Патент на изобретение №2376028
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) СПОСОБ УМЕНЬШЕНИЯ ОКИСЛИТЕЛЬНОГО ПОВРЕЖДЕНИЯ (ВАРИАНТЫ)
(57) Реферат:
Изобретение относится к медицине, а именно к фармакотерапии, и может быть использовано для уменьшения окислительного повреждения у млекопитающего в удаленном органе или клетке. Для этого вводят ароматические катионные пептиды, выбранные из группы 2′,6′-Dmt-D-Arg-Phe-Orn-NH2 (SS-05), 2′,6′-Dmt-D-Cit-Phe-Lys-NH2 (SS-29), Phe-D-Arg-2′,6′-Dmt-Lys-NH2 (SS-30), D-Arg-2′,6′-Dmt-Lys-Phe-NH2 (SS-31) и 2′,6′-Dmt-D-Arg-Phe-(2-аминогептановая кислота)-NH2 (SS-32) в эффективном количестве. Изобретение расширяет арсенал средств, обладающих антиоксидантной активностью и способных проникать не только через гематоэнцефалический барьер и клеточные мембраны, но и в митохондрии. 3 н. и 28 з.п.ф-лы, 2 табл., 17 ил.
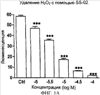
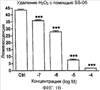
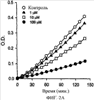
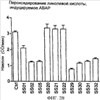
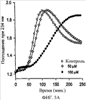
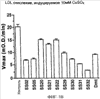
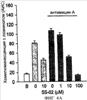
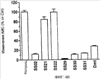
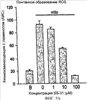
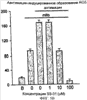
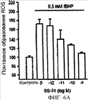
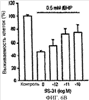
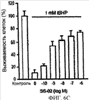
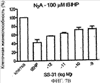
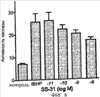
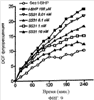
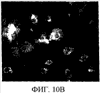
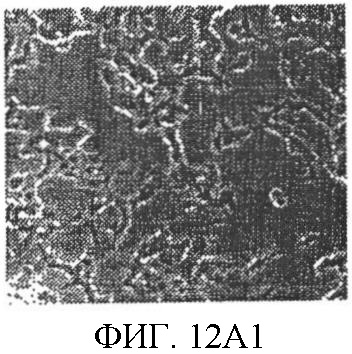
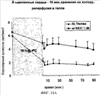
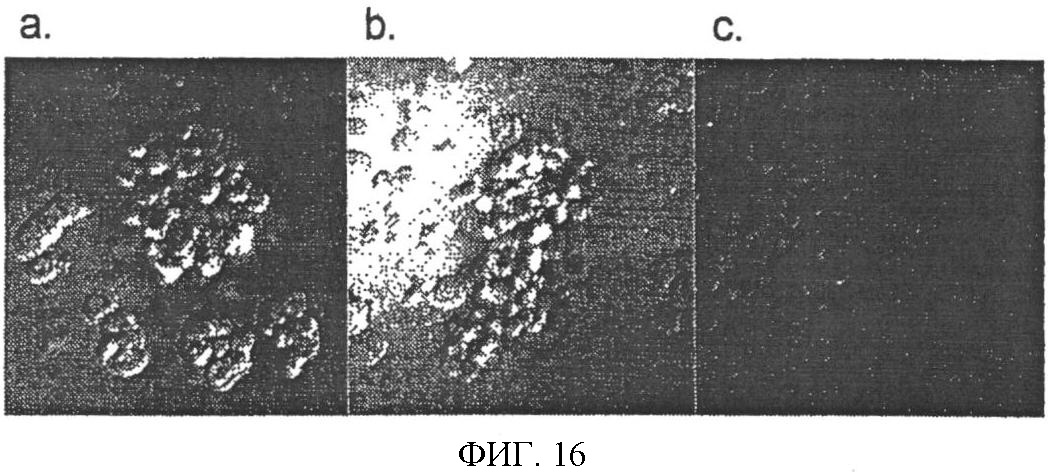
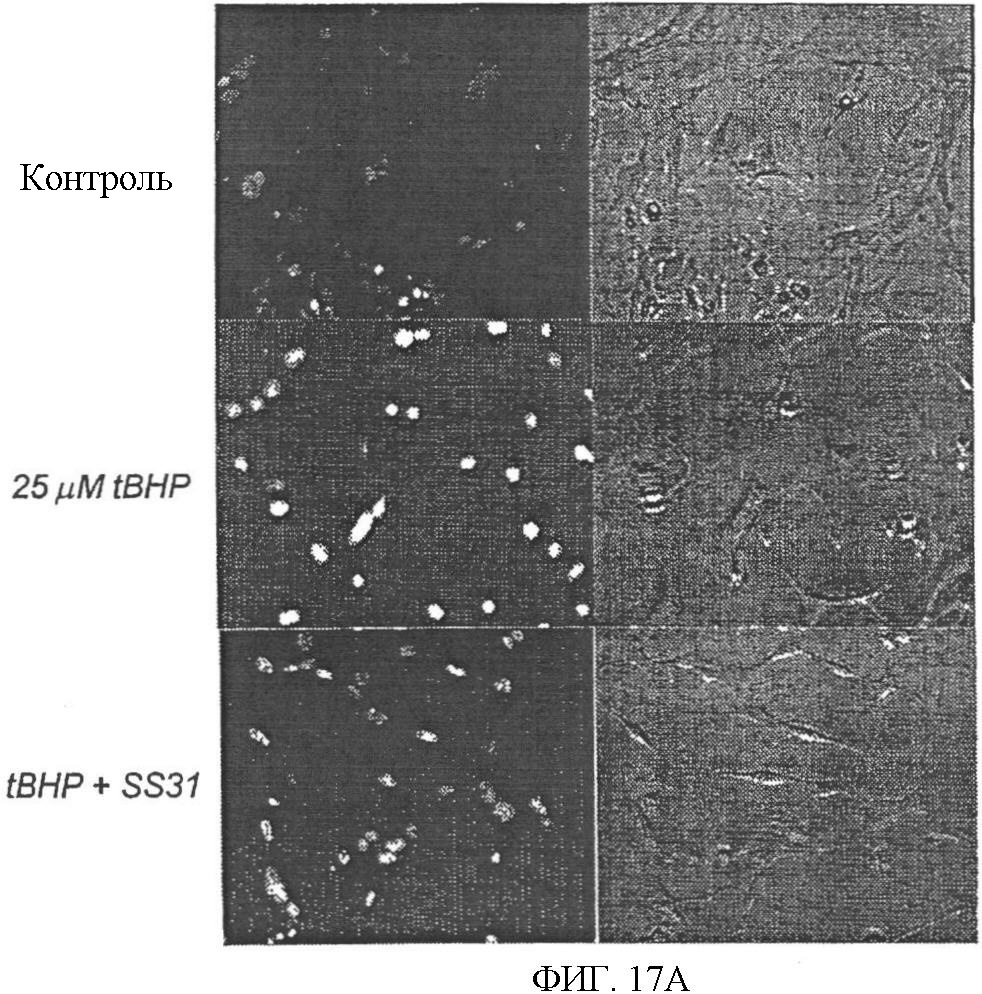
Уровень техники Митохондрии необходимы для выживания клеток как главные поставщики АТР посредством окислительного фосфорилирования. Однако дыхательная цепь митохондрий является также главным источником окислительных свободных радикалов. Например, образование радикалов может происходить как результат взаимодействия митохондриальных носителей электронов, таких как убихинон, с кислородом с получением супероксида. Супероксиды посредством дисмутации реагируют с пероксидом водорода, который может отщеплять гидроксильный радикал. Кроме того, супероксиды взаимодействуют с окисью азота, образуя при этом пероксинитрит и другие реакционноспособные оксиданты. Старение связывают не только с повышенным образованием реакционноспособных кислороднесущих окислителей (ROS), но также и с ослаблением эндогенных механизмов антиоксидантой защиты. Митохондрии в особенности уязвимы при окислительном стрессе, поскольку они непрерывно подвергаются действию ROS. Как следствие, разрушение митохондрий часто связывают со старением. Свободные радикалы, в том числе ROS, и реакционноспособные азотсодержащие их аналоги (RNS) вызывают различные неспецифические повреждения биологических молекул, включая липиды, протеины, РНК и ДНК. Такие повреждения этих веществ участвуют или приводят к различным клиническим расстройствам, например, атеросклерозу, преэклампсии, болезни Альцгеймера, болезни Паркинсона и артриту. Антиоксидантная терапия в принципе может замедлить процесс старения и быть благотворной в отношении множества заболеваний и состояний из числа тех, что указаны выше. Однако разработка конкретных методов лечения с помощью митохондрий сдерживается трудностями доставки молекул антиоксидантов к митохондриям in vivo. Например, такая молекула сначала должна быть доставлена через плазменный барьер в цитоплазму, а затем селективно направлена именно в митохондрии. Ни одно из существующих ныне доступных антиоксидантных средств не может быть доставлено непосредственно в митохондрии. Эндогенные антиоксиданты, супероксидцисмутаза и каталаза, в малой степени абсорбируются во рту, имеют короткое «время полужизни» и не проходят через барьер «кровь-головной мозг». Природные антиоксиданты (например, витамин Е, коэнзим Q, полифенолы) являются водонерастворимыми и имеют тенденцию накапливаться в клеточных мембранах и медленно преодолевают барьер «кровь-головной мозг». Таким образом, есть потребность в усовершенствованных способах уменьшения окислительного повреждения с помощью соединений, обладающих свойствами антиоксидантов, которые проходят через мембраны клеток. Кроме того, преимуществом таких веществ со свойствами антиоксидантов следует считать возможность их конкретного нацеливания в митохондрии. Сущность изобретения Эта и другие задачи изобретения могут быть решены с помощью настоящего изобретения, в котором обеспечивается способ уменьшения окислительного повреждения у нуждающегося в этом млекопитающего. Способ включает введение млекопитающему эффективного количества ароматического катионного пептида. Этот ароматический катионный пептид имеет (а), по крайней мере, один результирующий положительный заряд; (b) минимум три аминокислоты; (с) максимум около двадцати аминокислот; (d) соотношение между минимальным числом результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r), где 3pm представляет собой наибольшее число, которое меньше или равно r+1; (е) соотношение между минимальным числом ароматических групп (а) и общим числом результирующих положительных зарядов (pt), где 3а представляет собой наибольшее число, которое меньше или равно pt+1, за исключением того, что когда а является 1, pt может быть также 1; (f) по крайней мере, один аминокислотный остаток тирозина или триптофана. В другом варианте изобретение также обеспечивает способ уменьшения окислительного повреждения в удаленном органе млекопитающего. Этот способ включает введение в удаленный орган эффективного количества ароматического катионного пептида. Ароматический катионный пептид имеет (а), по крайней мере, один результирующий положительный заряд; (b) минимум четыре аминокислоты; (с) максимум около двадцати аминокислот; (d) соотношение между минимальным числом результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r), где 3pm представляет собой наибольшее число, которое меньше или равно r+1; (е) соотношение между минимальным числом ароматических групп (а) и общим числом результирующих положительных зарядов (pt), где 2а представляет собой наибольшее число, которое меньше или равно pt+1, за исключением того, что когда а является 1, pt может быть также 1; (f) по крайней мере, один аминокислотный остаток тирозина или триптофана. В другом варианте изобретение обеспечивает способ уменьшения окислительного повреждения у млекопитающего, нуждающегося в этом. Способ включает введение млекопитающему эффективного количества ароматического катионного пептида. Ароматический катионный пептид имеет (а), по крайней мере, один результирующий положительный заряд; (b) минимум три аминокислоты; (с) максимум около двадцати аминокислот; (d) соотношение между минимальным числом результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r), где 3pm представляет собой наибольшее число, которое меньше или равно r+1; (е) соотношение между минимальным числом ароматических групп (а) и общим числом результирующих положительных зарядов (pt), где 2а представляет собой наибольшее число, которое меньше или равно pt+1, за исключением того, что когда а является 1, (pt) может быть также 1; (f) по крайней мере, один аминокислотный остаток тирозина или триптофана. Еще в одном варианте изобретение обеспечивает способ уменьшения окислительного повреждения в удаленном органе млекопитающего. Способ включает введение в удаленный орган млекопитающего эффективного количества ароматического катионного пептида. Ароматический катионный пептид имеет (а), по крайней мере, один результирующий положительный заряд; (b) минимум три аминокислоты; (с) максимум около двадцати аминокислот; (d) соотношение между минимальным числом результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r), где 3pm представляет собой наибольшее число, которое меньше или равно r+1; (е) соотношение между минимальным числом ароматических групп (а) и общим числом результирующих положительных зарядов (pt), где 3а представляет собой наибольшее число, которое меньше или равно pt+1, за исключением того, что когда а является 1, pt может быть также 1; (f) по крайней мере, один аминокислотный остаток тирозина или триптофана. Еще в одном варианте изобретение обеспечивает способ уменьшения окислительного повреждения в клетке, нуждающейся в этом. Ароматический катионный пептид имеет (а), по крайней мере, один результирующий положительный заряд; (b) минимум три аминокислоты; (с) максимум около двадцати аминокислот; (d) соотношение между минимальным числом результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r), где 3pm представляет собой наибольшее число, которое меньше или равно r+1; (е) соотношение между минимальным числом ароматических групп (а) и общим числом результирующих положительных зарядов (pt), где 3а представляет собой наибольшее число, которое меньше или равно pt+1, за исключением того, что когда а является 1, pt может быть также 1; (f) по крайней мере, один аминокислотный остаток тирозина или триптофана. В другом варианте изобретение обеспечивает способ уменьшения окислительного повреждения в клетке, нуждающейся в этом. Ароматический катионный пептид имеет (а), по крайней мере, один результирующий положительный заряд; (b) минимум три аминокислоты; (с) максимум около двадцати аминокислот; (d) соотношение между минимальным числом результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r), где 3pm представляет собой наибольшее число, которое меньше или равно r+1; (е) соотношение между минимальным числом ароматических групп (а) и общим числом результирующих положительных зарядов (pt), где 2а представляет собой наибольшее число, которое меньше или равно pt+1, за исключением того, что когда а является 1, pt может быть также 1; (f) по крайней мере, один аминокислотный остаток тирозина или триптофана. Краткое описание графических материалов Фиг.1 (A) SS-02 и (В) SS-05 (в зависимости от концентрации) обеспечивают удаление H2O2. Фиг.2 (А) SS-02 (в зависимости от концентрации) ингибирует пероксидирование линолевой кислоты, вызываемое АВАР, и (В) SS-02, SS-05, SS-29, SS-30, SS-31, SS-32 и Dmt уменьшали скорость пероксидирования линолевой кислоты, вызываемого АВАР. Фиг.3 (А) SS-02 (в зависимости от концентрации) ингибирует LDL окисление, вызываемое 10 мМ CuSO4 и (В) SS-02, SS-05, SS-29, SS-30, SS-31, SS-32 и Dmt уменьшали скорость LDL окисления. Фиг.4 (A) SS-02 ингибирует продуцирование пероксида водорода в митохондриях, определенное хемилюминесценцией с люминолом в базальных условиях и при стимуляции антимицином. (В) SS-02, SS-29, SS-30 и SS-31 уменьшали спонтанное выделение пероксида водорода в изолированных (выделенных) митохондриях. Фиг.5 (А) SS-31 ингибирует спонтанное продуцирование пероксида водорода выделенными митохондриями и (В) SS-31 ингибирует продуцирование пероксида водорода, стимулированное антимицином. Фиг.6 SS-31 (в зависимости от концентрации) снижал внутриклеточные ROS (разновидности реакционноспособного кислорода) и (В) повышал выживаемость в N2A клетках, которые выдерживали при высоких дозах прооксидантного t-бутилгидропероксида (t-BHP; 0,5 мМ), (С) SS-02 (в зависимости от концентрации) также повышал клеточную выживаемость при выдерживании N2A-клеток с 1 мМ t-ВНР. Фиг.7 SS-31 (в зависимости от концентрации предотвращал потерю клетками жизнеспособности, вызванную воздействием низких доз t-BHP (0,05-0,1 мМ) в нейронах (A) SY-SY5Y и (В) N2A клетках. Фиг.8 SS-31 (в зависимости от концентрации) уменьшал долю клеток, показывающих повышенную активность каспазы после обработки N2A-клеток низкой дозой t-BHP в течение 12 часов. Фиг.9 SS-31 (в зависимости от концентрации) уменьшал скорость аккумуляции ROS в N2A клетках при обработке 0,1 мМ t-BHP в течение 4 часов. Фиг.10 SS-31 ингибировал пероксидирование липидов, вызываемое обработкой N2A клеток 1 мМ t-BHP в течение 1 часа. (А) необработанные клетки; (В) клетки, обработанные 1 мМ t-BHP в течение 3 часов; (С) клетки, обработанные 1 мМ t-BHP и 10 нМ SS-31 в течение 3 часов. Фиг.11 SS-31 предотвращал деполяризацию митохондрий и аккумуляцию ROS в N2A клетках, обработанных t-BHP. Фиг.12 SS-31 предотвращает апоптоз, вызванный низкой дозой t-BHP. Апоптоз оценивали с помощью конфокальной микроскопии с флуоресцентной пробой Hoechst 33342. (А1) Характерный образец клеток, не обработанных t-BHP. (A2) Изображение, полученное на основе флуоресценции, показывающее несколько клеток с плотными, фрагментированными хроматиновыми структурами, характерными для апоптозных ядер. (В1) Характерный образец клеток, обработанных 0,025 мМ t-BHP в течение 24 часов. (В2) Флуоресцентное изображение, показывающее увеличившееся число клеток с апоптозными ядрами. (С1) Характерный образец клеток, обработанных 0,025 мМ t-ВНР и 1 нМ SS-31 в течение 24 часов. (С2) Флуоресцентное изображение, показывающее уменьшившееся число клеток с апоптозными ядрами. (D) SS-31 (в зависимости от концентрации) уменьшал долю апоптозных клеток, полученных в результате 24-часовой обработки низкой дозой t-BHP (0,05 мМ). Фиг.13А SS-02 и SS-31 уменьшали пероксидирование липидов в изолированных сердцах морских свинок, подвергнутых тепловой реперфузии после кратковременной ишемии. Иммуногистохимический анализ 4-гидрокси-ноненол (HNE) – модифицированных белков в парафиновых срезах сердца морской свинки, аэробно перфузировали в течение 30 минут с буфером, затем подвергали 30-мин ишемии и реперфузировали с (а) буфером, (b) 100 нМ SS-02; (с) 100 нМ SS-20 и (d) 1 нМ SS-31, в течение 90 минут. Тканевые срезы инкубировали с анти-HNE антителом, (е) Фоновый контроль: окрашивание без первичного антитела. Фиг.13В SS-02 и SS-31 уменьшали пероксидирование липидов в изолированных сердцах морских свинок, подвергнутых тепловой реперфузии после кратковременной ишемии. Иммуногистохимический анализ 4-гидрокси-2-ноненол (HNE) – модифицированных белков в парафиновых срезах сердца морской свинки, аэробно перфузировали в течение 30 минут (а) буфером, (b) 100 нМ SS-02; (с) 100 нМ SS-20 и (d) 1 нМ SS-31, затем подвергали 30-минутной ишемии и снова перфузировали в течение 90 минут с соответствующими пептидами. Тканевые срезы инкубировали с анти-HNE антителом, (е) Фоновый контроль: окрашивание без первичного антитела. Фиг.14А SS-31 значительно увеличил коронарный ток в выделенных сердцах морских свинок, подвергнутых тепловой реперфузии после продолжительной (18 час) ишемии в условиях холода. Затемненная площадь относится к 18 час ишемии при 4°С. Фиг.14В Сердца морских свинок перфузировали кардиоплегическим раствором (St. Thomas раствор) без (а) или с (b) нМ SS-31 в течение 3 мин, затем подвергали 18-часовой ишемии на холоде (4°С), (с) фоновое окрашивание без первичного антитела. Сердца затем реперфузировали с буфером при 34°С в течение 90 мин. Фиг.14С SS-31 предотвращает апоптоз в эндотелиальных клетках и миоцитах в выделенных сердцах после продолжительной (18 час) ишемии на холоде. Сердца морских свинок реперфузировали кардиоплегическим раствором (St. Thomas раствор) без или с 1 нМ SS-31 в течение 3 минут и затем подвергали 18-часовой ишемии на холоде (4°С). Затем сердца реперфузировали буфером при 34°С в течение 90 мин. Апоптоз определяли с помощью TUNEL окрашивания (зеленый) и ядра визуализировали с помощью DAPI (голубой). Фиг.15А SS-31 увеличивает выживаемость островковых клеток, выделенных из поджелудочной железы мыши, которую определяли по митохондриальному потенциалу. Митохондриальный потенциал оценивали с помощью TMRM (красный) конфокальной микроскопией. Фиг.15В SS-31 уменьшает апоптоз и повышает жизнеспособность островковых клеток, выделенных из поджелудочной железы мыши, что определялось проточной цитометрией. SS-31 (1 нМ) добавляли ко всем буферным растворам, используемым при выделении на всем протяжении процесса. Апоптоз устанавливали при использовании аннексина Vu на основе некроза пропидиум-иодида (PI). Фиг.16 SS-31 уменьшает окислительное повреждение в панкреатических островковых клетках, вызываемое t-бутилгидроксипероксидом (t ВНР). Панкреатические островковые клетки мыши либо ничем не обрабатывали (а), либо обрабатывали 25 µМ t-BHP без (b) или с 1 нМ SS-31 (с). Митохондриальный потенциал определяли с помощью TMRM (красный), а разновидности реакционноспособного кислорода – с помощью DCF (зеленой) и конфокальной микроскопии. Фиг.17А SS-31 предохраняет допаминовые клетки от МРР+ токсичности. SN-4741 обрабатывали буфером, 50 µМ MPP+ или 50 µM SS-31 в течение 48 часов и факт алоптоза устанавливали флуоресцентной микроскопией с применением Hoechst 33342. Количество сконденсированных фрагментированных ядер существенно увеличилось при обработке МРР+. Конкурентная обработка SS-31 уменьшала число апоптозных клеток. Фиг.17В В зависимости от концентрации SS-31 предотвращал потерю допаминовых нейронов у мышей при обработке МРТР. Три дозы МРТР (10 мг/кг) давали мышам (n=12) в течение 12 часов. Ха 30 мин перед каждой МРТР инъекцией вводили SS-31, а также спустя 1 час и 12 часов после последней МРТР инъекции. Через неделю животных умерщвляли и стриатальные зоны мозга подвергали иммуноокрашиванию для определения тирозингидроксилазной активности (показано черным). Фиг.17С В зависимости от концентрации SS-31 увеличивал уровни стриатального допамина, ДОРАС (3,4-дигидроксифенилуксусная кислота) и HVA (гомованилловая кислота) у мышей, обработанных МРТР. Мышам (n=12) в течение 2 часов давали три дозы МРТР (10 мг/кг). SS-31 вводили за 30 мин до каждой МРТР инъекции, а также спустя 1 час и 12 часов после последней МРТР инъекции. Через неделю животных умерщвляли и определяли уровни допамина, ДОРАС и HVA с помощью жидкостной хроматографии под высоким давлением. Подробное описание изобретения Данное изобретение базируется на неожиданном открытии, что некоторые ароматические катионные пептиды уменьшают окислительное повреждение. Уменьшение степени окислительного повреждения является важным из-за свободных радикалов, таких как ROS и RNS, приводящих к пагубному неспецифическому повреждению липидов, белков, РНК и ДНК. Окислительное повреждение, вызываемое свободными радикалами, связывают с рядом заболеваний и состояний у млекопитающих. Пептиды Ароматические катионные пептиды, которые могут использоваться в данном изобретении, являются водорастворимыми и высокополярными. Несмотря на эти свойства, эти пептиды могут с легкостью проникать через клеточные мембраны. Ароматические катионные пептиды, используемые в данном изобретении, включают минимум три аминокислоты, предпочтительно – четыре аминокислоты, ковалентно связанных пептидными связями. Максимальное число аминокислот, содержащихся в ароматических катионных пептидах настоящего изобретения, составляет около двадцати аминокислот, ковалентно связанных между собой пептидными связями. Предпочтительно максимальное число аминокислот составляет около двенадцати, более предпочтительно – около девяти, самое предпочтительное – около шести. Оптимально, когда количество аминокислот, содержащихся в пептидах, равно четырем. Аминокислоты в ароматических катионных пептидах, используемых в настоящем изобретении, могут быть любыми аминокислотами. В используемом здесь смысле термин «аминокислота» применяется в отношении любой органической молекулы, которая содержит, по меньшей мере, одну аминогруппу и, по меньшей мере, одну карбоксильную группу. Предпочтительно, по меньшей мере, одна аминогруппа находится в Аминокислоты могут быть природного происхождения. Такие природные аминокислоты включают, например, двадцать наиболее распространенных левовращающих (L) аминокислот, обычно входящих в состав белков животного происхождения, например, аланин (Ala), аргинин (Arg), аспарагин (Asn), аспарагиновая кислота (Asp), цистин (Cys), глутамин (Glu), глутаминовая кислота (Glu), глицин (Gly), гистидин (His), изолейцин (Ilеu), лейцин (Leu), лизин (Lys), метионин (Met), фенилаланин (Phe), пролин (Pro), серии (Ser), треонин (Thr), триптофан (Trp), тирозин (Tyr) и валин (Val). Другие встречающиеся в природных источниках аминокислоты включают, например, аминокислоты, которые синтезируются в процессах метаболизма, не связанных с синтезом белков. Например, аминокислоты орнитин и цитруллин синтезируются в метаболических процессах организма животного при продуцировании мочевины. Пептиды, используемые в настоящем изобретении, могут содержать одну или несколько аминокислот неприродного происхождения. Аминокислоты неприродного происхождения могут быть L-, декстровращающими (D) или их смесями. Оптимально пептид не содержит аминокислот природного происхождения. Аминокислоты неприродного происхождения – это такие аминокислоты, которые не синтезируются в нормально протекающих в живом организме метаболических процессах и потому не встречаются в белках в природе. Кроме того, не встречающиеся в природных источниках аминокислоты, используемые в настоящем изобретении, предпочтительно не распознаются обычными протеазами. Аминокислоты неприродного происхождения могут содержаться в любом положении пептида. Например, аминокислота неприродного происхождения может быть на N-конце, С-конце или в любом положении между N-концом и С-концом. Аминокислоты неприродного происхождения могут, например, включать алкильные, арильные или алкиларильные группы. Примеры алкилсодержащих аминокислот включают Аминокислоты неприродного происхождения могут включать производные аминокислот природного происхождения. Производные аминокислот природного происхождения могут, например, включать присоединение одной или нескольких химических групп к аминокислоте природного происхождения. Например, одна или несколько химических групп могут быть присоединены в одном или нескольких 2′, 3′, 4′, 5′ или 6′ положениях ароматического кольца остатка фенилаланина или тирозина или в 4′, 5′, 6′ или 7′ положении бензольного кольца остатка триптофана. Такая группа может быть любой химической группой, которая может быть введена в ароматическое кольцо. Примеры таких групп включают разветвленные или неразветвленные алкилы, такие как метил, этил, н-пропил, изопропил, бутил, изобутил, или т-бутил, С1-С4 алкилокси (например, алкокси), амино, C1-C4 алкиламино и C1-C4 диалкиламино (например, метиламино, диметиламино), нитро, гидроксил, гало (например, фтор, хлор, бром или иод). Конкретными примерами не встречающихся в природе производных аминокислот природного происхождения являются норвалин (Nva), норлейцин (Nle) и гидроксипролин (Hyp). Другим примером модификации аминокислоты в пептиде, используемом в способах настоящего изобретения, является дериватизация карбоксильной группы остатка аспарагиновой кислоты или глутаминовой кислоты пептида. Примером дериватизации является амидирование аммиаком или первичным или вторичным амином, например, метиламином, этиламином, диметиламином или диэтиламином. Другим примером дериватизации является этерификация, например, метиловым или этиловым спиртом. Другая модификация включает дериватизацию аминогруппы лизина, аргинина или гистидина. Например, такие аминогруппы могут быть ацилированы. Приемлемые ацильные группы включают, например, бензоильную группу или алканоильную группу, включая какие-либо из C1-C4 алкильных групп, упомянутых выше, например, ацетильную или пропионильную группу. Аминокислоты неприродного происхождения предпочтительно устойчивы и более предпочтительно – нечувствительны по отношению к общеизвестным протеазам. Примеры аминокислот неприродного происхождения, которые являются устойчивыми или нечувствительными к протеазам, включают декстровращающую (D-) форму любой из вышеупомянутых природных L-аминокислот, а также L- и/или D-неприродные аминокислоты. D-аминокислоты обычно не встречаются в белках, хотя они обнаружены в некоторых пептидных антибиотиках, которые синтезируются другим, а не обычным путем рибосомального синтеза белков в клетке. В используемом здесь смысле считают, что D-аминокислоты являются аминокислотами неприродного происхождения. Для того чтобы была минимизирована чувствительность к протеазам, пептиды, используемые в способах настоящего изобретения, должны иметь менее пяти, предпочтительно менее четырех, более предпочтительно менее трех и наиболее предпочтительно – менее двух смежных L-аминокислот, распознаваемых обычными протеазами, независимо от того, относятся они к аминокислотам природного или неприродного происхождения. Благоприятно, когда пептид содержит только D-аминокислоты, а не L-аминокислоты. Если пептид содержит чувствительные к протеазам последовательности аминокислот, по крайней мере, одна из аминокислот предпочтительно является D-аминокислотой неприродного происхождения, что обеспечивает в результате устойчивость к протеазам. Пример протеазно-чувствительной последовательности включает две или более смежных основных аминокислоты, которые легко расщепляются известными протеазами, такими как эндопептидазы и трипсин. Примеры основных аминокислот включают аргинин, лизин и гистидин. Является важным, что, по меньшей мере, одна из аминокислот, содержащихся в ароматическом катионном пептиде, является остатком тирозина или триптофана или их производным. Важным является также и то, что ароматические катионные пептиды имеют минимальное количество результирующих положительных зарядов при физиологическом значении pH по сравнению с общим числом аминокислотных остатков в пептиде. Минимальное количество положительных зарядов при физиологических pH будет обозначаться далее как (pm). Общее количество аминокислотных остатков в пептиде будет обозначаться далее как (r). Минимальное число результирующих положительных зарядов, обсуждаемое далее, соответствует физиологическому pH. Термин «физиологический pH» в используемом здесь смысле относится к нормальному pH в клетках ткани или органов организма млекопитающего. Например, физиологический pH для человека обычно составляет приблизительно 7,4, а нормальное значение физиологического pH у млекопитающего может быть любым значением pH в диапазоне от около 7,0 до около 7,8. «Результирующий заряд» в используемом здесь смысле означает баланс между числом положительных зарядов и числом отрицательных зарядов, поддерживаемый аминокислотами, содержащимися в пептиде. Понятно, что в данном описании результирующий заряд определяют при физиологическом pH. Аминокислоты природного происхождения, которые имеют положительный заряд при физиологическом pH, включают L-лизин, L-аргинин и L-гистидин. Аминокислоты природного происхождения, которые имеют отрицательный заряд при физиологическим pH, включают L-аспарагиновую кислоту и L-глутаминовую кислоту. Как правило, пептид имеет положительно заряженную аминокислоту на N-конце и отрицательно заряженную карбоксильную группу на С-конце. Эти заряды взаимно погашают друг друга при физиологическом pH. Пример подсчета результирующего заряда: в пептиде Tyr-Arg-Phe-Lys-Glu-His-Trp-Arg имеется одна отрицательно заряженная аминокислота (например, Glu) и четыре положительно заряженных аминокислоты (например, два Arg остатка, один Lys и один His). Следовательно, вышеобозначенный пептид имеет общий (результирующий) положительный заряд, равный трем. В одном варианте настоящего изобретения ароматические катионные пептиды имеют соотношение между минимальным количеством результирующих положительных зарядов при физиологическом pH (pm) и общим числом аминокислотных остатков (r), где 3pm является наибольшим числом, которое меньше или равно r+1. В этом варианте соотношение между минимальным количеством результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r) является следующим:
В другом варианте ароматические катионные пептиды имеют соотношение между минимальным количеством результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r), где 2pm представляет собой наибольшее число, которое меньше или равно r+1. В этом варианте соотношение между минимальным количеством результирующих положительных зарядов (pm) и общим числом аминокислотных остатков (r) является следующим:
В одном варианте минимальное количество результирующих положительных зарядов (pm) и общее число аминокислотных остатков (r) равны друг другу. В другом варианте пептиды имеют три или четыре аминокислотных остатка и минимум один результирующий положительный заряд, предпочтительно минимум два результирующих положительных заряда и более предпочтительно минимум три результирующих положительных заряда. Важно также то, что ароматические катионные пептиды имеют минимальное количество ароматических групп по сравнению с общим количеством результирующих положительных зарядов (pt). Минимальное количество ароматических групп будет обозначаться далее как (а). Аминокислоты природного происхождения, которое имеют ароматическую группу, включают гистидин, триптофан, тирозин и фенилаланин. Например, гексапептид Lys-Gln-Tyr-Arg-Phe-Trp имеет результирующий положительный заряд, равный двум (за счет остатков лизина и аргинина), и три ароматические группы (за счет остатков тирозина, фенилаланина и триптофана). В одном варианте изобретения ароматические катионные пептиды, используемые в способах настоящего изобретения, имеют соотношение между минимальным числом ароматическим групп (а) и общим количеством результирующих положительных зарядов при физиологическом pH (pt), где 3а представляет собой наибольшее число, которое меньше или равно pt+1, за исключением того, что когда pt равно 1, а может быть также равно 1. В этом варианте соотношение между минимальным числом ароматических групп (а) и общим количеством результирующих положительных зарядов (pt) является следующим:
В другом варианте ароматические катионные пептиды имеют соотношение между минимальным числом ароматических групп (а) и общим количеством результирующих положительных зарядов (pt), где 2а представляет собой наибольшее число, которое меньше или равно pt+1. В этом варианте соотношение между минимальным числом ароматических аминокислотных остатков (а) и общим количеством результирующих положительных зарядов (pt) является следующим:
В другом варианте количество ароматических групп (а) и общее количество результирующих положительных зарядов (pt) равны. Карбоксильные группы, особенно концевые карбоксильные группы С-концевой аминокислоты, предпочтительно амидированы, например, аммиаком с образованием С-концевого амида. Альтернативно концевая карбоксильная группа С-концевой аминокислоты может быть амидирована первичным или вторичным амином. Первичный или вторичный амин может, например, быть алкил-, в частности разветвленный или неразветвленный C1-C4 алкил, или ариламином. Соответственно аминокислота на С-конце пептида может быть превращена в амидо, N-метиламидо, N-этиламидо, N,N-диметиламидо, N,N-диэтиламидо, N-метил-N-этиламидо, N-фениламидо или N-фенил-N-этиламид. Свободные карбоксильные группы остатков аспарагина, глутамина, аспарагиновой кислоты и глутаминовой кислоты, не встречающиеся на С-концах ароматических катионных пептидов настоящего изобретения, также могут быть амидированы независимо от их местонахождения внутри структуры пептида. Амидирование этих внутренних положений может быть осуществлено с помощью аммиака или первичных и вторичных аминов, описанных выше. В одном варианте ароматический катионный пептид, используемый в способах настоящего изобретения, является трипептидом, имеющим два результирующих положительных заряда и, по меньшей мере, одну ароматическую аминогруппу. В отдельном варианте ароматический катионный пептид, используемый в способах настоящего изобретения, является трипептидом, имеющим два результирующих положительных заряда и две ароматические аминокислоты. Ароматические катионные пептиды, используемые в способах настоящего изобретения, включают (но без ограничения) следующие примеры пептидов: Lys-D-Arg-Tyr-NH2, D-Tyr-Trp-Lys-NH2, Trp-D-Lys-Tyr-Arg-NH2, Tyr-His-D-Gly-Met, Tyr-D-Arg-Phe-Lys-Glu-NH2, Met-Tyr-D-Lys-Phe-Arg, D-His-Glu-Lys-Tyr-D-Phe-Arg, Lys-D-Gln-Tyr-Arg-D-Phe-Trp-NH2, Phe-D-Arg-Lys-Trp-Tyr-D-Arg-His, Gly-D-Phe-Lys-Tyr-His-D-Arg-Tyr-NH2, Val-D-Lys-His-Tyr-D-Phe-Ser-Tyr-Arg-NH2, Trp-Lys-Phe-D-Asp-Arg-Tyr-D-His-Lys, Lys-Trp-D-Tyr-Arg-Asn-Phe-Tyr-D-His-NH2, Thr-Gly-Tyr-Arg-D-His-Phe-Trp-D-His-Lys, Asp-D-Trp-Lys-Tyr-D-His-Phe-Arg-D-Gly-Lys-NH2, D-His-Lys-Tyr-D-Phe-Glu-D-Asp-D-His-D-Lys-Arg-Trp-NH2, Ala-D-Phe-D-Arg-Tyr-Lys-D-Trp-His-D-Tyr-Gly-Phe, Tyr-D-His-Phe-D-Arg-Asp-Lys-D-Arg-His-Trp-D-His-Phe, Phe-Phe-D-Tyr-Arg-Glu-Asp-D-Lys-Arg-D-Arg-His-Phe-NH2, Phe-Tyr-Lys-D-Arg-Trp-His-D-Lys-D-Lys-Glu-Arg-D-Tyr-Thr, Tyr-Asp-D-Lys-Tyr-Phe-D-Lys-D-Arg-Phe-Pro-D-Tyr-His-Lys, Glu-Arg-D-Lys-Tyr-D-Val-Phe-D-His-Trp-Arg-D-Gly-Tyr-Arg-D-Met-NH2, Arg-D-Leu-D-Tyr-D-Phe-Lys-Glu-D-Lys-Arg-D-Trp-Lys-D-Phe-Tyr-D-Arg-Gly, D-Clu-Asp-Lys-D-Arg-D-His-Phe-Phe-D-Val-Tyr-Arg-Tyr-D-Tyr-Arg-His-Phe-NH2, Asp-Arg-D-Phe-Cys-Phe-D-Arg-D-Lys-Tyr-Arg-D-Tyr-Trp-D-His-Tyr-D-Phe-Lys-Phe, His-Tyr-D-Arg-Trp-Lys-Phe-D-Asp-Ala-Arg-Cys-D-Tyr-His-Phe-D-Lys-Tyr-His-Ser-NH2, Gly-Ala-Lys-Phe-D-Lys-Glu-Arg-Tyr-His-D-Arg-D-Arg-Asp-Tyr-Trp-D-His-Trp-His-D-Lys-Asp, Thr-Tyr-Arg-D-Lys-Trp-Tyr-Glu-Asp-D-Lys-D-Arg-His-Phe-D-Tyr-Gly-Val-Ile-D-His-Arg-Tyr-Lys-NH2. В одном варианте пептиды, используемые в способах настоящего изобретения, обладают активностью агониста mu-опиоидного рецептора (например, активируют mu-опиоидный рецептор). Активация mu-опиоидного рецептора обычно повышает обезболивающий эффект. В некоторых случаях ароматический катионный пептид, обладающий активностью mu-опиоидного рецептора, является предпочтительным. Например, в процессе кратковременного лечения при остром заболевании или внезапно возникшем состоянии использование ароматического катионного пептида, который активирует mu-опиоидный рецептор, может оказаться благоприятным. Например, острые заболевания и состояния могут сопровождаться умеренной или сильной болью. В этих случаях анальгетический эффект ароматического катионного пептида может быть с успехом использован при лечении пациента-человека или млекопитающего, хотя ароматический катионный пептид, который не активирует mu-опиоидный рецептор, также может использоваться (вместе с анальгетиком или без него) по клинической необходимости. Альтернативно в других случаях ароматический катионный пептид, который не обладает активностью mu-опиоидного рецептора, является предпочтительным. Например, при длительном лечении хронического заболевания или состояния использование ароматического катионного пептида, который активирует mu-опиоидный рецептор, может быть противопоказано. В этих случаях потенциально вредные или аддиктивные эффекты ароматического катионного пептида могут мешать использованию ароматического катионного пептида, который активирует mu-опиоидный рецептор при лечении человека или другого млекопитающего. Потенциально вредные, побочные эффекты могут включать седативный эффект, запор, угнетение нервной системы или угнетение дыхания. В таких случаях ароматический катионный пептид, который не активирует mu-опиоидный рецептор, может быть подходящим лечебным средством. Примеры острых состояний включают сердечный приступ, удар и травму. Травматическое поражение может включать травму головного мозга и травму позвоночника. Примеры хронических заболеваний или состояний включают заболевание коронарной артерии и нейродегенеративные заболевания, например, те, которые описаны ниже. Пептиды, используемые в способах настоящего изобретения, которые имеют активность mu-опиоидного рецептора, обычно представляют собой пептиды, имеющие на N-конце остатки тирозина или производного тирозина (положение первой аминокислоты). Предпочтительные производные тирозина включают 2′-метилтирозин (Mmt); 2′,6′-диметилтирозин (2’6’Dmt); 3′,5′-диметилтирозин (3’5’Dmt); N, 2′,6′-триметилтирозин (Tmt) и 2′-гидрокси-6′-метилтирозин (Hmt). В конкретном предпочтительном варианте пептид, который обладает активностью mu-опиоидного рецептора, имеет формулу Tyr-D-Arg-Phe-Lys-NH2 (для удобства представленную как DALDA, что упоминается здесь как SS-01). DALDA имеет результирующий положительный заряд, равный трем, за счет аминокислот тирозина, аргинина и лизина, и имеет две ароматические группы за счет аминокислот фенилаланина и тирозина. Тирозин DALDA может представлять собой модифицированное производное тирозина, например, 2’6′-диметилтирозин, и тогда соединение имеет формулу 2’6′-Dmt-D-Arg-Phe-Lys-NH2 (например, Dmt1-DALDA, который обозначен здесь как SS-02). Пептиды, которые не обладают активностью mu-опиоидного рецептора, вообще не имеют остатка тирозина или производного тирозина на N-конце (т.е. в положении первой аминокислоты). Аминокислота на N-конце может быть кислотой природного происхождения или неприродного происхождения, отличной от тирозина. В одном варианте аминокислотой на N-конце является фенилаланин или его производное. Предпочтительными производными фенилаланина являются 2′-метилфенилаланин (Mmp), 2′,6′-диметилфенилаланин (Dmp), N,2′,6′-триметилфенилаланин (Tmp), 2′-гидрокси-6′-метилфенилаланин (Hmp). В другом предпочтительном варианте аминокислотным остатком на N-конце является аргинин. Примером такого пептида является D-Arg-2’6′-Dmt-Lys-Phe-NH2 (обозначаемый здесь как SS-31). Другой ароматический катионный пептид, который не обладает активностью mu-опиоидного рецептора, имеет формулу Phe-D-Arg-Dmt-Lys-NH2. Альтернативно, N-концевой фенилаланин может быть производным фенилаланина, например, 2′,6′-диметилфенилаланином (2’6′-Dmt). DALDA, содержащий 2′,6′-диметилфенилаланин в аминокислотном положении 1, имеет формулу 2′,6′-Dmp-D-Arg-Dmt-Lys-NH2. В предпочтительном варианте аминокислотная последовательность Dmt1-DALDA (SS-02) перестроена таким образом, что Dmt не находится в N-концевом положении. Пример такого ароматического катионного пептида, который не обладает активностью mu-опиоидного рецептора, соответствует формуле D-Arg-2’6’Dmt-Lys-Phe-NH2 (SS-31). DALDA, SS-31 и их производные могут также включать функциональные аналоги. Пептид расценивается как функциональный аналог DALDA или SS-31, если аналог имеет ту же функцию, что и DALDA или SS-31. Аналог может, например, быть вариантом замещения DALDA или SS-31, когда одна или несколько аминокислот замещены другой аминокислотой. Подходящие варианты замен в DALDA или SS-31 включают замены консервативных аминокислот. Аминокислоты могут быть сгруппированы в соответствии с их физико-химическими характеристиками следующим образом: (a) Неполярные аминокислоты: Ala(A)Ser(S)Thr(T)Pro(P)Gly(G) (b) Кислые аминокислоты: Asn(A)Asp(D)Glu(E)Gln(Q); (c) Основные аминокислоты: His(H)Arg(R)Lys(K); (d) Гидрофобные аминокислоты: Met(M)Leu(L)Ile(I)Val(V); (e) Ароматические аминокислоты: Phe(F)Tyr(Y)Trp(W)His(H). Замещения (замены) аминокислоты в пептиде другой аминокислотой из той же самой группы расценивается как консервативная замена и может сохранять физико-химические характеристики исходного пептида. Аминокислотные замены в пептиде другой аминокислотой из иной группы, по всей видимости, приводят к изменению характеристик исходного пептида. Примеры аналогов, используемых в настоящем изобретении, которые активируют mu-опиоидные рецепторы, включают (без ограничения) ароматические катионные пептиды, показанные в таблице 1.
Примеры аналогов, используемых в настоящем изобретении, которые не активируют mu-опиоидные рецепторы, включают (без ограничения) ароматические катионные пептиды, приведенные в Таблице 2.
Аминокислоты пептидов, приведенных в таблицах 1 и 2, могут быть как в L-конфигурации, так и в D-конфигурации. Способы уменьшения окислительного повреждения Вышеописанные пептиды могут быть использованы для уменьшения окислительного повреждения у млекопитающего, которое в этом нуждается. Млекопитающие, нуждающиеся в уменьшении окислительного повреждения, это млекопитающие, страдающие от заболевания, состояния или подвергнутые лечению, когда они связаны с окислительным повреждением. Как правило, окислительное повреждение вызывается свободными радикалами, такими как разновидности реакционноспособного кислорода (ROS) и/или разновидности реакционноспособного азота (RNS). Примеры ROS или RNS включают гидроксильный радикал (НО В одном варианте млекопитающее, нуждающееся в этом, может представлять собой млекопитающее, подвергшееся лечению, связанному с окислительным повреждением. Например, млекопитающее может быть подвергнуто реперфузии. Реперфузия означает восстановление кровотока к любому органу или ткани, в которых кровоток снижен или блокирован. Восстановление кровотока в процессе реперфузии дает дыхательный импульс и приводит к образованию свободных радикалов. Сниженный или заблокированный кровоток может быть следствием гипоксии или ишемии. Потеря или резкое снижение обеспечения кровью во время гипоксии или ишемии может происходить вследствие тромбоэмболического удара, коронарного атеросклероза или заболевания периферических сосудов. Множество органов и тканей могут подвергнуться ишемии или гипоксии. Примеры таких органов включают головной мозг, сердце, почки, кишечник и простату. Ткань, на которую оказывается такое воздействие, обычно является мышечной, например, сердечная мышца, скелетные мышцы или гладкие мышцы. Например, ишемия или гипоксия сердечной мышцы часто бывает вызвана атеросклеротическими или тромботическими блокадами, которые приводят к уменьшению или прекращению доступа кислорода в ткани сердца за счет артериального и капиллярного кровоснабжения сердца. Такие явления ишемии и гипоксии сердца могут вызывать боль и некроз задействованных мышц сердца и в конечном счете приводить к сердечной недостаточности. Ишемия или гипоксия скелетной мышцы или гладкой мышцы могут возникать по тем же самым причинам. Например, ишемия или гипоксия в кишечной гладкой мышце или скелетной мышце конечности могут также быть обусловлены атеросклерозом или тромбозом. Восстановление кровотока (реперфузия) может осуществляться любым способом, известным специалистам в данной области. Например, реперфузия ишемических тканей сердца может быть обусловлена ангиопластикой, шунтированием коронарной артерии или при использовании тромболитических лекарственных средств. Снижение окислительного повреждения, связываемое с ишемией/гипоксией, и реперфузия являются важными, потому что повреждение тканей, связанное с ишемией, гипоксией, обуславливают, например, инфаркт миокарда, удар и геморрагический шок. В другом варианте нуждающееся в уменьшении окислительного повреждения млекопитающее может быть млекопитающим с заболеванием или состоянием, связанным с окислительным повреждением. Окислительное повреждение может произойти в любой клетке, ткани или органе млекопитающего. Примеры клеток, тканей или органов включают, но не ограничиваются, эндотелиальные клетки, клетки нервной системы, кожу, сердце, легкие, почки и печень. Например, процессы пероксидирования липидов и воспаления связывают с окислительным повреждением вследствие болезни или состояния. Пероксидирование липидов означает окислительную модификацию липидов. Липиды могут содержаться в мембране клетки. Эта модификация мембранных липидов обычно приводит к изменению и/или повреждению функции мембраны клетки. Кроме того, пероксидирование липидов может также происходить и с липидами или липопротеинами, экзогенными для клетки. Например, липопротеины низкой плотности весьма чувствительны к липидному пероксидированию. Примером состояния, связанного с пероксидированием липидов, является атеросклероз. Снижение степени окислительного повреждения, связанное с атеросклерозом, является важным, так как атеросклероз имеет отношение, например, к сердечным приступам и заболевания и коронарной артерии, Воспалительный процесс означает активацию иммунной системы. Обычно иммунная система активируется антигенным веществом. Антигенное вещество может быть любым веществом, распознаваемым иммунной системой, и включает самовоспроизводящиеся частицы и частицы, создающиеся из чужеродного вещества. Примеры заболеваний или состояний, начинающихся с воспалительного процесса до самовоспроизводящихся частиц, включают артрит и рассеянный склероз. Примеры чужеродных частиц включают вирусы и бактерии. Вирус может быть любым вирусом, который активирует воспалительный процесс и связан с окислительным повреждением. Примеры вирусов включают вирусы гепатита А, В или С, вирус иммунодефицита человека, вирус гриппа и вирус диареи быка. Например, вирус гепатита может вызывать воспалительный процесс и образование свободных радикалов, повреждая тем самым печень. Бактерии могут быть любыми бактериями и включать как грамотрицательные, так и грамположительные бактерии. Грамотрицательные бактерии содержат липополисахарид в бактериальной стенке. Примеры грамотрицательных бактерий включают: Escherichia coli, Klebsiella pneumonia, Proteus species, Pseudomonas aeroginosa, Serratia и Bacteroides. Пример грамположительных бактерий включают пневмококки и стрептококки. Примером воспалительного процесса, связанного с окислительным стрессом, вызываемым бактериями, является сепсис. Обычно сепсис вызывается грамотрицательными бактериями, попавшими в кровоток. Повреждение печени, вызванное токсичным агентом, представляет собой другое состояние, связанное с воспалительным процессом и окислительным стрессом. Токсический агент может быть любым агентом, который вызывает повреждение печени. Например, токсичный агент может вызвать апоптоз и/или некроз клеток печени. Примерами таких агентов являются спирт, медикаменты, например, предписанные и непредписанные лекарственные средства (наркотики), взятые для лечения заболевания или состояния. Способы настоящего изобретения могут также использоваться для уменьшения окислительного повреждения, связанного с нейродегенеративным заболеванием или состоянием. Нейрогенеративное заболевание может повлиять на любую клетку, ткань, орган центральной и периферической нервной системы. Примерами таких клеток, тканей и органов являются головной мозг, спинной мозг, нейтроны, ганглии, Schwann клетки, астроциты, олигодендроциты и микроглия. Нейродегенеративное состояние может быть острым состоянием, например, как удар или травматическое поражение головного или спинного мозга. В другом варианте нейродегенеративное заболевание или состояние может быть хроническим нейродегенеративным состоянием. При хроническом нейродегенеративном состоянии свободные радикалы могут, например, вызвать повреждение белка. Примером такого белка является амилоидный Другие состояния, которые можно лечить в соответствии с настоящим изобретением, включают преэклампсию, диабет, симптомы и состояния, связанные со старением, например, дегенерацию желтого пятна, морщины. В другом варианте пептиды, используемые в настоящем изобретении, могут также использоваться для уменьшения окислительного повреждения в органе млекопитающего перед трансплантацией. Например, удаленный орган, если он подвергается реперфузии после трансплантации, может оказаться чувствительным к окислительному повреждению. Следовательно, пептиды могут использоваться для уменьшения окислительного повреждения от реперфузии трансплантированного органа. Удаленный орган может быть любым органом, пригодным для трансплантации. Примеры таких органов включают сердце, печень, почки, легкое и панкреатические островки. Удаленный орган помещается в подходящую среду, например, стандартный буферный раствор, обычно используемый в данной области. Например, удаленное сердце может быть помещено в кардиоплегический раствор, содержащий пептиды, описанные выше. Концентрацию пептидов в стандартном буферном растворе легко может определить специалист в данной области. Такие концентрации могут быть, например, между около 0,01 нМ до около 10 µМ, предпочтительно от около 0,1 нМ до около 10 µM, более предпочтительно от около 1 нМ до около 5 µМ и еще более предпочтительно от около 1 нМ до около 100 нМ. В другом варианте изобретение предусматривает способ уменьшения окислительного повреждения в клетке, которая нуждается в этом. Клетки, которым требуется уменьшение окислительного повреждения, это, в основном, такие клетки, в которых клеточные мембраны или ДНК клетки повреждены свободными радикалами, например, ROS и/или RNS. Примеры клеток, которые могут подвергаться окислительному повреждению, включают описываемые здесь клетки. Подходящие примеры клеток включают панкреатические островковые клетки, миоциты, эндотелиальные клетки, нейронные клетки, стволовые клетки и т.п. Эти клетки могут быть клетками культуры ткани. Альтернативно клетки могут быть получены от млекопитающего. В одном случае клетки могут быть повреждены из-за окислительного эффекта как результат инсульта. Такие инсульты, включают, например, заболевание или состояние (например, диабет или др.) или ультрафиолетовое облучение (например, солнце и т.д.). Например, панкреатические островковые клетки, повреждаемые при окислении как результат диабета, могут быть получены от млекопитающего. Пептиды, описываемые здесь, могут быть введены в клетку любым способом, известным специалистам в данной области. Например, пептиды можно инкубировать вместе с клетками в подходящих условиях. Такие условия может легко определить специалист в данной области. Вследствие уменьшения окислительного повреждения обработанные клетки могут оказаться способными к регенерации. Такие регенерированные клетки могут быть введены обратно млекопитающему как терапевтическое лечение заболевания или состояния. Как указано выше, одним таким состоянием является диабет. Полагают, что окислительное повреждение «уменьшается», если степень, мера окислительного повреждения у млекопитающего, в удаленном органе или клетке снижается после введения эффективного количества ароматических катионных пептидов, описанных выше. Полагают обычно, что окислительное повреждение уменьшается, если окислительное повреждение снижается, по меньшей мере, примерно на 10%, предпочтительно, по меньшей мере, примерно на 25%, более предпочтительно, по меньшей мере, примерно на 50%, еще более предпочтительно, по меньшей мере, примерно на 75% и самое предпочтительное, по меньшей мере, примерно на 90%. Синтез пептидов Пептиды, используемые в способах настоящего изобретения, могут быть синтезированы химическим путем с помощью любого из хорошо известных способов. Подходящие методы для синтезирования белков включают, например, такие, которые описаны Stuart and Young в «Solid Phase Peptide Synthesis», Second Edition, Pierce Chemical Company (1984) и в «Solid Phaze Peptide Synthesis», Methods EnzymoL, 289, Academic Press, Inc., Ne York (1997). Способы введения Пептид, который можно использовать в способах настоящего изобретения, вводят млекопитающему в количестве, эффективном для уменьшения окислительного повреждения. Эффективное количество определяют в процессе предклинических испытаний и клинических испытаний методами, знакомыми врачам и клиницистам. Эффективное количество пептида, используемого в способах настоящего изобретения, предпочтительно в виде фармацевтической композиции, может вводиться млекопитающему, нуждающемуся в этом, любым из многочисленных широко распространенных способов для введения фармацевтических композиций. Пептид может вводиться системно или локально. В одном варианте пептид вводят внутривенно. Например, ароматические катионные пептиды, которые могут использоваться в способах настоящего изобретения, могут вводиться с помощью быстрой внутривенной болюсной инъекции. Предпочтительно, однако, пептид вводят как внутривенную инфузию при постоянной скорости. Пептид можно впрыскивать непосредственно в коронарную артерию во время, например, ангиопластики или операции по коронарному шунтированию, или применять на коронарные стенты. Пептид может также вводиться отдельно, местно, интраназально, внутримышечно, подкожно или трансдермально. В предпочтительном варианте трансдермальное введение ароматических катионных пептидов способами настоящего изобретения осуществляют путем ионофореза, при котором заряженный пептид доставляется через кожу, обуславливая электрический ток. Другие методы введения включают интрацеребровентрикулярный или интратекальный методы. Интрацеребровентрикулярно означает введение через вентрикулярную систему головного мозга. Интратекально означает введение в пространство под арахноидной мембраной спинного мозга. Таким образом интрацеребровентрикулярное или интратекальное введение может быть предпочтительным при тех заболеваниях и состояниях, в которых задействованы органы или ткани центральной нервной системы. В предпочтительном варианте интратекальное введение используется при травмах спинного мозга. Пептиды, пригодные для использования в способах данного изобретения, можно также вводить млекопитающим путем непрерывной доставки (выделения), который хорошо известен в данной области. Введение методами непрерывной доставки представляет собой такой метод доставки лекарственного средства, который обеспечивает достижение (или поддержание) определенной концентрации лекарства в течение определенного временного периода. Концентрации определяют в сыворотке или плазме. Описание способов доставки соединения при условии контролируемого выделения может быть найдено в заявке РСТ Любой состав, известный в данной области фармацевтики, пригоден для введения ароматических катионных пептидов, которые можно использовать в способах настоящего изобретения. Для орального способа введения могут применяться жидкие или твердые составы. Примеры составов включают таблетки, желатиновые капсулы, пилюли, пастилки, эликсиры, суспензии, сиропы, пастилки для жевания и т.п. Пептиды могут быть смешаны с подходящим фармацевтическим носителем (наполнителем) или разбавителем, что понятно специалистам в данной области. Примеры носителей и наполнителей включают крахмал, молоко, сахар, некоторые виды клея, желатин, молочную кислоту, стеариновую кислоту или их соли, в том числе стеараты магния или кальция, тальк, растительные масла, смолы и гликоли. Для системного, интрацеребровентрикулярного, интратекального, местного, интраназального, подкожного или трансдермального введения в композициях ароматических катионных пептидов, используемых в способах настоящего изобретения, могут применяться обычные разбавители, носители или наполнители и т.п., которые хорошо известны в данной области как средства доставки пептидов. Например, такие составы могут включать одно или несколько следующих компонентов: стабилизатор, сурфактант (ПАВ), предпочтительно неионный ПАВ и необязательно соль и/или забуферивающий агент. Пептиды могут подаваться в виде водных растворов или в лиофилизированной форме. Стабилизатор может быть, например, аминокислотой, такой, к примеру, как глицин, или олигосахаридом, например, сахарозой, тетралозой, лактозой или декстраном. Альтернативно стабилизатор может представлять собой сахарный спирт, например маннитол; или их сочетание. Предпочтительно стабилизатор или сочетание стабилизаторов составляет от около 0,1% до около 10% вес. от веса пептида. Поверхностно-активное вещество (ПАВ) предпочтительно является неионным ПАВ, таким как полисорбат. Некоторые примеры подходящих сурфактантов включают Твин 20, Твин 80; полиэтиленгликоль или полиоксиэтилен полиоксипропиленгликоль, такой как Pluronic F-68 при концентрации от около 0,001% (вес/об.) до около 10% (вес/об.). Соль или забуферивающий агент могут быть любой солью или буферным агентом, например, хлоридом натрия и фосфатом натрия/калия соответственно. Предпочтительно буферный агент поддерживает pH фармацевтической композиции в пределах от около 5,5 до около 7,5. Соль и/или буферный агент используются также для поддержания осмолярности на уровне, приемлемом для введения человеку или животному. Предпочтительно соль или буферный агент содержатся в примерно изотонической концентрации от около 150 мМ до около 300 мМ. Композиции пептидов, которые могут использоваться в способах настоящего изобретения, могут также содержать одну или несколько обычных добавок. Примеры таких добавок включают солюбилизатор, например, глицерин, антиоксидант, например бензалконий-хлорид (смесь четвертичных производных аммония, известная как «quats», бензиловый спирт, хлоретон или хлорбутанол; анестезирующее средство, например, производное морфина, или изотонический агент и т.д., например из тех, что перечислены выше. Как дополнительная предосторожность в плане окисления или иной порчи, фармацевтические композиции могут храниться под азотом в запаянных сосудах с непроницаемой укупоркой. Млекопитающее, которое можно лечить согласно изобретению, может быть любым млекопитающим, например, это могут быть сельскохозяйственные животные, то есть свиньи, овцы, коровы, лошади, домашние животные – кошки, собаки, лабораторные животные – крысы, мыши, кролики. В предпочтительном варианте млекопитающее – это человек. Примеры Пример 1. [Dmt1]DALDA проникает через клеточную мембрану. Поглощение клетками [3H][Dmt1]DALDA изучали с использованием линии интестинальных эпителиальных клеток человека (Сасо-2) и анализировали с применением SH-SY5Y (клетки нейробластомы человека), НЕК 293 (клетки почки эмбриона человека) и CRFK клеток (клетки почечного эпителия). Монослои клеток выращивали на 12-луночных планшетах (5×105 клеток/ячейку) с коллагеновым покрытием в течение 3 дней. На 4-ый день клетки промывали дважды подогретым HBSS и затем инкубировали с 0,2 мл HBSS, содержащим 250 нм [3H][Dmt1]DALDA при 37°С или 4°С в течение разных промежутков времени в пределах 1 часа. [3H][Dmt1] стало возможным наблюдать в клеточном лизате уже спустя 5 мин, а стабильный уровень был достигнут через 30 мин. Общее количество [H][Dmt1]DALDA, выделенное в клеточный лизат после 1 часа инкубации, достигло почти 1% общего количества лекарства. Поглощение [3H][Dmt1]DALDA при 4°С было более медленным, чем при 37°С, но достигало 76,5% через 45 мин и 86,3% спустя 1 час. Интернализация [3H][Dmt1]DALDA не была ограничена Сасо-2 клетками, но наблюдалась также и в SH-SY5Y, НЕК-293 и CRFK клетках. Определили, что внутриклеточная концентрация [Dmt1]DALDA была примерно в 50 раз выше, чем внеклеточная концентрация. В другом эксперименте клетки инкубировали в диапазоне концентраций [Dmt1]DALDA (1 µM – 3 µМ) в течение 1 часа при 37°С. В конце инкубационного периода клетки промывали 4 раза HBSS и 0,2 мл 0,1 н NaOH с 1% SDS добавляли в каждую ячейку. Содержимое с клетками затем переносили в сцинтиляционные сосуды и подсчитывали радиоактивность. Чтобы разграничить радиоактивность от интернализации от связанной с поверхностью радиоактивности, была включена стадия отмывки кислотой. Перед лизисом клеток их инкубировали с 0,2 мл 0,2 М уксусной кислоты и 0,05 М NaCl в течение 5 мин на льду. Поглощение [Dmt1]DALDA Сасо-2 клетками оценивали конфокальной лазерной сканирующей микроскопией (CLSM) с использованием флуоресцентного анализа [Dmt1]DALDA (Dmt-D-Agr-Phe-dnsDap-NH2; где dnsDap= CLSM подтверждал поглощение флуоресцентного Dmt-D-Arg-Phe-dnsDap-NH2 в Сасо-2 клетках после инкубации с 0,1 µM [Dmt1, DnsDap4]DALDA в течение 1 часа при 37°С. Поглощение флуоресцентного пептида при 37°С и 4°С было сходным. По-видимому, флуоресценция проникает через цитоплазму, но исключается ядрами. Пример 2. Нацеливание [Dmt1]DALDA в митохондрии Чтобы исследовать межклеточное распределение [Dmt1]DALDA, был получен флуоресцентный аналог [Dmt1, AtnDap4]DALDA (Dmt-D-Arg-Phe-atnDap-NH2; где atn= Клетки также инкубировали с тетраметилродаминметиловым эфиром (TMRM, 25 нМ), красителем для окрашивания митохондрии, в течение 15 мин при 37°С. Клетки затем промывали три раза ледяным HBSS и наслаивали 200 µл PBS, осуществляли микроскопирование в течение 10 мин, применяя Nikon конфокальный лазерный сканирующий микроскоп с С-Апохроматным 63×/1,2 W объективом. Для [Dmt1, AtnDap4]DALDA возбуждение осуществляли при 350 нм с помощью УФ-лазера и измеряли эмиссию при 520 нм. Для TMRM возбуждение осуществляли при 536 нм и эмиссию определяли при 560 нм. CLSM показал поглощение флуоресцентного [Dmt1, AtnDap4]DALDA в Сасо-2 клетках после инкубации в течение примерно 15 мин при 37°С. Поглощение красителя ядрами было совершенно исключено, но голубой краситель показал окрашивание внутри цитоплазмы. Митохондрии были мечены красным с TMRM. Распределение [Dmt1, AtnDap4]DALDA по митохондриям показало частичное перекрывание [Dmt1, AtnDap4]DALDA распределения и TMRM распределения. Пример 3. Удаление пероксида водорода с помощью SS-02 и SS-05 (фиг.1) Действие SS-02 и SS-05 (Dmt-D-Arg-Phe-Orn-NH2) на H2O2 определяли люминолиндуцируемой хемилюминесценцией. 25 µM люминола и 0,7 IU пероксидазы хрена добавляли к раствору Н2О2 (4,4 нмоль) и пептида и регистрировали хемилюминесценцию с помощью Chronolong Model 560 агрегометра (Havortown, PA) в течение 20 минут при 37°С. Результаты показывают, что SS-02 и SS-05 в зависимости от концентрации ингибировали отклик люминола, наводя на мысль о том, что эти пептиды могут “убирать” H2O2. Пример 4. Ингибирование пероксидирования липидов (фиг.2) Пероксидирование линолевой кислоты индуцировали водорастворимым инициатором, АВАР (2,2′-азобис (2-амидинопропан)), и липидное пероксидирование определяли по образованию конъюгированных диенов, регистрируя это спектрофотометрически при 236 нм (В.Longoni, W.A.Pryor, P.Marchiafava. Biochem. Biophys. Res. Commun. 233, 778-780 (1997)). 5 мл 0,5 М АВАР и различные концентрации SS-02 инкубировали в 2,4 мл суспензии линолевой кислоты, пока скорость автоокисления не стала постоянной. Результаты показали, что SS-02 в зависимости от концентрации ингибировали пероксидирование линолевой кислоты. Различные пептиды добавляли в концентрации 100 µМ. Эти данные представляли в виде графического изображения образования диенов. За исключением SS-20 (Phe-D-Arg-Phe-Lys-NH2), SS-21 (Циклогексил-D-Arg-Phe-Lys-NH2) и SS-22 (Ala-D-Arg-Phe-Lys-NH2), все другие SS пептиды уменьшали скорость пероксидирования линолевой кислоты. Отмечается, что SS-20, SS-21 и SS-22 не содержали ни тирозинового остатка, ни диметилтиразинового остатка. SS-01, который содержит Tyr больше, чем Dmt, не так эффективен в предотвращении пероксидирования линолевой кислоты. SS-29 представляет собой Dmt-D-Cit-Phe-Lys-NHb, SS-30 представляет собой Phe-D-Arg-Dmt-Lys-NH2, SS-32 представляет собой Dmt-D-Arg-Phe-Ahp (2-аминогептановая кислота)-NH2. Пример 5. Ингибирование LDL окисления (фиг.3). Человеческий LDL (липопротеин низкой плотности) приготовили в свежем виде из запасов плазмы. LDL окисление индуцировали каталитически добавлением 10 мМ CuSO4 и образование конъюгированных диенов регистрировали при 234 нм в течение 5 часов при 37°С (B.Moosmaim and C.Behl, Mol Pharmacol 61, 260-268 (2002)). (А) Результаты показывают, что в зависимости от концентрации SS-02 ингибирует скорость LDL окисления. (В) Добавляли различные пептиды в концентрации 100 µ.М. За исключением SS-20 (Phe-D-Arg-Phe-Lys-NH2), SS-21 (Циклогексил-D-Arg-Phe-Lys-NH2) и SS-22 (Ala-D-Arg-Phe-Lys-NH2), все другие SS пептиды уменьшали скорость пероксидирования линолевой кислоты (снижали скорость образования конъюгированных диенов). Отмечается, что SS-20, SS-21 и SS-22 не содержали ни тирозинового остатка, ни диметилтиразинового остатка. SS-29 представляет собой Dmt-D-Cit-Phe-Lys-NH2, SS-30 представляет собой Phe-D-Arg-Dmt-Lys-NH2, SS-32 представляет собой Dmt-D-Arg-Phe-Ahp(2-аминогептановая кислота)-NH2. Пример 6. Образование пероксида водорода митохондриями, выделенными из клеток печени мыши (фиг.4). Из-за того, что митохондрии – главный источник образования ROS, проверяли действие SS-02 на образование H2O2 в изолированных митохондриях при базальных условиях, а также после обработки антимицином, комплексом III ингибитора. У мышей удаляли печень, собирали сырье и гомогенизировали в ледяном буфере и центрифугировали при 13800×g в течение 10 минут. Осадок промывали один раз и затем ресуспендировали в 0,3 мл промывочного буфера и помещали на лед до использования. H2O2 определяли хемилюминесценцией с люминолом, как описано ранее (Y.Li, H.Zhu, M.A.Trush, Biochim. Biophys. Acta 1428, 1-12 (1999)). 0,1 мг митохондриального протеина добавили к 0,5 мл калий-фосфатного буфера (100 µМ, рН 8,0) в отсутствие или в присутствии SS пептидов (100 мМ). Добавили 25 мМ люминола и 0,7 IU пероксидазы хрена и регистрировали хемилюминесценцию с Chronolong Model 560 агрегометр (Havertown, PA) в течение 20 минут при 37°С. Количество образовавшегося H2O2 определяли как площадь под кривой (AUC) через 20 мин и все данные приводили к AUC, полученной от одних митохондрий (без добавок). (A) Количество образовавшегося H2O2 существенно уменьшилось в присутствии 10 µM SS-02. Добавление антимицина (1 µM) значительно увеличило образование Н2О2 выделенными митохондриями, и это увеличение совершенно блокировалось 10 µМ Dmt1-DALDA (обозначаемого также в описании как dDALDA). (B) Количество образовавшегося H2O2 существенно уменьшалось пептидами SS-02, SS-29, SS-30 и SS-31. SS-21 и SS-22 не оказывали эффекта на образование H2O2. Отмечается, что SS-21 и SS-22 не содержат остатка тирозина или остатка диметилтирозина. Аминокислота Dmt (диметилтирозин) одна также ингибировала образование H2O2. Пример 7. SS-31 ингибирует образование Н2О2 выделенными митохондриями (фиг.5). Н2О2 определяли люминольной хемилюминесценцией, как описано ранее (Y.Li, H.Zhu, M.A.Trush, Biochim. Biophys. Acta 1428, 1-12 (1999)). 0,1 мг митохондриального белка добавили к 0,5 мл калий-фосфатного буфера (100 µМ, pH 8,0) в отсутствие или в присутствии SS-31. Добавили 25 µМ люминола и 0,7 IU пероксидазы хрена, и хемилюминесценцию регистрировали с Chronolong Model 560 агрегометром (Havertown, PA) в течение 20 мин при 37°С. Количество образованного Н2О2 определяли по площади под кривой (AUC) через 20 минут, все данные приводили к AUC, полученной на одних митохондриях (без добавок). (A) SS-31 в зависимости от концентрации уменьшал спонтанное образование H2O2 выделенными митохондриями. (B) SS-31 в зависимости от концентрации уменьшал образование H2O2, индуцируемое антимицином в выделенных митохондриях. Пример 8. SS-02 и SS-31 уменьшали внутриклеточные ROS и увеличивали выживаемость клеток (фиг.6). Для того чтобы показать, насколько заявленные пептиды эффективны применительно к целым клеткам, нейронные N2A клетки помещали в 96-луночные планшеты при плотности 1×104/ячейку и давали им расти в течение 2 дней до обработки tBHP (0,5 или 1 мМ) в течение 40 мин. Затем клетки дважды промывали и замещали одной средой или средой, дополнительно содержащей различные концентрации SS-02 или SS-31, в течение 4 часов. Внутриклеточные ROS измеряли карбокси – H2DCFDA (Molecular Probes, Portland, OR). Гибель клеток оценивали по анализу клеточной пролиферации (MTS анализ, Promega, Madison, WI). Инкубация с tBHP приводила к дозозависимому повышению внутриклеточных ROS (А) и уменьшала жизнеспособность (выживаемость) клеток (В и С). Инкубация этих клеток либо с SS-31, либо с SS-02 в зависимости от концентрации уменьшала внутриклеточные ROS(A) и повышала жизнеспособность клеток (В и С) и ЕС50 в нМ диапазоне. Пример 9. SS-31 предотвращает потерю клетками жизнеспособности (фиг.7). Нейронные N2A и SH-SY5Y клетки помещали в 96-луночный планшет при плотности 1×104/ячейку и выращивали их в течение 2 дней перед обработкой t-бутилгидроксипероксидом (tTBHP) (0,05-0,1 мМ) в отсутствие или в присутствии SS-31 (1012 M÷10-9 М) в течение 24 часов. Гибель клеток оценивали по анализу клеточной пролиферации (MTS исследование, Promega, Madison, WI). Обработка N2A и SH-SY5Y клеток низкими дозами t-BHP (0,05-0,1 мМ) в течение 24 часов приводила к снижению жизнеспособности клеток. (А) 0,05 мМ t-BHP вызывало 50% потерю жизнеспособности у N2A клеток и 30% у SH-SY5Y клеток. (В) 0,1 мМ t-BHP приводило к еще большему снижению жизнеспособности у SH-SY5Y клеток. Конкурентная обработка клеток с помощью SS-31 приводила к дозозависимому уменьшению t-BHP-индуцированной цитотоксичности. Полная защита от t-BHP достигалась при 1 нм SS-31. Пример 10. SS-31 уменьшал активность каспазы (фиг.8). N2A клетки выращивали в 96-луночных планшетах, обрабатывали t-BHP (0,05 мМ) в отсутствие или в присутствии SS-31 (10-11 М – 1018 М) при 37°С в течение 12-24 часов. Все процедуры обработки выполнялись в четырех повторностях. N2A – клетки инкубировали с t-BHP (50 мМ) с или без SS-31 при 37°С в течение 12 часов. Клетки осторожно удаляли из планшета вместе с клеточным отделяющимся раствором (Accutase, Innovative Cell Technologies, Inc., San Diego, (CA) и дважды промывали PBS. Активность каспазы оценивали с помощью FLICA набора (Immunochemistry Technologies LLC, Bloomington, MN). Согласно инструкциям производителя клетки ресуспендировали (приблизительно 5×106 клеток/мл) в PBS и метили pan-каспазным ингибитором FAM -VAD-FMK в течение 1 часа при 37°С при 5% СО2 и защите от света. Клетки затем промывали, чтобы удалить несвязавшийся реагент и зафиксировать. Интенсивность флуоресценции в клетках измеряли лазерным сканирующим цитометром (Beckman – Coulter XL, Beckman Coulter, Inc., Fullerton, CA) используя стандартные эмиссионные фильтры для зеленого (FL1). Для каждого прогона подбирали 100000 индивидуальных случаев и хранили в типовых файлах для анализа. Активация каспазы представляет собой инициирующий толчок для апоптозного каскада и наши результаты показали значительное повышение каспазной активности после инкубации SH-SY5Y клеток с 50 мМ t-BHP в течение 12 часов, который в зависимости от концентрации ингибировался повышающимися концентрациями SS-31. Пример 11. SS-31 уменьшал скорость аккумуляции ROS (фиг.9). Внутриклеточные ROS определяли с помощью флуоресцентной пробы DCFH-DA (5-(и -6)-карбокси-2′,7′-дихлордигидрофлуоресцеин ацетат). DCFH-DA поступает в клетки и затем деацетилируется до нефлуоресцентного DCFH. DCFH реагирует с ROS с образованием DCF, флуресцентного продукта. N2A клетки в 96-луночныом планшете промывали HBSS и нагружали 10 µМ DCFDA в течение 30 минут при 37°С. Клетки промывали 3 раза HBSS и выдерживали с 0,1 мМ t-BHP, без или вместе с SS-31. Окисление DCFH до DCF регистрировали в текущем времени с помощью флуоресцентного электронного считывателя (Molecular Devices), используя 485 нм для возбуждения и 530 нм для эмиссии. Скорость аккумуляции ROS в N2A клетках, обработанных 0,1 мМ t-BHP, ингибировалась добавлением SS-31 в зависимости от концентрации. Пример 12. SS-31 ингибировал пероксидирование липидов в клетках, подвергнутых окислительному повреждению (фиг.10). SS-31 ингибировал пероксидирование липидов, обработанных t-BHP. Липидное пероксидирование оценивали, измеряя HNE Michael аддуктов. 4-HNE является одним из основных альдегидных продуктов пероксидирования полиненасыщенных жирных кислот мембраны. N2A клетки были засеяны на стеклянный диск за 1 день до t-BHP обработки (1 µМ, 3 часа, 5% СО2) в присутствии или в отсутствие SS-31 (10-8-10-10 М). Клетки затем промывали дважды PBS и фиксировали 30 минут 4% параформальдегидом в PBS при комнатной температуре и затем промывали 3 раза PBS. Затем клетки пермеабилизовали, обработали кроличьими антителами против HNE и потом вторичным антителом (IgG козы к кроличьим антителам, конъюгированные с биотином). Клетки затем поднимали в Vectashield и присматривали в Leiss флуоресцентном микроскопе, используя возбуждение при длине волны 460±20 нм и удлиненный фильтр при эмиссии 505 нм. (А) Необработанные клетки; (В) клетки, обработанные 1 мМ t-BHP в течение 3 часов; (С) клетки, обработанные 1 мМ t-BHP и 10 нМ SS-31 в течение 3 часов. Пример 13. SS-02 ингибирует потерю митохондриального потенциала в клетках, подвергнутых действию пероксида водорода. Сасо-2 клетки обрабатывали t-BHP (1 µМ) в отсутствие или в присутствии SS-02 (0,1 µМ) в течение 4 часов, затем инкубировали с TMRM и проверяли под LSCM. В контрольных клетках митохондрии были четко видны в виде полосок по цитоплазме. В клетках, обработанных tBHP, TMRM флуоресценция очень сильно снижалась, что дает основания предполагать генерализованную деполяризацию. В противоположность этому конкурентная обработка SS-02 предохраняла против деполяризации митохондрий, вызываемой tBHP. Пример 14. SS-31 предотвращает потерю митохондриального потенциала и повышенную ROS аккумуляцию в N2A клетках, вызываемые выдерживанием с t-BHP (фиг.11). N2A клетки на стеклянном диске обрабатывали 0,1 мМ t-BHP с 1 нМ SS-31 или без него, в течение 6 часов. Клетки потом нагружали 10 µМ дихлорфлуоресцина (ex/em=485/530) в течение 30 минут при 37°С, 5% СО2. Затем клетки промывали HBSS три раза и окрашивали 20 нМ Mitotracker TMRM (ex/em=550/575 нм) в течение 15 минут при 37°С и исследовали конфокальной лазерной сканирующей микроскопией. Обработка N2A клеток с помощью t-BHP привела к потере TMRM флуоресценции, показывающей деполяризацию митохондрий. Был также сопутствующий всплеск – повышение DCF флуоресценции, показывающий повышение внутриклеточных ROS. Конкурентная обработка 1 нМ SS-31 предотвращала митохондриальную деполяризацию и снижала ROS аккумуляцию. Пример 15. SS-31 предотвращает апоптоз, вызываемый окислительным стрессом (Фиг.12). SH-SY5Y клетки выращивали в 96-луночном планшете, обрабатывали t-BHP (0,025 мМ) в отсутствие или в присутствии SS-31 (10-12 М – 10-9 М) при 37°С в течение 24 часов. Все виды обработки осуществляли в четырех повторностях. Затем клетки окрашивали 2 мг/мл Hoechst 33342 в течение 20 минут, фиксировали 4% параформальдегидом и оценивали с помощью Zeiss флуоресцентного микроскопа, (Axiovert 200 М), применяя Zeiss Acroplan X20 объектив. Морфологию клеток оценивали, используя длину волны при возбуждении 350±10 нм и удлиняющийся фильтр 400 нм для эмиссии. Получали снимки и анализировали их, применяя MetaMorph программное обеспечение (Universal Imaging Corp., West Chester, PA). Одинаково прокрашенные ядра подсчитывали, учитывая их как здоровые жизнеспособные нейроны, в то время как слипшиеся или фрагментированные ядра подсчитывались и расценивались как апоптозные. SS-31 предотвращает апоптоз, индуцированный низкой дозой t-BHP. Апоптоз оценивали конфокальной микроскопией с флуоресцентной пробой Hoechst 33342. (A1) репрезентативная область клеток, не обработанных t-BHP. (A2) Флуоресцентное изображение, показывающее несколько клеток с плотным фрагментированным хроматином, показывающим апоптозные ядра. (A3) репрезентативная область клеток, обработанных 0,025 мМ t-BHP в течение 24 часов. (А4) Флуоресцентное изображение, показывающее увеличившееся число клеток с апоптозными ядрами. (А5) Репрезентативная область клеток, обработанных 0,025 мМ t-BHP и 1 нМ SS-31 в течение 24 часов. (А6) Флуоресцентное изображение, показывающее сниженное число клеток с апоптозными ядрами. (В) SS-31 в зависимости от концентрации уменьшал долю апоптозных клеток, получившихся в результате обработки низкой дозой t-BHP (0,05 мМ) в течение 24 часов. Пример 16. SS-31 предотвращает пероксидирование липидов в сердцах, подвергнутых кратковременным операциям ишемия/реферфузия (Фиг.13). Выделенные сердца морских свинок перфузировали обратным способом в Langendorff аппарате и подвергли ишемии/реперфузии в различных диапазонах времени. Сердца после этого немедленно фиксировали и погружали в парафин. Иммуногистохимический анализ 4-гидрокси-2-ноненол (НМЕ)-модифицированных протеинов в парафиновых срезах осуществляли, используя антитела к HNE. (A) Иммуногистохимический анализ 4-гидрокси-2-ноненол (HNE)-модифицированных протеинов в парафиновых срезах сердец морских свинок, аэробно перфузированных 30 минут (а) буфером; (b) 100 нМ SS-02; (с) 100 нМ SS-20 и (d) 1 нМ SS-31, подвергнутых затем 30-минутной ишемии и реперфузированных в течение 90 минут с теми же пептидами. Тканевые срезы инкубировали с антителами, специфичными к HNE. (е) Фоновый контроль: окрашивание без первичных антител. (B) Иммуногистохимический анализ HNE-модифицированных протеинов в парафиновых срезах сердец морских свинок, аэробно перфузированных 30 минут буфером, затем подвергнутых 30 мин ишемии и реперфузированных (а) буфером; (b) 100 нМ SS-02; (с) 100 нМ SS-20 и (d) 1 нМ SS-31 в течение 90 минут с теми же пептидами. Тканевые срезы инкубировали с антителами, специфичными к HNE. (е) Фоновый контроль: окрашивание без первичных антител. Пример 17. SS-31 увеличивает коронарный кровоток и уменьшает пероксидирование липидов и апоптоз в сердцах, подвергнутых пролонгированной ишемии на холоду с последующей реперфузией в теплых условиях (Фиг.14). Выделенные сердца морских свинок реперфузировали обратным способом в Langendorff аппарате в кардиоплегическом растворе (St. Thomas раствор) в отсутствие или в присутствии SS-31 (1 нМ) в течение 3 минут, затем скрепляли их и хранили при 4°С в течение 18 часов. Затем повторно загружали в Langendorff аппарат и реперфузировали с Krebs-Henseleit раствором при 34°С в течение 90 минут. После этого сердца быстро фиксировали и погружали в парафин. (A) SS-31 существенно повышал коронарный кровоток в сердцах после 18-часового хранения на холоду в состоянии ишемии. Затемненная зона отражает 18-часовую ишемию на холоду. (B) Иммуногистохимический анализ HNE – модифицированных протеинов в парафиновых срезах сердец морских свинок, которые хранили без (а) или с (b) SS-31 (1 нМ). (с) Фоновое окрашивание без первичного антитела. (C) SS-31 предотвращает апоптоз в эндотелиальных клетках и миоцитах в выделенных сердцах морских свинок, подвергнутых тепловой реперфузии после продолжительной (18 час) ишемии на холоду. Апоптоз оценивали TUNEL окрашиванием (зеленый) и просматривали ядра, применяя DAPI (голубой). Пример 18. SS-31 повышает выживаемость островковых клеток, выделенных из поджелудочной железы мыши (Фиг.15). (A) SS-31 повышает митохондриальный потенциал в островковых клетках, выделенных из поджелудочной железы мыши. Отбирали мышечные поджелудочные железы, известными способами получали из них островковые клетки. В некоторых опытах SS-31 (1 нМ) добавляли ко всем буферам, используемым при выделении, на протяжении всего периода выделения. Потенциал митохондрий измеряли, используя TMRM (красный), и визуально оценивали конфокальной микроскопией. (B) SS-31 уменьшает апоптоз и увеличивает выживаемость островковых клеток, выделенных из поджелудочной железы мыши. Отбирали мышиные поджелудочные железы, известными способами получали из них островковые клетки. В некоторых опытах SS-31 (1 нМ) добавляли ко всем буферам, используемым при выделении, на протяжении всего процесса выделения. Апоптоз оценивали с помощью проточной цитометрии, используя аннексии V, а некроз – с помощью пропидиум иодида. Пример 19. SS-31 предохраняет панкреатические островковые клетки от окислительного повреждения (Фиг.16). Островковые клетки поджелудочной железы мыши не обрабатывали ничем (а), или обрабатывали 25 µM t-BHP без (b) или с 1 нМ SS-31 (c). Митохондриальный потенциал определяли с помощью TMRM (красный) и разновидности реакционноспособного кислорода определяли с помощью DCF (зеленый), используя конфокальную микроскопию. Пример 20. SS-31 предотвращает болезнь Паркинсона (Фиг.17). МРТП является нейротоксином, который избирательно разрушает стриатальные допаминовые нейроны, и может использоваться в качестве животной модели болезни Паркинсона. Мишенью МРР+, метаболита МРТР, являются митохондрии, МРР+ ингибирует комплекс I транспортной цепи электронов и повышает образование ROS. МРР+ используется в исследованиях на клеточных культурах, поскольку клетки не обладают способностью метаболизировать МРТР в активный метаболит. МРТР используется в исследованиях на животных. (A) SS-31 предохраняет допаминовые клетки от МРР+ токсичности. SN-4741 клетки обрабатывали буфером, 50 µM МРР+ или 50 µМ МРР+ и 1 нМ SS-31 в течение 48 часов, и факт апоптоза определяли флуоресцентной микроскопией с Hoechst 33342. Количество слипшихся фрагментов существенно увеличилось при MPP+ обработке. Конкурентная обработка SS-31 уменьшала количество апоптозных клеток. (B) SS-31 в зависимости от концентрации предотвращал гибель допаминовых нейронов у мышей, обработанных МРТР. Три дозы МРТР (10 мг/кг) задавались мышам (n=12) в течение 2 часов. SS-31 вводили за 30 мин до каждой инъекции МРТР и спустя 1 час и 12 часов после последней инъекции МРТР. Неделей позже животных умерщвляли и стриатальные зоны головного мозга подвергали иммуноокрашиванию для выделения тирозингидроксилазы. (C) SS-31 в зависимости от дозы приводил к повышению уровней стриатального допамина, DOPAC (3,4-дигидроксифенилуксусная кислота) и HVA (гомованилловая кислота) у мышей, получивших МРТР. Три дозы МРТР (10 мг/кг) задавались мышам (n=12) в течение 2 часов. SS-31 вводили за 30 минут до каждой инъекции МРТР и спустя 1 час и 12 часов после последней инъекции МРТР. Неделей позже животных умерщвляли и определяли уровни допамина, DOPAC и HVA жидкостной хроматографией под высоким давлением.
Формула изобретения
1. Способ уменьшения окислительного повреждения у млекопитающего, нуждающегося в этом, включающий введение млекопитающему эффективного количества ароматического катионного пептида, выбранного из группы, включающей 2. Способ по п.1, в котором пептид вводят орально. 3. Способ по п.1, в котором пептид вводят местно. 4. Способ по п.1, в котором пептид вводят интраназально. 5. Способ по п.1, в котором пептид вводят системно. 6. Способ по п.1, в котором пептид вводят внутривенно. 7. Способ по п.1, в котором пептид вводят подкожно. 8. Способ по п.1, в котором пептид вводят внутримышечно. 9. Способ по п.1, в котором пептид вводят интрацеребровентрикулярно. 10. Способ по п.1, в котором пептид вводят интратекально. 11. Способ по п.1, в котором пептид вводят трансдермально. 12. Способ по п.35, в котором трансдермальное введение осуществляют путем ионофореза. 13. Способ по п.1, в котором млекопитающее подвергают реперфузии. 14. Способ по п.37, в котором реперфузия является лечением ишемии. 15. Способ по п.38, в котором ишемия является следствием удара. 16. Способ по п.1, в котором млекопитающее страдает от сепсиса. 17. Способ по п.1, в котором млекопитающее страдает от воспалительного процесса. 18. Способ по п.41, в котором млекопитающее страдает от артрита. 19. Способ по п.1, в котором млекопитающее страдает от диабета. 20. Способ по п.41, в котором млекопитающее страдает от рассеянного склероза. 21. Способ по п.1, в котором млекопитающее страдает от повреждения печени. 22. Способ по п.45, в котором повреждение печени вызвано вирусной инфекцией. 23. Способ по п.45, в котором повреждение печени вызвано токсическим агентом. 24. Способ по п.1, в котором млекопитающее страдает от нейродегенеративного заболевания или состояния. 25. Способ по п.48, в котором нейродегенеративное заболевание или состояние является болезнью Паркинсона. 26. Способ по п.48, в котором нейродегенеративное заболевание или состояние является болезнью Альцгеймера. 27. Способ по п.48, в котором нейродегенеративное заболевание или состояние является болезнью Гентингтона. 28. Способ по п.48, в котором нейродегенеративное заболевание или состояние является Амиотрофическим Латеральным Склерозом (ALS). 29. Способ по п.1, в котором млекопитающее – человек. 30. Способ уменьшения окислительного повреждения в удаленном органе млекопитающего, включающий введение в удаленный орган эффективного количества ароматического катионного пептида, выбранного из группы, включающей 31. Способ уменьшения окислительного повреждения в клетке, нуждающейся в этом, включающий введение в клетку эффективного количества ароматического катионного пептида, выбранного из группы, включающей
РИСУНКИ
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 2, p.260-268. RU 2115660 C1, 20.07.1998. US 2003109457 A1, 12.06.2003. РЕБРОВА Т.Ю. и др. Вклад системы антиокислительных ферментов в реализацию картиопротекторного эффекта опиоидов приокислительном стрессе. Вопросы медицинской химии, 2001, т.47,
2, p.260-268. RU 2115660 C1, 20.07.1998. US 2003109457 A1, 12.06.2003. РЕБРОВА Т.Ю. и др. Вклад системы антиокислительных ферментов в реализацию картиопротекторного эффекта опиоидов приокислительном стрессе. Вопросы медицинской химии, 2001, т.47,  -положении относительно карбоксильной группы.
-положении относительно карбоксильной группы. -аминомасляную кислоту,
-аминомасляную кислоту,  -аминомасляную кислоту,
-аминомасляную кислоту,  -аминовалериановую кислоту и
-аминовалериановую кислоту и  -аминокапроновую кислоту. Примеры арилсодержащих аминокислот включают орто-, мета- и парааминобензойную кислоты. Примеры алкиларилсодержащих аминокислот включают орто-, мета- и парааминофенилуксусную кислоты и
-аминокапроновую кислоту. Примеры арилсодержащих аминокислот включают орто-, мета- и парааминобензойную кислоты. Примеры алкиларилсодержащих аминокислот включают орто-, мета- и парааминофенилуксусную кислоты и 
 ), радикал супероксидного аниона (O2
), радикал супероксидного аниона (O2