праймеров, которые предположительно должны были отжигаться на последовательность клонируемого гена. Праймеры спланированы исходя из последовательности предположительно наиболее близких по нуклеотидной последовательности генов карбоксипентидаз S. avermitilis и S. griseus, выбранных по гомологии N-концевой последовательности аминокислот зрелого фермента. Сравнение последовательности карбоксипептидаз из S. avermitilis и S. griseus (ORF – 1353 п.н.) показало уровень гомологии 77%. В консервативной области зрелого фермента уровень гомологии двух последовательностей ДНК составлял 82%, что позволяло амплифицировать консервативный район гена карбоксипептидазы КПSB даже при высоком проценте GC геномной ДНК. В то время как уровень гомологии в области, кодирующей про-часть (368 п.н.), более низок – около 50%. Вследствие этого попытка прямого клонирования полноразмерной карбоксипептидазы была неосуществима. Фрагмент гена, кодирующий зрелый фермент КПSB, получен в ходе следующих операций. При ПЦР амплификации с геномной ДНК проводят эффективный синтез специфического фрагмента ДНК, соответствующего предсказанной длине (0.780 kb). Для этого используют праймеры bik6a (gggatccgtcaggtgcagtactgcgcttccttgccga) и bik7a (GaattCACCAGCACGCCCGCGAGCACCTGAC) в условиях: 95°С – 10 сек, 64°С – 20 сек, 70С° – 2 мин, а также смесь Taq и Pfu политераз (Fermentas, Литва) с буфером для Pfu политеразы той же фирмы. Полученный фрагмент, соответствующий области внутри участка, кодирующего зрелый фермент, секвенируют. На основе полученной последовательности синтезируют серию праймеров, элонгация которых приводит к репликации ДНК фланговых последовательностей, лежащих за пределами фрагмента в соответствии со стратегией обращенного ПЦР (inverted PCR) (Nucleic Acids Res. 1988 August 25; 16(16): 8186). Далее, амплификацию фланкирующих участков гена осуществляют следующим образом. Геномную ДНК штамма S. bikiniensis ВКПМ Ас-1783 подвергают неполной рестрикции TaqI рестриктазой в соответствии с протоколом производителя (Fermentas, Литва). Реакцию проводят при 55°С и соотношении реагентов: 2 единицы активности фермента (TaqI) на 10 мкг геномной ДНК. В результате 20% всех сайтов рестриктазы подверглось расщеплению. Был получен препарат геномной ДНК, содержащий рестриктные фрагменты различного размера с преобладающей фракцией 1.0-1.5 kb. Далее геномную ДНК, подвергшуюся рестрикции, превращают в кольцевую форму за счет лигирования при низкой концентрации (0.005 мг/мл) с помощью ДНК лигазы (Fermentas) в стандартном буфере для ДНК лигазы (Fermentas) без добавления полиэтиленгликоля в условиях, предписанных производителем. Продуктами последующей ПЦР амплификации с использованием праймеров bik3b (cctgcagccccacttgtagttccagttgcggtt) и bik5b (gaattcGGCGGCTTCTACCCGCCCGACGAGGT) являются фрагменты различного размера 0.5 kb, 0.8 kb 1.0 kb, для одного из которых ПЦР реакция (0.8 kb) устойчиво воспроизводилась. Амплифицированный участок секвенируют. В результате определена последовательность нуклеотидов (SEQ ID NO 1), соответствующая зрелому ферменту КПSB. Сравнение полной нуклеотидной последовательности фрагмента ДНК SB27-995 фермента КПSB с соответствующим фрагментом гена КПSG показало 81% идентичности. Это свидетельствует о существенном отличии заявляемого фермента КПSB от его ближайшего аналога КПSG.
Формула изобретения
1. Фермент карбоксипептидаза KПSB, имеющий аминокислотную последовательность SEQ ID NO 2. 2. Штамм Streptomyces bikiniensis ВКПМ Ас-1783-продуцент карбоксипептидазы КПSВ по п.1. 3. Фрагмент ДНК SB27-995, кодирующий синтез зрелой формы карбоксипептидазы КПSB и имеющий нуклеотидную последовательность SEQ ID NO 1, либо соответствующую ей нуклеотидную последовательность в силу вырожденности генетического кода. 4. Способ микробиологического синтеза карбоксипептидазы КПSB, включающий культивирование в подходящих условиях штамма-продуцента этого фермента Streptomyces bikiniensis ВКПМ Ac-1783.
РИСУНКИ
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 360×t (мин)·v2 (мл), где
360×t (мин)·v2 (мл), где 
 0) для данной концентрации субстрата. Затем строят график зависимости
0) для данной концентрации субстрата. Затем строят график зависимости  O.D.225) от времени (t, с), выбирают прямолинейный участок полученной кривой, свидетельствующий об отсутствие субстратного истощения, и определяют значения
O.D.225) от времени (t, с), выбирают прямолинейный участок полученной кривой, свидетельствующий об отсутствие субстратного истощения, и определяют значения