Патент на изобретение №2388819
|
||||||||||||||||||||||||||
(54) СПОСОБ ПОЛУЧЕНИЯ ПЕРОКСИДАЗЫ ХРЕНА
(57) Реферат:
Изобретение относится к биотехнологии. Способ получения пероксидазы хрена включает гомогенизацию корней хрена, экстракцию фермента, концентрирование ультрафильтрацией и осаждение белков сульфатом аммония. Осадок белков диализуют против воды и 0,01-0,03 М раствора TEA-HCl буфера, с последующей очисткой пероксидазы от балластных белков на колонке с ДЭАЭ-целлюлозой в том же буфере. Затем продолжают очистку в 0,01-0,03 М растворе МОРS-NаОН буфера на колонке с КМ-целлюлозой при значениях pH и pK буферов 7,1-7,4 и 7,5-7,6 соответственно. Очищенный целевой продукт диализуют против воды и 0,01-0,03 М NaCl, а затем лиофилизируют. Способ обеспечивает упрощение получения высокоочищенной пероксидазы хрена и повышение ее выхода. 2 з.п. ф-лы.
Изобретение относится к биохимии и биотехнологии, а именно к получению высокоочищенной пероксидазы хрена адсорбционной хроматографией. Высокоочищенная пероксидаза хрена (критерий очистки А403/А275 Известен способ получения пероксидазы, включающий гомогенизацию корней хрена, экстракцию фермента водой или солевым раствором, фракционирование экстракта сульфатом аммония, очистку гель-фильтрацией, спиртовым осаждением, переосаждением хлоридом аммония, фильтрацией через сефадекс G-50, ионообменной хроматографией и диализом [1]. Недостатками этого способа являются сложность и продолжительность получения, сравнительно низкая чистота полученного препарата (RZ~2,7), трудность получения большого количества фермента. Известен также способ получения пероксидазы, включающий гомогенизацию корней хрена, экстракцию фермента, фракционирование экстракта сульфатом аммония, гель-фильтрацию [2]. Недостаток этого способа заключается в низком выходе фермента, низкой его чистоте. Задачей данного изобретения является повышение выхода высокоочищенного фермента при упрощении способа производства. Поставленная задача решается тем, что предложен способ получения пероксидазы хрена, включающий гомогенизацию корней хрена, экстракцию фермента, концентрирование ультрафильтрацией, осаждение белков сульфатом аммония, диализ осадка белков против 0,01-0,03 М раствора TEA (триэтаноламин)-HCl буфера, последовательную очистку пероксидазы от балластных белков на колонке с ДЭАЭ-целлюлозой в том же буфере, а затем в 0,01-0,03 М растворе MOPS(N-морфолинопропансульфоновая кислота)-NaOH буфера на колонке с КМ-целлюлозой, при значениях pH и pK буферов, близких к изоточке целевого продукта, и концентрации, обеспечивающей движение пероксидазы вниз по колонке, по принципу адсорбции-десорбции, со скоростью, меньшей, чем у части балластных белков, имеющих одноименный заряд с зарядом на колонке, тогда как другая их часть остается на старте вследствие ионного обмена; раствор очищенного целевого продукта диализуют против воды и 0,01-0,03 М NaCl с последующей лиофилизацией, причем фермент после гомогенизации корней экстрагируют водой, сульфат аммония вносят в концентрации 70-75% от насыщения, а значения pH и pK буферов составляют 7,1-7,4 и 7,5-7,6 соответственно. Отличием данного способа является одновременное использование одного и того же носителя для адсорбционной хроматографии целевого продукта и для очистки от балластных белков, при значениях pH, близких к изоточке целевого продукта, а также использование буферов небольшой концентрации со значениями pK, близкими к изоточке фермента, причем компоненты буферов имеют объемистые электронодонорные или электроноакцепторные группы, конкурирующие за адсорбент с группами макромолекулы пероксидазы. Пероксидазу хрена получают из сока корней хрена, собранных в самом начале цветения, в это время содержание фермента в корнях в 3-4 раза выше, по сравнению с прототипом. Техническим результатом решения задачи является получение пероксидазы с RZ 3,35 и активностью 1000 ед/мг фермента (субстрат 4-аминоантипирин) за одну стадию. Выход фермента 3,2 г из 100 кг корней, что в 6 раз выше по сравнению с прототипом. Предложенный способ позволяет уменьшить число стадий и за счет этого снизить время на получение целевого продукта и увеличить в 1,5-2 раза его выход. Известно, что адсорбция белков на каком-либо сорбенте связана с различными нековалентными взаимодействиями макромолекулярных групп с поверхностью этого сорбента. Однако их суммарная энергия мала по сравнению, например, со связыванием белок-ингибитор при аффинной хроматографии или ионными взаимодействиями на ионообменнике. Таким образом, адсорбционное взаимодействие маскируется другими, более сильными взаимодействиями поверхность белка-поверхность носителя. Поэтому, если предполагается очистить данный белок адсорбционной хроматографией, необходимо создать такие условия, чтобы балластные белки либо связывались на колонке, либо двигались по ней быстрее целевого белка, т.е. чтобы только данный белок мог медленно двигаться по принципу адсорбции-десорбции. Такие условия создаются, если использовать в качестве носителя для адсорбционной хроматографии целлюлозу, а для очистки от балластных белков использовать ДЭАЭ- и КМ-группы. Для того чтобы добиться одновременного движения целевого фермента по принципу адсорбционной хроматографии, а части балластных белков – вследствие отталкивания заряженного белка от одноименно заряженных групп, необходимо совместить эти группы и целлюлозный носитель, т.е. использовать ДЭАЭ- и КМ-целлюлозы. Если проводить хроматографию при значениях pH, близких к изоточке целевого фермента, часть балластных белков задерживается на старте вследствие ионного обмена, а часть быстро движется вниз из-за взаимодействия с одноименными зарядами ионообменника. Незаряженный или слабо заряженный фермент будет медленно двигаться, по сравнению с балластными белками, вниз и на ДЭАЭ- и на КМ-целлюлозах в случае конкуренции между группами фермента и компонентами буфера за сорбент. Буфер имеет: 1) небольшую концентрацию, поскольку при ее повышении конкуренция компонентов буфера может быть столь сильной, что целевой продукт будет двигаться со скоростью, сравнимой со скоростью движения балластных белков, и очистки не произойдет; 2) поскольку буфер должен обеспечить устойчивое значение pH при небольшой концентрации, он должен иметь pK, близкое к изоточке целевого продукта; 3) компоненты буфера должны иметь объемистые группы для эффективной конкуренции с поверхностными группами целевого продукта. Балластные белки будут заряжены при этом значении pH и, в зависимости от заряда, либо остаются на старте («ионный обмен»), либо движутся значительно быстрее целевого продукта (взаимодействие одноименных зарядов). Незаряженный или слабо заряженный целевой продукт будет отделяться и от белков, остающихся на старте, и от быстро движущихся белков. Лишь белки, имеющие изоточки, близкие к изоточке пероксидазы, могли бы связываться адсорбционно и тем самым уменьшать степень очистки, но, как показывает опыт, такие белки в соке из корней хрена отсутствуют или находятся в минорном количестве. Очистка целевого продукта: 1) белки из сока корней хрена после их гомогенизации экстрагируют водой, доводят ультрафильтрацией до начального объема сока, осаждают сульфатом аммония, диализуют против воды, а затем против 0,01-0,03 М ТЕА-HCl буфера, pH 7,1-7,4 и наносят последовательно на колонки с ДЭАЭ-целлюлозой в Cl–-форме и КМ-целлюлозой в Na+-форме, уравновешенные соответственно 0,01-0,03 М ТЕА-HCl и 0,01-0,03 М MOPS-NaOH буферами, pH 7,1-7,4 (pK буферов 7,5 и 7,6; изоточка пероксидазы 7,2); 2) за медленным движением коричневого кольца фермента следят визуально; 3) после выхода фермента из колонки с КМ-целлюлозой его диализуют последовательно против воды и 0,01-0,03 М NaCl, разбавляют до 2-5 мг/мл раствором 0,01-0,03 М NaCl и лиофилизируют; 4) измеряют значения RZ продукта и его активность по 4-аминоантипирину. Для измерения пероксидазной активности в кювету вносят 1,5 мл фенол-аминоантипиринового раствора (810 мг фенола растворяют в 40 мл воды, добавляют 25 мг 4-аминоантипирина и разбавляют водой до 50 мл), 1,4 мл раствора пероксида водорода (1 мл 30% H2O2 разбавляют водой до 100 мл; далее 1 мл этого раствора разбавляют 0,2 М калий-фосфатным буфером, pH 7,0 до 50 мл), 0,1 мл раствора фермента. Измерение оптической плотности проводят при 510 нм и температуре 25°С. Определяют скорость из линейной части кривой: ед/мг=( Концентрация фермента равна: мг фермента/мл=А403×0,44. Следующие примеры иллюстрируют эти положения. Пример 1. 100 кг корней хрена двухлетнего возраста, собранных в начале цветения, промывают водой, гомогенизируют, экстрагируют фермент водой, концентрируют ультрафильтрацией до начального объема сока, осаждают белок сульфатом аммония (70% насыщения). Осадок растворяют в минимальном количестве воды и диализуют против нее, а затем против 0,01 М ТЕА-HCl буфера, pH 7,1. Концентрацию белка доводят до 80 мг/мл концентрированием ультрафильтрацией (определение белка по Бредфорд или Лоури) и наносят на колонку (10×20 см) с ДЭАЭ-целлюлозой, уравновешенную 0,01 М ТЕА-HCl буфером, pH 7,1 и pK 7,5. После прохождения самотеком коричневого кольца его наносят на колонку того же объема с КМ-целлюлозой, уравновешенную 0,01 М MOPS-NaOH буфером, pH 7,1 и pK 7,5. После выхода фермента из колонки с КМ-целлюлозой его диализуют последовательно против воды и 0,01 М NaCl, разбавляют до 2-5 мг/мл раствором 0,01 М NaCl и лиофилизируют. Выход: 3,2 г пероксидазы с RZ 3,35 и активностью 1000 ед/мг фермента по 4-аминоантипирину. Пример 2. 100 кг корней хрена двухлетнего возраста, собранных в начале цветения, промывают водой, гомогенизируют, экстрагируют фермент водой, концентрируют ультрафильтрацией до начального объема сока, осаждают белок сульфатом аммония (75% насыщения). Осадок растворяют в минимальном количестве воды и диализуют против нее, а затем против 0,03 М ТЕА-HCl буфера, pH 7,4. Концентрацию белка доводят до 80 мг/мл концентрированием ультрафильтрацией и наносят на колонку (10×20 см) с ДЭАЭ-целлюлозой, уравновешенную 0,03 М ТЕА-HCl буфером, pH 7,4, pK 7,6. Далее хроматографируют целевой продукт, как в примере 1, на КМ-целлюлозе, но в 0,03 М MOPS-NaOH буфере, pH 7,4, pK 7,6. После выхода фермента из колонки с КМ-целлюлозой его диализуют последовательно против воды и 0,03 М NaCl, разбавляют до 2-5 мг/мл раствором 0,03 М NaCl и лиофилизируют. Выход: 3,2 г пероксидазы с RZ 3,35 и активностью 1000 ед/мг фермента по 4-аминоантипирину. Пример 3. 15 г лиофилизированного порошка пероксидазы с RZ 0,3 суспендируют в 0,01 М TEA-HCl буфере, pH 7,1 центрифугируют (20 мин, 3000 г) и 300 мл супернатанта наносят на колонку (10×20 см) с ДЭАЭ-целлюлозой, уравновешенную тем же буфером. После прохождения самотеком коричневого кольца его наносят на колонку того же объема с КМ-целлюлозой, уравновешенную 0,01 М MOPS-NaOH буфером, pH 7,1, pK 7,5. После выхода фермента из колонки с КМ-целлюлозой его диализуют последовательно против воды и 0,01 М NaCl, разбавляют до 2-5 мг/мл раствором 0,01 М NaCl и лиофилизируют. Выход составляет 1,5 г целевого продукта с RZ 3,35 и активностью 1000 ед/мг фермента по 4-аминоантипирину. Пример 4. 15 г лиофилизированного порошка пероксидазы с RZ 0,3 суспендируют в 0,03 М TEA-HCl буфере, pH 7,4, центрифугируют (20 мин, 3000 г) и 300 мл супернатанта наносят на колонку (10×20 см) с ДЭАЭ-целлюлозой, уравновешенную тем же буфером. После прохождения самотеком коричневого кольца его наносят на колонку того же объема с КМ-целлюлозой, уравновешенную 0,03 М MOPS-NaOH буфером, pH 7,4, pK 7,6. После выхода фермента из колонки с КМ-целлюлозой его диализуют последовательно против воды и 0,03 М NaCl, разбавляют до 2-5 мг/мл раствором 0,03 М NaCl и лиофилизируют. Выход составляет 1,5 г целевого продукта с RZ 3,35 и активностью 1000 ед/мг фермента по 4-аминоантипирину. Таким образом, предлагаемый способ в сравнении с прототипом позволяет получить целевой продукт с RZ 3,35 и активностью 1000 ед/мг фермента по 4-аминоантипирину. Выход фермента составляет 3,2 г на на 100 кг корней, что превышает выход по прототипу ~ в 6 раз. Кроме того, полученный целевой продукт обладает большей чистотой по сравнению с прототипом (RZ соответственно 3,35 и 2,7). Упрощение способа выражается в сокращении биохимических стадий с четырех до одной. Этот способ позволяет также выделять высокоочищенную пероксидазу из лиофильно высушенного низкоочищенного фермента. Источники литературы 1. Paul К.G. The Enzymes. New York, Acad. Press, 1963. 2. Патент РФ
Формула изобретения
1. Способ получения пероксидазы хрена, включающий гомогенизацию корней хрена, экстракцию фермента, концентрирование ультрафильтрацией, осаждение белков сульфатом аммония, диализ осадка, отличающийся тем, что осадок белков диализуют против воды и 0,01-0,03 М раствора TEA-HCl буфера с последующей очисткой пероксидазы от балластных белков на колонке с ДЭАЭ-целлюлозой в том же буфере, а затем в 0,01-0,03 М растворе MOPS-NaOH буфера на колонке с КМ-целлюлозой, при значениях pH и pK буферов, близких к изоточке целевого продукта 7,1-7,4 и 7,5-7,6 соответственно, очищенный целевой продукт диализуют против воды и 0,01-0,03 М NaCl, а затем лиофилизируют. 2. Способ по п.1, отличающийся тем, что фермент после гомогенизации корней экстрагируют водой. 3. Способ по п.1, отличающийся тем, что сульфат аммония вносят в концентрации 70-75% насыщения.
|
||||||||||||||||||||||||||
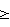 3,1 – RZ от Reinheitszahl – показатель чистоты) – один из наиболее востребованных ферментов для биохимии и биотехнологии. В частности, она широко используется в методах иммуноферментного анализа, например, для определения ВИЧ-инфекции, гепатитов и других социально значимых заболеваний человека.
3,1 – RZ от Reinheitszahl – показатель чистоты) – один из наиболее востребованных ферментов для биохимии и биотехнологии. В частности, она широко используется в методах иммуноферментного анализа, например, для определения ВИЧ-инфекции, гепатитов и других социально значимых заболеваний человека. А510/мин)/6,58×мг фермента/мл.
А510/мин)/6,58×мг фермента/мл. 2130070.
2130070.