Патент на изобретение №2166959
|
||||||||||||||||||||||||||
(54) СПОСОБ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ С ИСПОЛЬЗОВАНИЕМ ИНТЕРФЕРОНОВ ТИПА I
(57) Реферат: Изобретение относится к медицине, в частности к иммунологии и неврологии, и касается лечения аутоиммунных заболеваний. Для этого лечение включает стадию перорального введения интерферонов первого типа, причем каждое введение осуществляют в дозе от 50 до 25000 МЕ/кг. В этой связи предлагают лечить рассеянный склероз для снижения тяжести или частоты рецидивов, а также уменьшать аутоиммунное воспаление и ингибировать приступ аутоиммунного заболевания. Способ, в отличие от известного парентерального введения интерферонов, представляется удобным в использовании, эффективным и может применяться длительные сроки, насколько необходимо для лечения. 4 с. и 10 з.п. ф-лы, 10 ил. Настоящее изобретение относится к области неврологии, иммунологии и химии протеинов. Более конкретно, настоящее изобретение относится к новым способам лечения аутоиммунных заболеваний с использованием интерферонов первого типа. Острый экспериментальный аутоиммунный энцефаломиелит /EAE/ представляет собой воспалительный аутоиммунный процесс в центральной нервной системе, опосредованный T клетками, который напоминает такое заболевание человека, как рассеянный склероз. CR-EAE, хронический воспалительный аутоиммунный процесс центральной нервной системы, который напоминает рассеянный склероз человека /MS/ как в патологических, так и в клинических проявлениях, предоставляет модель для оценки вмешательств с целью изменения течения аутоиммунного заболевания человека. Миелиновый основной протеин /MBP/ представляет собой важный нейроантиген в патогенезе этого заболевания. CR-EAE у SJL/J мышей можно адаптивно переносить после внутривенной инъекции MBP пептида 89-100 специфической T-клеточной линии. Совместное культивирование MBP пептида 91-103 специфических T клеток с химически связанным пептидом придает T клеткам устойчивость к MBP при CR-EAE у SJL/J мышей. Парентеральное, например, четырехкратное введение природного крысиного интерферона /105 ед./ частично подавляет острый EAE у самцов крыс Льюиса и ингибирует пассивный гиперострый локализованный EAE при введении в тот же день, что и инокуляция иммуногена. Другие парэнтерально вводимые питокины, такие как TGF-  , могут ослабить клиническую картину заболевания и воспаления в мозгу и спинном мозгу при EAE. Парентерально вводимый природный IFN- , могут ослабить клиническую картину заболевания и воспаления в мозгу и спинном мозгу при EAE. Парентерально вводимый природный IFN- человека может понизить функции T клеток и выработку антител, зависимую от T клеток у людей. При пероральном введении природный IFN- человека может предотвращать экспериментальное развитие вирусных и паразитических инфекций у животных. Острый EAE предоставляет модель для демонстрации способности перорально вводимых иммуноактивных веществ влиять на течение аутоиммунного заболевания IFN- человека, представляет собой иммуноактивный протеин, который можно перорально вводить в низких дозах при лечении вирусных заболеваний у животных. IFN- человека другой интерферон первого типа усиливает функции супрессорных клеток ин витро при прогрессивном MS, а крысиный IFN-/ стимулам; активные ревматоидные артриты демонстрируют значительно пониженное продуцирование IFN-/ по сравнению с нормальными донорами. Такие другие аутоиммунные заболевания, как псориаз и атопические дерматиты, также демонстрируют уменьшение продуцирования интерферона. Лимфоциты периферической крови у пациентов с МS демонстрируют аналогичное неэффективное продуцирование интерферона типа 1 в ответ на вирусные или митогенные стимулы, параллельное с тяжестью заболевания. У больных рассеянным склерозом наблюдается также дефицит активности природных клеток-киллеров [NK], что коррелируется с тяжестью заболевания и может быть нормализовано за счет лечения IFN-.
Известные ненормальности в продуцировании или реакциях интерферона типа 1 при аутоиммунных заболеваниях побудили провести несколько небольших пилотных исследований с использованием интерферонов типа 1 в качестве терапевтических агентов для MS. В этих исследованиях использовали систематическое введение интерферона типа 1 в дозах 1 млн. ед. или более ежедневно без заметного клинического улучшения. Исследования парентерального введения IFN- при рассеянном склерозе с чередованием рецидивов и ремиссий позволяет предположить, что введение 1,6-8 млн. ед. IFN- подкожно трижды в неделю может уменьшить рецидивы на 40-50% и уменьшить воспаление мозга, о чем свидетельствуют серии MBI /изображения, полученные с помощью магнитного резонанса.
Известные способы лечения испытывают недостаток эффективных средств для лечения аутоиммунных заболеваний за счет перорального введения таких цитокинов, как интерфероны первого типа. Настоящее изобретение восполняет эту настоятельную потребность.
Целью настоящего изобретения является демонстрация того, что пероральное введение интерферона типа 1 ингибирует пролиферацию сенсибилизированных антигенов, подавляет клинические проявления рецидивов и уменьшает воспаление за счет изменения цитокинов в мышином CR-EAE.
Другой целью настоящего изобретения является иллюстрация действий интерферонов первого типа при пероральном введении до и после иммунизации GP-MBP при остром EAE.
Другой целью изобретения является подавление клинических проявлений приступов аутоиммунного заболевания, уменьшение патологических осложнений и ингибирование воспалительных питокинов при таких заболеваниях за счет перорального введения модификаторов биологических реакций, таких как интерфероны первого типа.
Целью настоящего изобретения является также демонстрация действия перорального введения крысиного IFN-/ на клинические проявления, гистологию и IFN- человека может понизить функции T клеток и выработку антител, зависимую от T клеток у людей. При пероральном введении природный IFN- человека может предотвращать экспериментальное развитие вирусных и паразитических инфекций у животных. Острый EAE предоставляет модель для демонстрации способности перорально вводимых иммуноактивных веществ влиять на течение аутоиммунного заболевания IFN- человека, представляет собой иммуноактивный протеин, который можно перорально вводить в низких дозах при лечении вирусных заболеваний у животных. IFN- человека другой интерферон первого типа усиливает функции супрессорных клеток ин витро при прогрессивном MS, а крысиный IFN-/ стимулам; активные ревматоидные артриты демонстрируют значительно пониженное продуцирование IFN-/ по сравнению с нормальными донорами. Такие другие аутоиммунные заболевания, как псориаз и атопические дерматиты, также демонстрируют уменьшение продуцирования интерферона. Лимфоциты периферической крови у пациентов с МS демонстрируют аналогичное неэффективное продуцирование интерферона типа 1 в ответ на вирусные или митогенные стимулы, параллельное с тяжестью заболевания. У больных рассеянным склерозом наблюдается также дефицит активности природных клеток-киллеров [NK], что коррелируется с тяжестью заболевания и может быть нормализовано за счет лечения IFN-.
Известные ненормальности в продуцировании или реакциях интерферона типа 1 при аутоиммунных заболеваниях побудили провести несколько небольших пилотных исследований с использованием интерферонов типа 1 в качестве терапевтических агентов для MS. В этих исследованиях использовали систематическое введение интерферона типа 1 в дозах 1 млн. ед. или более ежедневно без заметного клинического улучшения. Исследования парентерального введения IFN- при рассеянном склерозе с чередованием рецидивов и ремиссий позволяет предположить, что введение 1,6-8 млн. ед. IFN- подкожно трижды в неделю может уменьшить рецидивы на 40-50% и уменьшить воспаление мозга, о чем свидетельствуют серии MBI /изображения, полученные с помощью магнитного резонанса.
Известные способы лечения испытывают недостаток эффективных средств для лечения аутоиммунных заболеваний за счет перорального введения таких цитокинов, как интерфероны первого типа. Настоящее изобретение восполняет эту настоятельную потребность.
Целью настоящего изобретения является демонстрация того, что пероральное введение интерферона типа 1 ингибирует пролиферацию сенсибилизированных антигенов, подавляет клинические проявления рецидивов и уменьшает воспаление за счет изменения цитокинов в мышином CR-EAE.
Другой целью настоящего изобретения является иллюстрация действий интерферонов первого типа при пероральном введении до и после иммунизации GP-MBP при остром EAE.
Другой целью изобретения является подавление клинических проявлений приступов аутоиммунного заболевания, уменьшение патологических осложнений и ингибирование воспалительных питокинов при таких заболеваниях за счет перорального введения модификаторов биологических реакций, таких как интерфероны первого типа.
Целью настоящего изобретения является также демонстрация действия перорального введения крысиного IFN-/ на клинические проявления, гистологию и IFN- секрецию при экспериментальных аллергических невритах на ин виво модели острой воспалительной демиелинирующей радикулоневропатии или синдроме Гийена-Барре /Guillain – Barre’/.
В одном из вариантов настоящего изобретения предложен способ лечения аутоиммунных заболеваний у животных, который включает стадию перорального введения этим животным интерферона типа 1.
В другом варианте изобретения предложен способ снижения тяжести или частоты рецидивов рассеянного склероза у людей, который включает стадию перорального введения интерферона типа 1 указанному пациенту.
В еще одном варианте настоящего изобретения предложен способ уменьшения воспалений, связанных с аутоиммунным заболеванием у животных, который включает стадию перорального введения интерферона первого типа указанному животному.
И, наконец, в еще одном варианте настоящего изобретения предложен способ ингибирования приступов аутоиммунного заболевания, включающий стадию перорального введения указанному животному интерферона первого типа.
Другие аспекты, особенности и преимущества изобретения будут очевидны из следующего описания представленных предпочтительных вариантов изобретения, приводимых с целью раскрытия изобретения.
С целью разъяснения вышеперечисленных особенностей, преимуществ и целей настоящего изобретения, а также других, которые выяснятся в дальнейшем, а также с целью более детального разъяснения, более конкретные описания изобретения, кратко суммированного ранее, можно осуществить со ссылками на некоторые варианты изобретения, которые иллюстрируются в прилагаемых чертежах. Эти чертежи составляют часть описания. Однако следует отметить, что прилагаемые чертежи иллюстрируют лишь предпочтительные варианты изобретения и поэтому их не следует рассматривать как ограничивающие объем изобретения.
Фиг. 1 демонстрирует, что пероральное введение мышиного природного IFN-/ подавляет клинический рецидив при мышином CR-EAE. Двум группам по шесть животных инокулируют, а после первого приступа дают либо имитацию IFN, либо по 100 ед. мышиного природного IFN-/ трижды в неделю в течение 7 недель. Представлен один из двух представительных примеров. SEM/ср. квадратичная ошибка/ составляет менее 10%.
Фиг. 2 демонстрирует, что пероральное введение 5000 ед. крысиного IFN-/ ингибирует клиническое проявление при остром EAE. Четырем группам крыс Льюиса /по 6 в каждой/ вводят имитацию IFN 1000, 3000 или 5000 ед. IFN-/ каждой крысе в течение 7 дней, предшествующих иммунизации /-7/ и в течение последующих 21 дня /+14/. Результаты выражены как средний клинический показатель для каждой группы в каждый день заболевания после инокуляции секрецию при экспериментальных аллергических невритах на ин виво модели острой воспалительной демиелинирующей радикулоневропатии или синдроме Гийена-Барре /Guillain – Barre’/.
В одном из вариантов настоящего изобретения предложен способ лечения аутоиммунных заболеваний у животных, который включает стадию перорального введения этим животным интерферона типа 1.
В другом варианте изобретения предложен способ снижения тяжести или частоты рецидивов рассеянного склероза у людей, который включает стадию перорального введения интерферона типа 1 указанному пациенту.
В еще одном варианте настоящего изобретения предложен способ уменьшения воспалений, связанных с аутоиммунным заболеванием у животных, который включает стадию перорального введения интерферона первого типа указанному животному.
И, наконец, в еще одном варианте настоящего изобретения предложен способ ингибирования приступов аутоиммунного заболевания, включающий стадию перорального введения указанному животному интерферона первого типа.
Другие аспекты, особенности и преимущества изобретения будут очевидны из следующего описания представленных предпочтительных вариантов изобретения, приводимых с целью раскрытия изобретения.
С целью разъяснения вышеперечисленных особенностей, преимуществ и целей настоящего изобретения, а также других, которые выяснятся в дальнейшем, а также с целью более детального разъяснения, более конкретные описания изобретения, кратко суммированного ранее, можно осуществить со ссылками на некоторые варианты изобретения, которые иллюстрируются в прилагаемых чертежах. Эти чертежи составляют часть описания. Однако следует отметить, что прилагаемые чертежи иллюстрируют лишь предпочтительные варианты изобретения и поэтому их не следует рассматривать как ограничивающие объем изобретения.
Фиг. 1 демонстрирует, что пероральное введение мышиного природного IFN-/ подавляет клинический рецидив при мышином CR-EAE. Двум группам по шесть животных инокулируют, а после первого приступа дают либо имитацию IFN, либо по 100 ед. мышиного природного IFN-/ трижды в неделю в течение 7 недель. Представлен один из двух представительных примеров. SEM/ср. квадратичная ошибка/ составляет менее 10%.
Фиг. 2 демонстрирует, что пероральное введение 5000 ед. крысиного IFN-/ ингибирует клиническое проявление при остром EAE. Четырем группам крыс Льюиса /по 6 в каждой/ вводят имитацию IFN 1000, 3000 или 5000 ед. IFN-/ каждой крысе в течение 7 дней, предшествующих иммунизации /-7/ и в течение последующих 21 дня /+14/. Результаты выражены как средний клинический показатель для каждой группы в каждый день заболевания после инокуляции  SEM. Приводится один из представительных примеров. Данные для случаев введения 1000 и 3000 ед. не показаны, xp менее 0,02 между имитатором интерферона и 5000 ед. IFN-/ введенных животным.
Фиг. 3 демонстрирует, что пероральное введение hrIFN- крысам с острым EAE задерживает приступ, снижает тяжесть его, и скорость возврата клинического приступа. Трем группам по семь крыс Льюиса инокулируют MBP и CFA в 0 день и перорально вводят PBS 1000 ед. или 5000 ед. hrIFN- ежедневно, начиная с семи дней до иммунизации /-7/ и в течение 14 дней после иммунизации /+14/. Величины представляют экспериментальные клинические показатели при аллергическом неврите для каждой из групп по 7 животных SEM. Представлен один из двух представительных примеров.
Фиг. 4 демонстрирует, что пероральное введение hrIFN- крысам с острым EAE снижает кумулятивную клиническую тяжесть острых клинических приступов при аллергических невритах. Три группы по семь крыс Льюиса обрабатывают, как представлено на фиг. 3. Приведенные величины соответствуют кумулятивным клиническим показателям при аллергических невритах для каждой группы из 7 животных с 10 до 16 дня после инокуляции. xp менее 0.001, 5000 ед. IFN- против как для 1000 ед. IFN-, так и для PBS контроля.
Фиг. 5 демонстрирует, что пероральное введение 5000 ед. hrIFN- снижает количество воспалительных центров в спинном мозге по сравнению с контрольной группой и группой, которой вводят 1000 ед. интерферона. После умерщвления спинной мозг животных, представленных на фиг. 3, обрабатывают в соответствии с описанием в разделе методики. Результаты выражены как количество воспалительных центров для спинного мозга SEM. Представлен один из представительных примеров. xp менее 0,04, 5000 ед. IFN против 1000 ед. IFN или PBS контрольной группы.
Фиг. 6 демонстрирует, что пероральное введение 5000 ед. hrIFN- без подкожного введения снижает тяжесть клинических приступов. Трем группам крыс Льюиса по 6 в группе инокулируют MBP и CFA на 0 день, и без обработки вводят 5000 или инъектируют подкожно 5000 ед. hrIFN- ежедневно, начиная с семи дней до иммунизации /-7/ и в течение последующих 21 дня. /+14/. Величины представляют ежедневные клинические показатели экспериментальных аллергических невритов для каждой группы из 6 животных SEM. Представлен один из двух представительных примеров.
Фиг. 7 демонстрирует, что пероральное введение hrIFN- крысам с острым EAE снижает кумулятивную клиническую тяжесть острых клинических приступов при экспериментальных аллергических невритах. Три группы по шесть крыс Льюиса в каждой обрабатывают, как показано на фиг. 6. Величины представляют кумулятивные клинические показатели при экспериментальных аллергических невритах для каждой группы из 6 животных с 10 до 16 дня после инокуляции. xp менее 0,005, для животных, которым вводили, по сравнению с необработанными/ подкожно введенными обработанными животными.
Фиг. 8 демонстрирует, что пероральное введение 5000 ед. hrIFN- снижает число воспалительных центров в спинном мозге по сравнению с контрольной группой. После умерщвления спинной мозг животных, представленных на фиг. 6, обрабатывают, как описано ранее, и проводят независимую оценку для центров воспаления одним наблюдателем, которому не известен способ обработки. Результаты выражены как число воспалительных центров на спинной мозг SEM. Представлен один из двух представительных примеров. xp менее 0,04, 5000 ед. IFN введено перорально против 5000 ед., введенных подкожно.
Фиг. 9A и 9B демонстрируют, что пероральное введение крысиного интерферона –/ снижает клинические проявления заболевания у крыс с экспериментальными аллергическими невритами. Крысам Льюиса вводят либо 5000 ед. IFN-/ /интерферон/, либо моск-IFN /имитация/, начиная с 7 дней до иммунизации бычьим миелином периферических нервов. Клинический показатель оказывается значительно выше у моск(имитатор)- IFN крыс/ н = 11/, нежели у IFN-/ крыс /н=11/. В моск-IFN группе оказывается больше крыс, которые достигают клинического показателя 4 или выше, нежели в группе, которой вводили IFN-/ /моск-IFN 7 из 11 против IFN-/ 4 из 11 крыс/. /Фиг. 9A/ и соответственно больше крыс, которые теряют 25% или более веса тела /моск-IFN 4 из 11 против IFN-/ 2 из 11 крыс/ /фиг. 9B/.
Фиг. 10 демонстрирует, что пероральное введение крысиного интерферона –/ уменьшает воспаление и IFN- продуцирование при экспериментальном аллергическом неврите, Крысам Льюиса вводят перорально либо 5000 ед. IFN-/ либо моск-IFN, начиная с 7 дней до иммунизации бычьим миелином периферических нервов до момента умерщвления. Иммуноцитохимическое исследование люмбосакральных нервных корешков с использованием ED1 антител /1:1000/ и IFN- антител /1:10/ на 13 день после иммунизации во время приступа клинического заболевания показывает меньше ED1 позитивных макрофагов /фиг. 10A, 2B/ у IFN-/ обработанных крыс по сравнению со значениями у моск-IFN обработанных крыс /фиг. 10B, 10E/. Наряду со сниженным количеством ED1 позитивных макрофагов наблюдается уменьшение IFN- экспрессии у крыс, обработанных IFN-/ фиг. 100/ по сравнению с крысами, обработанными моск-IFN /фиг. 10F/. Bar = 20 микрон.
В способе настоящего изобретения модификация заболевания и ингибирование продуцирования воспалительных цитокинов, индуцированные за счет перорального введения цитокинов при EAE, предоставляет удобные, эффективные и длительного срока способы лечения аутоиммунных заболеваний, вызванных неизвестным антигеном у людей, в частности рассеянного склероза.
Настоящее изобретение направлено на способ лечения аутоиммунных заболеваний у животных, который включает стадию перорального введения интерферона типа 1 указанным животным. Обычно интерферон, который можно использовать в способе настоящего изобретения, является либо альфа-интерфероном, либо бета-интерфероном. Интерферон первого типа может быть получен из любого источника. Предпочтительно выбирать интерферон из группы, состоящей из человеческого рекомбинантного интерферона, крысиного интерферона и мышиного интерферона.
Обычно интерферон первого типа, вводимый в соответствии со способами настоящего изобретения, следует вводить в дозах, которые эффективно ингибируют приступ или рецидивы и т.д. аутоиммунных заболеваний. Так например, интерферон можно вводить в дозах от около 50 ME/кг до около 25000 ME/кг.
Способ настоящего изобретения можно использовать для любых животных с аутоиммунными заболеваниями. Более предпочтительно использовать способ настоящего изобретения для лечения людей.
Способами настоящего изобретения можно лечить широкий круг аутоиммунных заболеваний. Представительные примеры аутоиммунных заболеваний включают рассеянный склероз, ревматоидные артриты, сахарный диабет, псориаз, органоспецифические аутоиммунные заболевания, хроническую воспалительную демиелинизирующую полирадикулоневропатию и синдром Гийена-Барре.
Настоящее изобретение направлено также на способ уменьшения тяжести или частоты наступления рецидивов рассеянного склероза у людей, который включает стадию перорального введения указанному пациенту интерферона типа 1. Более того, настоящее изобретение направлено также на способ ингибирования приступов аутоиммунного заболевания, который включает стадию перорального введения указанному пациенту интерферона первого типа.
Нижеследующие примеры приведены с целью иллюстрации различных вариантов настоящего изобретения, и настоящее изобретение никоим образом не следует ограничивать экспериментальным аллергическим невритом.
Пример 1 SEM. Приводится один из представительных примеров. Данные для случаев введения 1000 и 3000 ед. не показаны, xp менее 0,02 между имитатором интерферона и 5000 ед. IFN-/ введенных животным.
Фиг. 3 демонстрирует, что пероральное введение hrIFN- крысам с острым EAE задерживает приступ, снижает тяжесть его, и скорость возврата клинического приступа. Трем группам по семь крыс Льюиса инокулируют MBP и CFA в 0 день и перорально вводят PBS 1000 ед. или 5000 ед. hrIFN- ежедневно, начиная с семи дней до иммунизации /-7/ и в течение 14 дней после иммунизации /+14/. Величины представляют экспериментальные клинические показатели при аллергическом неврите для каждой из групп по 7 животных SEM. Представлен один из двух представительных примеров.
Фиг. 4 демонстрирует, что пероральное введение hrIFN- крысам с острым EAE снижает кумулятивную клиническую тяжесть острых клинических приступов при аллергических невритах. Три группы по семь крыс Льюиса обрабатывают, как представлено на фиг. 3. Приведенные величины соответствуют кумулятивным клиническим показателям при аллергических невритах для каждой группы из 7 животных с 10 до 16 дня после инокуляции. xp менее 0.001, 5000 ед. IFN- против как для 1000 ед. IFN-, так и для PBS контроля.
Фиг. 5 демонстрирует, что пероральное введение 5000 ед. hrIFN- снижает количество воспалительных центров в спинном мозге по сравнению с контрольной группой и группой, которой вводят 1000 ед. интерферона. После умерщвления спинной мозг животных, представленных на фиг. 3, обрабатывают в соответствии с описанием в разделе методики. Результаты выражены как количество воспалительных центров для спинного мозга SEM. Представлен один из представительных примеров. xp менее 0,04, 5000 ед. IFN против 1000 ед. IFN или PBS контрольной группы.
Фиг. 6 демонстрирует, что пероральное введение 5000 ед. hrIFN- без подкожного введения снижает тяжесть клинических приступов. Трем группам крыс Льюиса по 6 в группе инокулируют MBP и CFA на 0 день, и без обработки вводят 5000 или инъектируют подкожно 5000 ед. hrIFN- ежедневно, начиная с семи дней до иммунизации /-7/ и в течение последующих 21 дня. /+14/. Величины представляют ежедневные клинические показатели экспериментальных аллергических невритов для каждой группы из 6 животных SEM. Представлен один из двух представительных примеров.
Фиг. 7 демонстрирует, что пероральное введение hrIFN- крысам с острым EAE снижает кумулятивную клиническую тяжесть острых клинических приступов при экспериментальных аллергических невритах. Три группы по шесть крыс Льюиса в каждой обрабатывают, как показано на фиг. 6. Величины представляют кумулятивные клинические показатели при экспериментальных аллергических невритах для каждой группы из 6 животных с 10 до 16 дня после инокуляции. xp менее 0,005, для животных, которым вводили, по сравнению с необработанными/ подкожно введенными обработанными животными.
Фиг. 8 демонстрирует, что пероральное введение 5000 ед. hrIFN- снижает число воспалительных центров в спинном мозге по сравнению с контрольной группой. После умерщвления спинной мозг животных, представленных на фиг. 6, обрабатывают, как описано ранее, и проводят независимую оценку для центров воспаления одним наблюдателем, которому не известен способ обработки. Результаты выражены как число воспалительных центров на спинной мозг SEM. Представлен один из двух представительных примеров. xp менее 0,04, 5000 ед. IFN введено перорально против 5000 ед., введенных подкожно.
Фиг. 9A и 9B демонстрируют, что пероральное введение крысиного интерферона –/ снижает клинические проявления заболевания у крыс с экспериментальными аллергическими невритами. Крысам Льюиса вводят либо 5000 ед. IFN-/ /интерферон/, либо моск-IFN /имитация/, начиная с 7 дней до иммунизации бычьим миелином периферических нервов. Клинический показатель оказывается значительно выше у моск(имитатор)- IFN крыс/ н = 11/, нежели у IFN-/ крыс /н=11/. В моск-IFN группе оказывается больше крыс, которые достигают клинического показателя 4 или выше, нежели в группе, которой вводили IFN-/ /моск-IFN 7 из 11 против IFN-/ 4 из 11 крыс/. /Фиг. 9A/ и соответственно больше крыс, которые теряют 25% или более веса тела /моск-IFN 4 из 11 против IFN-/ 2 из 11 крыс/ /фиг. 9B/.
Фиг. 10 демонстрирует, что пероральное введение крысиного интерферона –/ уменьшает воспаление и IFN- продуцирование при экспериментальном аллергическом неврите, Крысам Льюиса вводят перорально либо 5000 ед. IFN-/ либо моск-IFN, начиная с 7 дней до иммунизации бычьим миелином периферических нервов до момента умерщвления. Иммуноцитохимическое исследование люмбосакральных нервных корешков с использованием ED1 антител /1:1000/ и IFN- антител /1:10/ на 13 день после иммунизации во время приступа клинического заболевания показывает меньше ED1 позитивных макрофагов /фиг. 10A, 2B/ у IFN-/ обработанных крыс по сравнению со значениями у моск-IFN обработанных крыс /фиг. 10B, 10E/. Наряду со сниженным количеством ED1 позитивных макрофагов наблюдается уменьшение IFN- экспрессии у крыс, обработанных IFN-/ фиг. 100/ по сравнению с крысами, обработанными моск-IFN /фиг. 10F/. Bar = 20 микрон.
В способе настоящего изобретения модификация заболевания и ингибирование продуцирования воспалительных цитокинов, индуцированные за счет перорального введения цитокинов при EAE, предоставляет удобные, эффективные и длительного срока способы лечения аутоиммунных заболеваний, вызванных неизвестным антигеном у людей, в частности рассеянного склероза.
Настоящее изобретение направлено на способ лечения аутоиммунных заболеваний у животных, который включает стадию перорального введения интерферона типа 1 указанным животным. Обычно интерферон, который можно использовать в способе настоящего изобретения, является либо альфа-интерфероном, либо бета-интерфероном. Интерферон первого типа может быть получен из любого источника. Предпочтительно выбирать интерферон из группы, состоящей из человеческого рекомбинантного интерферона, крысиного интерферона и мышиного интерферона.
Обычно интерферон первого типа, вводимый в соответствии со способами настоящего изобретения, следует вводить в дозах, которые эффективно ингибируют приступ или рецидивы и т.д. аутоиммунных заболеваний. Так например, интерферон можно вводить в дозах от около 50 ME/кг до около 25000 ME/кг.
Способ настоящего изобретения можно использовать для любых животных с аутоиммунными заболеваниями. Более предпочтительно использовать способ настоящего изобретения для лечения людей.
Способами настоящего изобретения можно лечить широкий круг аутоиммунных заболеваний. Представительные примеры аутоиммунных заболеваний включают рассеянный склероз, ревматоидные артриты, сахарный диабет, псориаз, органоспецифические аутоиммунные заболевания, хроническую воспалительную демиелинизирующую полирадикулоневропатию и синдром Гийена-Барре.
Настоящее изобретение направлено также на способ уменьшения тяжести или частоты наступления рецидивов рассеянного склероза у людей, который включает стадию перорального введения указанному пациенту интерферона типа 1. Более того, настоящее изобретение направлено также на способ ингибирования приступов аутоиммунного заболевания, который включает стадию перорального введения указанному пациенту интерферона первого типа.
Нижеследующие примеры приведены с целью иллюстрации различных вариантов настоящего изобретения, и настоящее изобретение никоим образом не следует ограничивать экспериментальным аллергическим невритом.
Пример 1Индуцирование экспериментального аутоиммунного энцефаломиелита Хронически рецидивирующую форму EAE индуцируют у 7-10 недельных самок мышей SJL/J по способу Brown and McFarlin, Lab.Invest. /1981/ 45:278-284, модифицированному Miller et al., J.Immunol. /1987/ 138:3776-3784. Короче, каждой мыши подкожно вводят в выбритый правый бок 0,3 мл эмульсии, содержащей 1 мг сингенного мышиного гомогената спинного мозга /MSCH/ в 0,15 мл фосфатного буферированного физиологического раствора и 0,03 мг Mycobacterium tuberculoses homines H37Ra /MT/ [Difeo Labs. Detroit M1/ в 0,15 мл неполного адьюванта Фреунда /IFA/. Через семь дней мышам вводят аналогичную инъекцию в левый бок. Начальные клинические признаки заболевания наблюдаются между 13 и 25 днем после иммунизации. Клиническую тяжесть повторяющихся приступов оценивает наблюдатель /вслепую/: 0 = нет признаков заболевания, 1 = минимальная или умеренная слабость задних конечностей; 2 = умеренная слабость задних конечностей или умеренная атаксия; 3 = умеренная до сильной слабость задних конечностей; 4 = сильная слабость задних конечностей до умеренной атаксии; 5 = параплегия при более чем умеренной слабости четырех конечностей; 6 = параплегия при сильной слабости четырех конечностей или сильная атаксия. Животных оценивают вслепую в течение 6-8 недель и кумулятивный еженедельный показатель рассчитывают, усредняя три показателя в неделю /понедельник, среда, пятница/ для каждой группы животных. Пример 2 Пероральное введение цитокина После начала приступа животным перорально вводят варьирующиеся дозы /1-100 ед./ мышиного природного IFN- / /Цитоимунный мышиный IFN-/ 4,0 105 IRU /мл, Lee Biomolecular Research, Inc., San Diego, CA/, или имитатор мышиного IFN-/ /Цитоиммунный менее 2 IRU/мл, Lee Biomolecular Research, Inc., Diego, CA/ полученный идентично IFN-/
Пример 3 105 IRU /мл, Lee Biomolecular Research, Inc., San Diego, CA/, или имитатор мышиного IFN-/ /Цитоиммунный менее 2 IRU/мл, Lee Biomolecular Research, Inc., Diego, CA/ полученный идентично IFN-/
Пример 3Получение основного миелинового протеина морских свинок /GР-МВР/ 20 г спинного мозга морских свинок добавляют к 4,7 мл метанола при -20oC и гомогенизируют в тканевом гомогенизаторе. Добавляют 9,4 мл хлороформа при -20oC, гомогенизируют в течение еще 2 минут и перемешивают в течение 60 минут при комнатной температуре; затем еще 60 минут при 4oC. Полученный материал фильтруют через сложенную вдвое стерильную марлю, через охлажденную воронку Бюхнера и фильтровальную бумагу N 1 Ватман. Тканевую лепешку промывают 20 мл ацетона при -20oC, и снова суспендируют в 40 мл холодной ddH2O; затем перемешивают в течение 30 минут при 4oC, фильтруют второй раз и снова суспендируют в 10 мл холодной ddH2O. Суспензию обрабатывают ультразвуком для облегчения образования суспензии осадка в ddH2O; pH суспензии ткань/H2O доводят до pH 3 за счет 1н. HCl, и перемешивают в течение ночи при 4oC. После перемешивания в течение ночи pH доводят до 3 и вращают при 10000 g в течение 15 минут при 4oC. Надосадочную жидкость фильтруют через бумагу N 1 Ватман и жидкий фильтрат сохраняют. pH полученного фильтрата доводят до 9 за счет 10 н. NaOH и перемешивают в течение 60 минут при 4oC; затем вращают при 10000 g в течение 15 минут при 4oC. Определяют объем надосадочной жидкости и добавляют сульфат аммония до конечной концентрации 27,7 г/100 мл; надосадочную жидкость перемешивают на холоде в течение 30 минут, а затем вращают при 10000 g в течение 15 минут при 4oC и снова суспендируют в 1 мл ddH2O. Повторно суспендированный осадок диализуют в низкомолекулярной /6-8000 MB/ трубке для диализа против ddH2O. Концентрацию протеина определяют на A280 с помощью УФ спектроскопии. Для проверки чистоты препарата миелинового основного протеина приготавливают 7-20% SOS-PACE градиентный гель и окрашивают 0,3 М CuCl2. Чистота препарата миелинового основного протеина морской свинки /GP-MBP/ продемонстрирована полосой 18 кД. Пример 4 Препарат клетки лимфатического узла и селезенки Животных умерщвляют через 6-8 недель после начала перорального введения, и удаляют дренирующие паховые узлы и клетки селезенки, и суспензии отдельных клеток получают, протирая через сито из нержавеющей стали с диаметром ячеек 90 меш. Осуществляют лизис эритроцитов в клеточной суспензии селезенки с 2 мл АСК раствора, добавленного к осадку и реакции дают возможность протекать в течение 5 минут при комнатной температуре. Пример 5 Пролиферация T клеток ин витро Спустя семь дней после клинического рецидива, мышей умерщвляют, дренирующие паховые лимфатические узлы извлекают и культивируют ин витро для определения антиген-специфических T клеточных пролиферативных реакций. Стимуляцию антигеном осуществляют, используя в качестве антигена либо 0 либо 100 мкг/мл /GP-MBP или MT/ и митогенную стимуляцию Con A при 2,5 мкг/мл за счет инкубирования клеток лимфатических узлов при 2 10 клеток/ячейку в RPM1 /Gibco, Grand Island, NY/, дополненном 10% сывороткой плода теленка /FCS/ Whittaker Bioproducts Walkersville MD/, 1% пируватом натрия /Gibco, Crand, Island, NY /, 1% глутамина “Gibcol/, 1% пенициллин/стрептомицина и 50 мкл 2-меркаптоэтанола). Пластины инкубируют при 5% содержании CO2 и увлажнении при 37oC в течение 4 дней. В этот момент клетки обрабатывают 2 мкКи титрированного [3H] dTd и собирают спустя 18 часов с помощью автоматизированного сборника. Захват [3Н] dTd определяют с помощью сцинтилляционного счетчика /жидкостного/ Бекмана. Культивирование ведут в трех параллельных опытах, и полученные результаты выражают как  CPM.
Пример 6 CPM.
Пример 6Гистология После умерщвления извлекают спинной мозг и иммерсионно фиксируют в 10% нейтральном буферированном формалине минимум в течение двух недель. После фиксации мозг рассекают в горизонтальной плоскости с интервалами примерно 3 мм и обрабатывают парафином. Парафиновые блоки рассекают на 6-8 микрон и поэтапные секции окрашивают гематоксилином и зозином и Luxol-fast blue /PAS/ гематоксилином, и исследуют в оптическом микроскопе. Секции мозга оценивают независимо для определения центров воспаления /слепым методом/, без информации о характере обработки животного до умерщвления. Образцы спинного мозга для каждого животного подготавливают идентичным образом, и определяют количество центров воспаления на секцию / более 20 воспалительных клеток/ в паренхиме. Пример 7 Анализ цитокинов Клетки селезенки животных, обработанных имитацией или 100 ед. мышиного IFN- / культивируют с Con A /2,5 мкг/мл/ при 1 106 клеток/мл в 75 см2 склянках для культуры тканей в течение 48 часов во влажном инкубаторе с 5% CO2/95% воздуха при 37oC. Надосадочную жидкость собирают через 24 и 48 часов после Con A активации, и после центрифугирования замораживают при -70oC. Содержание интерлейкинов определяют, используя твердофазный ELISA анализ. Анти-IL-2, анти-IL-10 или антиIFN-/Phar Mingen San Diego, CA/ инкубируют на поливиниловых микротитровальных пластинах с 96 ячейками в 0,01М карбонатном буфере /pH 9,6/ в течение ночи при 4oC. Пластины блокируют 3% BSA в фосфатном буферированном физиологическом растворе в течение 3 часов, 100 мкл надосадочных жидкостей добавляют в различных разбавлениях, которые оттитрованы для линейного участка кривой поглощение/концентрация с трехкратным повторением и инкубируют в течение 1 часа при комнатной температуре. После того, как пластины пять раз промывают фосфатным буферированным физиологическим раствором Твина /0,05%/, в течение 60 минут добавляют 100 мкл коньюгированных с переоксидазой интерлейкиновых моноклональных антител /с эпитопными детерминантами, отличными от первого антитела, которое было использовано для покрытия поливиниловой пластины/, в концентрации 1:1000. Затем добавляют О-фенилендиаминдигидрохлорид, и поглощение определяют при 450 нм. Получают стандартные кривые для различных количеств различных интерлейкино. Статистический анализ осуществляют, используя т-тест Стьюдента.
Пример 8Пероральное введение мышиных интерферонов перового типа подавляет клинические признаки заболевания, воспаления и ингибирует пролиферацию, связанную с MBP Пероральное введение 1-10 единиц IFN первого типа оказывает иммунологический эффект, но оказывается неадекватным при давлении клинических рецидивов. Поэтому две группы по шесть иммунизованных SJL/J мышей обрабатывали имитатором IFN или 100 ед. мышиного природного IFN- / трижды в неделю, начиная с 30 дня после выздоровления после первого приступа. Групповые клинические показатели после первого приступа заметно не отличались для различных групп. Клинические рецидивы наблюдались примерно через 40 дней после инокуляции. Клинические показатели демонстрируют значительные различия в их наступлении для группы, которой перорально вводили имитатор IFN по сравнению с группой, которой, вводили 100 ед. мышиного природного IFN-/ менее 0,03, см. фиг. 1/. Два основных рецидива в группе с имитацией IFN происходят в течение семи недель, причем наблюдается усиленный неврологический дефицит. У группы, которой вводили перорально природный мышиный IFN-/ наблюдался замедленный единичный приступ без остаточного неврологического дефицита. Пероральное введение мышиного природного IFN-/ уменьшает тяжесть и снижает групповой показатель между начальным приступом и рецидивом.
Con A активация клеток дренирующих паховых лимфатических узлов оказывается ингибированной у животных, которым вводили перорально имитацию IFN по сравнению с животными, которым вводили 100 ед. мышиного природного IFN-//88222 срм 1910 по сравнению с 38095 срм 3160/. Клетки лимфатических узлов животных, которым вводили имитацию IFN, тестированные с МВР, вырабатывают интенсивную пролиферативную реакцию, но эта реакция существенно ингибируется у животных, которым вводили мышиный природный IFN-/ /14052 срм 842 против 448 срм 50/.
Для определения того, существует ли ингибирование на другой сенсибилизированный антиген, исследуют антиген-специфическую пролиферацию дренирующих паховых лимфатических узлов на второй сенсибилизированный антиген, Mycobacterium tuberculoses homines /MT/, компонент MSCH иноклюма. Клетки лимфатических узлов животных, которым вводили имитацию IFN, вырабатывают интенсивную пролиферативную реакцию на МТ, но эта реакция существенно снижена у животных, которым вводили мышиный природный IFN-/ /52401 срм 857 против 5214 срм 808/.
Животных исследовали также гистологически через 65 дней после иммунизации. Оказалось, что наблюдается заметно меньше воспалительных центров у группы, которой вводили IFN /0,5 0,1/ по сравнению с контрольной группой, которой вводили имитацию IFN /1,8 1,1/р менее 0,05/. Таким образом, мышиные природные интерфероны первого типа при пероральном введении могут подавлять клинические проявления заболевания, уменьшать воспаления и ингибировать пролиферацию в ответ на MBP и MT.
Пример 9Подавление рецидивов при пероральном введении IFN- / коррелируется со снижением IFN- секрецииИнтенсивность заболевания EAE ассоциируется с IFN- секрецией после Con A стимуляции клеток селезенки. Поэтому объединенные клетки селезенки стимулируют /как для животных, перорально обработанных имитатором IFN /н=5/, так и для перорально обработанных 100 ед. /н= 5/ мышиного IFN-/ Con A /2,5 мкг/мл/ при 1106 клеток/мл в течение двух дней, и надосадочные жидкости анализируют в твердофазном ELISA/IL-2, IL-10, IFN- . Пероральное введение IFN-/ последовательно снижает IFN- медиатор воспаления /эксп. 1, имитатор, селезенка 2500 нг/мл 300 против 100 ед., селезенка x1100 нг/мл 200; эксп. N 2, имитатор селезенки 2180 нг/мл 100 ед., селезенки против 247 x143 нг/мл 13, xp менее 0,001 по сравнению с контролем-имитацией IFN/. Наблюдается также снижение IL-2, фактора роста T клеток и увеличение продуцирования IL-10, хотя эти изменения не достигают статистической значимости.
Настоящее изобретение иллюстрирует, что пероральное введение интерферонов первого типа может модифицировать биологическую реакцию CP-EAE при введении после того, как животное выздоравливает после первого приступа. При введении в адекватных дозах, пероральное введение интерферонов типа 1 подавляет клинические рецидивы, снижает воспаления и ингибирует пролиферацию активированных клеток из дренирующих паховых лимфатических узлов, природных участков для высокой частоты активированных T клеток при подкожном введении антигена. Таким образом, интерфероны типа 1 оказываются активными при пероральном способе введения со значительными иммунологическими эффектами в подверженных воздействию иммунных участках. Пероральное введение IFN последовательно снижает Con A индуцированную IFN- секрецию в клетках селезенки. Подавление пролиферации для MBP может происходить даже после промежутка времени 6 недель после инокуляции и спустя 4 недели после начального клинического приступа. Таким образом, пероральное введение цитокинов без защитной матрицы для предотвращения переваривания протеина в ротоглотке и остальной части алиментарного канала способно ингибировать пролиферацию для сенсибилизированных ранее антигенов.
Ослабление установившейся или новой иммунной реакции является важным терапевтическим моментом. Пероральное введение миелиновых антигенов животных может улучшить клиническое течение CR-EAE у крыс и морских свинок и уменьшить патологические изменения на миленовые антигены у CR-EAE. Пероральное введение миелиновых протеинов может понизить тяжесть приступов у мужчин DR2-MS пациентов и частоту MBP специфических клеточных линий у обработанных миелином индивидуумов. Однако в наблюдениях за пациентами и в опытах на животных наблюдается лишь частичное подавление клинических или патологических заболеваний, что дает возможность предположить, что пероральное введение других иммуноактивных веществ может превосходить миелиновые антигены.
Иммуномодуляторный механизм перорального введения интерферона первого типа неизвестен. Модуляторные эффекты Co A активированных лимфоцитов на митогенные реакции клеток нормальных респондеров могут быть аннулированы за счет добавления античеловеческий лейкоцит IFN сыворотки ин витро; это может предотвратить продуцирование ингибирующих факторов, индуцируемых IFN-, например, растворимого супрессора иммунных реакций /SIRS/ и полученного из макрофагов супрессорного фактора 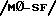 . После такой мутагенной стимуляции могут потребоваться периферализованные T клетки для продуцирования интерферона. Однако пероральное введение низких доз IFN- мышaм не приводит к детектиуемым уровням IFN- в крови, в противоположность с внутрибрюшинным введением, его действие не может быть блокировано циркулирующими анти-IFN антителами. Нейтропенический эффект перорально введенного IFN можно заменить инъекцией клеток крови, но не сыворотки, животным-реципиентам. Анализ мРНК цитокинов CNS при EAE предполагает, что IL-2, IL-6 и IFN- повышены при остром заболевании, но в процессе стабилизации симптомов количество этих цитокинов уменьшается при одновременном повышении уровней IL-10, которые могут регулировать Th 1 клетки. Поэтому IFN типа 1 могут быть иммуномодуляторные молекулы, которые индуцируют супрессорные факторы, такие как IL-10, ингибируя реакцию на иммунизованные антигены, такие как MBP и MT.
Итак, пероральное введение таких модификаторов биологических реакций, как интерфероны первого типа, представляющие потенциально нетоксичные, удобные и непрерывные средства, ингибирующие иммунную реакционноспособность. Модификаторы биологических реакций, вводимые перорально, обеспечивают дополнительные средства для лечения аутоиммунных заболеваний.
Пример 10 . После такой мутагенной стимуляции могут потребоваться периферализованные T клетки для продуцирования интерферона. Однако пероральное введение низких доз IFN- мышaм не приводит к детектиуемым уровням IFN- в крови, в противоположность с внутрибрюшинным введением, его действие не может быть блокировано циркулирующими анти-IFN антителами. Нейтропенический эффект перорально введенного IFN можно заменить инъекцией клеток крови, но не сыворотки, животным-реципиентам. Анализ мРНК цитокинов CNS при EAE предполагает, что IL-2, IL-6 и IFN- повышены при остром заболевании, но в процессе стабилизации симптомов количество этих цитокинов уменьшается при одновременном повышении уровней IL-10, которые могут регулировать Th 1 клетки. Поэтому IFN типа 1 могут быть иммуномодуляторные молекулы, которые индуцируют супрессорные факторы, такие как IL-10, ингибируя реакцию на иммунизованные антигены, такие как MBP и MT.
Итак, пероральное введение таких модификаторов биологических реакций, как интерфероны первого типа, представляющие потенциально нетоксичные, удобные и непрерывные средства, ингибирующие иммунную реакционноспособность. Модификаторы биологических реакций, вводимые перорально, обеспечивают дополнительные средства для лечения аутоиммунных заболеваний.
Пример 10Индуцирование острого экспериментального аутоиммунного энцифаломиелита. EAE индуцируют у 8-10 недельных самок крыс Льюиса /Harlan Sprague Dawley, Indianapolis, IN/, используя эмульсию, содержащую равные части миелинового основного протеина в PBS и полного адьюванта Фреунда /CFA/; Mycobacterium tubereuloses homines H37Pa /MT/ /Difco Labs, Detroit, MI/. Крыс Льюиса иммунизуют подкожными инъекциями в подошву задней лапы 0,1 мл эмульсии, содержащей 50 мкг и 500 мкг MT в адьюванте Фреунда. Начальные клинические признаки болезни наблюдаются между 9 и 11 днями после иммунизации. Клиническую тяжесть начального приступа подразделяют следующим образом /оценка вслепую/: 0 = нет заболевания, 1 = отвислый хвост; 2 = небольшая слабость задних конечностей; 3 = умеренная слабость задних конечностей; 4 = полная слабость задних конечностей и недержание мочи. Средний дневной клинический показатель рассчитывают, определяя средний показатель для каждой группы в каждый день приступа. Кумулятивный клинический показатель экспериментального аллергического неврита был рассчитан, причем он включал полную клиническую тяжесть заболевания, частоту заболевания и длительность клинических признаков в единый сравнительный показатель в результате суммирования дневных показателей по всем крысам в обработанной группе между началом и концом приступа и деления на полное число крыс в обработанной группе. Статистический анализ осуществляют для объединенных данных из по крайней мере двух экспериментов. Пример 11 Введение За семь дней до иммунизации /-7/ и в течение 21 дня после начала /+14/ крысам перорально вводили либо имитацию IFN, либо 1000, 3000 или 5000 ед. крысиного природного IFN- / /цитоиммунный крысиный IFN-+ 4,0105 IRU /мл. Lee Biomolecular Research, Inc. San Diego, CA/ или человеческий рекомбинантный IFN- /hrIFN- / / IFN-11 3,0106 EgH/мл, Schering Corp., Kenilworth, NJ/, используя шприц с gauge boll point иглой /Thomas Seientifie, Swedesboro, NJ/, помещенный в заднюю часть ротоглотки или инъектируемый подкожно с PBS плацебо или 5000 ед. hr IFN- ежедневно. Лиофилизированные IFN препараты воссоздают с DPBS и разбавляют 1/600 до 1/3000. Имитацию природного препарата получают идентично, вводя крысам природный IFN-/ но неиндуцированный Newcastle вирусом.
Получение миелинового основного протеина морских свинок /GP-MBP/, т.е. гомогенизацию и дилапидацию, кислотное элюирование MBP, препараты клеток лимфатических узлов и селезенки, анализы ин витро T клеточной пролиферации и цитокинов осуществляют, как указано ранее.
Пример 12Гистология После умерщвления спинной мозг извлекают и имерсионно фиксируют в 10% нейтральном буферированном формалине в течение минимум двух недель. После фиксации хорды рассекают по всей горизонтальной плоскости с примерно 3 мм интервалами и обрабатывают парафином. Парафиновые блоки рассекают на секции 6-8 мк и поэтапно секции /н=5/ окрашивают гематоксилином, зозином и Luxol – fast blue /PAS/ гематоксилином, и исследуют под оптическим микроскопом. Секции хорд оценивают независимо на предмет определения центров воспаления /слепым методом/ без информации о характере обработки животного до умерщвления. Образцы тканей спинного мозга отбирают идентичным образом для каждого животного, и подсчитывают количество воспалительных центров на секцию /более 20 воспалительных клеток/ в паренхиме. Статистический анализ осуществляют, используя т-тест Стьюдента. Пример 13 Природный крысиный интерферон альфа-бета модифицирует клинический характер заболевания, ингибирует пролиферативные реакции и уменьшает воспаления GP-МВР. Четыре группы по шесть восьми- десятинедельных крыс Льюиса иммунизуют равными частями MBP и CFA, приступы после этого начинаются к 9 дню и продолжаются до 16 дня. За семь дней до иммунизации /-7/ с через 21 день после этого /+14/, каждой группе животных перорально вводят либо имитатор либо 1000, 3000 или 5000 ед. крысиного IFN- / первого типа ежедневно в 0,1 мл PBS. Всех животных оценивают на предмет клинических симптомов заболевания вплоть до 16 дня после иммунизации. Результаты исследований /слепых/ ежедневных групповых клинических показателей демонстрируют значительную разницу в клинических проявлениях у животных, которым вводили имитацию интерферона, и животных, которым вводили 5000 ед. IFN-/, особенно на 13-16 день, но не для животных, которым вводили 1000 или 3000 единиц. У крыс, которым вводили 5000 ед. крысиного природного IFN-/ пик заболевания был менее острым, нежели у группы, которой вводили имитатор интерферона, и эта группа выздоравливала более быстро и вернулась к базовой линии быстрее, нежели группа, обработанная имитатором /фиг. 2, данные для 1000 и 3000 ед. не представлены/. Клинические кумулятивные показатели экспериментального аллергического реврита для животных, обработанных 5000 ед. крысиного природного IFN- , были значительно ниже, нежели у контрольных, которым вводили имитатор IFN /0,8 0,2 при 5000 ед; против 1,20,2 для имитатор IFN, p менее 0,02/. Проводили также гистологические исследования животных через 16 дней после иммунизации. Наблюдается меньше воспалительных очагов у крыс, обработанных IFN-/ /266/по сравнению с группой, которой вводили имитацию интерферона /502/, хотя это не имеет статистической значимости из-за малого числа исследованных образцов спинного мозга на группу /н=3/ p менее 0,06/.
Пролиферация Con A в дренирующих подколенных лимфатических узлах ингибируется с 162091234 срм у животных с имитатором интерферона до 8120765 срм у животных, которым вводили 5000 ед. IFN-/. Клетки дренирующих подколенных лимфатических узлов животных, обработанных имитатором IFN и 5000 ед. IFN были стимулированы иономицином + PMA и демонстрируют уменьшение пролиферации от имитатора /20505 срм505/ до 5000 ед. /6111 срм636/ для обработанных животных. В дренирующих подколенных лимфатических узлах животных, обработанных имитатором и интерфероном, не наблюдалось существенной разницы в MBP или MT пролиферации. Не наблюдалось также ингибирования в селезенке или недренирующих лимфатических узлах брыжжейки Con A или иономицин /PMA у IFN обработанных животных.
Таким образом, образцы специфических интерферонов типа 1 ингибируют тяжесть острых клинических заболеваний при введении в соответствующих дозах. Ингибирование пролиферации за счет перорально введенного IFN-/ может быть связано с непосредственным действием IFN, который попадает в поток крови через кишечник или непосредственно. Ин витро обработка интерфероном типа 1 клеток дренирующих подколенных лимфатических узлов и клеток селезенки, взятых от иммунизованных, но имитационно обработанных животных, демонстрирует эффекты, аналогичные обработке ин виво. Не наблюдается четкого воздействия на Con A пролиферацию для клеток дренирующих подколенных лимфатических узлов или клеток селезенки, экспонированных IFN /1-100 ед./ ин витро, в противоположность ин виво пероральному введению IFN, при котором пролиферация дренирующих подколенных лимфатических узлов была понижена. Таким образом, косвенный эффект IFN первого типа может осуществляться за счет лимфоидных тканей, связанных с кишечником.
Пример 14Модификация острого крысиного EAE за счет перорального введения человеческого рекомбинантного интерферона альфа. Так как человеческий интерферон первого типа может демонстрировать перекрестно-специфическую активность у мышей, морских свинок, гнотобиотных телят, лошадей и свиней, и кошек, был использован рекомбинантный человеческий интерферон hrIFN- / однородный материал, который может обеспечить более эффективную иммуносупрессию на единицу активности, нежели природные препараты. В течение семи дней до иммунизации /-7/ и в течение последующих 21 дня /+14/ каждой группе животных вводили либо PB, либо 1000 или 5000 ед. IFN- ежедневно. Три группы по семь восьми- десятинедельных крыс Льюиса иммунизовали, в результате чего у них наблюдался приступ, начиная с 10 дня, который продолжался до 16 дня. Всех животных наблюдали для оценки клинического характера заболевания до 16 дней после иммунизации. Как видно на фиг. 3, животные, которым вводили PBS и 1000 ед. hrIFN- демонстрировали серьезные острые приступы, начиная с 10 дня, и достигали пиковых значений показателей на 14 день. Животные, которым вводили 5000 ед. IFN-, демонстрировали задержку наступления приступов, пониженную тяжесть в пике и более раннее выздоровление от приступа. Кумулятивные клинические показатели экспериментального аллергического неврита у животных, обработанных 5000 ед. hrIFN- , были значительно ниже, нежели у животных, которым вводили PBS /контроль/ или 1000 ед. /фиг. 4/. Таким образом, пероральное введение hrIFN- подавляет наступление клинических приступов EAE у крыс Льюиса при введении до инокуляции. После умерщвления на 16 день после иммунизации проводили также гистологическое исследование животных. Оказалось, что значительно меньше очагов воспаления приходится на группу, которой вводили по 5000 ед. IFN- по сравнению с PBS контрольной группой или группой, которой вводили 1000 ед. IFN- /фиг. 5/.
Пример 15Пероральное (но при неравных дозах) или парентеральное введение человеческого рекомбинантного интерферона-альфа модифицирует клинический характер болезни Пероральное введение hrIFN- модифицирует клинический характер болезни и уменьшает воспаление спинного мозга. Исследовали относительную эффективность эквивалентных количеств перорально и парентерально вводимого hrIFN-. Три группы по шесть крыс Льюиса были иммунизованы и либо оставлены необработанными, либо им вводили 500 ед. hrIFN-, или им инъектировали подкожно 5000 ед. hrIFN- в течение семи дней, предшествующих иммунизации /-7/, и в течение последующих 21 дня /+14/. Всех животных оценивали по клиническим проявлениям заболевания до 18 дня после иммунизации. У крыс, которым перорально вводили по 5000 ед. hrIFN- нaблюдалось менее тяжелое течение заболевания в пике и более быстрое выздоровление по сравнению с необработанной группой /фиг. 6/. На деле необработанные и подкожно обработанные группы имели весьма сходные показатели клинических кривых, что дает возможность предположить, что подкожное введение hrIFN- оказывает слабое или вовсе не оказывает влияния на клинический характер заболевания. Кумулятивные клинические показатели при экспериментальном аллергическом неврите демонстрируют существенные различия в клинических проявлениях у необработанных/подкожно инъектированных животных по сравнению с животными, которым дозы вводили перорально /фиг. 7/. Не существует заметных различий между необработанными и подкожно инъектированными животными. Через 18 дней после иммунизации животных умерщвляли и проводили гистологическое исследование. Оказаслоь, что значительно меньше очагов воспаления наблюдается в случае 5000 ед. интерферона по сравнению с 5000 ед., подкожно введенными /фиг. 8/.
Аналогичные результаты получены для перорально вводимого и подкожно вводимого PBS. В этом случае четырем группам по шесть крыс Льюиса вводили либо перорально PBS, либо подкожно PBS, либо перорально 5000 ед. hrIFN- либo подкожно 5000 ед. hrIFN- в течение семи дней до иммунизации /-7/ и в течение 21 последующего дня /+14/. Всех животных иммунизуют в день 0. Животных оценивают по характеру клинических симптомов до 16 дня после иммунизации и затем умерщвляют. Кумулятивные клинические показатели экспериментального аллергического неврита оказываются значительно меньше у животных, которым вводили hrIFN- /1,00,2/ по сравнению с теми, которым вводили PBS /2,50,4/ /p менее 0,005/. Не наблюдается существенной разницы между теми животными, которым подкожно вводили имитацию /1,70,4/ и теми, которым вводили подкожно 5000 ед. hrIFN-, хотя даже обработанные подкожным введением животные должны были иметь более высокие кумулятивные клинические показатели при экспериментальном аллергическом неврите. Таким образом, подкожное введение hrIFN- не может модифицировать возникновение острого клинического заболевания при введении в клинически превентивных дозах перорально.
Пример 16Пероральное введение IFN- ингибиpует индуцируемое митогеном IFN- и в дренирующих подколенных лимфатических узлах.
Интенсивность заболевания EAE была связана с IFN- секрецией после Con A стимуляции. Поэтому клетки селезенки и дренирующих подколенных лимфатических узлов /на 18 день после иммунизации от животных, которым перорально вводили имитатор интерферона или 5000 ед. перорально hrIFN- стимулировали Con A /2,5 мкг/мл/ в течение двух дней и надосадочную жидкость анализировали с помощью ELISA. Представленные в таблице 1 результаты демонстрируют, что пероральное введение hrIFN- снижает IFN- медиатор воспаления в дренирующих подколенных лимфатических узлах, но не в селезенке.
Таблица 1Ингибирование клинического проявления заболевания за счет перорального введения IFN коррелирует со снижением секреции IFN- в дренирующих подколенных лимфатических узлах. LN имитация LN 5000 селезенка, имитация селезенки, 5000 IFN- /нг/мл461 60 9632* 36075 31062Клетки селезенки и дренирующих подколенных лимфатических узлов /на 18 день/ от крыс, которым вводили перорально имитатор IFN и 5000 ед. IFN иммунизованных крыс культивируют с Con A /2,5 мкг/мл/ при концентрации 1 106 клеток/мл в течение 48 часов. Собирают надосадочные жидкости через 48 часов после активации Con A и замораживают при -70oC после центрифугирования. Содержание IFN- определяют, используя твердофазный ELISA анализ. Полученные результаты выражают в нг/мл. Представлены объединенные данные из двух экспериментов. LN 5000 сравнивают с LN имитацией xр менее 0,05.
Перорально введенные интерфероны первого типа, в противоположность идентичным дозам, введенным подкожно, раскрыты в настоящем изобретении как модификаторы биологических реакций на MBP в случаях острого EAE у крыс Льюиса, если их вводят перед сенсибилизацией и клиническим приступом. Перорально введенные интерфероны типа 1 частично модифицируют клинические приступы, уменьшают число воспалительных центров в спинному мозгу, снижают неспецифическую пролиферацию за счет Con A и иономицин/PMA и снижают продуцирование IFN- в дренирующих подколенных лимфатических узлах. Это дает возможность предположить, что IFN- более активны при пероральном способе введения, и оказывают определяемый иммунологический эффект, и подтверждает, что специфические цитокины способны ингибировать клинические заболевания при введении через желудочно-кишечный тракт.
Неожиданно оказалось, что в настоящем изобретении работают как человеческий рекомбинантный IFN-, тaк и видоспецифический крысиный природный IFN. Природные IFN представляют собой смеси 14 отдельных подвидов, включая IFN-11 подтип, который может быть лишь небольшим компонентом природного типа. Человеческий IFN демонстрирует некоторую перекрестно-видовую активность, однако исключительное использование IFN-11 подтипа может привести к большему количеству за счет ингибиторной активности/полное количество единиц активности наиболее важной компоненты для иммуносупрессии у крыс.
Антипролиферативное действие перорально вводимых IFN оказывается больше в дренирующих подколенных лимфатических узлах, нежели в недренирующих подколенных лимфатических узлах. Пероральное введение IFN- также ингибирует продуцирование IFN- в дренирующих подколенных лимфатических узлах. Дренирующие подколенные лимфатические узлы представляют природные дренирующие участки для подкожного введения антигенов и поэтому предположительно являются резервуаром высокой частоты сенсибилизированных MBP-специфических T клеток. Пероральное введение цитокинов может преимущественно влиять на пролиферацию и продуцирование цитокинов в местах иммунной активацци по сравнению с системным введением. Ингибирование IFN- секреции в активированном региональном иммунном отделе за счет IFN- может уменьшить воспалительный эффект за счет MBP-специфических клеток в CNS.
Пероральное введение IFN- оказывается эффективным при ингибировании EAE по сравнению с идентичными дозами, вводимымии подкожно. Ин витро IFN не ингибирует Con A пролиферацию в популяции LN в противоположность с ин виво пероральным введением. Так, способ введения оказывается критическим и может определять иммунологический механизм для IFN. Протеины, которые могут не пережить транзит через алиментарный канал, могут все еще демонстрировать иммуноингибиторную активность за счет связанных с кишечником лимфоидных тканей /GALT/ в ротоглотке. Поэтому пероральное введение обеспечивает альтернативную систему доставки лекарств для терапевтических цитокинов при лечении автоиммунных заболеваний за счет выработки иммунорегуляторных клеток через иммунную систему кишечника.
Иммуномодуляторный механизм перорально введенного интерферона первого типа неизвестен. Модуляторный эффект Con A активированных лимфоцитов на митоненные реакции клеток нормальных респондеров можно аннулировать, добавляя античеловеческий лейкоцит IFN сыворотку ин витро, что может модифицировать продуцирование ингибирующих факторов, индуцируемых IFN- , например растворимый супрессор иммунной реакции /STRS/ и супрессорный фактор, полученный из макрофага 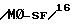 . Периферализованные T клетки могут потребоваться для продуцирования интерферона после такой митогенной стимуляции. Однако пероральное введение низких доз IFN мышам не приводит к детектируемым уровням IFN в крови, в противоположность случаям внутрибрюшинных инъекций; нельзя этот эффект блокировать и циркулирующими анти-IFN антителами. Нейтропенический эффект перорально вводимых IFN можно перенести за счет инъекции кровяных клеток но не сыворотки животным-реципиентам. Аналогично, клетки можно индуцировать в пейеровых бляшках после перорального введения антигена мышам, что снижает гуморальную реакцию ин витро. Поэтому IFN первого типа может быть иммуномодуляторной молекулой, продуцируемой активированными T и другими иммунными клетками, которые индуцируют супрессорные факторы, такие, как и M . Периферализованные T клетки могут потребоваться для продуцирования интерферона после такой митогенной стимуляции. Однако пероральное введение низких доз IFN мышам не приводит к детектируемым уровням IFN в крови, в противоположность случаям внутрибрюшинных инъекций; нельзя этот эффект блокировать и циркулирующими анти-IFN антителами. Нейтропенический эффект перорально вводимых IFN можно перенести за счет инъекции кровяных клеток но не сыворотки животным-реципиентам. Аналогично, клетки можно индуцировать в пейеровых бляшках после перорального введения антигена мышам, что снижает гуморальную реакцию ин витро. Поэтому IFN первого типа может быть иммуномодуляторной молекулой, продуцируемой активированными T и другими иммунными клетками, которые индуцируют супрессорные факторы, такие, как и M -SF, ингибирующие реакции на иммунизованные антигены, такие как MBP.
Настоящее изобретение демонстрирует, что пероральная IFN- терапия подходит для рецидивирующего-ремитирующего рассеянного склероза и других хронических не неврологических автоиммунных заболеваний, так как перорально введенный IFN- по-видимому, более эффективен, нежели эквивалентные дозы парентерально введенного IFN-. Так, пероральное введение модификаторов биологической реакции цитокинов, таких как IFN- обеспечивает эффективные средства для модификации клинических приступов в ответ на антигены при введении перед сенсибилизацией. Пероральный способ введения является удобной системой доставки лекарств, которая позволяет использовать значительно более низкие дозы цитокинов, минимизирует побочные эффекты и повышает эффективность.
Пример 17 -SF, ингибирующие реакции на иммунизованные антигены, такие как MBP.
Настоящее изобретение демонстрирует, что пероральная IFN- терапия подходит для рецидивирующего-ремитирующего рассеянного склероза и других хронических не неврологических автоиммунных заболеваний, так как перорально введенный IFN- по-видимому, более эффективен, нежели эквивалентные дозы парентерально введенного IFN-. Так, пероральное введение модификаторов биологической реакции цитокинов, таких как IFN- обеспечивает эффективные средства для модификации клинических приступов в ответ на антигены при введении перед сенсибилизацией. Пероральный способ введения является удобной системой доставки лекарств, которая позволяет использовать значительно более низкие дозы цитокинов, минимизирует побочные эффекты и повышает эффективность.
Пример 17Иммунизация Самок крыс Льюиса /Harlan/ 150-170 г весом иммунизуют 20 мг /влажный вес/ периферического нервного миелина, эмульгированного в равном объеме полного адьюванта Фреунда /CFA/, который содержит 10 мг. M. tuberculoses на мл адьюванта. 200 мкл иммуногена инъектируют в подошву правой задней лапы. Миелин периферических нервов выделяют из бычьих спинальных корешков /Pel-Freeze, TX/ способом Norton. Methods Enzymol., 31 /par A/, 435, 1975. Пример 18 Введение интерферона Начиная за семь дней до иммунизации и до умерщвления на 20 день после иммунизации, крысам вводят либо имитацию IFN, либо 5000 ед. крысиного природного IFN- /. Цитоиммунный крысиный IFN-/ 1,5106 ед/мл. Lee Biomolecular Researeh Inc., San Diego, CA/. Лиофилизированный IFN восстанавливают за счет стерильной воды, разбавленной в фосфатном буферированном солевом растворе /PBS/ и вводят в ежедневной дозе 0,1 мл, используя шприц с иглой 20 gauge ball point/Thomas Seientific, Swedesboro, NJ/, помещенный в заднюю часть ротоглотки.
Пример 19Клинические оценки Крыс взвешивают ежедневно, начиная с 7 дня после иммунизации, и проводят клиническую оценку, используя следующую схему: 0, норма; 1 = вялый хвост; 3 умеренный парапорез; 4=серьезный парапорез; 5 = параплегия; 6 = параплегия, включающая передние конечности; 7 = параплегия, включающая передние конечности и нарушения дыхания; 8 = агония или смерть в соответствии с определением Hahn et al. Аста Neuropathol. 82 : 60 – 65 /1991/. Потерю веса рассчитывают для каждого животного как разницу между весом в момент умерщвления и максимальным весом и выражают как процент от максимального веса. Пример 20 Пролиферационный анализ При умерщвлении на 20 день после иммунизации извлекают дренирующие подколенные лимфатические узлы и селезенку. Суспензии отдельных клеток получают, пропуская ткань через сетку из нержавеющей стали с диаметрами ячейки 90 мкм. Лизис эритроцитов осуществляют в суспензии клеток селезенки, добавляя 2 мл ACK раствора к осадку. Клетки из селезенки и дренирующих лимфатических узлов собирают для каждой из обрабатываемых групп для ин витро культуры. Полные популяции клеток селезенки или лимфатических узлов инкубируют при 2 105 кл/мл с добавлением M.tuberculosis при 10 мкг/мл, SP26 при 1 мкг/мл /SP26 представляет собой нейротогенный пептид из 26 аминокислот/ Rostami et al., J. Neuroimmunol. , 30 : 145 – 151 /1990/, синтезированный в Department of Analitical Chemistry at the Uneversity of Texas/, Конканавалин A /Con A/ при 2,5 мкг/мл /Sigma Chemical Co., St.Louis, MO/ в RPMI (Gibco, Crand Island, NY), иономицина при 100 нг/мл /Calbiochem, La Jolla, CA/ в сочетании с пальмитиловым сложным эфиром миристиновой кислоты /PMA/ при 1 нг/мл /Сигма/, дополненным 10% сывороткой плода теленка /FCS, Whittaker Bioproducts, Walkersville, MD/, 1 мМ пирувата натрия /Gibco, Grand Island, NY/, 2 мМ глутамина /Gibco/, 100 ед. пенициллин/стрептомицина и 50 мкМ 2-меркаптоэтанола. Пластины инкубируют в 5% CO2 во влажной среде при 37oC в течение 4 дней. В этот момент клетки обрабатывают 2 мкКи, меченой тритилом /3Н/ dTd, и собирают спустя 18 часов с помощью автоматического сборника. Захват /3Н/ dTd определяют с помощью жидкостного сцинтилляционного счетчика Бекмана. Культуры ведут по три экземпляра и результаты выражают как CMP.
Пример 21Анализ цитокинов Клетки объединенных дренирующих подколенных лимфатических узлов и объединенных селезенок от животных, которых обрабатывали имитацией интерферона или 5000 ед. крысиного IFN с экспериментальным аллергическим невритом культивируют с Con A /2,5 мкг/мл, M. Tuberculosis /10 мкг/мл/ или PS26 /1 мкг/мл/ при 1 106 клеток/мл в 75 см2 склянке для культуры тканей в течение 48 часов во влажной среде инкубатора, содержащей 5% CO2/ 95% воздуха при 37oC. Надосадочную жидкость собирают после центрифугирования, и замораживают при -70oC. Содержание IFN- определяют, используя твердофазный ELISA анализ. Анти-IFN- /Pharmingen, San Diego, CA/ инкубируют на поливиниловых микротитровальных пластинах с 96 ячейками 0,01М карбонатным буфером /pH 9,6/ в течение ночи при 4oC. Пластины блокируют 3% альбумином бычьей сыворотки /BSA/ в PBS в течение 3 часов. Добавляют по 100 мкл надосадочной жидкости в различных разбавлениях, оттитрованных для линейного участка кривой поглощение/концентрация/ по три эксперимента и инкубируют в течение 1 часа при комнатной температуре. Затем пластины промывают 5 раз PBS Tween /0,05% и добавляют в течение 60 минут 100 мкл коньюгированного с пероксидазой IFN- мoнoклoнaльнoгo антитела /с эпитопными детерминантами, отличающимися от первого антитела, использованного для нанесения на поливиниловую пластину/ в концентрации 1: 1000. Затем к пероксидазному субстрату добавляют О-фенилендиаминдигидрохлорид, и поглощение измеряют при 450 нм. Получают стандартные кривые для различных количеств IFN-.
Пример 22Гистология Крыс умерщвляют за счет перфузии физиологического раствора при анестезии фенобарбиталом на 20 день после иммунизации. Хвост удаляют и делят. Дистальную часть фиксируют в 2,5% параформальдегид /2,5% глутаральдегиде в PBS в течение 4-6 часов, обрабатывают осмием в течение ночи в 2% тетроксиде осмия, дегидратируют и вводят в Epon. От каждого животного приготавливают от 4 до 5 блоков нижних люмбосакральных корешков. Представительные 0,5 мк срезы каждого из блоков окрашивают толуидином синим. Гистологические исследования демиелинизации и воспалений в каждом корешке осуществляют, используя следующую шкалу оценок: /Hahn et al., 1991/: демиелинизация; 1 + = нескольно демиелинизированных аксонов первивенулярно или рассеяно; 2+ = много центров перивенулярной демиелинизации; +3 = интенсивное демиелинизирование перивенулярно и конфлюентно; воспаление; 1+ = несколько рассеянных моноядерных воспалительных клеток часто субпериневрально 2+ = перивенулярное образование “манжетки” с моноядерными воспалительными клетками; 3+ = интенсивное многоцентровое перивенулярное образование “манжеток” и широкое распространение эндоневрального воспаления. Показатели рассчитывают для каждого животного, разделив общее количество демиелинизирования и воспалений на число нервных корешков в срезах. Слайды считывают с помощью исследователя /MP/, который не знает протокола иммунизации и обработки, предшествующих получению образцов. Проксимальную часть конского хвоста / 2 см/ замораживают в метилбутане, охлажденном жидким азотом и хранят при -70oC. Восьмимикронные поперечные срезы получают в криостате, фиксируют в эфире охлажденным льдом и используют для иммуноцитохимических исследований.
Пример 23Иммуноцитохимические исследования Иммуноцитохимические исследования осуществляют, используя следующие антитела: ED1 1:1000 /макрофаги/, W3/13 /лимфоциты; Harlan Bioproduets for Seience, IN), CDll b/c 1: 100 /CR3 на макрофагах, гранулоцитах, дендритных клетках; Pharmingen, CA/, IFN- 1:10/ Genzyme, MA/. Восьмимикронные замороженные срезы блокируют в сыворотке нормальных кроликов в течение 30 минут, затем инкубируют с первичным антителом в течение 1 часа, промывают PBS, инкубируют с биотинилированным вторичным антителом в течение 1 часа, промывают в PBS, инкубируют с ABC пероксидазой /Vectastain/ в течение 5 минут, промывают и проявляют диаминобензидином и перекисью водорода. Блокирование 0,01% перекисью водорода предшествует инкубированию с первичными антителами к IFN- Интенсивность и распределение окрашивания нервных корешков оценивают как 1 + слабое; 2 + умеренное и 3 + сильное.
Пример 24Клинические симптомы Крысы начинают терять вес на 10 день после иммунизации, а первые неврологические симптомы появляются на 12 день, тогда как выздоровление начинается на 16 день или позже. У 11 из 22 крыс /50%/ развивается тяжелый экспериментальный аллергический неврит с клиническим показателем 4 /тяжелый парапорез/ или выше, и 2 из 11 животных, которым вводят имитацию, умирают на 14 и 15 день после иммунизации, тогда как ни одно из животных, которым вводили ШАТ- / не погибло. Клинический показатель в группе с имитацией IFN /н= 11, показатель 4,12,4; экспериментальный аллергический неврит /был значительно выше, нежели в группе крыс, которым вводили IFN-/ /н=11, показатель 3,31,4; p менее 0,03, тест Уилкинсона/. Большее количество крыс в группе с имитацией IFN демонстрировало клинический показатель 4 или выше /фиг. 9A/. Аналогично, большее количество крыс из группы, которой вводили имитатор IFN потеряло более 25% веса, нежели из группы, которой вводили IFN-/ /фиг. 9B/.
Пример 25Клеточная пролиферация и цитокины Пролиферация клеток селезенки на Con A у животных, которым вводили IFN- / была снижена по сравнению с животными, которым вводили имитатор IFN /селезенка: у группы с имитатором 11811415194 против крыс с IFN-/ 83594, среднее SEM, p менее 0,006, т-тест/.
Продуцирование IFN- после стимуляции за счет P26 клетками лимфатических узлов и селезенки у крыс, которым вводили имитацию IFN была гораздо выше, нежели продуцирование IFN- клетками лимфатических узлов и селезенки у крыс, которым вводили IFN-/ по данным ELISA, но разница не достигает статистической значимости /лимфатические узлы: 18,92,4 нг/мл против 11,93, нг/мл; селезенка: 11,90,4 нг/мл против 6,3 1,8 нг/мл, среднее SEM, у крыс с имитацией против крыс IFN-/ p<0,08/. После иммунизации Con A или M. tuberculosis не наблюдалось никаких различий в продуцировании IFN-Пример 26 Гистология Гистологические исследования включенных в Epo срезов, показывает, что демиелинирование значительно менее серьезно у крыс, которым вводили IFN- / /показатель 0,880,11 среднее +SD/ по сравнению с крысами, которым вводили имитатор IFN /показатель 1,10,26; p менее 0,05, т-тест/, но воспаление в этих группах заметно не отличается /показатель 1,00,2 против 1,180,24/.
Иммуноцитохимия с CDllbc антителами показывает гориздо большее число воспалительных клеток как первазикулярно, так и диффузно инфильтрующих нервные корешки конского хвоста как у крыс, которым вводили имитатор IFN, так и у крыс, которым вводили IFN-/, когда их умертвили на 20 день. Основная часть этих клеток была фагоцитарно ED1 позитивными макрофагами, и не наблюдалось заметной разницы между группами животных которым вводили имитатор IFN или IFN-/ в степени воспалений.
Группы крыс, обработанных либо крысиным /н=3, либо имитатором IFN /н=3/, умертвили раньше в ходе заболевания на 13 день после иммунизации. Иммуноцитохимические исследования должны были оценить IFN- продуцирование ин ситу, которое видимо только до 14 дня для экспериментального аллергического неврита, а также оценить воспаление. Инфильтрация ED1 позитивных макрофагов и W 3/13 позитивных лимфоцитов была гораздо менее сильной у крыс, которым вводили IFN-/, нежели у крыс, которым вводили имитатор IFN /контроль/. IFN- окраска присутствовала на участках плотного воспаления на поверхности воспалительных клеток. Фиг. 10 демонстрирует, что IFN- окрашивание заметно меньше у крыс, которым вводили параллельно с менее интенсивным воспалением.
Пероральное введение видо-специфических IFN-/ по сравнению с имитатором IFN снижает тяжесть экспериментального аллергического неврита, вызывает заметное уменьшение клинического показателя заболевания и приводит к меньшей потере веса. Гистологические данные, полученные на 20 день после иммунизации, когда уже началось выздоровление, показывают меньшую демиелинизацию у группы, обработанной IFN-/ без изменения степени воспаления. На 13 день после иммунизации, ин ситу, IFN- продуцирование было понижено наряду с воспалениями, по данным иммуноцистохимии. Исследования пролиферации клеток дренирующих лимфатических узлов и селезенки дает пониженную пролиферацию для Con A у IFN-/ обработанных крыс с экспериментальным аллергическим невритом по сравнению с крысами, которым вводили имитатор IFN. Анализ цитокинов выявил сниженное, продуцирование IFN- после стимуляции SP26, антигенов миелина периферических нервов, способным индуцировать экспериментальный аллергический неврит. В целом, настоящее изобретение демонстрирует, что пероральное введение IFN-/ снижает тяжесть экспериментального аллергического неврита за счет уменьшения продуцирования IFN-. Таким образом, настоящее изобретение демонстрирует иммуномодуляторный эффект перорального введения IFN-/ на иммунную систему хозяина.
Механизм действия перорального введения IFN-/ на иммунную систему неизвестен, но может включать действие на пейеровые бляшки в лимфоидных тканях, связанных с кишечником /CALT/, где могут вырабатываться регуляторные клетки. IFN- может индуцировать иммунорегуляторные факторы, полученные из CD8 + T клеток, которые ответственны за модификацию заболевания. Снижение продуцирования IFN-, связанное с пероральным введением IFN-/ предполагает возможное ингибирование функций системных T 1-подобных хелперных T клеток, обнаруженных при EAE, которые продуцируют IFN-. Это приводит к снижению T клеточной энцефалитогенетичности активно или пассивно индуцированного заболевания.
Как было показано, парентеральное введение – IFN- усиливает как индуцированный миелином, так и опосредованный T клетками экспериментальный аллергический неврит при усиленных клинических признаках и гистологических нарушениях, а также повышает оксидативное прорастание макрофагов, тогда как противоположный эффект наблюдается при парентеральном введении анти IFN- антитела. IFN- индуцирует также MHC класса 1 и 11 экспрессию на клетках Schwann ин витро. IFN- воспалительный цитокин, выделяемый Th1 CD4+T клетками, может усиливать MHC класса 11 экспрессию на макрофагах и эндотелиальных клетках и активировать макрофаги, которые играют важную роль в фагоцитозе миелина. Сниженная IFN- экспрессия и уменьшенное воспаление наблюдаются при приступах клинической болезни у животных, которым вводили IFN-/. Гистологическая оценка в конце процесса болезни, когда началось выздоровление, демонстрирует снижение демиелинизирования, но не воспаления у животных, которым вводили IFN-/. Снижение IFN- и уменьшение воспаления на ранних стадиях и уменьшение демиелинизирования на поздних стадиях заболевания предполагают критическую роль, IFN- в патогенезе экспериментального аллергического неврита. Невозможность уменьшить воспаление на поздних стадиях заболевания может быть связана с частичным подавлением IFN- продуцирования, влиянием других цитокинов и медиаторов воспаления, а также индуцированием слишком тяжелого экспериментального неврита.
В итоге, вводимый перорально интерферон является модификатором биологических реакций, эффективным для модификации заболевания в другое экспериментальное автоиммунное заболевание, экспериментальный аллергический неврит. Результаты настоящего изобретения в экспериментальном аллергическом неврите, демонстрирующие ослабление болезни у крыс, которым перорально вводят IFN-/ иллюстрируют, что пероральное введение IFN-/ должно быть полезно при лечении иммуно-опосредствованных невропатий у человека, таких как синдром Гийена-Барре или хроническая воспалительная демиелинизирующая полирадикулоневропатия /CIDP/.
Все патенты и публикации, упомянутые в этом описании, являются указателями предшествующего уровня для специалистов имеющим отношение к настоящему изобретению. Все патенты и публикации включены в качестве ссылок.
Специалисту легко понять, что настоящее изобретение хорошо адаптировано для осуществления целей и достижения указанных результатов и преимуществ. Приведенные примеры, а также способы, процедуры, обработки, молекулы и конкретные соединения, описанные здесь, являются представительными или предпочтительными вариантами, являются примерами и никоим образом не ограничивают объем изобретения. Те изменения и другие применения, которые очевидны специалистам, включены в суть изобретения, определенную объемом формулы изобретения.
Формула изобретения
РИСУНКИ
MM4A – Досрочное прекращение действия патента СССР или патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе
Дата прекращения действия патента: 12.04.2009
Извещение опубликовано: 20.07.2010 БИ: 20/2010
|
||||||||||||||||||||||||||