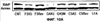Патент на изобретение №2376018
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) ЛЕЧЕНИЕ ПРОЛИФЕРАТИВНЫХ ЗАБОЛЕВАНИЙ С ИСПОЛЬЗОВАНИЕМ АНТИСМЫСЛОВОГО ОЛИГОМЕРА IAP И ХИМИОТЕРАПЕВТИЧЕСКОГО ПРЕПАРАТА
(57) Реферат:
Изобретение относится к области медицины и касается лечения пролиферативных заболеваний с использованием антисмыслового олигомера IAP и химиотерапевтического препарата. Сущность изобретения включает способ лечения больного, страдающего пролиферативным заболеванием с использованием антисмыслового олигомера SEQ ID NO:151 или его фармацевтически приемлемой соли и химиотерапевтического препарата. Преимущество изобретения заключается в повышении эффективности лечения. 27 з.п. ф-лы, 25 ил., 9 табл.
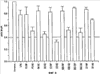
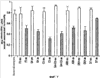
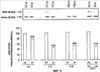
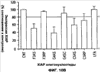
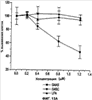
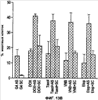
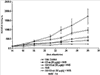
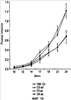
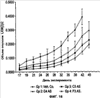
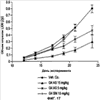
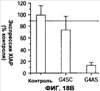
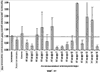
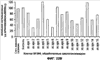
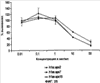
Уровень техники Изобретение относится к лечению рака и других пролиферативных заболеваний. Одним из путей гибели клеток является апоптоз или программируемая клеточная смерть. Апоптоз зачастую является естественной стадией развития и существования здоровых тканей. Этот процесс может происходить настолько быстро, что его трудно зафиксировать. Известно, что вступление клеток в апоптоз играет критическую роль в таких процессах как, например эмбриогенез, вирусный патогенез, развитие рака, аутоиммунные нарушения и нейродегенеративные заболевания. Предполагают, что нарушение процесса апоптоза имеет место при развитии рака, аутоиммунных заболеваниях, таких как системная красная волчанка и рассеянный склероз, и вирусных инфекциях, включая инфекции, ассоциированные с вирусом герпеса, поксвирусом и аденовирусом. В последние годы стала ясна важная роль апоптоза при раке. Идентификация в конце 70-х годов онкогенов, стимулирующих деление клеток, дала начало практически всеобщей концентрации внимания на клеточной пролиферации, что являлось доминирующей темой исследований в онкобиологии на протяжении многих лет. Господствовавшая долгое время догма постулировала, что способы противораковой терапии предпочтительно должны быть направлены на быстро делящиеся по сравнению с нормальными «раковые» клетки. Это объяснение не было полностью удовлетворительным, поскольку некоторые медленно растущие опухоли легко поддаются лечению, в то время как многие опухоли, характеризующиеся быстрым делением клеток, оказываются исключительно устойчивыми к противораковой терапии. Прогресс в области онкологии привел к появлению в онкобиологии новой парадигмы, согласно которой новообразование рассматривается как неспособность клетки осуществлять нормальный путь программируемой клеточной смерти. Нормальные клетки получают постоянную обратную связь от своих соседей посредством разнообразных факторов роста и совершают «самоубийство» при нарушении контекста межклеточных взаимодействий. Раковые клетки некоторым образом способны игнорировать эти команды и продолжают неуместное деление. В настоящее время считается, что многие способы противораковой терапии, включая лучевую терапию и многие химиотерапевтические схемы, как считалось действующие посредством повреждения клеток, в действительности представляют собой пусковые факторы апоптоза. Как нормальные типы клеток, так и раковые клетки проявляют широкий спектр восприимчивости к факторам, запускающим апоптоз, однако детерминанты этой устойчивости находятся до сих пор на стадии изучения. Многие типы нормальных клеток претерпевают временную остановку роста в ответ на сублетальную дозу излучения или цитостатика, в то время как находящиеся по соседству раковые клетки входят в апоптоз. Этот дифференцированный эффект при определенной дозе открывает ключевые терапевтические возможности, обеспечивающие успешное лечение онкологических заболеваний. Исходя из этого, не вызывает удивления тот факт, что устойчивость раковых клеток к апоптозу, вероятнее всего, является главной причиной неэффективности лечения рака. Идентифицированы несколько эндогенных белков, которые являются мощными ингибиторами апоптоза, включая семейства белков IАР и Всl-2 у млекопитающих. Некоторые представители IАР семейства непосредственно ингибируют терминальные эффекторные каспазы, например каспазу-3 и каспазу-7, принимающих участие в осуществлении смерти клетки, наряду с ключевой инициаторной митохондриальной каспазой, каспазой-9, играющей важную роль в опосредовании клеточной смерти, вызванной химиотерапией. Белки IАР являются единственными известными на сегодняшний момент эндогенными ингибиторами каспаз и таким образом играют центральную роль в регуляции апоптоза. Установлено, что белки IАР играют важную роль в развитии некоторых видов рака и предполагаемая хромосомная транслокация, затрагивающая конкретный IАР (с1АР2/Н1АР1) была идентифицирована при MALT-лимфоме. Недавно у больных острой миелогенной лейкемией была выявлена взаимосвязь между повышением уровня XIAP, неблагоприятным прогнозом и коротким периодом выживания. Более того, отмечалась значительная гиперэкспрессия гена XIAP во многих линиях опухолевых клеток, депониированных НИР (Национальный институт рака). Таким образом, существует необходимость в усовершенствованных противоопухолевых препаратах, в особенности препаратов, способных индуцировать в клетках процесс апоптоза и обходить анти-апоптозные сигналы, возникающие в таких клетках. Краткое описание изобретения. Настоящее изобретение относится к способам индуцирования апоптоза в клетке. Способы, описанные в настоящем изобретении, могут быть использованы для лечения рака и других пролиферативных заболеваний. Настоящее изобретение относится к способу лечения пациентов, страдающих пролиферативными заболеваниями, такими как рак, посредством введения антисмыслового олигонуклеотида IAP и химиотерапевтического препарата. Химиотерапевтический препарат и антисмысловой олигонуклеотид IAP вводят одновременно или с интервалом, не превышающим 28 дней (например, 21 день, 14 дней, 7 дней, 1 день или 1 час), в дозах, которые в совокупности достаточны для оказания удовлетворительного лечебного эффекта. Антисмысловые олигонуклеотиды IAP сокращают количество продуцируемого белка IAP, что позволяет клетке, которая нормально экспрессирует IAP, войти в апоптоз. Указанный процесс осуществляется посредством введения олигонуклеотидов, которые специфично гибридизуются с одним и более полинуклеотидом, кодирующим IAP. Специфичная гибридизация олигонуклеотида согласно настоящему изобретению с полинуклеотидом, кодирующим IAP (например, РНК или ДНК), препятствует нормальной функции вышеуказанного полинуклеотида, кодирующего IAP, что приводит к сокращению продукции IAP. Молекула нуклеиновой кислоты, которая модулирует функцию нуклеиновой кислоты-мишени путем специфической гибридизации с указанной кислотой-мишенью, обычно называется «антисенс-препарат». Несмотря на то, что может быть использован любой антисмысловой олигонуклеотид IАР, уменьшающий экспрессию IАР, в одном аспекте настоящего изобретения олигонуклеотид насчитывает от 8 до 30 нуклеотидов в длину и содержит, по меньшей мере, восемь последовательно идущих нуклеотидов последовательности, выбранной из SEQ ID NO: 1-99, 143, 147, 151, 163-260, 287, 289, и 300-460. В отдельных способах реализации олигонуклеотид содержит последовательность, выбранную из SEQ ID NO: 1-99, 143, 147, 151, 163-260, 287, 289, и 300-460. Желательно, чтобы олигонуклеотид состоял (или преимущественно состоял) из одной или более вышеуказанных SEQ ID NO. Например, олигонуклеотид может содержать последовательность, выбранную из SEQ ID NO 97, 98, 99, 143, 147, 151, 287, и 289; SEQ ID NO 300-389, или SEQ ID NO 390-460. В наиболее предпочтительном способе реализации, настоящее изобретение, предусматривает олигонуклеотид, который содержит 11 остатков ДНК (дезоксирибонуклеотидов), фланкированных с каждой стороны (находящихся между) четырьмя 2′-O-метилированными остатками РНК (остатки 2′-O-метил РНК) и состоящий из одной из следующих последовательностей: 5′-UUGGT TTCCAATGTGUUCU-3″ (SEQ ID NO: 155); 5′-ACACGACCGCTAAGAAACA-3′ (SEQ ID NO: 16); 5′-ACAGGACTACCACTTGGAA-3′ (SEQ ID NO: 157); 5″-UGCCAGTGTTGATGCUGAA-3′ (SEQ ID NO: 27); 5’GCUGAGTCTCCATATUGCC-3″ (SEQ ID NO: 141); 5′-UCGGGTATA TGGTGTCUGA-3′ (Номер SEQ ID: 41); 5′-AAGC ACTGCACTTGGUCAC-3′ (SEQ ID NO: 47); 5′-CCGGCCCAAAACAAAGAAG-3′ (SEQ ID NO: 51); 5′-ACCCTGGATACCATTUAGC-3′ (Номер SEQ ID: 63); 5′-UGUCAGTACA TGTTGGCUC-3′ (Номер SEQ ID NO: 161); и 5′-UGCACCCTGGATGCCAUUU-3′ (Номер SEQ ID NO: 151). В другом способе реализации настоящего изобретения антисмысловой олигонуклеотид IAP насчитывает до 30 нуклеотидов в длину и содержит, по меньшей мере, 8 последовательных нуклеотидов из последовательности, выбранной из SEQ ID NO: 461-490. Другие олигонуклеотиды IAP, которые могут назначаться в сочетании с химиотерапевтическим препаратом, – это олигонуклеотиды, которые гибридизуются при жестких условиях с полинуклеотидом, кодирующим полипептид IАР и отобранным из белков NAIP (Bircl), HIAP1 (cIAP2, API2, MIHC, hITA), HIAP2 (cIAPI, MIHB), XIAP (hILP, hILP1, MIHA, AP13), сурвивина (TIAP, MIHD, AP14), ливина (KIAP, ML-IAP, cIAP3, HIAP3) и hILP2 (Ts-IAP, TIAP. Олигонуклеотид, который используют в способе согласно настоящему изобретению, может содержать, по меньшей мере, одну модифицированную связь (например, фосфоротиоатную, метилфосфонатную, фосфотриэфирную, фосфородитиоатную или фосфоселенатную связи), модифицированный нуклеотид (например, 5-метил цитозин) и/или модифицированную остаток сахара (например, 2′-O-метоксиэтильная группа или 2′-O-метильная группа). В одном из способов реализации изобретения олигомер представляет химерный олигомер (например, олигонуклеотид, содержащий остатки ДНК, связанные вместе фосфоротиоатными или фосфодиэфирными связями, и фланкированные, по меньшей мере, одной, двумя, тремя или четырьмя 2′-O-метилированными остатками РНК, связанными между собой фосфоротиоатными связями). В другом своем аспекте настоящее изобретение отностся к способу стимуляции процесса апоптоза в клетке. Этот способ включает стадию введения в клетку антисмыслового олигонуклеотида IAP и химиотерапевтического препарата одновременно, либо с интервалом не более 28 дней в таких количествах, которые в совокупности достаточны для стимуляции апоптоза. Клетка может находиться ех vivo или in vitro. В одном способе реализации изобретения в качестве клетки выступает раковая клетка (например, раковая клетка человека) или клетка лимфоидного или миеломного происхождения. Рак может представлять собой, например острый лейкоз, острый лимфолейкоз, острый миелобластный лейкоз, острый миелоцитарный лейкоз, промиелоцитарная лейкемия, острый миеломоноцитарный лейкоз, острый эритролейкоз, хронический лейкоз, хронический миелоцитарный лейкоз, миелодиспластический синдром, хронический лимфолейкоз, истинная полицитемия, лимфома, болезнь Ходжкина, макроглобулинемия Вальденстрома, фибросаркома, липосаркома, хондросаркома, остеогенная саркома, хордома, ангиосаркома, эндотелиосаркома, лимфангиосаркома, лимфангиоэндотелиосаркома, синовиома, мезотелиома, опухоль Юинга, лейомиосаркома, рабдомиосаркома, рак кишечника, рак поджелудочной железы, рак груди, рак яичника, рак простаты, папиллярная карцинома, плоскоклеточный рак, базально-клеточный рак, аденокарцинома, рак потовой железы, рак сальной железы, цистаденокарцинома, медуллярный рак, рак бронха, почечноклеточный рак, гепатома, карцинома желчных протоков, хориокарцинома, семинома, тератокарцинома, опухоль Вильма, рак шейки матки, рак тела матки, рак яичка, рак легких, мелкоклеточный рак легких, рак желчного пузыря, эпителиальная карцинома, глиома, астроцитома, медуллобластома, краниофарингиома, эпендимома, пинеалома, гемангиобластома, невринома слухового нерва, олигодендроглиома, шваннома, менингиома, меланома, нейробластома или ретинобластома. При лечении рака может быть желательно назначать дополнительно один или несколько химиотерапевтических препаратов, препарат, влияющий на биологический ответ и/или химиосенсибилизирующие средства. Желательно, чтобы введение одного или нескольких вышеперечисленных препаратов проводилось в пределах 28 дней со дня введения олигонуклеотида. Химиотерапевтический препарат и олигонуклеотид можно вводить одним путем либо различными путями. Несмотря на то, что допустим любой способ введения, обеспечивающий адекватное количество препарата в нужном месте, предпочтительными являются внутривенный и внутриопухолевый пути введения. В другом своем аспекте настоящее изобретение относится к фармакологическому составу, который содержит химиотерапевтический препарат и антисмысловой олигонуклеотид IАР, где оба компонента представлены в таких количествах, которые в совокупности достаточны для оказания лечебного эффекта у пациента, страдающего пролиферативным заболеванием (например, раком). При необходимости, фармакологический препарат может дополнительно содержать другие компоненты (например, коллоидную дисперсную систему). Изобретение также относится к способу лечения пациентов, страдающих пролиферативным заболеванием (например, таким как рак или лимфопролиферативные процессы), который предусматривает введение больному химиотерапевтического препарата и каталитической молекулы РНК или экспрессионного вектора, кодирующего данную молекулу РНК, где оба компонента вводятся одновременно или с интервалом, не превышающим 28 дней в дозе, которая в совокупности достаточна для оказания лечебного эффекта. В предпочтительном способе реализации изобретения связующие фрагменты каталитической молекулы РНК содержат, по меньшей мере, восемь последовательных нуклеотидов, соответствующих антисмысловому олигонуклеотиду IАР (например, любую последовательность нуклеотидов, из приведенных в таблицах 2, 3, 7, 8 и 9). Молекула РНК предпочтительно содержит структуру типа «головки молотка», но допустима, также, структура типа «шпильки», дельта-вируса гепатита, интрона группы 1, структур типа VS РНК или РНК RNaseP. Изобретение также относится к способу лечения пациента, страдающего раком или лимфопролиферативным заболеванием, который предусматривает введение указанному пациенту химиотерапевтического препарата и двухцепочечной молекулы РНК, содержащей от 21 до 29 нуклеотидов, из которых, по меньшей мере, восемь последовательно идущих нуклеотидов соответствуют антисмысловому олигонуклеотиду IAP (например, любая последовательность, из приведенных в таблицах 2, 3, 7, 8 и 9). Химиотерапевтический препарат и двухцепочечная молекула РНК вводятся одновременно или с интервалом, не превышающим 28 дней, в количествах, которые в совокупности достаточны для оказания благоприятного лечебного эффекта. В другом родственном аспекте настоящее изобретение также оотносится к способу лечения пациента, страдающего раком или лимфопролиферативным заболеванием, который предусматривает введение указанному пациенту химиотерапевтического препарата и двухцепочечной молекулы РНК, состоящей из 50-70 нуклеотидов, первый домен которой состоит из 21-29 нуклеотидов, содержащих, по меньшей мере, восемь последовательных нуклеотидов, соответствующих антисмысловому олигонуклеотиду IPA (например, любая последовательность из приведенных в таблицах 2, 3, 7, 8, и 9); второй домен, которой комплементарен первому домену, а петля домена расположена между первым и вторым доменами таким образом, что первый и второй домены способны формировать двойную спираль молекулы РНК. Химиотерапевтический препарат и двухцепочечная молекула РНК вводятся одновременно или с интервалом не более 28 дней в количествах, которые в совокупности достаточны для оказания благоприятного лечебного эффекта. Изобретение предусматривает несколько наборов. Один из наборов включает 1) антисмысловой олигонуклеотид IAP, имеющий в длину от восьми до тридцати нуклеотидов, 2) химиотерапевтический препарат и 3) инструкцию по введению антисмыслового олигонуклеотида IAP и химиотерапевтического препарата пациентам с пролиферативным заболеванием в количествах, которые в совокупности достаточны для оказания благоприятного лечебного эффекта у пациента. Другой комплект включает: 1) антисмысловой олигонуклеотид IAP, имеющий в длину от восьми до тридцати нуклеотидов, 2) инструкцию по введению антисмыслового олигонуклеотида IAP и химиотерапевтического препарата пациентам с пролиферативным заболеванием в количествах, которые в совокупности достаточны для оказания благоприятного лечебного эффекта. Кроме того, еще один комплект включает в себя 1) состав, представляющий собой предмет изобретения (описан выше) и 2) инструкции по назначению состава пациентам с пролиферативным заболеванием в количествах, которые в совокупности достаточны для оказания благоприятного лечебного эффекта у пациента. Под «олигонуклеотидом» подразумевается соединение, которое содержит цепочку, состоящую, по меньшей мере, из восьми нуклеотидов, соединенных между собой группами сцепления (связями). В это определение входят естественные и синтетические олигонуклеотиды, как модифицированные, так и немодифицированные, а также миметики олигонуклеотидов, например пептидно-нуклеиновые кислоты (ПНА), кольцевые нуклеиновые кислоты (LNA) и арабинонуклеиновые кислоты (АНК). Олигонуклеотид согласно настоящему изобретению может содержать многочисленные нуклеотиды и связи, включая подробно описанные ниже, в разделе «Олигонуклеотиды и другие нуклеотидные олигомеры». «Белок», «полипептид» или «полипептидный фрагмент» обозначает любую цепочку, состоящую из двух и более аминокислот независимо от модификации, происшедшей после трансляции (например, гликозилирования или фосфорилирования), полностью или частично состоящую из природных полипептидов или пептидов, или искусственно синтезированных полипептидов/пептидов. «Апоптоз» обозначает процесс смерти клетки, в ходе которого погибающая клетка демонстрирует ряд хорошо известных биохимических признаков, включающих образование пузырьков в клеточной мембране, сморщивание сомы клетки, конденсацию хроматина и фрагментацию ДНК. Под термином «ген IAP» подразумевается ген, кодирующий полипептид, имеющий, по меньшей мере, один BIR-домен и способный регулировать (ингибировать или стимулировать) процесс апоптоза в клетке или в тканях при его введении любыми способом интрацеллюлярной или экстрацеллюлярной доставки (см., например, патент США Под «Белком IAP» или «полипептидом IAP» подразумевается полипептид или его фрагмент, кодируемый геном IAP. Полипептиды IAP включают NAIP (Birc1), НIАР1 (cIAP2, API2, MIHC, hIТА), HIAP2 (сIАР1, MINB), XIAP (hILP, hILPI, MIHA, API3), сурвивин (TIAP, MIHD, AP14), ливин (KIАР, ML-IAP, HIAP3), и hILP2 (Ts-IAP, TIAP). Под «биологической активностью» подразумевается любая активность, вызванная полипептидом IАР в условиях in vivo и in vitro. Под «стимуляцией апоптоза» подразумевается увеличение числа клеток, находящихся в состоянии апоптоза, в данной клеточной популяции (например, раковых клеток, лимфоцитов, фибробластов и любых других клеток). Подразумевается, что степень стимуляции апоптоза, происходящей под воздействием апоптоз-стимулирующего препарата в каждом отдельном исследовании отличается, но квалифицированный специалист сможет определить статистически значимое усиление уровня апоптоза, свидетельствующее о том, что олигонуклеотид стимулирует апоптоз, который до этого был ингибирован белком IАР. Предпочтительно, чтобы под «стимуляцией апоптоза» подразумевалось, что количество клеток, входящих в состояние апоптоза составляет, по меньшей мере, 10%, предпочтительно 25% или даже 50%, причем наиболее желательным является, по меньшей мере, двукратное усиление апоптоза по сравнению с клетками, в которые не вводили олигомер согласно настоящему изобретению, но которые в остальном подвергались идентичному воздействию. Предпочтительно, образец, подвергающийся мониторингу, представляет собой образец культуры клеток, которые обычно претерпевают неполноценный апоптоз (например, раковые клетки). Способы, используемые для определения изменения степени апоптоза (например, стимуляции или сокращения) описаны ниже. Под олигонуклеотидом, который «ингибирует экспрессию» гена-мишени (например, IAP), подразумевается такой олигонуклеотид, который сокращает количество мРНК-мишени, или белка, кодируемого данной мРНК, по меньшей мере, на 5%, предпочтительно на 10%, 25% или даже 50% по отношению к не обработанным образцам. Способы, используемые для измерения уровня мРНК и белков хорошо известны, примеры методик описаны ниже. Под «гибридизацией» подразумевается образование водородных связей, которые могут быть в конформации Уотсона-Крика, Hoogsteen-a или обратной конформации Hoogsteen-a между комплиментарными нуклеотидами. Например, аденин или тимин являются комплиментарными нуклеотидами, которые образуют пару благодаря формированию водородных связей. Под «пролиферативным заболеванием» подразумевается заболевание, которое вызвано или развилось в результате неадекватно интенсивного деления клеток, неадекватно низкого уровня апоптоза или обоих этих процессов одновременно. Одним из примеров пролиферативного заболевания является рак. Примеры рака включают все без ограничения лейкозы (например, острый лейкоз, острый лимфолейкоз, острый миелобластный лейкоз, острый миелоцитарный лейкоз, промиелоцитарная лейкемия, острый миеломоноцитарный лейкоз, острый эритролейкоз, хронический лейкоз, хронический миелоцитарный лейкоз, миелодиспластический синдром, хронический лимфолейкоз, истинная полицитемия, лимфома (лимфогранулематоз, лимфома не Ходжкина), макроглобулинемия Вандельстрема, болезнь Н-цепей и солидные опухоли, такие как саркомы и карциномы (например, фибросаркома, миксосаркома, липосаркома, хондросаркома, остеогенная саркома, хордома, ангиосаркома, эндотелиосаркома, лимфангиосаркома, синовиома, мезотелиома, саркома Юинга, лейомиосаркома, рабдомиосаркома, рак толстой кишки, рак поджелудочной железы, рак молочной железы, рак яичника, рак предстательной железы, плоскоклеточный рак, базальноклеточный рак, аденокарцинома, рак потовой железы, рак сальной железы, папиллярный рак, папиллярная аденокарцинома, цистаденокарцинома, медуллярный рак, рак бронха, почечноклеточный рак, гепатома, карцинома желчных протоков, хориокарцинома, семинома, тератокарцинома, опухоль Вильма, рак шейки матки, рак тела матки, рак яичка, рак легких, мелкоклеточный рак легких, рак желчного пузыря, эпителиальная карцинома, глиома, астроцитома, медуллобластома, краниофарингиома, эпендимома, пинеалома, гемангиобластома, невринома слухового нерва, олигодендроглиома, шваннома, менингиома, меланома, нейробластома или ретинобластома. К пролиферативным процессам относятся также лимфопролиферативные заболевания. Предпочтительно, олигонуклеотид, который используют в способе согласно настоящему изобретению, находясь в клетке, которая неспособна нормальным путем войти в состояние апоптоза, оказывает влияние на его стимуляцию и/или снижает уровень мРНК IАР либо содержания белка. Предпочтительно, чтобы это стимулирование уровня апоптоза составляло, по меньшей мере 10% по отношению к контрольным образцам, предпочтительней – 25%, и наиболее предпочтительно – 100% и более. Желательно, чтобы олигонуклеотид, который используют в способе согласно настоящему изобретению, содержал от 8 до 30 нуклеотидов. В конкретных примерах осуществления изобретения, по меньшей мере восемь последовательных нуклеотидов относятся к последовательности, выбранной из SEQ ID NO: 1-99, 143, 147, 151, 163-260, 287, 289, 300-490. Олигонуклеотид согласно настоящему изобретению может также дополнительно содержать, например, 20, 40, 60, 85, 120 или более последовательных нуклеотидов, комплементарных полинуклеотиду, кодирующему полипептид IAP. Олигонуклеотид (или его фрагмент) может содержать измененный (модифицированный) полипептидный остов. Известны фосфоротиоатные, фосфородитиоатные и другие модифицированные остовы. Олигонуклеотид может также содержать одну или несколько искусственных связей. Под «пациентом» подразумевается любое животное (в том числе и человек). К числу животных, которые могут подвергаться лечению с использованием способов, комплектов и составов, описанных в настоящем изобретении, относятся лошади, собаки, кошки, свиньи, козы, кролики, хомяки, обезьяны, морские свинки, крысы, мыши, ящерицы, змеи, овцы, крупный рогатый скот, рыбы и птицы. Под «химиотерапевтическим препаратом» подразумевается препарат, который убивает раковые клетки или замедляет их рост. Соответственно, химиотерапевтическими препаратами считаются как цитотоксические, так и цитостатические препараты. Типичными химиотерапевтическими препаратами являются таксаны (например, паклитаксель, доксетаксель, RPR 109881A, SB-T-1213, SB-T-1250, BMS-275183, BRT 216, DJ-927, MAC-321, IDN5109 и IDN5390), алкалоиды барвинка (например, винкристин, винбластин, виндезин, винфлунин, винорелбин и ангидровинбластин), доластины (доластин-10, доластин-15, ILX651, TZT-1027, симплостатин 1, симплостатин 3 и LU103793), криптофицины (например, эпотилон А, эпотилон В, дезоксиэпотилон В и эпотилон В пактам), элеутеробин, дискодермолид, 2-эпидискодермолид, 2-десметилдисеодермолид, 5-гидроксиметилдискодермолид, 19-десаминокарбонилдискодермолид, 9(13)-циклодискодермолид и лаулималид. Другие препараты приведены в Таблице 1. Под «модулятором биологического ответа» подразумевается препарат, который стимулирует или восстанавливает способность иммунной системы сопротивляться болезни. Некоторые (но не все) препараты, модулирующие биологический ответ, способны замедлять рост опухолевых клеток и поэтому также относятся к химиотерапевтическим препаратам. Примерами препаратов, модулирующих биологический ответ, являются интерфероны (альфа, бета, гамма), интерлейкин-2, ритуксимаб и трастузумаб. Под «химиосенсибилизатором» подразумевается препарат, который делает опухолевые клетки более чувствительными к воздействию химиотерапевтических препаратов. Под «лимфопролиферативным заболеванием» подразумевается заболевание, при котором имеет место патологическая пролиферация клеток лимфатической системы (например, Т-клеток и В-клеток) Под «рибозимом» подразумевается РНК, которая обладает ферментативной активностью, обладающая специфичностью и способностью к расщеплению молекулы РНК-мишени. Рибозимы можно использовать для снижения экспрессии полипептида. Способы использования рибозимов для снижения экспрессии полипептидов описаны, например, у Turner et al. (Adv. Exp. Med. Biol. 465: 293-301,2000) и Norris et al. (Adv. Exp. Med. Biol. 465: 293-301, 2000). Под «репортерным геном» подразумевается ген, кодирующий полипептид, экспрессия которого может быть оценена. Такие полипептиды включают, помимо прочего, глюкуронидазу, люциферазу, хлорамфеникол трансацетилазу и бета-галактозидазу. Под «промотором» подразумевается полинуклеотид, контролирующий транскрипцию. «Функционально связанный» означает, что первый нуклеотид непосредственно примыкает ко второму полинуклеотиду, контролирующему транскрипцию первого, если при этом со вторым нуклеотидом связаны соответствующие молекулы (например, белки – активаторы транскрипции). Другие особенности и преимущества изобретения будут представлены в описании модификаций и формуле изобретения. Краткое описание фигур Фиг.1A-1L представляют собой диаграммы, показывающие действие антисмыслового олигонуклеотида XIAP на экспрессию белка XIAP по отношению к общему содержанию белка (тотальному белку) (Фиг.1А, 1C, 1Е, 1G, 1I и 1К). На фиг.1В, 1D, 1F, 1Н, 1J и 1L приведены значения концентрации тотального белка для каждой трансфекции нуклеотидом по сравнению с результатами контрольной трансфекции без нуклеотида, проводившейся с целью стандартизации результатов для вышеописанного белка XIAP. На Фиг.2F-2C представлены диаграммы, показывающие влияние различных антисмысловых XIAP олигонуклеотидов, применявшихся по отдельности или в комбинации, на (количество) РНК XIAP (Фиг.2А) и белок (Фиг.2В). На Фиг.2С представлена диаграмма значений концентрации тотального белка для каждой трансфекции олигонуклеотида по сравнению с результатами контрольной трансфекции, использовавшихся для стандартизации результатов белка XIAP, показанных на Фиг.2В. На Фиг.3 и 4 представлены диаграммы, показывающие действие смеси 4Х4 олигонуклеотидов остова (mixed backbone nucleotides, MBO) FG8 или Е12 в концентрации 31 нМ (Фиг.3) или 63 нМ (Фиг.4). Клетки рака легких линии Н460 подвергали трансфекции на протяжении 18 часов в течение одного, двух или трех дней олигонуклеотидами MBO в концентрации 125 нМ и Липофектамином 2000. В определенное время производили отбор образцов для вестерн-блот анализа. Производили сканирующую денситометрию, после чего уровни белка XIAP стандартизировали до значений GAPDH и сравнивали с контрольной группой трансфекции, принятой за 100%. Указанные значения в процентах выражают % нокдауна белка XIAP по отношению к специфичным контрольным обработкой смесью нуклеотидов (scrambled control). На Фиг.5A-5D представлены графики, отражающие влияние антисмыслового олигонуклеотида XIAP на жизнеспособность клетки (Фиг.5А, 5С и 5D) и химиосенсибилизацию в присутствии адриамицина (Фиг.5В). На Фиг.6 представлена диаграмма, показывающая олигонуклеотид – опосредованную специфичную понижающую регуляцию (снижение уровня) ХIАР мРНК в клетках 1-1460 in vitro. На чертеже изображены уровни ХIАР мРНК в клетках Н640, на которые воздействовали только Липофектамином 2000 (LFA) или Липофектамином 2000 в комбинации с 1,2 На Фиг.7 представлена диаграмма, показывающая уровень XIAP РНК в клетках Н460 после трансфекции их различными PS-XIAP олигонуклеотидами. Клетки рака легких человека линии Н460 подвергали трансфекции 1 микро Моль PS-олигонуклеотида и Липофектамином 2000 в течение 6 часов. После этого осуществляли сбор клеток для анализа способом Takman. На Фиг.8 представлена диаграмма, показывающая уровень XIAP РНК в клетках Н460 через 9 часов после трансфекции 4Х4 олигонуклеотидами MOB. Клетки Н460 подвергали трансфекции 4Х4 олигонуклеотидами MOB (от 62,5 нМ до 1 µМ) и Липофектамином 2000 в течение 9 часов. После этого осуществляли сбор клеток анализа способом Takman. На Фиг.9 представлена диаграмма, показывающая нокдаун белка XIAP в клетках Н460 через 24 часа после трансфекции 4Х4 МВО. Клетки Н460 в течение 24 часов подвергали трансфекции 4Х4 МВО в концентрации 1 µМ и Липофектимином 2000. После этого осуществляли сбор клеток для анализа способом вестерн-блота. Клетки подвергали сканирующей денситометрии, уровни белка XIAP стандартизировали по уровню актина и сравнивали со специфичными контрольными трансфекциями олигонуклеотидами (sm, rm), значения в которых принимались за 100%. Фиг.10А и 10В представляют собой схематические иллюстрации, изображающие специфическую понижающую регуляцию (снижение уровня) белках XIAP в клетках Н460 in vitro, опосредованную антисенс-олигонуклеотидами. На схеме показаны уровни белка XIAP в клетках Н460, на которые воздействовали только Липофектамином 2000 (LFA) либо Липофектамином 2000 в сочетании с F3, G4, или С5 XIAP олигонуклеотидами в концентрации 1,2 µМ, или при соответствующих контрольных трансфекциях (RP, SC). Уровни белка XIAP анализировали с помощью вестерн-блот анализа (Фиг.10А), количество белка определяли с денситометрии (Фиг.10В). Значения уровней XIAP стандартизировали по уровню клеточного актина и сравнивали с уровнем контрольной группы (CNT). Фиг.11А и 11В представляют собой схематические иллюстрации, показывающие влияние олигонуклеотидов XIAP на активацию каспаз. Показано действие XIAP-олигонуклеотидов F3, G4, или С5 или соответствующих им RP и SC ODN контрольных образцов в концентрации 1,2 µМ, на экспрессию прокаспазы-3, PARP (как полноразмерной – 116 кДа, так и процессированной – 85 кДа)) (Фиг.11А) и на уровни белков Bcl 2 и Вах (Фиг.11В) в клетках Н460, подвергшихся трансфекции по сравнению с контрольными образцами. Экспрессия белков оценивалась с помощью вестерн-блот анализа. Уровни белков Bcl 2 и Вах стандартизировали по уровню клеточного актина и производили их количественную оценку с помощью денситометрии. Соотношение Bcl 2/Bax представлено в виде средней величины двух или трех независимых экспериментов, причем это соотношение в контрольных образцах клеток (CNT) принималось за 1. На Фиг.12А и 12В представлены схематические иллюстрации, показывающие специфическую индукцию апоптоза XIAP-олигонуклеотидами. Индукцию апоптоза определяли в клетках Н460, которые подвергали воздействию AS олигонуклеотида XIAP G4, в концентрации 1,2 µМ, SC олигонуклеотида G4 или в необработанных контрольных образцах (CNT). На Фиг.12 показан процент клеток с суб-G0/G1 (апоптозным) содержанием ДНК при анализе с помощью окрашивания пропидия йодидом (РI) и проточной цитометрии. На Фиг.12В показана морфология ядра клеток Н460, при окраске DAPI, после обработки олигонуклеотидами. Стрелки указывают на клетки, имеющие характерную апоптическую морфологию в виде коденсации или фрагментации ядерной ДНК. На Фиг.13А представлена диаграмма, показывающая влияние обработки AS XIAP – олигонуклеотидом G4 на жизнеспособность клеток Н640. Клетки подвергали воздействию LFA или комплексов LFA-олигонуклеотид с AS или SC олигонуклеотидами G4 в возрастающих концентрациях, после чего спустя 24 часа после воздействия производили оценку жизнеспособности клеток анализом МТТ. Данные представляют собой среднее арифметическое ± SD трех независимых экспериментов. На Фиг.13В представлена диаграмма, показывающая процент мертвых клеток Н460 после воздействия на них LFA и комплексов с AS или SC олигонуклеотидами G4 в количестве 0,4 µМ в присутствии или отсутствии доксорубицина (DOX), таксола, винорельбина (VNВ) или этопозида (Etop), который определяли способом МТТ. Данные представляют собой среднее арифметическое ± SD трех независимых экспериментов. На Фиг.14 представлена диаграмма, показывающая относительный рост опухоли Н460 у мышей, получавших ASXIAP 2×2 МВО в сочетании с винорельбином. Производили внутриопухолевое введение олигонуклеотида в 50 опухолевых масс у мышей с подкожными ксенотрансплантатами клеток Н460. Данное лечение сочетали с введением винорельбина. На Фиг.15 представлена диаграмма, показывающая средний размер Н460 опухолей у мышей, систематически получавших AS PS-олигонуклеотиды XIAP. Систематическое ведение AS PS-олигонуклеотидов XIAP мышам SCID-RAG2 с подкожными ксенотрансплантатами клеток Н460 привело к сокращению размера опухолей по сравнению с контрольной группой. На Фиг.16 представлена диаграмма, показывающая размер опухоли, образованной клетками MDA-MB-435/LCC6 рака молочной железы человека (LCC-клетки) у мышей, систематически получавших AS PS-олигонуклеотиды XIAP. Систематическое введение (и.п.) AS PS-олигонуклеотидов XIAP самкам мышей SCID-RAG2 с ксеноимплантатом LCC-клеток в грудных жировых складках привело к уменьшению размера опухолей по сравнению с контрольной группой. На Фиг.17 представлена схематическая иллюстрация, показывающая влияние олигонуклеотидов G4 на рост опухоли и уровень белка XIAP в опухоли in vivo. Было обнаружено противоопухолевое действие систематически вводимых, чистых AS или SC G4 нуклеотидов XIAP на рост подкожных ксенотрансплантатов клеток Н460 у самцов мышей SCID-RAG2. Все данные представлены в виде среднего арифметического ± SEM (n=6 мышей/группу). На Фиг.18А и 18В представлены схематические иллюстрации, показывающие уровни экспрессии белка XIAP в ксенотрансплантатах опухоли Н460 у мышей SCID-RAG2 после 21-дневного ведения AS или SC олигонуклеотидов С4 или только носителя (контроль), которые анализировали с помощью вестерн-блотов и количественно определяли с помощью денситометрии. Затем уровень XIAP стандартизировали по уровню клеточного актина. Все данные представлены в виде среднего арифметического ± SD (n=3). На Фиг.19А и 19В представлены микрофотографии, показывающие действие in vivo олигонуклеотидов G4 на гистопатологию опухоли Н460, имплантированной мышам SCID-RAG2, через 21 день после системного введения им AS или SC G4 олигонуклеотидов XIAP в дозе 15 мг/кг. На Фиг.19А изображены срезы опухоли, окрашенные гематоксилином и эозином. На Фиг.19В показана иммуногистохимия экспрессии убиквитина в срезах опухоли. Представлены репрезентативные микрофотографии. Маркеры на внутренней шкале соответствуют 100 µм. На Фиг.20А и 20В представлены диаграммы, отображающие повышение эффективности винорельбина (VNB), в сочетании с олигонуклеотидами XIAP. Противоопухолевую активность VNB по отношению к ксенотрансплантатам опухоли Н460 в сочетании с G4 AS или SC G4 олигонуклеотидами XIAP или без них, определяли у мышей SCID- RAG2. На фиг.20А показано противоопухолевое действие изолированно вводившихся препаратов, на фиг.20В – противоопухолевое действие комбинации VNB и олигонуклеотида G4. Все данные представлены в виде среднего арифметического ± SEM (n=6 мышей/группу). На Фиг.21 представлена диаграмма, показывающая действие олигонуклеотидов НIАР1 на уровень НIАР1 РНК. На Фиг.22А и 22В представлены схематические иллюстрации, на которых изображены денситометрические профили результатов вестерн-блот анализа, показывающие действие олигонуклеотидов НIАР1 на способность клеток блокировать повышающую регуляцию (увеличение количества) белка НIАР1, индуцированную циклогексимидом. На Фиг.23 представлена диаграмма, изображающая действие олигонуклеотидов НIАР1 на цитотоксичность, оцененное по тотальному белку. На Фиг.24 представлена диаграмма, показывающая подтверждение специфичности последовательности для олигонуклеотида НIАР1 АРО 2. На Фиг.25 представлена диаграмма, показывающая действие олигонуклеотидов НIАР1 на химиосенсибилизацию устойчивых к химиотерапии глиобластом SF295. Подробное описание изобретения Изобретение относится к способам стимуляции апоптоза в клетках, посредством введения нуклеотидного олигомера, согласно настоящему изобретению в сочетании с химиотерапевтическим препаратом, например цитотоксическим препаратом, цитостатическим препаратом или препаратом, модулирующим биологический ответ. Предпочтительным образом, оба препарата вводят одновременно или с промежутком не более 28 дней друг от друга. В некоторых способах реализации изобретения химиотерапевтический препарат представляет собой вещество, которое разрушает или стабилизирует микротрубочки. В качестве химиотерапевтического препарата можно, например, использовать таксан или алкалоид барвинка. При необходимости можно дополнительно назначать химиосенсибилизатор, т.е. вещество, которое делает пролиферирующие клетки более восприимчивыми к химитерапии. Для создания лекарственного препарата можно использовать любую комбинацию вышеупомянутых компонентов. Этот лекарственный препарат можно использовать при лечении пролиферативных заболеваний, таких как рак или лимфопролиферативные заболевания, а также их симптомов. Комбинация олигонуклеотид/химиотерапевтический препарат, согласно настоящему изобретению при лечении рака или пролиферативного заболевания может также применяться в сочетании с лучевой терапией. Активация апоптоза в раковых клетках предлагает совершенно новый и потенциально успешный способ улучшения биологического ответа пациента на традиционную химио- или лучевую терапию. ХIАР представляет собой наиболее действенный представитель семейства генов IАР в отношении его способности непосредственно ингибировать каспазы и подавлять апоптоз. Мы изучили влияние понижающей регуляции (снижения экспрессии или количества, супрессия) белка ХIАР с помощью антисенс (АS) олигонуклеотида на рост клеток не мелкоклеточного рака легких человека (NCI-H460) in vivo и in vitro. В культивируемых клетках рака легких человека (Н460) AS олигонуклеотид G4 был идентифицирован как наиболее сильное соединение, оказывающее эффективное супрессивное действие на ХIАР мРНК, изменяя ее содержание на 55% и уровень белка на 60% по результатам RT-PCR в режиме реального времени и вестерн-блот анализа соответственно. Он также индуцировал смерть 60% клеток при концентрации 1,2 µМ. В противоположность этому, олигонуклеотид G4 при рандомизированном контроле (Scrambled control) вызвал незначительное снижение уровня XIAP и менее 10% смерти клеток. Воздействие на клетки G4 AS при концентрациях 1,2 µМ вызвало апоптоз, который выявляли по деградации про-каспазы-3 и белка PARP, сопровождавшейся значительной степенью концентрации и фрагментации ядерной ДНК. Более того, AS олигонуклеотид XIAP сенсибилизирует клетки Н460 к цитотоксическому действию доксорубицина, таксола, винорельбина и этопозида. В животных моделях нам удалось продемонстрировать, что G4 AS в дозе 15 мг/кг оказывает значительный ингибирующий эффект на специфический рост ксенотрансплантатов опухоли Н460 человека у мышей группы SCID/RAG2 с иммунодефицитом при его системном введении интраперитонеально. Системное введение AS ODN сопровождалось 85% супрессией белка XIAP в опухолевых ксенотрансплантатах. Комбинация 15 мг/кг G4 AS и 5 мг/кг винорельбина в значительной степени ингибирует рост опухоли, причем в большей степени, чем каждый из данных препаратов по отдельности. Эти исследования свидетельствуют о том, что супрессия белка XIAP представляет собой сильный сигнал, приводящий к смерти клеток карциномы легких и способна индуцировать апоптоз in vitro, а также ингибировать рост опухоли in vivo. Эти исследования подтверждают мнение о том, что белки IАР являются подходящей мишенью для онкотерапии при мелкоклеточном раке легких человека, а также и при других солидных опухолях. Лечение Лечение может проводиться в местах, где обычно проводится онкотерапия: дома, в кабинете врача, в клинике, в поликлиническом отделении, или в стационаре. Как правило, лечение начинается в стационаре, так чтобы врач мог непосредственно наблюдать за его ходом и вносить все необходимые поправки. Длительность терапии зависит от типа рака, возраста и состояния пациента, стадии и типа заболевания и того, как отвечает организм данного пациента на проводимое лечение. Введение препарата можно осуществлять с различными интервалами (например, ежедневно, раз в неделю или раз в месяц). Лечение можно проводить в виде периодических циклов, которые включают перерывы, во время которых организм пациента имеет возможность образовать новые здоровые клетки и восстановить свои силы. В зависимости от типа рака и стадии его развития указанное лечение можно использовать для замедления распространения рака, его роста, для уничтожения или обезвреживания клеток, которые могут распространяться из первичной опухоли в другие органы, для облегчения симптомов, обусловленных раком или предотвращения развития первичного рака. В контексте настоящего изобретения термины «рак» и «злокачественное новообразование» означают скопление клеток, которые патологически размножаются. Злокачественный рост характеризуется бесконтрольностью и прогрессированием и происходит при условиях, которые в норме либо не вызывают, либо останавливают деление нормальных клеток. Традиционная фармацевтика можно использовать для приготвления соответствующих составов и лекарственных форм для введения препарата пациентам, страдающим заболеванием, вызванным избыточной пролиферацией клеток. Препарат можно назначать до того, как у пациента появится симптоматика. При этом можно использовать любой путь введения, например парентеральный, внутривенный, интраартериальный, подкожный, внутриопухолевый, внутримышечный, интракраниальный, интраорбитальный, глазной, интравентрикулярный, внутрипеченочный, интракапсулярный, интратекальный, интрацистернальный, интраназальный, в виде аэрозоля, суппозиториев или форм для орального применения. Например, лекарственные формы могут представлять собой жидкие растворы или суспензии; для орального применения можно использовать форму в виде таблеток или капсул, для интраназальных форм – в виде порошков, каплей в нос или аэрозолей. Широко известные методы изготовления лекарственных форм приведены, например, в “Remington: The Science and Practice of Pharmacy” Ed. A. R. Gennaro, Lippincourt Williams & Wilkins, Philadelphia, PA, 2000. Лекаственные формы для парентерального введения могут, например, содержать наполнители, стерильную воду или физраствор, полиалкиленгликоли, например полиэтиленгликоль, масла растительной природы или гидрогенизированые нафталины. Биосовместимый, подвергающийся биологическому разложению лактидный полимер, лактидный/полигликолидный полимер или сополимер полиоксиэтилен-полиоксипропилен можно использовать для контроля высвобождения ингредиентов (в организме). Другие удобные системы парентеральной доставки компонентов, оказывающих модулирующее действие на IАР, могут включать частицы сополимера этиленвинилацетата, осмотические насосы, имплантируемые инфузионные системы и липосомы. Ингаляционные лекарственные формы могут содержать наполнители, например, полиоксиэтилен-9-лауриловый эфир, гликохолат и дезоксихолат или представлять собой масляные растворы в виде каплей в нос или геля. Лекарственные формы можно вводить больным людям в количествах, обеспечивающих терапевтический эффект (например, предотвращающих, устраняющих или ослабляющих патологическое состояние) в отношении заболевания или состояния. Предпочтительная дозировка олигомера согласно настоящему изобретению зависит от таких параметров, как тип и объем нарушения, общее состояние отдельно взятого пациента, наполнители, входящие в препарат и способ введения. Как описано выше, при желании лечение нуклеотидным олигомером, относящимся к данному изобретению, можно сочетать с другими способами лечения пролиферативных заболеваний (например, лучевой терапией, хирургическими методами или химиотерапией). Для любого из вышеописанных способов применения, олигонуклеотид, относящийся к настоящему изобретению, предпочтительно должен вводиться внутривенно или подводиться непосредственно к области, в которой требуется индуцировать апоптоз (например, с помощью инъекции). Олигонуклеотиды и другие нуклеотидные олигомеры По меньшей мере два типа олигонуклеотидов индуцируют расщепление РНК РН-азой Н: полидезоксинуклеотиды с фосфодиэфирными (РО) или фосфоротиоатными связями (PS). Несмотря на то, что 2′-OMe-RNK последовательности проявляют высокую степень сродства к РНК-мишеням, эти последовательности не являются субстратом для РН-азы Н. Предпочтительным является нуклеотид, в основе которого находится 2′-модифицированный олигонуклеотид с олигодезоксинуклеотидными «брешами» (участками отсутствия нескольких нуклеотидов) и некоторыми или всеми связями, модифицированными в фосфоротиоатные, что обусловливает его резистентность к нуклеазе. Наличие метил-фосфонатных модификаций увеличивает сродство олигонуклеотида к РНК-мишени и, таким образом, снижает IC50. Указанная модификация также повышает резистентность модифицированного олигонуклеотида к нуклеазе. Очевидно, что способы и реагенты согласно настоящему изобретению могут быть использованы в сочетании с любой потенциально разработанной технологией, включая ковалентно-закрытые множественные антисенс-олигонуклеотиды (CMAS) (Moon et Biochem J. 346: 295-303, 2000; PCT Publication Как известно, нуклеозид представляет собой комбинацию основания с остатком сахара. Основная часть нуклеозида, как правило, представляет собой гетероциклическое основание. Два наиболее распространенных класса таких гетероциклических оснований – пурины и пиримидины. Нуклеотиды – это нуклеозиды, которые дополнительно содержат фосфатную группу, присоединенную ковалентными связями к сахарному компоненту нуклеозида. В нуклеозидах, которые содержат пентофуранозильный сахар, фосфатная группа может быть связана по 2′, 3′ или 5′ гидроксильным радикалам сахара. При формировании олигонуклеотидов, фосфатные группы соединяются ковалентными связями соответствующие нуклеозиды друг с другом, образуя линейное полимерное соединение. В свою очередь, соответствующие концы этой линейной полимерной структуры могут в дальнейшем соединяться между собой, формируя циклическую структуру. Как правило, предпочтение отдается открытым линейным структурам. В составе олигонуклеотидной структуры фосфатные группы, как правило, рассматривают как формирующие остов олигонуклеотида. Нормальная связь или остов РНК и ДНК представляет собой 3′-5′ фосфодиэфирную связь. Специфические образцы предпочтительных нуклеосновных (нуклеотидных) олигомеров, которые могут быть использованы в данном изобретении, включают олигонуклеотиды с модифицированными остовами или искусственными межнуклеозидными связями. По определению, данному в настоящей спецификации, нуклеосновные олигомеры с модифицированным остовом включают такие (возможно модифицированные) нуклеотиды, которые сохранили в своем остове атом фосфора, и нуклеотиды, не имеющие в остове этого атома. В соответствии с целями, определенными в настоящем описании, модифицированные олигонуклеотиды, не имеющие атома фосфора в нуклеозидном остове, тоже считаются нуклеотидными (нуклеосновными) олигомерами. Нуклеотидные олигомеры с модифицированным остовом включают, например, фосфоротиоаты, хиральные фосфоротиоаты, фосфородитиоаты, фосфотриэфиры, аминоалкил-фосфотриэфиры, метил- и другие алкильные фосфонаты, включая 3′-алкилен фосфонаты и хиральные фосфонаты, фосфинаты, фосфоамидаты (фосфоамиды), включая 3′-аминофосфорамидат (амид) и аминоалкилфосфорамидаты (амиды), тионофосфорамидаты (амиды), тоноалкилфосфонаты, тионоалкилфосфотриэфиры и боранофосфаты с нормальными 3′-5′ связями, их аналогами с 2′-5′ связями и боранофосфаты с инверсной полярностью, где соответствующие пары нуклеозидных звеньев связаны между собой 3′-5′ к 5′-3′ или 2′-5′ к 5′-2′. Сюда включены также различные соли, смешанные соли и свободные кислоты. Образцы патентов США, в которых даются указания по приготовлению вышеупомянутых фосфорсодержащих связей соединений включают, но не ограничиваются Патентами США 3,687, 808; 4,469, 863; 4,476, 301; 5,023, 243; 5,177, 196; 5,188, 897; 5,264, 423; 5,276, 019; 5,278, 302; 5,286, 717; 5,321, 131; 5,399, 676; 5,405, 939; 5,453, 496; 5,455, 233; 5,466, 677; 5,476, 925; 5,519, 126; 5,536, 821; 5,541, 306; 5,550, 111; 5,563, 253; 5,571, 799; 5,587, 361; и 5,625, 050, каждый из которых включен путем ссылки. У нуклеотидных олигомеров с модифицированными олигонуклеотидными остовами, не включающими атом фосфора, сам остов сформирован короткоцепочечными алкильными или циклоалкильными межнуклеозидными связями, сочетанием гетероатома и алкила, либо циклоалкильными межнуклеозидными связями, одной или несколькими короткоцепочечными гетероатомными или гетероциклическими межнуклеозидными связями. Указанные олигомеры включают олигомеры с морфолиновыми связями (образованными из гликозидной части нуклеозида), силоксановые остовы; сульфидные, сульфоксидные и сульфоновые остовы; формацетиловые и тиоформацетиловые остовы; метилен формацетиловые и тиоформацетиловые остовы, алкен-содержащие остовы, сульфаматные остовы, метиленеминовые и метиленгидразиновые остовы; сульфонатные и сульфонамидные остовы, амидные остовы и другие остовы, состоящие из комплексных компонентов N, О, S и СН2. Образцы патентов США, в которых описывается процесс создания вышеупомянутых олигонуклеотидов, представлены патентами США В других нуклеотидных олигомерах, полисахарид и межнуклеозидная связь, т.е. остов, замещены новыми группами. Основания в цепи олигонуклеотидов сохраняют, чтобы обеспечить гибридизацию с белком IAP. Одим из таких нуклеотидных олигомеров представляет собой пептидную нуклеиновую кислоту (ПНК). В соединениях ПНК полисахаридный остов олигонуклеотида замещен амидсодержащим остовом, в частности аминоэтилглициновым остовом. Основания сохранены и соединены прямым или непрямым образом с атомами аза-азота амидной части остова. Способы получения и использования этих олигомеров описаны, например, в работе «Пептидные нуклеиновые кислоты: протоколы и приложения» (“Peptide Nucleic Acids: Protocols and Applications”) Ed. P. E. Nielsen, Horizon Press, Norfolk, United Kingdom, 1999. Образцы патентов США, в которых описывается получение ПНК, включая помимо прочего патенты США В конкретных примерах реализации изобретения нуклеотидные олигомеры имеют фосфоротиоатные остовы и нуклеозиды с остовом, включающим гетероатом, в частности -CH2-NH-O-CH2-, -СН2-(СН3)-O-СН2– (известный как метиленовый(метилиминовый) или MMI-остов), -CH2-O-N(CH3)-CH2-, -CH2-N(CH3)-N(CH3)-CH2 и -O-N(CH3)-CH2-CH2-. В других примерах реализации настоящего изобретения, олигонуклеотиды имеют остовы, в структуру которых входит морфолин, описанные в патенте США Нуклеотидные олигомеры могут также содержать одну или несколько замещенных полисахаридных частей. Нуклеотидные олигомеры в положении 2′ содержат один из следующих фрагментов: ОН; F; O-, S-, или N-алкил; О, S-, или N-алкенил; S- или N-алкинил либо О-алкил-O-алкил, где алкил, алкенил и алкинил могут замещаться или не замещаться на алкил С1-С10, или алкенил и алкинил С2-С10. Особенно предпочтительными являются O[(СН2)nО]mСН3, O(СН2)nОСН3, O(CH2)nNH2, O(СН2)nСН3, O(CH2)nONH2 и (CH2)nON[(CH2)nCH3)]2, где n и m – числа от 1 до 10. Другие предпочтительные нуклеотидные олигомеры содержат в положении 2′ один из следующих фрагментов: С1-С10 низший алкил, замещенный низший алкил, алкарил, аралкил, O-алкарил или O-аралкил, SH, SСН3, OCN, Cl, Br, CN, СF3, ОСF3, SОСН3, NH2, гетероциклоалкил, гетероциклоалкарил, аминоалкиламин, полиалкиламин, замещенный силил, группу, расщепляющую РНК, репортерную группу, интеркалирующий агент, группу, улучшающую фармакокинетические свойства нуклеотидного олигомера и другие заместители, обладающие подобными свойствами. Предпочтительными модификациями являются 2′-O-метил и 2′-метоксиэтоксин (2′-O-СН2СН2OСН3), известные также, как 2′-O-(2-метоксиэтил) или 2′-МОЕ). Другая предпочтительная модификация представляет собой 2′-диметиламинооксиэтокси (например, O(СН2)2ON(СН3)2), известный также, как 2′-DMAOE. Другие модификации включают 2′-аминопропокси (2′-OCH2CH2CH2NH2) и 2′-фтор (2′-F). Подобные модификации могут быть образованы в других положениях олигонуклеотида или другого нуклеосновного олигомера, в частности по положению 3′ полисахарида в положении 3′ конечного нуклеотида или в положении 2′-5′ связанных олигонуклеотидов и положении 5′ конечного нуклеотида. Патенты США, которые описывают создание таких структур с модифицированным полисахаридом, включают помимо прочего, патенты США Нуклеотидные олигомеры могут также содержать модифицированные или замещенные нуклеотиды. В контексте настоящего изобретения «неизмененные» или «естественные» нуклеотиды включают пуриновые основания аденин (А) и гуанин (G) и пиримидиновые основания Тимин(Т), цитозин (С) и урацил (U). Модифицированные нуклео-основания (нуклеотиды) включают синтетические и естественные нуклеозиды, такие как 5-метилцитозин (5-me-С), 5-гидроксиметил цитозин, ксантин, гипоксантин, 2-аминоаденин, 6-метил и другие алкиловые производные аденина и гуанина; 2-пропил и другие алкиловые производные аденина и гуанина; 2-тиоурацил, 2-тиотимин и 2-тиоцитозин; 5-пропинилурацил и цитозин; 6-азо урацил, цитозин и тимин; 5-урацил (псевдоурацил); 4-тиоурацил; 8-гало, 8-амино, 8-тиол, 8-тиоалкил, 8-гидроксил и другие 8-замещенные аденины и гуанины; 5-гало (например, 5-бромо), 5-трифторметил и другие 5-замещенные урацилы и цитозины; 7-метилгуанин и 7-деазааденин; 3-деазагуанин и 7-деазааденин. Другие нуклеотиды включают, в частности раскрытые в патенте США Другая модификация нуклеотидного олигомера, согласно настоящему изобретению, включает химическое присоединение к нуклеотидному олигомеру одной или нескольких компонентов или конъюгатов, которые улучшают активность, распределение в клетке или попадание олигонуклеотидов в клетку. Такие части включают, помимо прочего, липидные компоненты, например такие, как холестериновая часть (Letsinger et al., Proc. Natl. Acad. Sci. USA, 86:6553-6556, 1989), холевая кислота (Manoharan et al.., Bioorg. Med. Chem. Let, 4:1053-1060, 1994), тиоэфир, например гексил-S-тритилтиол (Manoharan et al., Ann. N.Y. Acad. Sci., 660:306-309, 1992; Manoharan et al.., Bioorg. Med. Chem. Let., 3:2765-2770, 1993), тиохолестерол (Oberhauser et al., Nucl. Acids Res., 20:533-538: 1992), алифатическая цепь, например, додекандиоловый или ундециловый остатки (Saison-Behmoaras et al., EMBO J., 10:1111-1118, 1991; Kabanov et al., FEBS lett, 259:327-330, 1990; Svinarchuk et al., Biochimie, 75:49-54, 1993), фосфолипид, например, ди-гексадецил-рак-глицерол или триэтиламмоний 1,2-ди-О-гексадецил-рак-глицеро-3-Н-фосфонат (Manoharan et al., Tetrahedron Lett., 36:3651-3654, 1995; Shea et al., Nucl. Acids Res., 18:3777-3783, 1990), полиамин или полиэтиленгликолевая цепь (Manoharan et al., Nucleosides & Nucleotides, 14: 969-973,1995) или адамантан-уксусная кислота (Manoharan et al., Tetrahedron Lett., 36:3651-3654, 1995), пальмитиловая часть (Mishra et al., Biochim. Biophys. Acta, 1264:229-237, 1995), октадециламин или гексиаламино-карбонил-оксихолестероловая группа (Crooke et al., J. Pharmacol. Exp. Ther., 277:923-937, 1996.) Примерами патентов США, описывающих получение таких конъюгатнуклеотидных, являются патенты США Настоящее изобретение также охватывает нуклеотидные олигомеры, представляющие собой химерные соединения. «Химерные» нуклеотидные олигомеры – это нуклеосновные олигомеры, в частности олигонуклеотиды, содержащие два или более химически различных участка, каждый из которых состоит, по меньшей мере, из одного мономерного блока, т.е. нуклеотида, если речь идет об олигонуклеотиде. Эти нуклеоосновные олигомеры, как правило, содержат, по меньшей мере, один участок, где олигонуклеотид модифицирован таким образом, что в результате олигонуклеотид приобретает, повышенную резистентность к расщеплению к действиям нуклеазы, улучшенную способность проникать в клетку, и/или повышенное сродство к связыванию с нуклеиновой кислотой-мишенью. Дополнительный участок нуклеотидного олигомера может служить субстратом для ферментов, расщепляющих РНК:ДНК или РНК:РНК гибриды. Например, РНКаза Н является клеточной эндонуклеазой, которая расщепляет нить РНК или РНК-ДНК дуплекс. Активация РНКазы, следовательно, приводит к расщеплению РНК-мишени, таким образом, значительно усиливая эффективность ингибирования экспрессии гена нуклеотидным олигомером. Как следствие, аналогичные результаты можно получить с использованием более коротких олигомеров, в случае если используют химерные нуклеотидные олигомеры, по сравнению с фосфоротиоатными дезоксиолигонуклеотидами гибридизирующимися с тем же самым участком-мишенью. Химерные нуклеотидные олигомеры согласно настоящему изобретению могут быть сформированы как сложные структуры, состоящие из одного и более нуклеотидных олигомеров, как описано выше. Такие нуклеосновные олигомеры, в частности олигонуклеотиды, также известны как гибриды или «гапмеры» (gapmers). Получение таких гибридных структур описано в патентах США Нуклеотидные олигомеры согласно настоящему изобретению могут быть получены общепринятым методом твердофазного синтеза. Оборудование для такого синтеза поставляется несколькими фирмами, в числе которых, например, Applied Biosystems (Foster City, CA). Для проведения указанного синтеза могут быть дополнительно или в качестве альтернативы использованы любые, известные в этой области средства. Широко известно применение аналогичных методик для получения олигонуклеотидов, например фосфоротиоатов и алкильных производных. Нуклотидные олигомеры, относящиеся к настоящему изобретению, могут также быть смешаны, инкапсулированы, конъюгированы или каким-либо другим способом связаны с другими молекулами, молекулярными структурами или смесями химических соединений, например такими, как липосомы, рецептор-аффинные молекулы, оральные, ректальные, местные и прочие лекарственные формы, способствующие проникновению, распределению и/или абсорбции. Получение таких лекарственных средств, способствующих проникновению, распределению и абсорбции, описаны в патентах США Нуклеотидные олигомеры согласно настоящему изобретению включают любые фармацевтически приемлемые соли, эфиры, или соли этих эфиров или любое другое вещество, способное образовать (прямым или косвенным способом) их биологически активный метаболит или радикал. Соответственно, область охвата настоящего изобретения также можно распространить на пролекарства и фармацевтически премлемые соли веществ, которые относятся к настоящему изобретению и другие биологические эквиваленты. Термин «пролекарство» относится к терапевтическому агенту, который готовят в неактивной форме и который претерпевает превращение в активную форму (т.е. лекарство) внутри организма или клетки под воздействием эндогенного белка или химических веществ и/или условий. В частности, пролекарства олигонуклеотидов согласно настоящему изобретению могут быть приготовлены в виде SATE- [(S-ацетил-2-тиоэтил) фосфатных] производных согласно способам, изложенным в заявках WO 93/24510 и WO 94/26764. Термин «фармацевтически приемлемая соль» относится к солям, которые сохраняют желательную биологическую активность исходного (родоначального) соединения и при этом не оказывают нежелательных токсических эффектов. Фармацевтически премлемые основно-аддитивные соли образованы с металлами или аминами, например, соли щелочных и щелочноземельных металлов или органических аминов. Примерами металлов, используемых в качестве катионов, могут служить натрий, калий, магний, кальций и им подобные. Примерами аминов – N, N’-дибензилэтилендиамин, хлоропрокаин, холин, диэтаноламин, дициклогексиламин, этилендиамин, N-метилглюкамин и прокаин (см., например, Berge et al., J.Pharma Sci., 66:1-19, 1977). Основно-аддитивные соли кислотных соединений получают традиционным способом путем взаимодействия свободной кислоты с достаточным количеством соответствующего основания для получения соли. Обратно форму свободной кислоты можно получить путем контактирования соли с соответствующей кислотой и выделения свободной кислоты известными способами. Свободные кислоты отличаются от соответствующих солей некоторыми конкретными физическими свойствами, такими как, например, растворимость в полярных растворителях, но в то же время соли эквивалентны соответствующим свободным кислотам в отношении целей настоящего изобретения. В данном контексте «фармацевтическая соль присоединения» включает фармацевтически приемлемую соль одного из компонентов состава, относящегося к настоящему изобретению, в форме кислоты. Такие соли включают органические или неорганические кислые соли аминов. Предпочтительными кислыми солями являются гидрохлориды, ацетаты, салицилаты, нитраты и фосфаты. Другие подходящие фармацевтически приемлемые соли хорошо известны специалистам в этой области и включают основные соли разнообразных неорганических и органических кислот, таких как, например, из неорганических: соляная кислота, бромоводородная кислота, серная кислота или фосфорная кислота; из органических – карбоновые, сульфо- или фосфо-кислоты и N-замещенные сульфаминовые кислоты, например, уксусная кислота, пропионовая кислота, гликолевая кислота, сукциниловая кислота, малеиновая кислота, гидроксималеиновая кислота, метилмалеиновая кислота, фумаровая кислота, яблочная кислота, тартаровая кислота, молочная кислота, щавелевая кислота, гликаровая кислота, глюконовая кислота, глюкуроновая кислота, лимонная кислота, бензойная кислота, коричная кислота, миндальная кислота, салициловая кислота, 4-амино-салициловая кислота, 2-феноксибензойная кислота, 2-ацетоксибензойная кислота, эмбоновая кислота, никотиновая или изоникотиновая кислота; или аминокислоты, такие как 20-альфа-аминокислоты, участвующие в синтезе белков в природе, например, глютаминовая кислота или аспарагиновая кислота, и фенолуксусная кислота, метансульфоновая кислота, этансульфоновая кислота, 2-гидроксиэтансульфоновая кислота, этан-1,2-дисульфоновая кислота, бензолсульфоновая кислота, 4-метилбензолсульфоновая кислота, нафталин-2-сульфоновая кислота, 5-дисульфоновая кислота, 2- или 3-фосфоглицериновая кислота, глюкоза-6-фосфат, N-циклогексилсулфамовая кислота (с формированием цикламатов), или с другими кислотными органическими соединениями, такими как аскорбиновая кислота. Фармацевтически приемлемые соли указанных соединений также можно приготовить с использованием фармацевтически приемлемых катионов. Подходящие фармацевтически примелемые катионы хорошо известны специалистам и включают щелочные, щелочноземельные катионы, катионы аммония и четвертичного аммония. Возможно также использование карбонатов или гидрокарбонатов. Для олигонуклеотидов и других нуклеотидных олигомеров фармацевтически приемлемые соли включают (i) соли, образованные катионами, такими как натрий, калий, азот, магний, кальций, полиамины, например спермин и спермидин и т.д.; (ii) кислотно-аддитивные соли, образованные неорганическими кислотами, например соляной кислотой, бромоводородной кислотой, серной кислотой, фосфорной кислотой, азотной кислотой и им подобными; (iii) соли, образованные органическими кислотами, такими как, например уксусная кислота, оксалиновая кислота, тартаровая кислота, сукциниловая кислота, малеиновая кислота, фумаровая кислота, клюконовая кислота, лимонная кислота, муравьиная кислота, аскорбиновая кислота, бензойная кислота, дубильная кислота, пальмитиновая кислота, альгиновая кислота, полиглютаминовая кислота, нафталенсульфониевая кислота, метансульфониевая кислота, р-толуенсульфониевая кислота, полигалактуроновая кислота и им подобные; и (iv) соли, образованные элементарными анионами, например анионами хлора, брома и йода. Настоящее изобретение также охватывает фармакологические составы и препараты, в состав которых входят нуклеотидные олигомеры согласно настоящему изобретению. Фармакологические препараты согласно настоящему изобретению могут вводиться несколькими путями, в зависимости от площади, подлежащей лечению и от того, какое требуется лечение – системное или местное. Препарат можно вводить местно (включая глазное введение и введение через слизистые (в том числе вагинальный и ректальный пути введения), такими путями как: легочный, например посредством ингаляций или вдыхания порошков или аэрозолей посредством распылителя; интратрахеальный и интраназальный, эпидермальный и трансдермальный) оральный или парентеральный способы. Парентеральное введение включает внутривенные, интраартериальные, подкожные, интраперитонеальные или внутримышечные инъекции и инфузии; или интракраниальное введение, например интратекальное или интравентрикулярное. Фиксированные нуклеиновые кислоты Фиксированные нуклеиновые кислоты (Locked Nucleic acids, LNA) представляют собой нуклеотидные олигомеры, которые тоже могут быть использованы согласно настоящему изобретению. LNA содержат в позициях 2′-O, 4′-С метиленовые мостики, которые ограничивают гибкость рибофуранозного кольца аналога нуклеотида и фиксируют его в ригидной бициклической конформации N-типа. LNA проявляют повышенную устойчивость к определенным экзо- и эндонуклеазам, активируют РНК-азу Н и могут быть включены практически в любой нуклеотидный олигомер. Более того, LNA-содержащие нуклеотидные олигомеры могут быть получены с помощью стандартных протоколов синтеза фосфорамидитных соединений. Дополнительную информацию относительно LNA можно найти в публикациях РСТ Арабинонуклеиновые кислоты Арабинонуклеиновые кислоты также могут быть задействованы в способах и реактивах согласно настоящему изобретению. ANA представляют собой нуклеотидные олигомеры, имеющие в своем составе сахар D-арабинозу вместо естественной D-2′-дезоксирибозы. Немодифицированные аналоги ANA обладают сродством к РНК, аналогичным фосфоротиоатам. При модификации арабинозы из фтором (2’F-ANA), повышается сродство к связыванию и происходит более эффективный селективный гидролиз связанной РНК в дуплексах ANA/PHK и F-ANA/PHK. Эти аналоги могут быть стабилизированы внутри клетки с помощью присоединения на их концах простых L-сахаров. Использование ANA в терапии в настоящее время обсуждается, например, у Damna et al., Nucleosides & Nucleic Acids 20: 429-440, 2001. Способы доставки нуклеотидных олигомеров Согласно настоящему изобретению чистые олигонуклеотиды способны проникать в опухолевые клетки и ингибировать экспрессию IAP. Однако было бы желательно использовать состав, который способствует доставке олигонуклеотида или других нуклеотидных олигомеров к клеткам (см., например, патенты США Рибозимы Каталитические молекулы РНК или рибозимы, которые содержат антисмысловую последовательность IAP согласно настоящему изобретению можно использовать для ингибирования экспрессии полинуклеотида IAP in vivo. Включение последовательностей рибозимов в антисмысловые РНК сообщает им РНК-расщепляющую активность, увеличивая, таким образом, активность всей конструкции. Разработка и использование РНК-специфичных рибозимов описаны у Haseloff et al., Nature 334:585-591, 1988 и публикациях патентных заявок США Изобретение также включает каталитическую молекулу РНК, которая в связующем сегменте содержит антисмысловую РНК, насчитывающую от восьми до девятнадцати последовательно идущих нуклеотидов, соответствующих любой из последовательностей, приведенных в Таблицах 1, 2, 3 и 7. В предпочтительных вариантах реализации данного изобретения каталитические молекулы нуклеиновой кислоты обладают структурой типа «головки молотка» (hammerhead), но допустима, также, структура типа «шпильки» (hairpin), дельта-вируса гепатита, интрона группы 1 или RN-азыР РНК (связанной с напрявляющей последовательностью РНК) и структур типа Neurospora VS РНК. Примеры таких структур типа «головки молотка» описаны в Rossi et al., AIDS Research and Human Retroviruses, 8:183, 1992. Примеры структур типа «шпильки головки» описаны в патентах США Интерференция РНК Нуклеотидные олигомеры согласно настоящему изобретению могут быть включены в двойную спираль молекул РНК для нокдауна экспрессии IАР, опосредованного интерференцией РНК (PHK-i). PHK-i представляет собой способ снижения клеточной экспрессии индифидуальных белков (см. обзор у TuschI, Chembiochem 2: 239-245, 2001; Sharp, Genes & Devel. 15: 485-490, 2000; Hutvagner and Zamore, Curr. Opin. Genet. Devel. 12: 225-232, 2002; and Hannon, Nature 418: 2002). При PHK-i подавление экспресии гена (silencing), как правило, осуществляется после процесса транскрипции благодаря присутствию в клетке двунитевой РНК (dsPHK). Эта dsPHK внутри клетки расщепляется на более короткие фрагменты, называемые siPHK. Введение siPHK в клетку либо путем трансфекции dsPHK, либо путем экспрессии siPHK с помощью плазмидной системы экспрессии все шире используется для вариантов фенотипа с отсутствующей функциональностью (loss-of-function phenotypes) в клетках млекопитающих. В одном способе реализации настоящего изобретения двунитевая молекула РНК (dsPHK) содержит последовательность, состоящую из восьми-девятнадцати последовательных нуклеотидов нуклеотидного олигомера согласно настоящему изобретения. Молекула dsPHK может представлять собой две отдельные нити РНК, которые образуют дуплекс, либо единственную нить РНК, образующую дуплекс с самой собой (тип «маленькой шпильки» (sh РНК)). Как правило, dsPHK состоят из 21-22 пар нуклеотидов, но могут быть, при желании, короче или длиннее (до 29 нуклеотидов). dsPHK можно получить с помощью стандартных технологий (например, путем химического синтеза или in vivo путем транскрипции). Имеются коммерчески доступные наборы, предоставляемые, например, Ambion (Austin, ТХ) и Epicentre (Madison, WI). Способы экспрессирования dsPHK в клетках млекопитающих описаны у Brummelkamp et al. Science 296: 550-553, 2002; Paddison et al. Genes & Devel. 16: 948-958, 2002; Paul et al. Nature Biotechnol. 20: 505-508, 2002; Sui et al. Proc. Natl. Acad. Sci. USA 99: 5515-5520, 2002; Yu et al. Proc. Natl. Acad. Sci. USA 99: 6047-6052, 2002; Miyagishi et al. Nature Biotechnol. 20: 497-500, 2002; and Lee et al. Nature Biotechnol. 20: 500-505, 2002, каждый из которых включен путем ссылки. shPHK имеют структуру «ствол-петля», возможно с 3′ UU-выступами. Несмотря на возможные вариации, размер самого «ствола» составляет от 21 до 31 bp (предпочтительно от 25 до 29 bp). Для внутриклеточной экспрессии shPHK в клетку вводят плазмидный вектор, содержащий промотор полимеразы III H1-РНК, tPHK или U6 и сайт клонирования для встраивания последовательности РНК, образующей структуру «ствол-петля», а в качестве сигнала терминации транскрипции можно использовать – 4-5-тимидин. Промоторы полимеразы III, как правило, обладает четко определяемыми старт- и стоп-сайтами, а у их транскриптов отсуствуют сегменты poly (A). Сигналы терминации для этих промоторов содержат участок политимидина, а транскрипт, как правило, расщепляется после второго уридина. Расщепление в этом сайте формирует 3’UU выступ в экспрессированной sh РНК, который идентичен 3′ выступам синтезированных молекул siPHK. Дополнительные способы экспрессирования shPHK в клетках млекопитающих описаны в ссылках, приведенных ниже. Химиотерапевтические препараты Применение нуклеотидных олигомеров в комбинации с одним или несколькими химиотерапевтическими препаратами, оказывающими разрушающее или стабилизирующее действие на микротрубочки, особенно эффективно при лечении рака и других злокачественных новообразований. Препараты, разрушающие микротрубочки (например, алкалоиды барвинка) и препараты, стабилизирующие микротрубочки (например, таксаны), подробно описаны ниже. Алкалоиды барвинка и родственные соединения Алкалоиды барвинка, которые можно использовать в комбинации с нуклеотидными олигомерами, согласно настоящему изобретению включают в себя винкристин, винбластин, виндезин, винфлунин, винорельбин и ангидровинбластин Доластины – это олигопептиды, которые сначала конкурируют с тубулином в домене связывания алкалоидов барвинка. Эти вещества могут также использоваться в сочетании с другими нуклеотидными олигомерами согласно настоящему изобретению для лечения рака и других новообразований. Доластины включают доластин-10 (NCS 376128), доластин-15, ILX635, TZT-1027, симплостатин 1, симплостатин 3 и LU103793 (цемадотин). Криптофицины (например, криптофицин 1 и криптофицин 52 (LY355703)) связывают тубулин в домене связывания Vinca-алкалоида и индуцируют остановку в G2/M и апоптоз. Любой из этих препаратов можно использовать в сочетании с нуклеотидным олигомером согласно настоящему изобретению для лечения рака и других новообразований. Другие препараты, разрушающие микротрубочки, которые можно использовать в сочетании с нуклеотидными олигомерами согласно настоящему изобретению для лечения рака и других новообразований, описаны в патентах США Таксаны и другие препараты, стабилизирующие микротрубочки В сочетании с нуклеотидными олигомерами согласно настоящему изобретению для лечения рака и других новообразований можно использовать такие таксаны, как паклитаксел, доксетаксел, RPR SB-T-1213, SB-T-1250, BMS-275183, BRT 216, DJ-927, IDN5109, и IDN5390. Аналоги таксана (например, BMS-184476, BMS-188797) и функционально родственные не-таксаны (такие как эпотилоны (например, эпотилон А, эпотилон В (ЕР0906), дезоксиэпотилон В и эпотилон В пактам (BMS-247550)), элеутеробин, дискодермолид, 2-эпи-дискодермолид, 2-дес-метилдискодермолид, 5-гидроксиметилдискодермолид, 19-дес-аминокарбонилдискодермолид, 9(13)-циклодискодермолид и лаулималид), также могут применяться в способах и составах согласно настоящему изобретению. Другие препараты, стабилизирующие микротрубочки, которые можно применять в сочетании с нуклеотидными олигомерами согласно настоящему изобретению для лечения рака и других новообразований, описаны в патентах США Другие химиотерапевтические препараты, которые можно вводить совместно с антисенс-нуклеотидными олигомерами IAP, перечислены в Таблице 1.
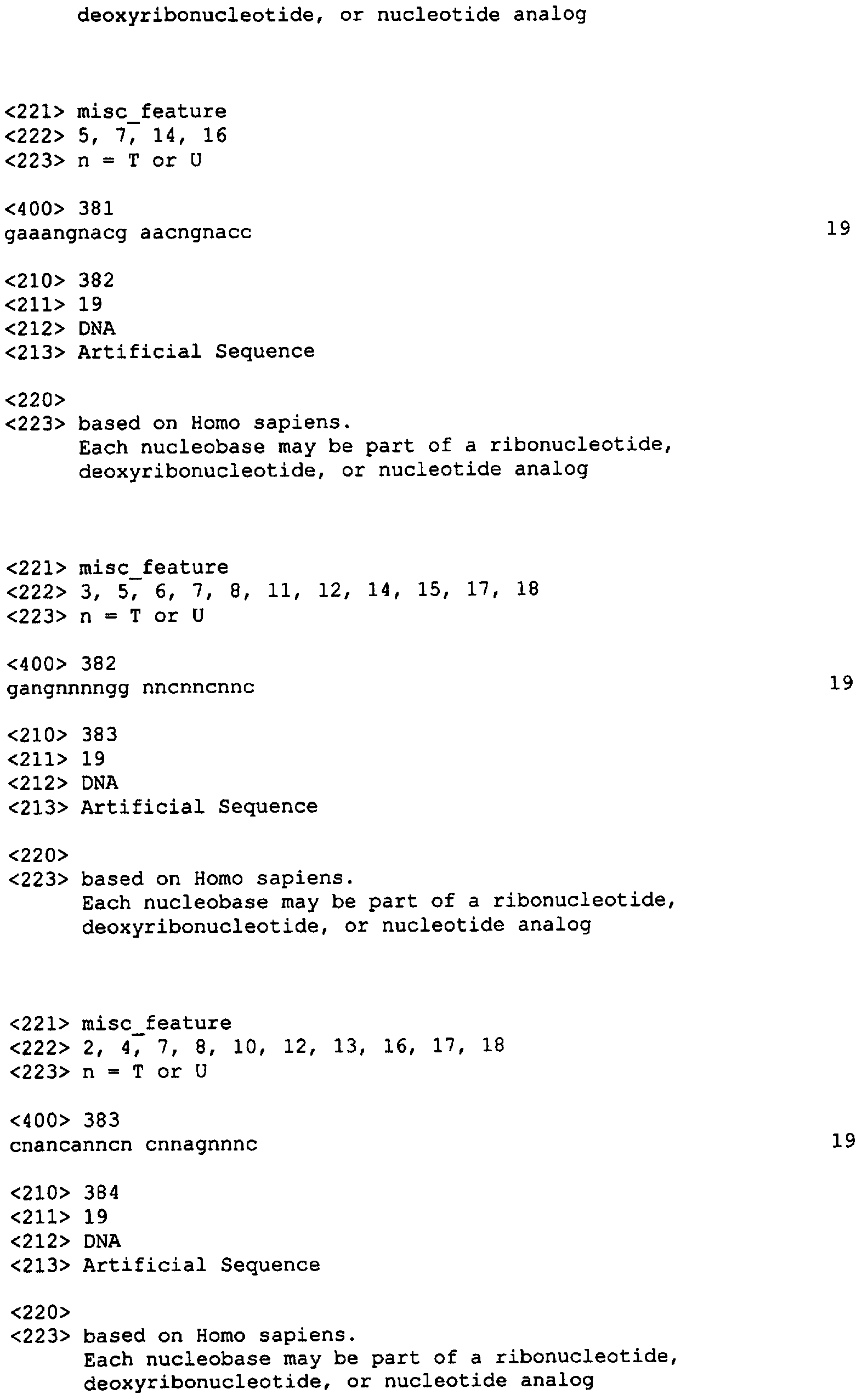
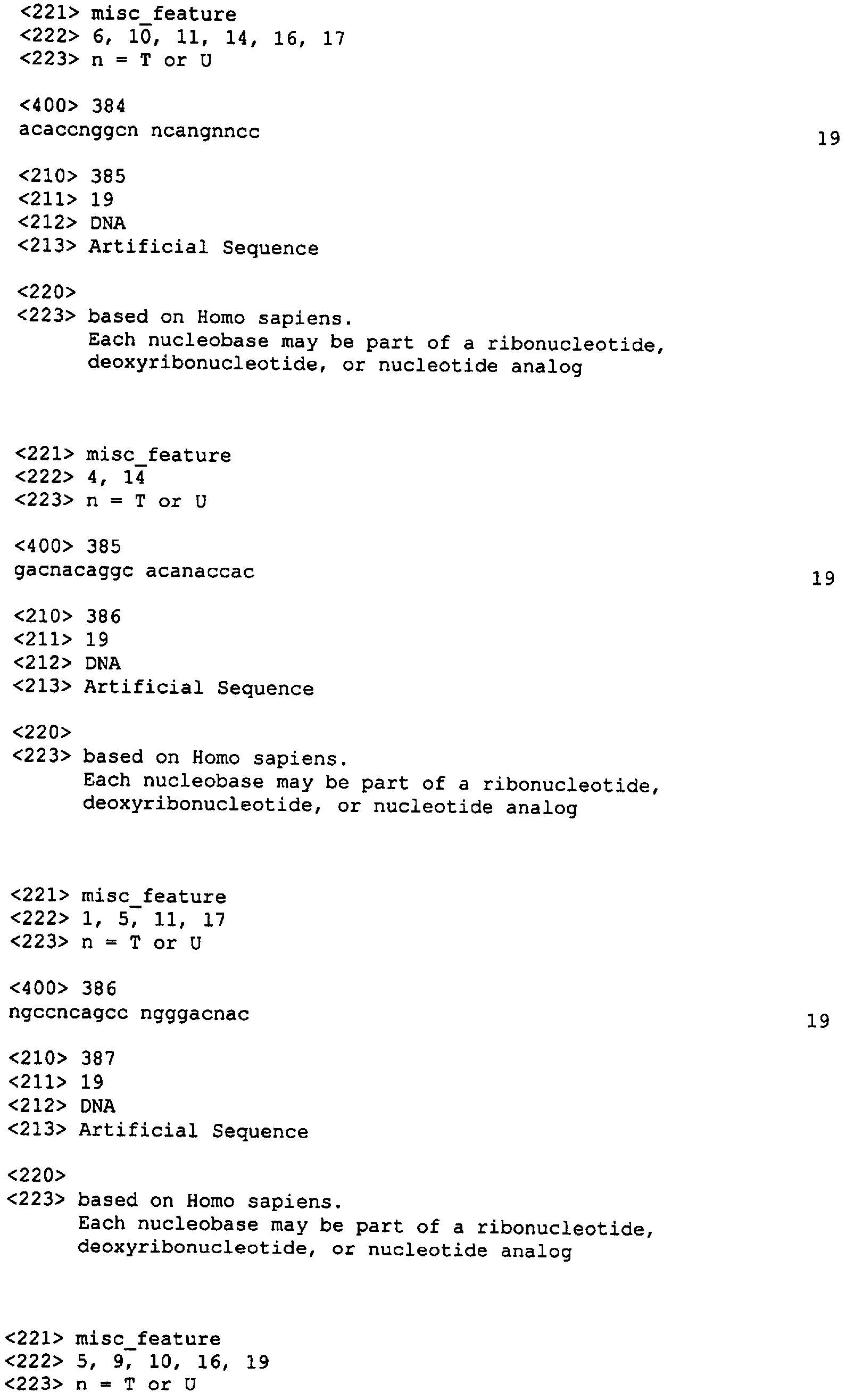
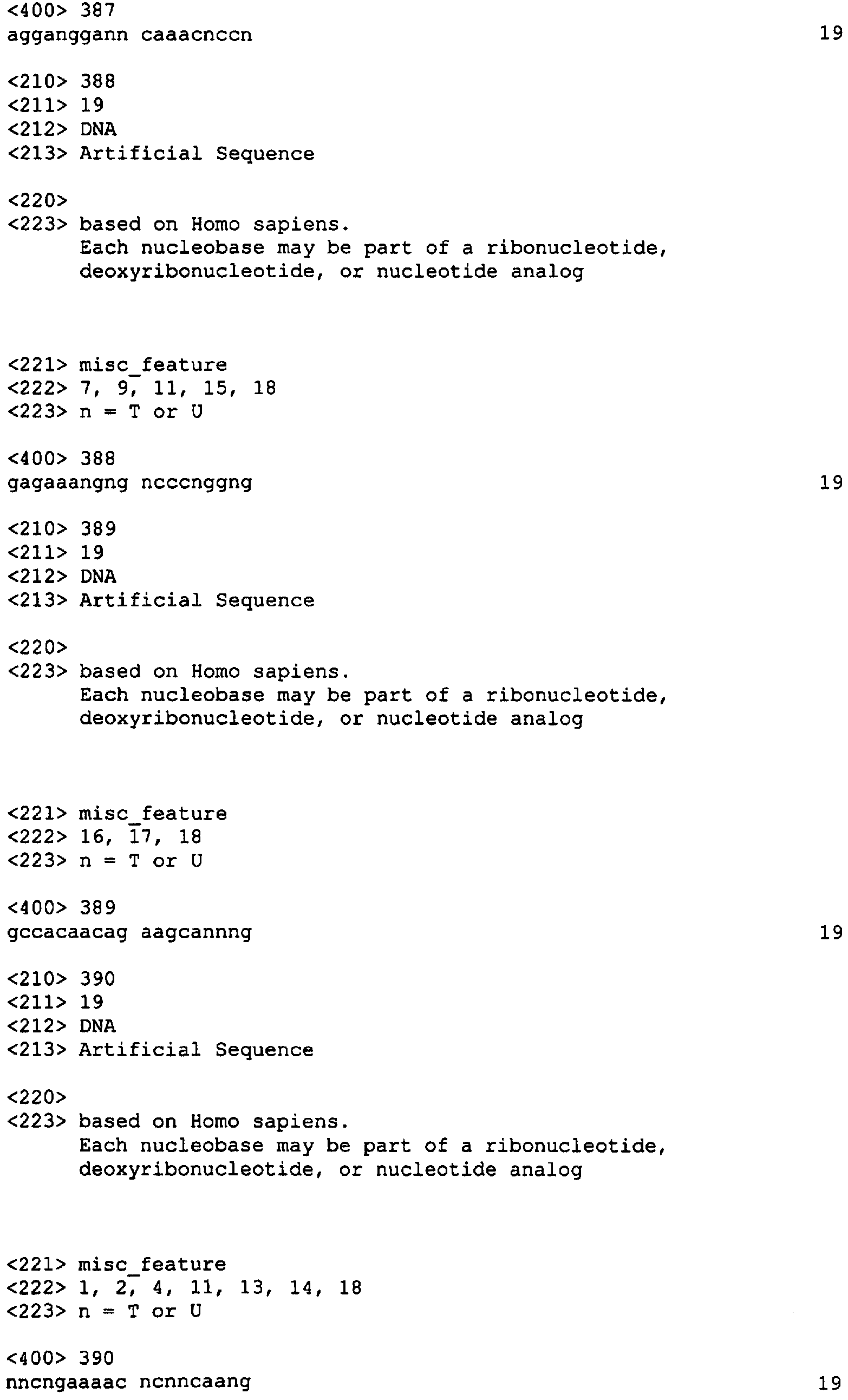
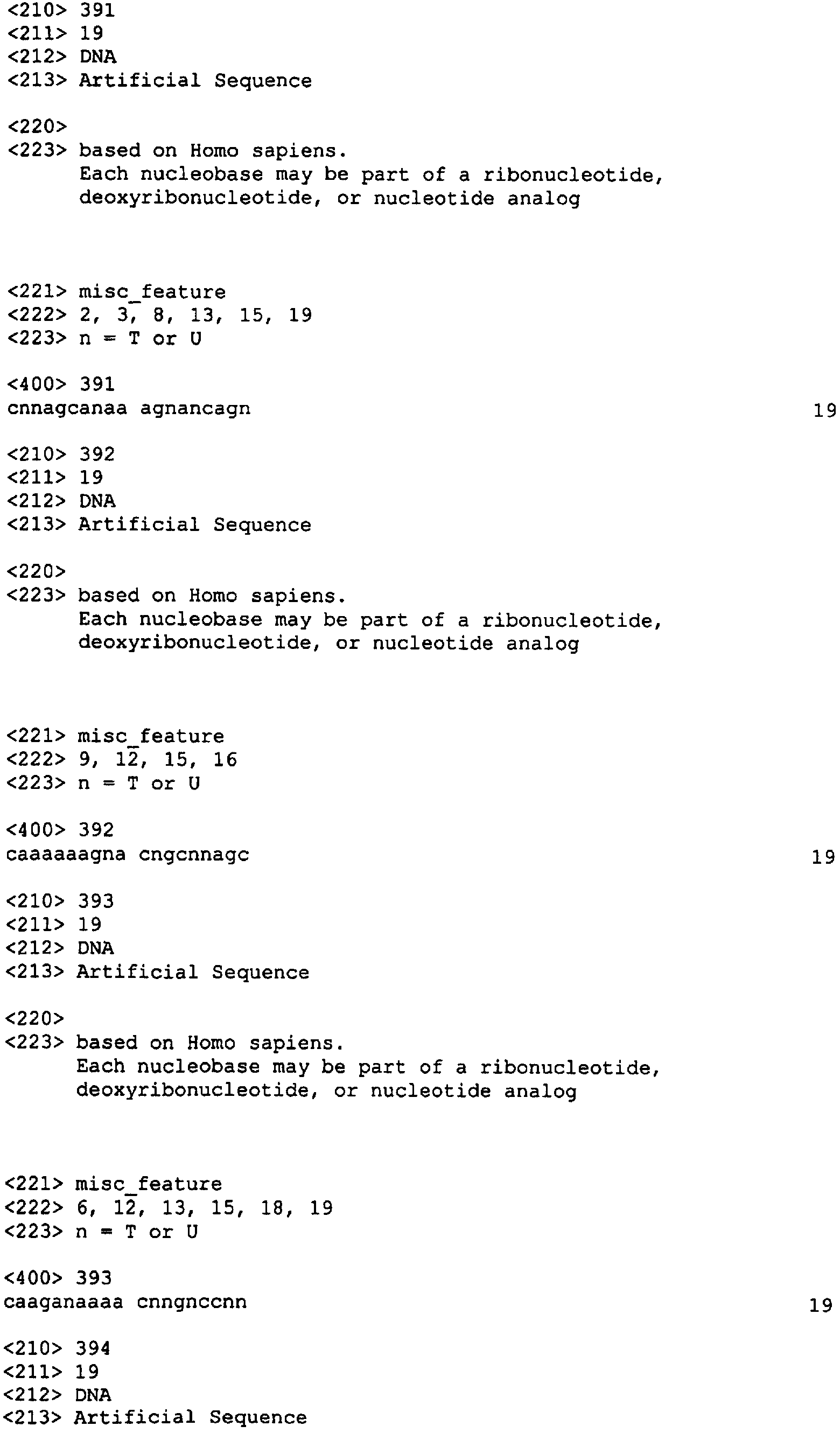
Приведенные ниже примеры служат исключительно для иллюстрации изобретения и ни в коей мере не ограничивают область охвата настоящего изобретения. Пример 1: Выбор нуклеотидного олигомера На основании нижеуказанных критериев нами было отобрано 96 и 98 неперекрывающихся последовательностей человеческого XIAP и HIAPI, соответственно из 19 нуклеотидов, в случае XIAP мы отобрали 96 последовательностей (каждая длиной в 19 нуклеотидов) (Номера последовательностей SEQ ID NO: от 1 до 96; Таблица 2) из участка последовательности, расположенные приблизительно на расстоянии около 1 кб против направления транскрипции от стартового кодона и примерно 1 кб по направлению транскрипции от стоп-кодона кДНК. Это покрыло примерно 50% кодирующей области и прилежащие последовательности 3′-UTR и 5′-UTR (например, 96 19-меров занимают 1,8 кб последовательности, в то время как целевая область составляет примерно 3,5 кб в длину и состоит из 1,5 кб кодирующей области плюс 1 кб UTR последовательностей с каждой стороны).
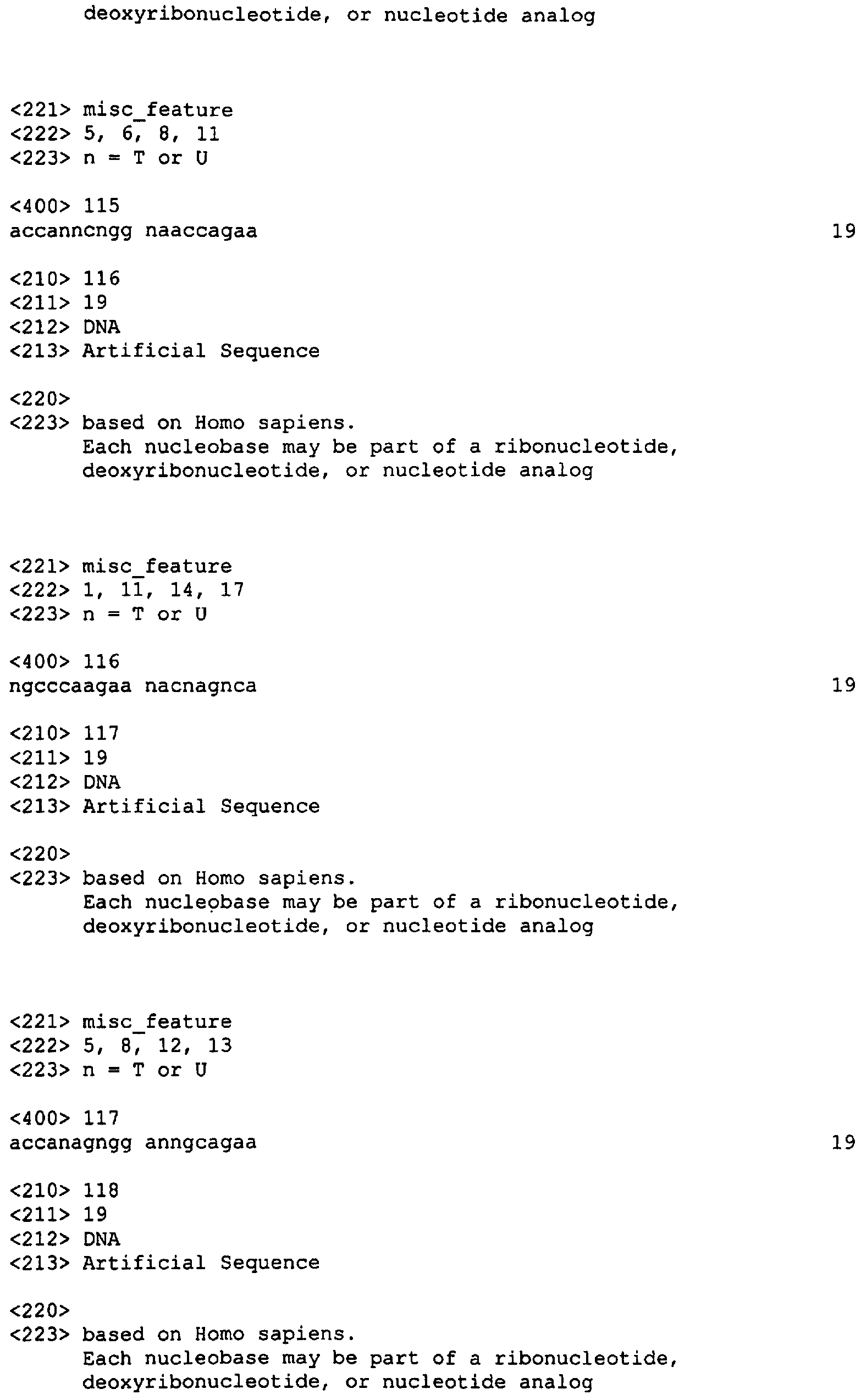
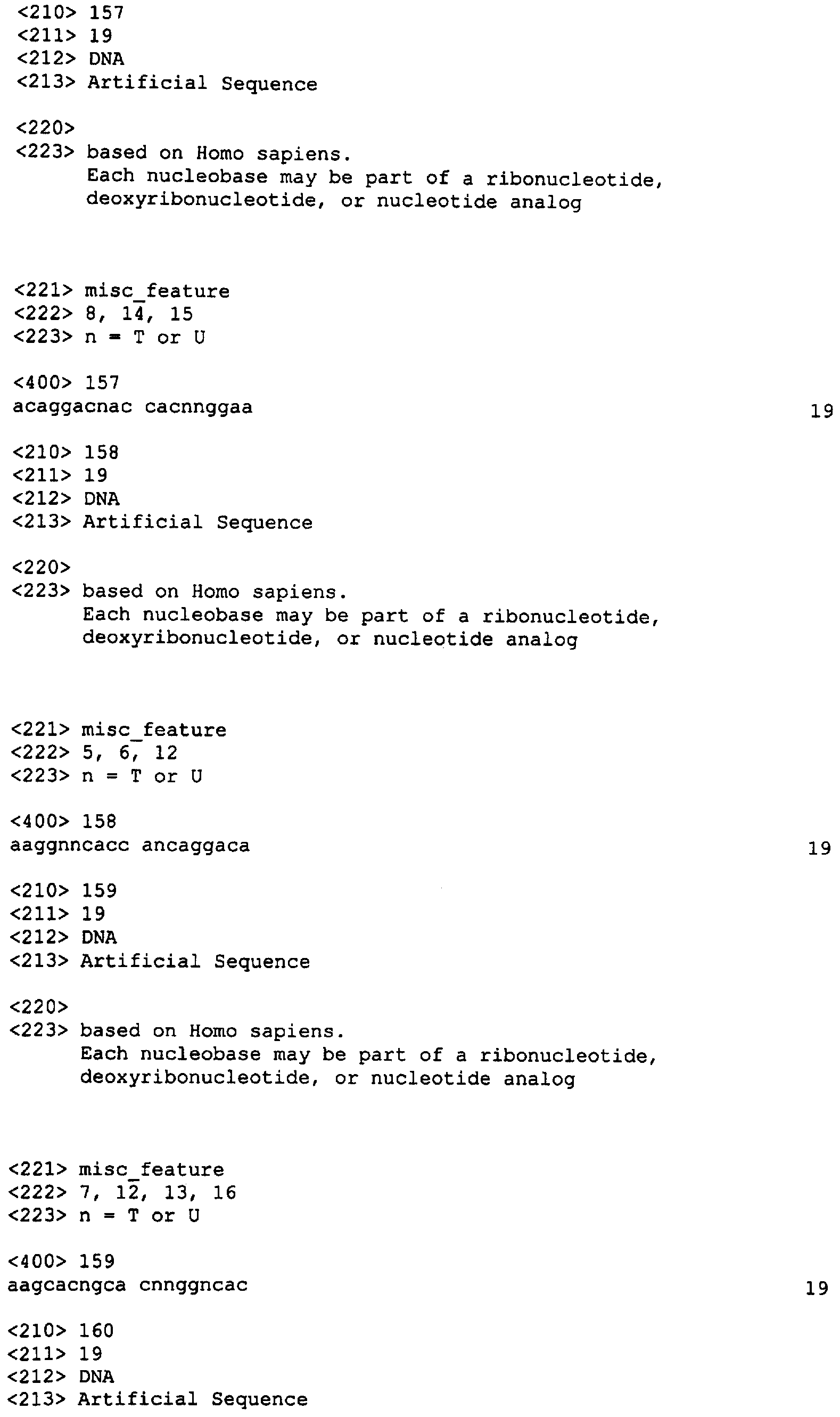
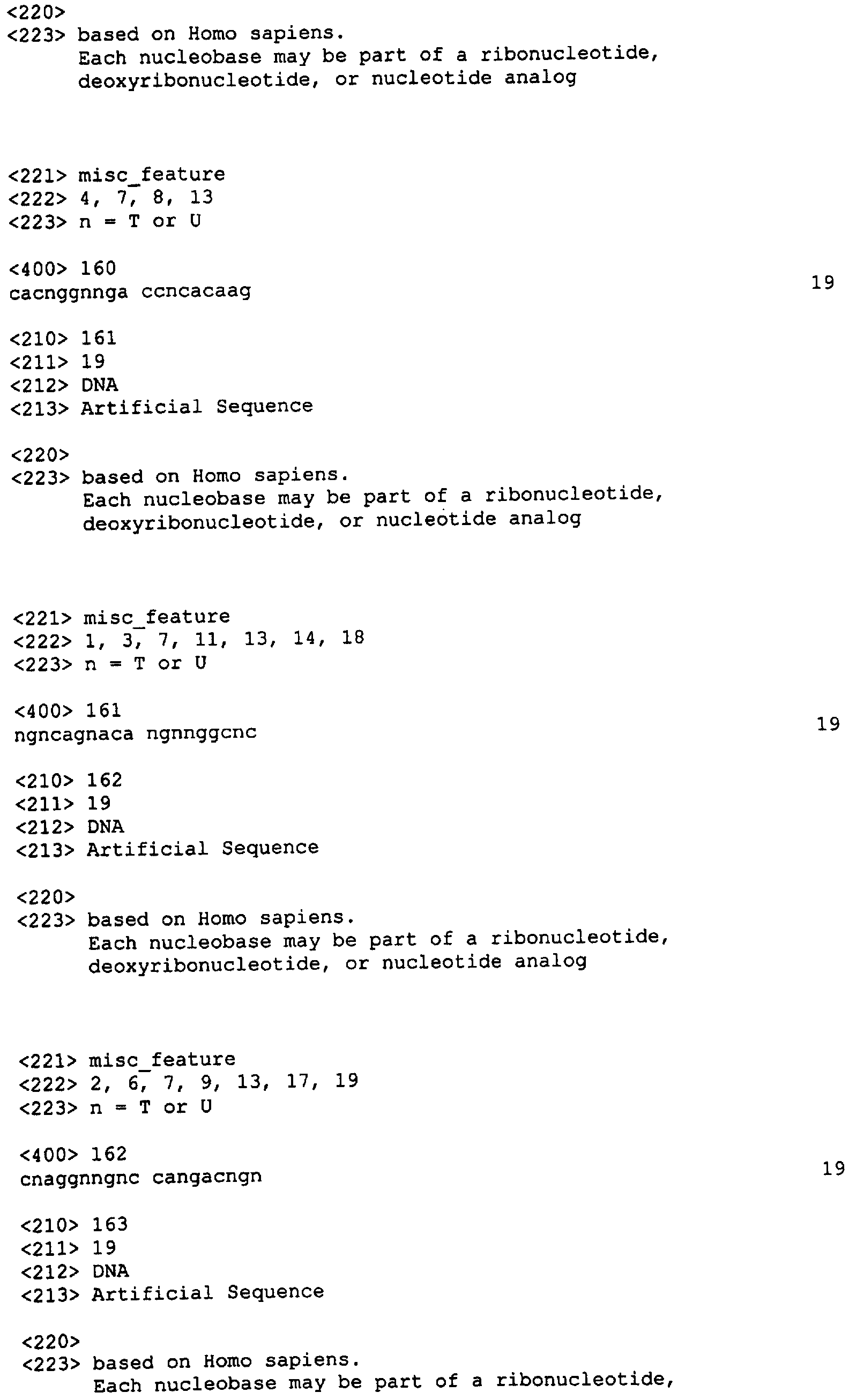
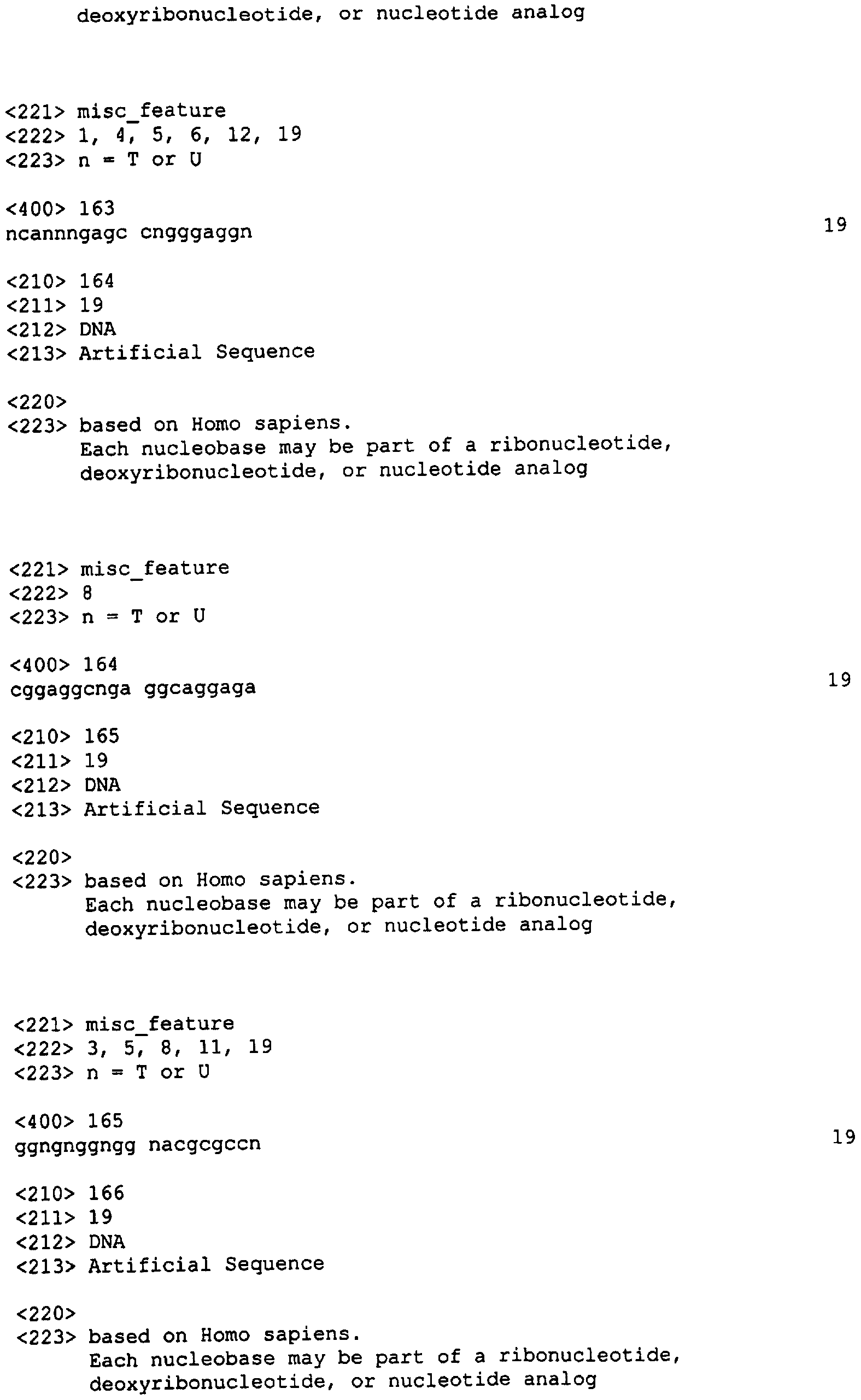
Следует заметить, что в нуклеотидных олигомерах каждый нуклеотид, указанный в данном описании изобретения, может независимо представлять собой остаток ДНК или РНК, например 2′-O-метилированный или 2′-O-метоксиэтилированный остаток РНК. Нуклеотидная последовательность SEQ ID NO: 3 может быть, например, 5′-CAGATATATATGTA АСАСТ-3′, 5′-CAGATATATATGTAA CACU-3′, или 5′-mCmAGATATATATGTAACAmCmU-3′ (где mХ представляет 2′-O-метилированный Х остаток). В данной области известны другие модифицированные нуклеотиды. Связи могут быть фосфодиэфирными (РО), фосфоротиоатными (PS) или метилфосфонатными (МР). В качестве остова может быть любой остов, позволяющий осуществлять гибридизацию нуклеотидного олигомера с целевым IAP полинуклеотидом. В настоящем описании приводятся примеры таких остовов. В других вариантах реализации изобретения нуклеотидные олигомеры включают акридин-защищенные связи, компоненты холестерола или псоралена, С5-пропиниловые пиримидины. Приемлемые модификации нуклеотидных олигомеров согласно настоящему изобретению включают как вышеописанные, так и указанные в публикации заявки на патент США Примеры нуклеотидных олигомеров приведены ниже в Таблице 3 (где mX – 2′-O-метилированный Х остаток РНК).
Пенетратин и его применение при опосредованном введении нуклеотидных олигомеров в клетки описаны в Заявке на патент РСТ Аналогичный подход к идентификации был предпринят при создании анти-HIAP I нуклеотидных олигомеров. Первоначально были отобраны 98 нуклеотидных олигомеров из 19 остатков (SEQ ID NOs: 163-260; Таблица 4). Из этих 98 нуклеотидных олигомеров специфичных к последовательности НIАРI, для дальнейшей оценки были отобраны пятнадцать (SEQ ID NOs: 163-170, 173, 179, 202, 222, 223, 247 и 259). Эти пятнадцать отобранных нуклеотидных олигомеров включали четыре нуклеотидных олигомера, нацеленных на кодирующий участок (SEQ ID NOs; 202, 222, 223, и 247), один олигонуклеотид, нацеленный на 3’UTR (SEQ ID NO: 259), семь нуклеотидных олигомеров нацеленных на (SEQ ID NOs: 166-170, 173, и 179; один из семи нуклеотидных олигомеров, перекрывал старт-кодон) и три нуклеотида (SEQ ID NOs: 163-165), направленные на интронный сегмент 5’UTR.
Критерии отбора нуклеотидных олигомеров Отбор испытуемых нуклеотидов производили с использованием программы OLIGO (ранее распространяемой компанией National Biosciences Inc.) с учетом следующих критериев: 1) содержание GC не более 75%, и содержания AT не более 75%; 2) предпочтительны нуклеотидные олигомеры, не содержат четыре и более последовательно идущих G-остатков (в связи с имеющимися данными о токсическом эффекте, причем один их таких олигомеров был отобран для контроля токсичности); 3) предпочтительны нуклеотидные олигомеры, не обладающие способностью формировать устойчивые димеры или структуры типа «шпилька»; 4) предпочтительны последовательности, располагающиеся в зоне сайта инициации трансляции. В дополнение к вышесказанному доступные участки тРНК-мишени предварительно определили с помощью алгоритма предсказания вторичной структуры РНК MFOLD (M. Zuker, D. H. Mathews & D. H. Turner, «Алгоритмы и термодинамика предсказания вторичной структуры РНК: практическое руководство»; Algorithms and Thermodynamics for RNA Secondary Structure Prediction: A Practical Guide. In: RNA Biochemistry and Biothechnology, J. Barciszewski & B.F.C. Clark, Ed., NATO ASI Series, Kluwer Academic Publishers, (1999). Субоптимальные укладки молекулы РНК со значениями свободной энергии, находящимися в пределах отличия в 5% от прогнозируемой наиболее устойчивой укладки тРНК, рассчитывали с помощью «окна» из 200 нуклеотидов, в пределах которого остаток мог бы найти комплиментарное основание для формирования связи между основаниями. Открытые участки молекулы, которые не сформировали связь между парой оснований, суммировали с каждым субоптимальным вариантом укладки, а участки, которые постоянно определялись как открытые, считали более доступными для связывания с нуклеотидными олигомерами. Дополнительные нуклеотидные олигомеры, которые только частично соответствовали вышеперечисленным критериям, также отбирали, как возможные кандидаты в случае, если они распознавали вычисленный открытый участок m-РНК-мишени. Пример 2: Синтез олигонуклеотидов Способность нуклеотидных олигомеров ингибировать экспрессию IAP исследовали с помощью олигонуклеотидов, использовавшихся в качестве типичных нуклеоосновных олигомеров. Олигонуклеотиды синтезировали в компании IDT (Integrated DNA Technologies, USA), в виде химерных олигонуклеотидов «второго поколения», состоящих из центральной части, представленной фосфодиэфирными группами ДНК, фланкированными с обеих сторон двумя 2′-O-метилированными остатками РНК и фосфоротиоатной связью между ними. Олгонуклеотиды размещали в 96-луночном планшете, а также соответствующих пробирках, содержащих минимум 12 OD нуклеотидного олигомера, что должно было обеспечить достаточное количество материала для трансфекций (более чем 100 тестов с 96-луночным иммунологическим планшетом), при которых в качестве метода определения использовали количественные методы TaqMan PCR или ELISA. При удачном результате трансфекций производили ресинтез олигонуклеотидов с тремя, вместо имевшихся двух, флануирующих остатков РНК, чтобы дополнительно повысить стабильность/устойчивость к нуклеазе. В дополнение в целях подтверждения результатов для некоторых мишеней, обеспечивавших в результате наивысшую активность, синтезировали соответствующие контрольные образцы (например, олигонуклеотиды со случайно расположенным 4-нуклеотидным несоответствием и олигонуклеотиды с обратной полярностью). Пример 3: Скрининговые исследования и оптимизация нуклеотидных олигомеров Наш подход к идентификации нуклеотидных олигомеров, способных ингибировать экспрессию IAP заключался в исследовании вышеописанных библиотек олигонуклеотидов на наличие специфического снижения («нокдауна») РНК и/или белка-мишени для специфического гена IAP. Для определения уровня РНК и белка в клетках можно использовать любое количество стандартных исследований. Например, уровни РНК можно измерять с помощью нозерн-блот анализа или RT-PCR технологии. Уровень белка можно измерять, например, стандартным вестерн-блот анализом или с помощью методов иммунопреципитации. В качестве альтернативы клетки, в которые была введена нуклеиновая кислота IAP, могут быть исследованы на предмет жизнеспособности согласно способам, описанным, например, в патентах США Для оценки изменений уровней белка после воздействия олигонуклеотидами мы использовали количественную TaqMan PCR (описана ниже). Мы использовали способ ELISA для определения уровня белка XIAP и вестерн-блот анализ для определения уровня Н1АР1. Условия трансфекций оптимизировались путем использования Липофектимина Плюс или Липофектамина 2000 (Life Technologies, Canada) на клетках рака желчного пузыря Т24, клетках мелкоклеточного рака легких, или Липофектина на клетках глиобластомы SF-295, путем использования смысловых нуклеотидов с конъюгированным флуоресцентным маркером (флуоресцентно-маркированные нуклеотиды) (5′-mGmAGAAGATGACTGGTAAmCmA-3′; SEQ ID NO: 261) белка XIAP из области старт-кодона в качестве контроля. Результаты визуализировали и оценивали с помощью эпи-флуоресцентной микроскопии. В клетках Т24 дальнейшие трансфекции оптимизировали с помощью использования способности известного олигонуклеотида подавлять экспрессию сурвивина (Li et al., Nat. Cell Biol. 1: 461-466, 1999) (5′-U/TGTGCTATTCTGTGAAU/TU/T-3′ SEQ ID NO: 262). Оптимизацию условий трансфекции производили на основании результатов TaqMan теста, для нокдауна РНК сурвивина, определявшегося с помощью PCR-праймеров и флюоресцентного зонда, подробно описанному ниже. Было обнаружено, что оптимальными условиями для попадания олигонуклеотидов в клетки являются: 940 нМ олигонуклеотида, 4 µл реактива PLUS 0,8 µл Липофектимина при общем объеме 70 µл с выдержкой 3 часа. После этого применяли вышеописанные условия для отслеживания нокдауна белка XIAP при использовании библиотеки олигонуклеотидов на клетках Т24. Нокдаун НIАР1 исследовали на клетках SF-295, так как в них легко определяется и распознается белок HIAP 1 (70 кДа), в то время как многие другие линии клеток не экспрессируют достаточно высокий уровень белка или он не распознается среди большого количества белка HIAP 2, имеющего сходные размеры (68 кДа). ПЦР в реальном времени (Real-time, RT-PCR) РНК экстрагировали из клеток, лизированных в буфере RLT (QIAGEN, Valencia, СА) и очищенных в колонках/наборах QIAGEN RNeasy). Количественную PCR в реальном времени производили на установке Perkin-Elmer ABI 7700 Prism PCR. Обратная транскрипция РНК и ее амплификация происходила в соответствии с протоколом TaqMan Universal PCR Master Mix от РЕ Biosystems при использовании праймеров и зондов, разработанных для специфичного распознавания XIAP, HIAP1, сурвивина или GAPDH. Для человеческого сурвивина прямым праймером являлся 5′-TCTGCTTCAAG GAGCTGGAA-3′ (SEQ ID NO: 263), обратным – 5′-GAAAGG AAAGCGCAACCG-3′ (SEQ NO: 264), зондом – 5′-(FAM)AGCGAGATGACGACCCCATAGAGGAACATA(TAMRA)-3′ (SEQ ID NO: 265). Для человеческого HIAP1 прямым праймером являлся 5′-TGGAGATGATCCATG GGTTCA-3′ (SEQ ID NO: 266), обратным – 5′-GAACTCCTGTC CTTTAATTCTTATCAAGT-3′ (SEQ ID NO: 267) и зондом – 5′-(FAM)CTCACACCTTGGAAACCACTTGGCATG(TAMRA)-3′ (SEQ ID NO: 268). Для человеческого ХIАР прямым праймером являлся 5′-GGTGATAAAGTA AAGTGCTTTCACTGT-3′ (SEQ ID NO: 269), обратным праймером – 5′-TCAGTAGTTCTTACCAGACACTCCTCAA-3′ (SEQ ID NO: 270) и зондом -CAACATGCTAAATGGTATCCAGGGTGCAAATATC(TAMRA)-3′ (SEEQ ID NO: 271). Для человеческого GAPDH прямым праймером являлся 5′-GAAGGTGAAGGTCGGAGTC-3′ (SEQ ID NO: 272), обратным – 5′-GAAGATGGTGATGGGATTC-3′ (SEQ ID NO: 273) и зондом 5′(JOE)CAAGCTTCCCGTTCTCAGCC (TAMRA)-3′ (SEQ ID NO: 274). FAM – это 6-карбоксифлюоросцеин, JOE – 6-карбокси-4,5-дихлоро-2,7-диметоксифлюоросцеин и TAMRA – 6-карбокси-N,N,N’,N’-тетраметилродамин. FAM и JOE относятся к 5′ репортер-красителям, а TAMRA-является 3′-«гасящим» красителем. Относительный количественный анализ экспрессии гена выполняли в соответствии с описанием, данным в руководстве РЕ Biosystems при использовании GAPDH в качестве внутреннего стандарта. Для относительного количественного анализа уровней IAP мРНК и GAPDH мРНК использовали сравнительный метод циклического порога (Ct). Вкратце, измерения флуоресценции в режиме реального времени, проводили в ходе каждого цикла PCR, пороговые значения цикла (Ct) вычисляли путем определения точки, при которой флуоресценция превышала пороговое значение, равное 30-кратному фоновому (baseline) стандартному отклонению (SD). Среднее фоновое значение и фоновое SD вычисляли, начиная от фонового значения третьего цикла и заканчивая фоновым значением за три цикла до того, как сигнал начинал экспоненциально возрастать. Праймеры и/или зонды PCR для специфических IAP разрабатывали так, чтобы они могли покрывать, по крайней мере, одну пограничную зону экзон-интрон, разделенную 1 кб или более геномной ДНК для сокращения потенциальной возможности амплификации и детекции согнала от геномной ДНК. Специфичность сигнала и его предполагаемое «загрязнение» неспецифичным сигналом от ДНК подтверждали обработкой некоторых образцов РНК ДН-азой или РН-азой перед осуществлением обратной транскрипции или этапами PCR. Вестерн-блот анализ НIАР1 и ХIАР ELISA. Стандартный колориметрический анализ ХIАР ELISA производили с использованием очищенных на основании сродства кроличьих поликлональных антител к ХIАР, выступавших в качестве иммобилизованных антител и детектировали с помощью моноклональных антител к ХIАР (MBL, Japan) и биотинилированных антител к мышиному Ig и стрептовидином, конъюгированным с пероксидазой хрена и ТМВ (тетраметилбензолом) в качестве субстрата. В альтернативном варианте для измерения уровня белков ХIАР и НIАР1 можно использовать поликлональные антитела XIAP и НIАР1 соответственно. НIАР1 определяли с помощью вестерн-иммуноблот анализа с использованием очищенных антикрысиных кроличьих антител к НIАР1, применявшихся в качестве первичных антител и выявляли с помощью ECL (Amersham) на рентгеновской пленке со вторичным антителом против IG кролика, конъюгированным с пероксидазой хрена, и хемилюминисцентным субстратом. Анти-НIАР1 поликлональные антитела получали против GST-белка слияния – крысиного НIАР1. Это антитело дает перекрестную реакцию и с человеческими, и с мышиными белками НIАР1 и HIAP2. Пример 4: Антисенс XIAP олигонуклеотиды снижают РНК и экспрессию полипептида ХIАР. Синтезированную библиотеку ХIАР, включающая 96 олигонуклеотидов сначала проверяли для выявления снижения уровня белка ХIАР. В частности, клетки Т24 высеяли в лунки 96-луночного иммунологического планшета (1,5×104 клеток в лунке) в 1-й день и культивировали в среде Маккоя (МасСоу) в течение 24 часов. На 2 день производили трансфекцию клеток антисмысловым нуклеотидом XIAP по методике, описанной выше (олигонуклеотиды маркировали в соответствии с положением на планшете, например от А1 до Н12 и включали два повтора А13 и В13, содержавших лиофилизированные гранулы ДНК, прилипшие к герметизирующей мембране). Олигонуклеотиды растворяли в 10 µл/на лунку бессывороточной, не содержащей антибиотики среде МакКоя, после чего добавляли реактив PLUS. Липофектамин растворяли в 10 µл/на лунку бессывороточной, не содержащей антибиотики среде Маккоя и обе смеси инкубировали в течение 15 минут при комнатной температуре. После этого смеси соединяли и снова инкубировали в течение 15 минут при комнатной температуре. В это время удаляли полную среду и к культурам клеток добавляли 50 µл/на лунку бессывороточной, не содержащей антибиотики среды МакКоя. Смеси для проведения трансфекции добавляли в лунки, после чего клетки инкубировали в течение трех часов. После этого в каждую лунку добавляли по 30 µл бессывороточной, не содержащей антибиотики среды и 100 µл/на лунку полной среды, не содержащей антибиотики с 20% содержанием сыворотки теленка. На 3 день измеряли уровни XIAP РНК с помощью количественной PCR в режиме реального времени по методике, описанной выше. На 4 день измеряли уровни белка XIAP с помощью ELISA (Фиг.1А, 1C, 1Е, 1G, 1I и 1К) общего (тотального) клеточного белка биохимическим способом (Фиг.1В, 1D, 1F, 1Н, 1J и 1L), что использовали для стандартизации уровня белка XIAP). Результаты сравнивали с образцом, подвергшимся холостой трансфекции (вводили трансфекционный препарат, но без добавления олигонуклеотида ДНК, после чего проводились те же процедуры, что и для других образцов). В ходе временных экспериментов было определено, что оптимальная длительность нокдауна белка должна составлять от 12 до 24 часов. Библиотеку олигонуклеотидов изучали, также, с целью выявления снижения уровней РНК в течение оптимального периода времени с использованием TaqMan-специфичных праймеров PCR и флюоресцентных зондов, описанных выше. В ходе временных экспериментов было определено, что оптимальное уменьшение уровня тРНК происходит через 6-9 часов. Эти результаты хорошо согласуются с результатами исследования белка. В ходе первого скрининга (выполнявшегося в суб-оптимальную временную точку, когда уровни XIAP возвращаются к нормальным значениям, возможно, благодаря разрастанию нетрансфицированных клеток) было идентифицировано 16 антисмысловых олигонуклеотидов (Таблица 1: С2, Е2, Е3, F3, С4, D4, Е4, F4, G4, С5, D5, В6, F6, D7, D8, F8) из 96 протестированных, которые продемонстрировали некоторое снижение уровней белка XIAP по отношению к общему (тотальному) белку при сравнении с уровнями белка при холостой трансфекции, проводившейся без олигонуклеотида (Фиг.1А, 1C, 1Е, 1G, 1I и 1К). Снижение уровня общего белка происходило для каждого из этих 16 олигонуклеотидов, что свидетельствует о токсическом или цитостатическом эффекте этих олигонуклеотидов. (Фиг.1В, 1D, 1F, 1Н, 1J, 1L). Олигонуклеотиды В9 и С9 продемонстрировали явное снижение уровня общего белка при отсутствии относительного падения уровней белка XIAP. 16 антисмысловых олигонуклеотидов, продемонстрировавших снижение относительного уровня белка XIAP по сравнению с холостой трансфекцией, были повторно исследованы по отдельности или в комбинации совместно с контрольным олигонуклеотидом (D2), на предмет их способности вызывать нокдаун белка XIAP в наиболее оптимальной временной точке (12 часов), определенной в ходе вышеописанных временных исследований (Фиг.2В). Указанные нуклеотидные олигомеры исследовали также на предмет их способности снижать уровни XIAP тРНК на 12 часу эксперимента, стандартизированного по отношению к уровню GARPDH и в сравнении с холостой трансфекцией. Была также определена общая концентрация белка на 12-м часу эксперимента (Фиг.2С). Наблюдали хорошую корелляцию между способностью олигонуклеотида снижать уровень белка XIAP (Фиг.2В) и способностью снижать уровни XIAP тРНК (Фиг.2А). Кроме того, не наблюдали большой потери общего белка в этой ранней временной точке, причем снижение XIAP mРНК белка предшествовало снижению общего белка, проявлявшемуся в более поздних временных точках. Олигонуклеотиды, продемонстрировавшие снижение уровней белка XIAP или его mРНК более, чем на 50% при использовании изолированно или в комбинации двух олигонуклеотидов, в соотношении 1:1, определяли как лучшие олигонуклеотиды и подвергали дальнейшим исследованиям. Из этих 16 олигонуклеотидов 10 (Е2, Е3, F3, Е4, F4, G4, С5, В6, D7, F8) проявили устойчивую способность снижать уровни белка XIAP и РНК более чем на 50%, в зависимости от используемых условий трансфекции, или при использовании их в комбинации (как для С5 и G4). Более того, 16 олигонуклеотидов, проявивших антисмысловую активность, группировались по 4 различных зоны-мишени тРНК XIAP, а прилежащие нуклеотидные олигомеры, которые тоже вызывали некоторый эффект нокдауна. У нуклеотидных олигомеров, котороые в качестве мишени имели участки последовательности между вышеуказаными зонами, или островками чувствительности наблюдали очень незначтельную антисмысловую активность или ее полное отсутствие. По-видимому, эти участки представляют собой зоны открытой конформации тРНК, которые доступны нуклеотидным олигомерам внутри клетки. Два нуклеотидных олигомера Е3 и F3, контактируют с ХIАР непосредственно выше по направлению транскрипции от старт-кодона в области между IRES и сайтом инициации трансляции и частично перекрывают конец IRES элемента. С2, D2 и Е2 направлены на область ХIАР, расположенную выше по транскрипции от минимального элемента IRES, что также является свидетельством того, что минимальная зона IRES – это высокоструктурированная зона РНК, труднодоступная для нуклеотидных олигомеров in vivo. Все остальные нуклеотидные олигомеры комплементарны части кодирующей зоны, включая кластер активности в позициях 856-916 последовательности XIAP (Е4, F4 и G4) и меньшим по размерам изолированным зонам, как было показано на примере нуклеотидных олигомеров, например С5 и D5. Часть 96 нуклеотидных олигомеров, приведенных в Таблице 5, была подвернута повторному скринингу для выявления способности вызывать нокдаун XIAP mPHK в клетках NCI-H460 на 9 часу после трансфекции. Данные приведены в Таблице 5.
Мы также определяли, способны ли 4×4 МВО (все остатки PS и ДНК, фланкированные с обеих сторон четырьмя 2′-O-метилированными остатками РНК) вызывать нокдаун белка XIAP в клетках Н460. Как показано на Фиг.3 и 4, 4×4 MB олигуонуклеотида Е12 и другого олигонуклеотида, FG8, были эффективны в таких малых дозах, как 31 нМ. Пример 5: Повышение цитотоксичности и хемосенсибилизации антисмысловыми нуклеотидными олигомерами XIAP. Нами были проведены исследования относительно способности антисмысловых онуклеотидных олигомеров XIAP хемосенсибилизировать высокоустойчивые к медикаментозному лечению клетки Т24 к традиционным химиотерапевтическим препаратам, таким как адриамицин или цисплатин. Антисмысловые олигонуклеотиды отбирали таким образом, чтобы представлять несколько различных зон-мишеней XIAP и испытывали на предмет их цитотоксического эффекта изолированно или в сочетании с другими олигонуклеотидами или препаратами. Пять антисмысловых олигонуклеотидов XIAP были исследованы на способность убивать или хемосенсибилизировать Т24 клетки рака мочевого пузыря, после чего проводили сравнение с действием, полученным от трех соответствующих рандомизированных контрольных олигонуклеотидов. Клетки Т24 были трансфицированы антисмысловыми олигонуклеотидами ХIАР, рандомизированными (scrambled) олигонуклеотидами и материалом, в котором олигонуклеотиды отсутствовали (холостой контроль) либо трансфекция клеток вообще не проводилась. Через 20 часов после трансфекции клетки исследовались на жизнеспособность (за исключением клеток, участвовавших в холостом контроле) при помощи анализа cWST-1 тетразолиевым красителем, где WST-1 тетразолиевый краситель восстанавливался в окрашенный продукт распада формазана в метаболически активных клетках (Фиг.5А). Наступление цитотоксического эффекта, индуцированное олигонуклеотидом Е4, исследовали визуально, путем изучения клеток Т24, не подвергавшихся воздействию, после холостой трансфекции, или трансфицированных Е4, случайно отобранными олигонуклеотидами Е4, олигонуклеотидами Е4 с обратной полярностью или олигонуклеотидами Е4, с несовподающими основаниями. Через 20 часов после трансфекции исследовали морфологию клеток (Фиг.5D). Признаки токсичности были выявлены только в клетках, трансфицированных антисмысловыми олигонуклеотидами Е4. Для изучения влияния нуклеотидных олигомеров на хемосенсибилизацию клеток Т24 к цисплатину или адриамицину, исследовали способность нуклеотидов убивать эти клетки в присутствии фиксированной дозы адриамицина (0,5 µг/мл). Клетки сначала трансфицировали олигонуклеотидом, затем добавляли адриамицин на 20 часов. Жизнеспособность измерялась с помощью WST-1 в конце 20 часового воздействия лекарственным препаратом (Фиг.5В). Результаты показаны на Фиг.5С в виде процента жизнеспособности по сравнению с образцами, подвергавшимися воздействию только нуклеотидным олигомером. Все пять исследуемых нуклеотидных олигомеров (F3, Е4, G4, С5, D7), а также комбинации Е4+С5 и G4+C5, убивали клетки Т24, оставляя только 10-15% жизнеспособных клеток через 24 часа после трансфекции по сравнению с клетками, подвергшимися холостой трансфекции (без олигонуклеотидов) или тремя соответствующими случайно изменными (scrambled) контрольными олигонуклеотидами для F3 (5′-mCmAmGAGATTT CATTTAAmCmGmU-3′; SEQ ID NO: 275), E4 (5′-mCmUmACGCTCGCCATCGTmUmCmA-3′; SEQ ID NO: 276), и C5 (5′-mUmGmCCCAAGAATACTAGmUmCmA-3′; SEQ ID NO: 277) (Фиг.5А и 5С). Следовательно, токсичность является последовательность-специфичной для нуклеотидных олигомеров, которые снижают уровень белка ХIАР и не связяна с последовательность неспецифичными хомоческими свойствами олигонуклеотидов. Эта цитотоксичность может быть обусловлена сочетанием эффекта нокдауна белка ХIАР (и ожидаемой утратой анти-апоптозной защиты, которую обеспечивает ХIАР), а также цитотоксичностью процедуры трансфекции как таковой. Дополнительное введение фиксированной дозы адриамицина или цисплатина в конце трехчасового периода трансфекции привело к дальнейшему снижению жизнеспособности клеток в отношении некоторых испытуемых олигонуклеотидов, дальнейшему снижению жизнеспособности на 40% через 20 часов после трансфекции для нуклеотидных олигомеров F3, D7 и комбинации G4+C5 (Фиг.5В) по сравнению с соответствующими значениями, полученными при воздействии олигонуклеотидами (Фиг.5С). Значения на Фиг.5В (олигонуклеотид плюс препарат) сравнивались со значениями монолечения олигонуклеотидом (Фиг.5С), который принимали за 100% для каждого ODN. Показаны только результаты хемосенсибилизации к адриамицину; аналогичные результаты были получены при хемосенсибилизации клеток цисплатином. При использовании фиксированных доз, холостая трансфекция и трансфекция случайно измененными олигонуклеотидами, в отличие от случаев введения адриамицина, не показала какого либо снижения выживаемости клеток (Фиг.5В). Хемосенсибилизация проявилась только в случаях, когда уровни XIAP были снижены специфическим антисмысловым олигонуклеотидом. Пример 6: Эффекты понижающей регуляции антисмысловых олигонуклеотидов на ХIАР mРНК в клетках Н460. Антисмысловые олигонуклеотиды (2×2 МВО, состящие из двух 2′-O-метилированных остатков РНК, соединенных фосфоротиоатными связями фланкирующих с обеих сторон на центральную часть, состоящую из 15 фосфодиэфирных остатков ДНК) исследовали с использованием метода PCR в режиме реального времени с целью изучения их влияния на ХIАР mРНК в клетках Н460. При такой конфигурации нуклеотидные олигомеры F3, G3, С5, АВ6 и DE4 уменьшали уровень mРНК на 50-70% по сравнению с необработанным контролем, в то время как D7 AS нуклеотидные олигомеры сокращали уровень mРНК на 30% (Фиг.6). Напротив, контрольные нуклеотидные олигомеры и вводимый изолированно препарат (LFA) по отдельности снижали уровень mРНК менее, чем на 20% по сравнению с необработанным контролем (Фиг.6). Для проведения дальнейших исследований in vivo и in vitro были отобраны нуклеотидные олигомеры F3, G4 и С5. Дополнительный нокдаун ХIАР mРНК, отмечавшийся при проведении анализа TaqMan, изображен на Фиг.7 и 8. Пример 7: Влияние понижающей регуляции (супрессии) антисмысловых олигонуклеотидов на белок ХIАР Мы охарактеризовали влияние нуклеоосновных олигомеров F3, G4 и С5, имеющих конфигурацию олигонуклеотидов, на экспрессию белка ХIАР с помощью вестерн-блот анализа (Фиг.9, 10А и 10В). Олигонуклеотиды G4 AS проявили самый сильный супрессирующий эффект на белок XIAP, понижая уровни его содержания на 62% через 24 часа после окончания трансфекции в концентрации 1,2 µМ (Фиг.10А и 10В). F3 AS олигонуклеотиды в концентрации 1,2 µМ понижали уровень белка XIAP на 50%, тогда как С5 AS олигонуклеотиды не продемонстрировали последовательность специфичного эффекта по сравнению с контрольными образцами (Фиг.10В). В дополнительных исследованиях было выявлено, что AS олигонуклеотиды Е12 и FG8 в значительной степени снижали уровни белка XIAP (Фиг.9). Пример 8: Индукция апоптоза AS олигонуклеотидами XIAP После того, как было определено, что AS нуклеотидные олигомеры XIAP способны снижать жизнеспособность клеток рака мочевого пузыря линий Н460 и Т24, мы определили, была ли наблюдавшаяся смерть клеток обусловлена индукцией апоптоза. Как показано на Фиг.11А, в клетках Н460, подвергшихся обработке олигонуклеотидами F3 или G4 AS в концентрации 1,2 µМ, происходила активация и деградация белка про-каспазы-3, сопровождающаяся снижением на 40% и 60% уровней белка соответственно, по сравнению с необработанными контрольными клетками. Также был обнаружен предполагаемый продукт расщепления PARP каспазой-3 (Фиг.11А). В противоположность этому, SC контрольные образцы олигонуклеотидов F3 и G4 в концентрации 1,2 µМ не оказывали никакого влияния на каспазу-3 или экспрессию белка PARP (Фиг.11А). В клетках Н460, обработанных AS олигонуклеотидами F3 и G4 в кнцентрации 1,2 µМ, и соответствующих контрольных образцах соотношение Dcl-2:Bax оставалось неизменным. Для выявления содержания гипо-диплоидной ДНК в клетках Н460, подвергшихся обработке AS олигонуклеотидами G4 и окрашенными PI, использовали метод поточной цитометрии (Фиг.12А). В клетках Н460, подвергшихся обработке AS олигонуклеотидами G4 или SC олигонуклеотидами контроля в концентрации 1,2 µМ количество клеток с гипо-диплоидным содержанием ДНК составило 40,8 и 22,1% соответственно по сравнению с 16,6% для клеток группы необработанного контроля. Для выявления морфологических изменений в ядре клеток Н460, подвергшихся воздействию AS олигонуклеотидами G4 в концентрации 1,2 µМ, использовали окрашивание DAPI. Как показано на Фиг.12В, клетки, подвергшиеся воздействию AS олигонуклеотидами G4, претерпевают морфологические изменения, характерные для апоптоза, включая конденсацию хроматина и фрагментацию ядерной ДНК. Только немногие клетки имели аналогичные морфологиечские изменения в образцах клеток, обработанных SC G4. Пример 9: Ингибирование клеточного роста и сенсибилизация клеток Н460 к противораковому препарату AS олигонуклеотидами Для изучения биологического эффекта нуклеотидных олигомеров, связанного с понижающей регуляцией экспрессии XIAP и апоптозом, рост клеток Н460, подвергшихся воздействию AS олигонуклеотидами G4, исследовали с помощью МТТ анализа. Через сорок восемь часов после трансфекции AS олигонуклеотиды G4 в концентрации 1,2 µМ сократили клеточный рост в дозо-зависимой форме, демонстрируя 55% снижение по отношению к необработанному контролю (Фиг.13А). В противоположность этому, ингибирующий рост эффект SC олигонуклеотидов G4 или изолированного препарата был сравнительно низким – менее 10% по отношению к соответствующему необработанному контролю. Для определения обладает ли понижающая регуляция экспрессии Х1АР потенциалом для сенсибилизации клеток Н460 к химиотерапевтическим препаратам проводилось комбинированное лечение AS олигонуклеотидами G4 и одним из следующих противораковых препаратов: доксорубицин (DOX), таксол, винорельбин (VNB) и этопозид (Etop). На Фиг.13В показано, что применение каждой из комбинаций привело по меньшей мере к аддитивному цитотоксическому эффекту в отношении клеточной смертности по сравнению с лечением либо AS олигонуклеотидом G4 либо противораковым препаратом, взятыми по отдельности. Пример 10: Противоопухолевый эффект AS олигонуклеотидов G4 в отношении ксенотрансплантатов опухолей Н460 и LCC6 Сначала определяли, сокращает ли объем опухолевого роста внутриопухолевая инъекция антисмысловых 2×2 МВО XIAP в подкожные ксенотрансплантаты рака легких человека Н460 у мышей-носителей SCID-RAG2. Лечение началось через 14 дней после инокуляции опухолевых клеток (инъекция в плечевую область 106 клеток) путем иъекции МВО в пальпируемую опухолевую массу три раза в неделю на протяжении двух недель (50 микрограмм 2′-O-метил РНК олигонуклеотидов из расчета на грамм опухоли). Винорельбин (VNB), также известный как навельбин (NVB) (15 мг/кг), вводили путем инъекции на 17 и 24 дня. Размер опухоли определяли при помощи штангенциркуля три раза в неделю. В конце периода лечения (24 дня) средний относительный рост опухоли у мышей, получавших лечение комбинацией C5+G4 AS МВО и VNB, сократился на ~70% по сравнению с мышами, получавшими лечение случайно измененными (SC) контрольными образцами МВО и VNB. Лечение С5 AS МВО и VNB привело к сокращению размера опухоли на ~60% по сравнению с рандомизированным контролем (Фиг.14). Начальный этап системных исследований PS-олигонуклеотидов был разработан без участия каких-либо химотерапевтических препаратов. Мышам SCID-RAG2 вводили клетки Н460 рака легких человека (инъекция в плечо 106 клеток) после чего через три для после инокуляции начинали лечение G4 и F3 AS PS-олигонуклеотидами, а также случайно измененными олигонуклеотидами, составлявшими группу контроля. Инъекции нуклеотидного олигомера назначались из расчета 12,5 мг/кг три раза в неделю в течение трех недель. В конце периода лечения средний размер опухоли в группах, получавших лечение любым из AS олигонуклеотидов G4 или F3, составил на ~50% меньше, чем в группе, получавшей лечение олигонуклеотидами группы SC контроля (Фиг.15). Ааналогичная схема лечения была испытана на самках мышей SCID-RAG2 с ортотопическим трансплантатом клеток рака молочной железы человека MDA-MB-435/LCC6. Две недели спустя после лечения (35-й день) объем опухоли у мышей, получавших лечение F3, С5, G4 AS олигонуклеотидами составил на 70%, 60% и 45% меньше соответственно, чем в контрольных группах с наполнителем (Фиг.16). Нами было проведено дополнительное исследование противоопухолевого эффекта G4 AS олигонуклеотидов у мышей-носителей SCID-RAG2 ксенотрансплантата Н460 не мелкоклеточного рака легких человека, имплантированного подкожно. Опухоли в контрольной группе, в которые вводили физраствор, выросли до размера 0,75 см3 за период около 24 дней (Фиг.17). Лечение олигонуклеотидами было начато через 3 дня после инокуляции опухолевых клеток. AS олигонуклеотиды G4 (от 5 до 15 мг/кг) вводили в соответствии со схемой лечения, т.е. в виде инъекций один раз в день на 3-7, 10-14 и 17-21 дней. Лечение AS олигонуклеотидами G4 в дозе 5 или 15 мг/кг в значительной степени замедлило рост опухоли: на 24 день средний размер опухоли составил 0.75, 0.45 и 0.29 см3 в контрольной группе, группах 5 и 15 мг/кг соответственно (Фиг.18А). Ингибирование роста опухоли имело дозо-зависимый характер. Размеры опухоли у мышей, получавших лечение AS олигонуклеотидами G4 в дозе 15 мг/кг, были значительно меньше, чем в контрольной группе и составляли 39% от среднего размера опухолей в контрольных группах. Напротив, введение SC олигонуклеотидов G4 в дозе 15 мг/кг не оказало какого-либо терапевтического действия (Фиг.17). Ни одна из особей мышей, получавших лечение олигонуклеотидами, не проявила никаких признаков интоксикации, обе дозы олигонуклеотидов хорошо переносились. Доза, составляющая 15 мг/кг, была выбрана для дальнейшего комбинированного лечения с противоопухолевыми препаратами. Пример 11: Сокращение экспрессии XIAP в опухолях Н460, подвергавшихся воздействию AS олигонуклеотидами G4 С тем, чтобы установить ингибирующее действие AS олигонуклеотидов G4 на опухолевый рост с экспрессией белка XIAP, исследовали изменения экспрессии XIAP в конце лечения, проводившегося in vivo AS и SC олигонуклеотидами G4 в дозе 15 мг/кг. На 21 или 24 день после инокуляции опухоли, когда ее размеры достигли 1 см3 (Фиг.17), опухоли были собраны и лизаты гомогенизированных опухолей были использованы для вестерн-блот анализа. Были использованы XIAP и Пример 12: Гистопатология образцов опухоли Для оценки того, приводит ли введение олигонуклеотида XIAP AS к непосредственной гибели опухолевых клеток, мы провели гистологическое исследование опухолей после лечения, как для изучения их морфологии, так и иммуноокрашивания на убиквитин (Фиг.19А и 19В). Опухоли, на 21 или 24 день после инокуляции подвергавшиеся воздействию AS олигонуклеотидами G4, SC олигонуклеотидами в дозе 15 мг/кг или физраствором, удаляли оперативным путем, из них брали срезы и окрашивали гематоксилином и эозином. Результаты свидетельствуют о том, что опухоли у животных, которым вводили указанный AS олигонуклеотид XIAP, содержали повышенное число погибших клеток, что подтверждалось морфологически их аморфной структурой и конденсацией ядерного материала (Фиг.19А). Деградация белков в значительной степени является убиквитин-протеасомо-зависимой; во время апоптоза наблюдали повышенную экспрессию убиквитина. Поэтому мы решили исследовать экспрессию убиквитина в срезах опухолей, окрашенных гематоксилином и эозином. Как показано на Фиг.19В, опухоли у мышей, получавших лечение AS олигонуклеотидами XIAP, продемонстрировали более интенсивное иммунохимическое окрашивание относительно опухолей контрольной группы или мышей, получавших лечение SC ODN. Эти данные свидетельствуют о том, что в клетках опухолей, обработанных AS нуклеотидными олигомерами XIAP больше свободного убиквитина и/или убиквитинированного белка, чем в контрольных образцах опухолей. Пример 13: Комбинированное лечение AS олигонуклеотидами G4 и винорельбином Для оценки того, оказывает ли комбинированное лечение AS нуклеотидными олигомерами G4 и винорельбином (VNB), химиотерапевтическим препаратом, используемым для онкотерапии рака легких какие-либо синергические эффекты, сравнивали терапевтическую эффективность VNB в присутствии или отсутствии AS или SC олигонуклеотидов G4. Схемы лечения начинали с 3-го дня после инокуляции опухолевых клеток. На Фиг.20А представлена эффективность in vivo для VNB, который давали мышам-носителям опухолей Н460 в дозах 5 мг/кг и 10 мг/кг, по сравнению с контрольной группой, в которой применяли физраствор. Каждая из двух схем вызывала значительное подавление роста опухоли в дозо-зависимой манере без проявления выраженных признаков токсичности (например, потеря массы тела). Когда введение AS олигонуклеотидов G4 (15 мг/кг) сочеталось с VNB (5 мг/кг) при лечении опухолей Н460 наблюдали еще более выраженное замедление опухолевого роста по сравнению с лечением одним из препаратов (Фиг.20В). Опять же, у мышей не было выявлено ни одного значительного признака токсичности (например, потеря веса). Средние размеры опухоли у мышей, получавших лечение 5 мг/кг VNB в присутствии или отсутствии AS или SC олигонуклеотидов G4, сравнивали на 29-й день (Фиг.20А и 20В). Среднеарифметический размер опухоли в группе VNB и AS олигонуклеотид G4 составил 0,22±0,03 см3, что было значительно ниже, чем среднеарифметический размер опухоли у животных, получавших монотерапию 5 мг/кг VNB или комбинацию VNB/ SC олигонуклеотид G4 (0.59±0.04 и 0.48±0.05 см3, соответственно). Способы Результаты, приведенные в примерах 5-13, были получены при использовании следующих способов. Синтез олигонуклеотидов Библиотека, содержащая более 96 неперекрывающихся химерных олигонуклеотида или нуклеотидов со смешанным остовом (МВО), составляли из 19-мерных антисмысловых олигонуклеотидов МВО, которые синтезировали как 2×2 МВО олигонуклеотиды, состоящие из 2 фланкирующих с двух сторон 2′-O-метилированных остатков РНК, соединных фосфоротиоатными связями с центральной частью, состоящей из 15 фосфордиэфирных остатков ДНК. Каждый конечный продукт обессоливали способом хроматографии Sephadex G-25 (IDT Inc., Coralville, IA). Указанная химерная крыловидная конфигурация и сочетание фосфоротиоатных и фосфодиэфирных связей (упоминаемых, как 2×2 PS/PO), обеспечила адекватную стабильность, одновременно сокращая неспецифическую токсичность, обусловленную фосфоротиоатными связями. Фосфоротиоатные нехимерные (ДНК) антисмысловые олигонуклеотиды для исследований in vivo и in vitro были полностью синтезированы Trilink Biotech, очищены – способом RP-HPLC. Скрининг антисмысловых олигонуклеотидов Клетки рака мочевого пузыря, которые подвергались трансфекции комплексами олигонуклеотид-липофектин (1-1,2 µМ) в течение 24-28 часов, исследовали для определения способности каждого из олигонуклеотидов вызывать нокдаун белка XIAP. При положительных результатах производили повторное определение способности вызывать нокдаун белка XIAP (i) на 12-18 часу трансфекции вестерн-блот анализом и (ii) XIAP mPHK на 6-9 часу трансфекции с помощью количественной RT-PCR (см. ниже) в клетках рака мочевого пузыря Т24 и клетках рака легких Н460. Отобранные олигонуклеотиды идентифицировали и подвергали дальнейшим исследованиям. Идентифицированные химерные олигонуклеотиды 2×2 PS/PO проявили дозозависимую способность к снижению уровней XIAP mPHK на 6-9 часу трансфекции в диапазоне концентраций 400-1200 нМ. Типичные олигонуклеотиды представлены в Табл.6.
Линия опухолевых клеток и модель ксенотрансплантатов у животных (животных ксенотрансплантатов) Линия клеток не мелкоклеточного рака легких человека NCI-H460(H460) получили от АТСС и культивировали в RPMI 1640 с добавлением 10% FCS во влажной атмосфере, содержащей 5% СО2. Клетки использовали в экспоненциальной фазе роста до 25 пассажей in vitro. Самцов мышей SCID-RAG2 (7-9 недель, 23-26 г) брали из колонии молодняка Объединенной зоологической лаборатории агенства по изучению рака Британской Колумбии и содержали в асептических условиях. Модель опухоли, состоящей из клеток NCI-H460, у мышей SCID-RAG2 формировали путем подкожной имплантации 1×106 клеток NCI-H460 на спине. Обработка клеток антисмысловым и противоопухолевым препаратами За день до проведения трансфекции клетки Н460 помещали на 6- или 96-луночные культуральные планшеты. В клетки вводили фосфоротиоатные антисмысловые олигонуклеотиды в комбинации с Липофектимином (Life Technologies) в виде комплексов липосома-олигонуклеотид. В последующие 4,5 или 6 часов трансфекционная среда заменялась средой RPMI, содержащей 10% FBS, после чего клетки подвергались инкубации еще 24 или 48 часов. Количественный анализ RT-PCR в режиме реального времени Общая РНК клеток Н460, подвергавшихся воздействию комплексами липосома-олигонуклеотид в течение 6 часов, немедленно извлекали с помощью колонок “RNeasy mini spin columns» и обработки ДН-азой (QIAGEN, Valencia, CA). Специфическую mPHK XIAP измеряли с помощью количественного анализа RT-PCR в режиме реального времени. Прямые и обратные праймеры XIAP (600 нМ), и зонд (200 нМ) (5′-GGTGATAAA GTAAAGTGCTTTCACTGT-3′ (SEQ ID NO 293), 6FAM-CAACATGCTA AATGGTTCCAGGGTGCAAATATC-TAMRA (SEQ ID NO: 294), and 5′-TCAGTAGTTCTTACCAGACACTCCTCAA-3′ (SEQ ID NO: 295) были разработаны таким образом, чтобы они захватывали границы экзонов 3-4 и 4-5. Один из праймеров, а также зонд, были разработаны таким образом, чтобы они перекрывали экзон-интронную границу для блокировки детекции ложного сигнала геномных ДНК. Осуществлялась обратная транскрипция РНК и амплификация PCR с помощью комплекта TaqMan EZ RT-PCR (PE/ABI, Foster City, CA) и системы для определения последовательностей (секвентатора) ABI Prism 7700 (PE/ABI). Условия термоциклирования (периодического изменения температуры) на этапе RT составляли 50°С в течение 2 мин, 60°С – 30 мин и 95°С – 5 мин, после чего следовали 45 циклов PCR (94°С для 20 сек и 60°С для 1 мин циклов). Уровни XIAP mPHK для каждого образца рассчитывали по отношению к клеткам группы необработанного контроля. Уровни XIAP mPHK определяли способом циклического порога (Ct), где пороговая величина составляла 30Х величины SD фонового значения, а уровни XIAP нормировали по уровню содержания GAPDH с помощью праймеров PE/ABI и зонда. Вестерн-блот анализ Клетки или ткани образцов опухоли подвергали лизису лизирующим буфером (50 мМ Tris, 150 мМ NaCl, 2.5 мМ EDTA, 0.1% SDS, 0.5% натрия дезоксихолат, 1% NP-40, 0.02% натрия азид) на льду, содержащим ингибиторы протеазы (Таблетки ингибитора протеазы Complete-Mini; Boehringer Mannheim GmBH, Mannheim, Германия). После инкубации на льду в течение 30 минут образцы центрифугировали при скорости 10000 об/мин 15 минут и сохраняли при температуре -20°С. Содержание белка в экстрактах лизата определяли с помощью анализа DC Bio-Rad (Detergent-compatible Bio-Rad assay, Bio-Rad Labs, Hercules, CA). Равные количества белка (40 µг/lane) сепарировали на 12% SDS-полиакриламидном геле или 4-15% градиентном SDS-полиакриламидном готовом геле (Bio-Rad) и переносили на нитроцеллюлозные мембраны. Использовали первичные антитела против XIAP, Bcl-2 (DAKO, Glostrup, Denmark), Bax (Sigma, St. Louis, МО), Измерения клеточного роста и жизнеспособности или смерти Ингибирование роста клеток Н460 определяли способом колориметрического МТТ анализа жизнеспособности/пролиферации клеток. Клетки обрабатывали раствором, содержащим липосомно-нуклеотидные комплексы в течение 4,5 часов, затем инкубировали 48 часов при температуре 37°С в среде, не содержащей транфекционный препарат или олигонуклеотиды с/без противоопухолевых препаратов. МТТ (25 µг на лунку) добавляли в каждую лунку, после чего планшеты инкубировали в течение 3 часов при температуре 37°С. После этапа инкубации окрашенный формазаном продукт растворяли путем добавления 200 мл DMSO. Считывание информации с планшетов производилось с помощью ридера с длиной волны 570 нм. Процент выживших клеток в лунках, обработанных олигонуклеотидами, стандартизировали по значениям необработанного контроля. Все анализы проводили троекратно. Анализ апоптоза проточной цитометрией Клетки обрабатывали липосомно-олигонуклеотидными комплексами в течение 4,5 часов и инкубировали 48 часов в среде, не содержащей трансфекционный препарат (реагент) при температуре 37°С. После инкубации производили сбор клеток, их дважды промывали пробным буфером (0.5% глюкоза в PBS без содержания Са++ и Мg++), после чего фиксировали в холодном 70% этиловом спирте при температуре 4°С не менее 18 часов. Образцы центрифугировали при скорости 3000 об/мин в течение 10 минут, затем формировали суспензию в типовом буферном растворе, содержащем 50 µг/мл пропидия йодида (PI) и 400 ед/мл РН-азы А. Образцы инкубировали в течение 30 минут при комнатной температуре и 30 минут на льду, после чего производили проточно-цитометрический анализ. Для создания гистограмм, по которым определяли распределение клеток по фазам клеточного цикла, использовали программу EXPO (Applied Cytometry Systems, Sacramento, СА). Субфракция клеток G1/G0 была принята за типичные апоптозные клетки. Ядерная морфология Клетки подвергали воздействию липосомно-олигонуклеотидными комплексами в течение 4,5 часов, после чего инкубировали 48 часов в среде, не содержащей трансфекционный препарат или олигонуклеотиды при температуре 37°С. После сбора клеток их окрашивали 0,10 µг/мл DAPI (4′, 6-диамидино-2-2-фенилиндол) в течение 30 минут при комнатной температуре. Клетки помещалиь на стекло, цитоспинировали и исследовали под микроскопом Leica с 40Х объективом при УФ флуоресцентным освещением. Изображения переводили в цифровой формат с помощью программы Imagedatabase V. 4.01 Software (Leica, Германия). Противоопухолевое действие in vivo. Эксперименты по эффективности проводились на самцах мышей-носителей опухоли NCI-H460 с иммунодефицитом, составляющих группу RAG2, или самках мышей-носителей опухоли LCC6, относящихся к той же группе. Лечение начинали на 3 день после инокуляции. Физраствор (контрольная группа), олигонуклеотиды G4 AS (5 или 15 мг/кг) или олигонуклеотиды G4 SC (5 или 15 мг/кг) вводили ежедневно по 5 доз в неделю на протяжении трех недель. Винорельбин (VNB, 5 или 10 мг/кг) вводили в вену хвоста отдельно либо в комбинации с олигонуклеотидами на 3, 7, 11 или 17 день после инокуляции. Когда олигонуклеотиды вводили в комбинации с VNB, лекарственную терапию проводили через четыре часа после введения ODN. Осмотр мышей проводили ежедневно. Для выявления возможной токсичности использовали показания массы тела и признаки стресса (например, летаргия, взъерошенная шерсть, атаксия). Животных с изъязвившимися опухолями или опухолями, объем которых превышал 1 см3, умерщвляли. Данные, полученные при измерении опухолей цифровым штангенциркулем, переводили в средний размер опухоли (см3) с помощью формулы: 1/2 [длина(см) × [ширина(см)]2. Средний размер опухоли для одной мыши использовали для вычисления группового среднего размера опухоли ± SE (n=6 мышей), полученного в результате минимум двух независимых экспериментов для каждой группы. Обработка опухоли и ткани Опухоли мышей извлекали на 21 или 14 день после инокуляции и лечения. Одну порцию опухолевой ткани фиксировали в формалине. Из пропитанных парафином тканей изготавливали срезы, которые подвергали патогистологическому макроисследованию после окраски гематоксилин-эозином и иммуногистохимическому исследованию на выявление экспрессии убиквитина. Оставшиеся порции опухоли гомогенизировали в лизирующем буфере для проведения вестерн-блот анализа (см. выше). Статистический анализ Для определения статистической значимости между двумя группами лечения использовали тест Стьюдента. Множественное сравнение производили с помощью однофакторного дисперсионного анализа (ANOVA) и post-hoc теста, в ходе которых было произведено сравнение групп с различными вариантами лечения с использованием критерия Shelle (Statistics release 4.5, StatSoft Inc., Tulsa, OK). Данные принимали за значимые при величине Р<0,05. Пример 14: Снижение экспрессии HIAP РНК и полипептида под воздействием антисмыслового олигонуклеотида НIАР1 Библиотеку, включающая 15 антисмысловых олигонуклеотидов, исследовали на наличие нокдауна белка, полученного при вестерн-блот анализе и нокдауна РНК, полученного при анализе TaqMan с использованием праймеров и зондов, описанных выше в Примере 3 при двух различных условиях. Уровни НIАР1 РНК могут быть определены/при стандартном Нозерн-блот анализе или способах RT-PCR. Антисмысловые олигонуклеотиды вводили в клетку при базовых условиях или в условиях индукции циклогексимидом (СНХ, лечение субтоксическими дозами на протяжении 24 часов). Циклогексимид (СНХ) приводит к 10-100 кратному усилению индукции HIAP mPHK в зависимости от обрабатываемой линии клеток. Это, в свою очередь, ведет к повышению уровня белка НIАР1, как видно из результатов вестерн-блот анализа (полоса 70 кДа). Этот эффект обусловлен двумя различными механизмами действия. Первый заключается в том, что СНХ активирует NFkB, известный фактор транскрипции НIАР1, путем блокирования синтеза de novo неустойчивого белка IkB, который является ингибитором NFkB. Этот эффект имитируется пуромицином, другим ингибитором синтеза белка и TNF-альфа (фактором некроза опухолей альфа), который индуцирует сигнальный каскад ведущий к фосфорилированию, убиквитинированию и распаду IkB. Только СНХ ведет к дальнейшей стабилизации НIАР1 mPHK, как видно по снижению скорости исчезновения переноса НIАР1 в присутствии актиномицина D, что необходимо для блокирования повторной транскрипции, а также СНХ, в отличии от результатов, полученных при применении комбинации актиномицина D и пуромицина или TNF-альфа. Клетки глиобластомы SF295 трансфицировали липофектином и олигонуклеотидами (случайно изменный сурвивин, без олигонуклеотида, антисмысловые АРО 1- АРO15) либо не подвергали обработке. Через 6 часов после трансфекции из клеток выделяли РНК, измеряли уровень НIАР1 mPHK способом количественной PCR (анализ TaqMan) и производили усреднение значений до GAPDH mPHK, причем значения трансфекции олигонуклеотидом сурвивин, отобранным способом случайной выборки, принимали за 1,0. Результаты этого эксперимента, представляющие собой сведения трех отдельных экспериментов, показаны на Фиг.21. Клетки с олигонуклеотидом сурвивина, изменным случайным образом, холостой трансфекцией и нетрансфицированные клетки продемонстрировали одинаковые уровни НIАР1 mРНК. Из 15 олигонуклеотидов семь (АРО 1, 2, 7, 8, 9, 12, 15) показали практически 50% снижение уровня по сравнению с холостой трансфекцией или контрольной трансфекцией случайно изменным сурвивином (5′-mUmAmAGCTGTTCTATGTGmUmC-3′; SEQ ID NO: 296) (Фиг.21). Некоторые олигонуклеотиды вызвали индукцию в НIАР1 mРНК, которая, возможно, являлась стрессовой реакцией на не специфический токсический олигонуклеотид. Антисмысловой олигонуклеотид, тем не менее, может быть эффективен в отношении нокдауна уровней белка НIАР1 даже при возрастании уровня его экспрессии, если олигонуклеотид способен влиять на процесс трансляции. Действие антисмыслового олигонуклеотида HIAP 1 на экспрессию белка НIАР1 и mРНК также исследовали на клетках, в которых была индуцирована экспрессия НIАР1. Производили трансфекцию клеток SF295 олигонуклеотидами или их холостую трансфекцию. После этого трансфицированные клетки подвергали воздействию 10 µг/мл циклогексимида на протяжение 24 часов с целью индукции 70 кДа НIАР1 mРНК и белка. Уровни белка определяли вестерн-блот анализом с анти-НIАР1 поликлональным антителом, после чего производили стандартизацию результатов по отношению к актину при повторном зондировании тех же самых блотов. Развертки результатов вестерн-блот анализа показаны на Фиг.22А. Результаты денситометрии приведены в виде диаграммы в соотношении с результатами холостой трансфекции (принятыми за 100%) на Фиг.22В. На уровне 50% проведена линия, облегчающая идентификацию наиболее эфективных антисмысловых олигонуклеотидов. Сам процесс трансфекции (например, холостая трансфекция или трансфекция случайно изменным сурвивином), как показал вестерн-иммуноблот анализ, вызывает индукцию белка НIАР1 по сравнению с необработанными образцами. Из 15 исследованных нуклеотидных олигомеров, шесть (АРО 1, 2, 7, 8, 12, и 15) проявили высокую активность или обладали значительной активностью в отношении как белка, так и mРНК, не вызывая при этом повышения НIАР1 mРНК, вызванного стрессом, как было в случаях с использованием нуклеотидных олигомеров АРО 4, 6, 11, 13, 14 (Фиг.21), и контрольными олигонуклеотидами до АРО 2 (несоответствия или обратная полярность, см. ниже описание и Фиг.23 и 24). Следует отметить, что АРО 6 также проявил признаки токсичности, как видно по общему возрастанию тотального белка (Фиг.23). Для дальнейшего исследования эффективности антисмысловых нуклеотидных олигомеров Н1АР1 в условиях индукции циклогексимидом, изменения в НIАР1 mРНК измеряли с помощью анализа Taqman PCR в реальном времени через 6 часов после трансфекции АРО 2, который непсредственно связывается с Alu повторами в интроне НIАР1 и приводит, таким образом, к сильнейшей блокаде СНХ-индуцированной повышающей регуляции (стимуляции) НIАР1 mРНК и белка. Контролем в этом эксперименте служили три олигонуклеотида, соответствующие АРО 2: одна случайная последоватеьность (тот же состав нуклеотидов, но произвольный порядок; 5′-AAGGGCGGCGGAGTGAGAC-3′; SEQ ID NO: 297), один олигонуклеотид с обратной полярностью (тот же состав нуклеотидов, тот же порядок последовательности, но в противоположном направлении, 5′-AGAGG ACGGAGTCGGAGGC-3′; SEQ ID NO: 298) и одна последовательность мисматчами (содержащая 4 ошибочно-спаренных нуклеотидов из 19, 5′-CGGAGCGTGAGGATGGAGA-3′; SEQ ID NO: 299). Трансфекция клеток антисмысловым АРО 2 вызвала снижение mРНК на 50% по сравнению с контрольным случайно изменным сурвивином и идеально соответствовала результатам в отношении белка, в то время как SC контроль для АРO2 (Н1 sc аро 2 на Фиг.24) не вызвал абсолютно никаких изменений уровней НIАР1 mРНК (измерения проводились дважды в ходе двух независимых экспериментах). Однако контрольные группы с ODN, содержащим неспаренные основания (Н1 mm аро 2) и обратной полярностью (Н1 RV аро 2) проявили 6-7 кратную индукцию НIАР1 mРНК в течение 6 часов. Эти олигонуклеотиды больше не оказывали воздействия на НIАР1, как ожидалось, но, возможно, продолжали связываться с Alu повторами благодаря дегенерации и повторяемой природы этих последовательностей. Таким образом, существует вероятность того, что эти два контрольных образца токсичны по отношению к клеткам и вызывают стрессовую реакцию, которая приводит к индукции НIАР1. Этот эффект может также наступить при использовании олигонуклеотида АРO2, но в этом случае АРO2 вызывает, кроме того, и деградацию индуцированной НIАР1 mРНК, который приводит к ее относительному снижению в сравнении с контролем со случайно изменным сурвивином, а также к относительному снижению белка НIАР1 после трансфекции и обработкой СНХ, в сравнении с контролем случайно изменным сурвивином. Шесть антисмысловых олигонуклеотида НIАР1 включают два олигонуклеотида, очень эффективных в отношении интронной последовательности (АРО 1, с АРО 2, где АРО 2 проявлял более высокую активность). Эти олигонуклеотиды могут быть использованы для лечения рака или аутоиммунных заболеваний. Олигонуклеотиды, направленные против интронной последовательности действуют только на пре-mРНК (мишень с очень коротким периодом существования), а не на зрелую форму НIАР1. Как правило, интроны не являются мишенями для антисмысловых последовательностей, за исключением случаев, когда антисмысловой олигонулеотид препятствует сплайсингу, повреждая интрон-экзонную границу или акцепторную точку сплайсинга. Это чаще приводит к пропуску зоны экзона, чем к деградации матрицы, вызванному РНазой. Оба механизма оказали бы положительное влияние на стимуляцию апоптоза, поскольку пропуск мог бы привести к потере экзона, кодирующего два первых, важных BIR-домена НIАР1. Антисмысловой ODN АРО 2, также направлен на 18 из 19 последовательных нуклеотидов интрона сурвивина, но при этом мы не наблюдали потери белка сурвивина. При этом отмечали только снижение НIАР1 после воздействия олигонуклеотидами, что свидетельствовало о специфичности антисмыслового олигонуклеотида НIАР1. Эти антисмысловые олигонуклеотиды направлены на Alu последовательности интрона НIАР1 и, потенциально, многих других генов, и индуцируют смерть раковых клеток (см. ниже), что может быть обусловлено понижающей регуляцией НIАР1 и некоторых других критических генов и, таким образом, представляют терапевтическую ценность при условии отсутствия токсического эффекта на здоровые клетки. Раковые клетки, по имеющимся данным, содержат больше Alu-содержащих транскриптов и, следовательно, могут обладать большей чувствительностью к индукции апоптоза олигонуклеотидами, направленными на Alu. Более того, этот убивающий эффект олигонуклеотидов АРO1 и АРО 2 может быть обусловлен сочетанием эффекта действия на Alu и НIАР1 одновременно. Этот двойной эффект может в результате проявляться в виде предотвращения нормальной реакции на стресс в виде индукции НIАР1 через NFkB, когда клетка подвергается воздействию определенных токсических агентов. Эта реакция на стресс, вероятнее всего, является частью анти-апоптозной программы раковой клетки. Блокируя экспрессию НIАР1, мы препятствуем развитию этой антиапоптозной реакции на стресс и ускоряем гибель раковой клетки. Пример 15: Повышение токсичности и хемосенсибилизации антисмысловыми олигонуклеотидами НIАР1 Также нами производилась оценка действия антисмысловых олигонуклеотидов НIАР1 на хемосенсибилизацию клеток SF295. Клетки трансфицировали одним из трех различных антисмысловых олигонуклеотидов (АРО 7, АРО 15 и SC APO2 (контроль)). Через 24 часа после трансфекции олигонуклеотидами клетки инкубировали с адриамицином еще 24 часа, после чего производилась оценка выживаемости клеток и WST-1 (способом WST-1). Кривые выживаемости WST-1 для клеток SF295, трансфицированных вышеописанными олигонуклеотидами НIАР1 и затем подвергшихся воздействию возрастающими концентрациями адриамицина, приведены на Фиг.25. Два олигонуклеотида, вызвавшие снижение НIАР1 mРНК также привели к снижению выживаемости при лечении адриамицином по сравнению с клетками, обработанными олигонуклеотидами, которые не вызывали снижение уровня НIАР1 mРНК. Следовательно, снижение уровня НIАР1 олигонуклеотидами или какими-либо другими способами может приводить к хемосенсибилизации линии клеток глиобластомы, которая отличается высокой резистентностью к цитотоксическому действию многих химиотерапевтических препаратов. Дополнительные 89 последовательностей антисмысловых нуклеотидных олигомеров, которые могут быть задействованы в способах настоящего изобретения, приведены в Таблице 7. Последовательности, которые на 100% идентичны НIАР1 и HIAP2 человека, а также последовательности, включающие одну или две ошибок спаривания нуклеотидов, выделены жирным шрифтом.
Нами также был проанализирован НIАР2 человека на наличие последовательностей, подходящих для использования в качестве антисмысловых нуклеотидных олигомеров. Идентифицированные последовательности представлены в Таблице 8.
Другие антисмысловые олигонуклеотиды IАР, включая описанные в Таблице 2 Патента США
Другие последовательности для антисмысловых олигонуклеотидов IAP, пригодные для примения вспособе, согласно настоящему изобретению, описаны, например, в Патентах США Другие примеры осуществления изобретения Все публикации и патентные заявки, упомянутые в настоящей спецификации, включены путем ссылки в том же объеме, как если бы каждая независимая публикация или патентная заявка были указаны конкретно по отдельности путем ссылки. Хотя настоящее изобретение было описано в связи с конкретными примерами реализации изобретения, очевидно, что возможны дальнейшие его модификации и настоящая заявка охватывает любые варианты, аспекты или воплощения изобретения не отступая в общих чертах от основных принципов изобретения и включая отклонения от настоящего изобретения, находящиеся в рамках известных или традиционных способов, применяемых в области, к которой относится указанное изобретение, и которые могут быть применены к основным признакам изобретения, сформулированным далее.
Формула изобретения
1. Способ лечения больного, страдающего пролиферативным заболеванием, который включает введение указанному пациенту 2. Способ по п.1, отличающийся тем, что указанный химиотерапевтический препарат представляет собой антиметаболический агент. 3. Способ по п.2, отличающийся тем, что указанный антиметаболический агент представляет собой гемцитабин, капецитабин, фторурацил, клофарабин, цитарабин или флюдарабин. 4. Способ по п.1, отличающийся тем, что указанный химиотерапевтический препарат представляет собой карбоплатин. 5. Способ по любому из пп.1-4, отличающийся тем, что указанному пациенту дополнительно вводят второй химиотерапевтический препарат. 6. Способ по п.5, отличающийся тем, что указанный второй химиотерапевтический препарат представляет собой противоопухолевый антибиотик. 7. Способ по п.6, отличающийся тем, что указанный противоопухолевый антибиотик представляет собой доксорубицин или идарубицин. 8. Способ по п.5, отличающийся тем, что указанный второй химиотерапевтический препарат представляет собой таксан. 9. Способ по п.5, отличающийся тем, что указанный таксан представляет собой паклитаксель или доцетаксель. 10. Способ, по любому из пп.1-9, отличающийся тем, что интервал между введением указанного нуклеотидного олигомера и указанного химиотерапевтического препарата составляет не более 28 дней, предпочтительно не более 24 ч, более предпочтительно не более 1 ч, или указанный нуклеотидный олигомер и указанный химиотерапевтический препарат вводят одновременно. 11. Способ по любому из пп.1-9, дополнительно включающий введение указанному пациенту химиосенсибилизирущего препарата или препарата, модулирующего биологический ответ. 12. Способ по любому из пп.1-9, отличающийся тем, что указанное пролиферативное заболевание представляет собой рак. 13. Способ по п.12, отличающийся тем, что указанный рак представляет собой острый лейкоз, острый лимфолейкоз, острый миелобластный лейкоз, острый миелоцитарный лейкоз, промиелоцитарную лейкемию, острый миеломоноцитарный лейкоз, острый эритролейкоз, хронический лейкоз, хронический миелоцитарный лейкоз, миелодиспластический синдром, хронический лимфолейкоз, истинную полицитемию, лимфому, болезнь Ходжкина, макроглобулинемию Вальденстрома, фибросаркому, липосаркому, хондросаркому, остеогенную саркому, хордому, ангиосаркому, эндотелиосакому, лимфангиосаркому, лимфангиоэндотелиосаркому, синовиому, мезотелиому, опухоль Юинга, лейомиосаркому, рабдомиосаркому, рак кишечника, рак поджелудочной железы, рак груди, рак яичника, рак простаты, папиллярную карциному, плоскоклеточный рак, базально-клеточный рак, аденокарциному, рак потовой железы, рак сальной железы, цистаденокарциному, медуллярный рак, рак бронха, почечноклеточный рак, гепатому, рак желчных протоков, хориокарциному, семиному, тератокарциному, опухоль Вильма, рак шейки матки, рак тела матки, рак яичка, рак легких, мелкоклеточный рак легких, рак желчного пузыря, эпителиальную карцинома, глиому, астроцитому, медуллобластому, краниофарингиому, эпендимому, пинеалому, гемангиобластому, невриному слухового нерва, олигодендроглиому, шванному, менингиому, меланому, нейробластому или ретинобластому. 14. Способ по любому из пп.1-9, отличающийся тем, что указанный нуклеотидный олигомер по существу состоит из последовательности SEQ ID NO:151. 15. Способ по любому из пп.1-9, отличающийся тем, что указанный нуклеотидный олигомер состоит из последовательности SEQ ID NO:151. 16. Способ по любому из пп.1-9, отличающийся тем, что указанный нуклеотидный олигомер содержит по меньшей мере одну модифицированную связь. 17. Способ по п.16, отличающийся тем, что указанная модифицированная связь представляет собой фосфоротиоатную связь 18. Способ по любому из пп.1-9, 16, отличающийся тем, что указанный нуклеотидный олигомер содержит по меньшей мере один модифицированный остаток сахара. 19. Способ по п.18, отличающийся тем, что модифицированный остаток сахара представляет собой 2′-O-метил- или 2′-0-метоксиэтил- модифицированный остаток. 20. Способ по любому из пп.1-9, 16, 18, отличающийся тем, что указанный нуклеотидный олигомер содержит, по меньшей мере, одно модифицированное основание. 21. Способ по п.20, отличающийся тем, что указанное модифицированное основание представляет собой 5-метил цитозин. 22. Способ по любому из пп.1-9, 16, 18, 21, отличающийся тем, что указанный нуклеотидный олигомер представляет собой химерный нуклеотидный олигомер. 23. Способ по п.22, отличающийся тем, что нуклеотидный олигомер содержит остатки ДНК, которые соединены между собой фосфоротиоатными связями, причем указанные остатки ДНК фланкированы с каждой стороны, по меньшей мере, одним 2′-O-метилированным или 2′-O-метоксиэтилированным остатком РНК, предпочтительно, по меньшей мере, тремя 2′-O-метилированными или 2′-O-метоксиэтилированными остатками РНК, предпочтительно, четырьмя 2′-O-метилированными или 2′-O-метоксиэтилированными остатками РНК. 24. Способ по п.23, отличающийся тем, что указаные остатки РНК соединены между собой фосфоротиоатными связями, причем указанные остатки РНК соединены с указанными остатками ДНК фосфоротиоатными связями. 25. Способ по п.22, отличающийся тем, что нуклеотидный олигомер содержит остатки ДНК, которые соединены между собой фосфородиэфирными связями, причем указанные остатки ДНК фланкированы с каждой стороны по меньшей мере двумя 2′-O-метилированными или 2′-O-метоксиэтилированными остатками РНК, соединенными фосфоротиоатными связями, предпочтительно по меньшей мере тремя 2′-O-метилированными или 2′-O- метоксиэтилированными остатками РНК. 26. Способ по п.1, отличающийся тем, что указанный нуклеотидный олигомер представляет собой соль Na19 и содержит одиннадцать остатков ДНК, фланкированных с каждой стороны четырьмя 2′-O-метилированными остатками РНК, причем указанный нуклеотидный олигомер состоит из последовательности 27. Способ по любому из пп.1-9, 16, 18, 21, 23-26, отличающийся тем, что указанный нуклеотидный олигомер вводят парэнтерально. 28. Способ по любому из пп.1-9, 16, 18, 21, 23-26, отличающийся тем, что указанный нуклеотидный олигомер находится в форме для внутривенного введения.
РИСУНКИ
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 7, p.2826-2836. Ji J. et al. Human ovariancancer and cisplatin resistance: possible inhibitor of apoptosis protein. Pros. National Acad. Sci. USA, v. 142,
7, p.2826-2836. Ji J. et al. Human ovariancancer and cisplatin resistance: possible inhibitor of apoptosis protein. Pros. National Acad. Sci. USA, v. 142,  М олигонуклеотидов F3, G4, С5, АВ6, DE4 или D7, или соответствующие контрольные трансфекции олигонукелотидами обратной полярности (RP) или смесью олигонуклеотидов (Scrambled control, SC). На протяжение 6 часов трансфекции производилась в режиме реального времени оценка относительного количества XIAP мРНК с помощью RT-PCR. Все данные представлены в виде среднего арифметического ± стандартное отклонение (SD) троекратных измерений, произведенных в ходе репрезентативного эксперимента. Уровень XIAP мРНК клеток, не подвергавшихся никакому воздействию (контрольная группа) определяли в идентичных экспериментальных условиях и принимали за 1.
М олигонуклеотидов F3, G4, С5, АВ6, DE4 или D7, или соответствующие контрольные трансфекции олигонукелотидами обратной полярности (RP) или смесью олигонуклеотидов (Scrambled control, SC). На протяжение 6 часов трансфекции производилась в режиме реального времени оценка относительного количества XIAP мРНК с помощью RT-PCR. Все данные представлены в виде среднего арифметического ± стандартное отклонение (SD) троекратных измерений, произведенных в ходе репрезентативного эксперимента. Уровень XIAP мРНК клеток, не подвергавшихся никакому воздействию (контрольная группа) определяли в идентичных экспериментальных условиях и принимали за 1.
 (BioKeys)
(BioKeys) BC5 as
BC5 as -актин антитела против белка человека, что позволило определить уровни человеческого XIAP из образцов опухолевых клеток без примеси XIAP из мышиных клеток. Уровень белка XIAP в клетках опухолей, обработанных AS олигонуклеотидами G4, значительно снизился примерно до 85% от опухолей контрольной группы (Р<0,005) (Фиг.18А и 18В). Опухоли, обработанные AS олигонуклеотидами G4, сократились в размерах на 24% по отношению к размеру опухолей контрольной группы. Эти результаты свидетельствуют о том, что ингибирование роста опухоли Н460 AS олигонуклеотидами G4 кореллирует с понижающей регуляцией экспрессии белка XIAP.
-актин антитела против белка человека, что позволило определить уровни человеческого XIAP из образцов опухолевых клеток без примеси XIAP из мышиных клеток. Уровень белка XIAP в клетках опухолей, обработанных AS олигонуклеотидами G4, значительно снизился примерно до 85% от опухолей контрольной группы (Р<0,005) (Фиг.18А и 18В). Опухоли, обработанные AS олигонуклеотидами G4, сократились в размерах на 24% по отношению к размеру опухолей контрольной группы. Эти результаты свидетельствуют о том, что ингибирование роста опухоли Н460 AS олигонуклеотидами G4 кореллирует с понижающей регуляцией экспрессии белка XIAP.