Патент на изобретение №2349601
|
||||||||||||||||||||||||||
(54) ГОНАДОТРОПНЫЙ ГОРМОН, ПОЛУЧЕННЫЙ ИЗ БЕСПОЗВОНОЧНЫХ, И ЕГО СИНТЕЗ
(57) Реферат:
Изобретение относится к биотехнологии, конкретно к получению гормонов, и может быть использовано в разведении беспозвоночных. Из беспозвоночного Asterina pectinifera выделяют гонадотропный гормон, представляющий собой пептид с молекулярной массой 4500-4900 и состоящий из двух субъединиц, белковая структура которого связана SS-мостиками, образованными между остатками SH цистеинов, содержавшихся в субъединицах. Смешивание и окисление этих двух субъединиц после их синтеза позволяет получить гонадотропный гормон, обладающий гонад-стимулирующей активностью. 3 н. и 1 з.п. ф-лы, 8 ил., 1 табл.
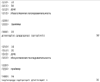
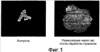
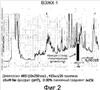
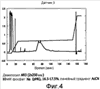
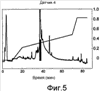
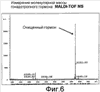
Область техники, к которой относится изобретение Изобретение относится к гонадотропному гормону, полученному из беспозвоночных, таких как Asterina pectinifera и т.п., и его синтезу. Уровень техники Существует два вида гонадотропных гормонов позвоночных животных, которые управляют развитием половых органов и развитием, созреванием и высвобождением яйцеклеток и сперматозоидов. Сделано значительное продвижение в изучении гонадотропных гормонов позвоночных животных, например, рыб и т.п. Структурное подобие этих гормонов хорошо сохраняется от рыб к людям, и эти гормоны являются белковыми гормонами с гетеродимерной структурой, включающей Было подтверждено, что гонадотропные гормоны позвоночных оказывают межвидовое воздействие (непатентная ссылка 1, патентные ссылки 1 и 2) на позвоночных животных, но не на беспозвоночных. Гонадотропные гормоны беспозвоночных не были настолько изучены, как гонадотропные гормоны позвоночных. Гонадотропный гормон (GSS) был экстрагирован из радиального нерва Asterina pectinifera, то есть беспозвоночного, и, как было подтверждено, вызвал размножение из овариального компонента (непатентные ссылки 2 и 3). Сообщается, что гонадотропный гормон стимулирует овариальные фолликулярные клетки, окружающие яйцеклетки в яичнике, к синтезу и секреции гормона, индуцирующего созревание ооцита (1-метиладенина), непосредственно влияющего на яйцеклетки (непатентная ссылка 4). Патентная ссылка 1: Japan patent No. 2967945. Патентная ссылка 2: Japan patent Application Public Disclosure No. H06-107689. Непатентная ссылка 1: Ed., The Japanese society for comparative endocrinology, “Biological Science of Hormone 5, Hormone and Reproduction (II)”, Japan Scientific Societies Press, pp. 41-47, 1979. Непатентная ссылка 2: Ed., The Japanese society for zoology, “Problems in Modern Zoology 4, Oocytes and Sperms”, Tokyo University Press, pp. 21-37, 1975. Непатентная ссылка 3: Shirai H., Gonad-Stimulating and Maturation-Inducing Substance, “Method in Cell Biology” Academic Press, vol. 27, pp. 73-88, 1986. Непатентная ссылка 4: Mita M. & Nagahama Y., Involvement of G-proteins and adenylate cyclase in the action of gonad-stimulating substance on starfish ovarian follicle cells, Developmental Biology, 1991, 144, 262-8. Задачи, решаемые настоящим изобретением Так как гонадотропные гормоны оказывают стимулирующий эффект на созревание, их важно применять для культивирования материнского живого организма. Настоящее изобретение основано на инновационном раскрытии структуры гонадотропного гормона беспозвоночного и может найти перспективное применение в культивировании беспозвоночных, например, креветок, крабов, морских ракушек и т.п. Способы решения задач Авторы изобретения успешно проанализировали структуру гонад-стимулирующего гормона, секретируемого нервными клетками Asterina pectinifera. Изобретатели впервые показали, что гонадотропный гормон – это пептид с молекулярной массой 4500-4900, состоящий из субъединиц с молекулярной массой 2000-2400 и 2400-2600, и что его структура образована SS-мостиками, сформированными между SH-остатками цистеинов, содержащимися в этих двух субъединицах, и подтвердили, что пептид, полученный при смешивании и окислении двух синтезированных субъединиц, имел гонад-стимулирующую активность. Результат, проанализированный авторами настоящего изобретения, может открыть новые возможности для широкого диапазона прикладных областей, чтобы наладить массовое производство гормона и др. А именно, настоящее изобретение представляет собой гонадотропный гормон беспозвоночного, включающий следующие два пептида, в которых SS-мостики сформированы между 6 цистеинами этих двух пептидов: (a) пептид, имеющий аминокислотную последовательность SEQ ID NO: 1, или аминокислотную последовательность, включающую делецию, замену или добавление одной или нескольких аминокислот (то есть 2-3) в указанной аминокислотной последовательности кроме аминокислот 4 и 16 (Cys) указанной аминокислотной последовательности, и обладающий гонад-стимулирующей активностью; (b) пептид, имеющий аминокислотную последовательность SEQ ID NO: 2, или аминокислотную последовательность, включающую делецию, замену или добавление одной или нескольких (то есть 2-3) аминокислот в указанной аминокислотной последовательности кроме аминокислот 10, 11, 15 и 24 (Cys) указанной аминокислотной последовательности, и обладающий гонад-стимулирующей активностью. Также настоящее изобретение представляет собой гонадотропный гормон беспозвоночного, полученный путем смешивания и окисления двух следующих пептидов: (a) пептид, имеющий аминокислотную последовательность SEQ ID NO: 1, или аминокислотную последовательность, включающую делецию, замену или добавление одной или нескольких (то есть 2-3) аминокислот в указанной аминокислотной последовательности (предпочтительно аминокислоты кроме Cys); и обладающий гонад-стимулирующей активностью; (b) пептид, имеющий аминокислотную последовательность SEQ ID NO: 2, или аминокислотную последовательность, включающую делецию, замену или добавление одной или нескольких (то есть 2-3) аминокислот в указанной аминокислотной последовательности (предпочтительно аминокислоты кроме Cys), и обладающий гонад-стимулирующей активностью. Эти два пептида могут быть получены путем очистки (a) нервного органа, выделенного из беспозвоночного, или (b) определенного органа, который, как сообщалось, обладает гонад-стимулирующей активностью, основанной на гонад-стимулирующей активности беспозвоночного в качестве индикатора. Кроме того, настоящее изобретение представляет собой способ получения пептидов с гонад-стимулирующей активностью, включающий смешивание и окисление двух указанных пептидов. Эти два пептида могут быть получены любым способом, то есть они могут быть химически синтезированными или могут быть получены при помощи генной инженерии. Настоящее изобретение представляет собой ДНК, кодирующую любой из двух указанных пептидов ((c) и (d)). Пептиды выделяют из организмов-хозяев или среды культивирования, при этом эти организмы-хозяева трансформируют вектором, содержащим две вышеупомянутые ДНК, культивируют и выращивают в культуральной среде. Пептид с гонад-стимулирующей активностью может быть получен путем смешивания и окисления пептидов. Кроме того, настоящее изобретение представляет собой ДНК, которая содержит ДНК, кодирующую два указанных пептида ((c) и (d)), с более чем 70% гомологии с нуклеотидной последовательностью SEQ ID NO: 3. Пептиды c гонад-стимулирующей активностью могут быть получены путем культивирования и выращивания организма-хозяина, трансформированного вектором, содержащим ДНК. Кроме того, пептиды могут быть выделены из организмов-хозяев или их среды культивирования, при этом пептиды могут быть смешаны и окислены. Краткое описание чертежей Фиг.1 – фотография, показывающая критерии гонад-стимулирующей активности. Правая сторона показывает, что яйцеклетки зародились и гонад-стимулирующая активность присутствует. Фиг.2 показывает профиль очистки гонадотропного гормона высокоэффективной жидкостной хроматографией. Фиг.3 показывает профиль очистки гонадотропного гормона высокоэффективной жидкостной хроматографией. Фиг.4 показывает профиль очистки гонадотропного гормона микродозовой высокоэффективной жидкостной хроматографией. Фиг.5 показывает профиль очистки гонадотропного гормона микродозовой высокоэффективной жидкостной хроматографией. Фиг.6 показывает результат анализа очищенного гонадотропного гормона (гетеродимера) на масс-спектрометре. Фиг.7 показывает результат анализа субъединиц гонадотропного гормона на масс-спектрометре. Фиг.8 показывает профиль очистки синтезированного гонадотропного гормона высокоэффективной жидкостной хроматографией. Подробное описание изобретения Последовательность ДНК гена гонадотропного гормона беспозвоночного согласно настоящему изобретению включает нуклеотидную последовательность (351 основание) SEQ ID NO:3, ДНК, кодирующую два указанных пептида ((c) и (d)), или последовательности нуклеотидов, которая является гомологичной более чем на 70% нуклеотидной последовательности SEQ ID NO:3. В докладе по проекту анализа генома человека консорциума генома человека (Science 2001, 291 (5507), 1304-1351) предложен критерий автоматической аннотации гена в последовательности генома с помощью компьютерной программы (Otto system), причем этот критерий, основанный на гомологии к полинуклеотидной последовательности с известной генетической информацией, должен обладать более чем 92%-ной идентичностью для того, чтобы точно определить область гена в геноме. В том же докладе предлагается критерий гомологии полинуклеотида на 70%, для отыскания геномов, увеличенных генной дупликацией (более поздняя эволюция могла ввести генную мутацию и могла содействовать частичным изменениям нуклеотидной последовательности). Поэтому нуклеотидная последовательность с гомологией больше чем 70%,и предпочтительно, больше чем 92%, к SEQ ID NO:3 в настоящем изобретении может функционировать как ген гонадотропного гормона беспозвоночного, который применяется к тесно связанным, но различным разновидностям животных. Аминокислотная последовательность гонадотропного гормона беспозвоночного согласно настоящему изобретению включает 116 аминокислот, показанных в SEQ ID NO: 4, или аминокислотную последовательность, в которой одна или несколько (например, 2-3) аминокислот (предпочтительно аминокислоты кроме Cys) делетированы, заменены или добавлены. В аминокислотной последовательности (SEQ ID NO: 4) последовательность, начиная с аминокислоты 1, метионина (M), до аминокислоты 29, глицина (G), является сигнальной последовательностью, последовательность, начиная с аминокислоты 30, глутамина (E), до аминокислоты 48, серина (S), является субъединицей А (GSS-A), последовательность, начиная с аминокислоты 49, лизина (K), до аминокислоты 92, аргинина (R), является субъединицей C (GSS-C), и последовательность, начиная с аминокислоты 93, серина (S), до аминокислоты 116, цистеина (C), является субъединицей B (GSS-B). Гонадотропный гормон беспозвоночного согласно настоящему изобретению содержит пептид, сформированный связыванием GSS-A и GSS-B 1 к 1, при этом пептид содержит SS-мостики, окислительно сформированные между цистеинами (аминокислоты 4 и 16 из SEQ ID NO: 1 и аминокислоты 10, 11, 15 и 24 из SEQ ID NO: 2), содержащимися в каждом пептиде. Максимальная возможность образования SS-мостиков между GSS-A и GSS-B составляет 12. Гетеродимерная структура с мостиковой конструкцией между пептидами GSS-A и GSS-B может быть собрана в виде гонадотропного гормона после удаления сигнальной последовательности из цепи пептида, синтезированной на основе гена, с последующим заключительным удалением GSS-C части от мостиковой структуры. Однако GSS-C часть все еще может также иметь физиологическую активность согласно недавней научной публикации. Так как все гонадотропные гормоны беспозвоночных согласно настоящему изобретению секретируются из нервных тканей, источник гонадотропных гормонов беспозвоночных от других видов, кроме Asterina pectinifera, был ограничен нервными тканями исходных видов. Если другие виды животных, как сообщается, обладают гонад-стимулирующей активностью в других тканях и органах, эти ткани и органы могут использоваться как источники для экстракции. Беспозвоночные включают много полезных морских видов, таких как кишечнополостные, например кораллы, иглокожие, например морские ежи и морские слизняки, моллюски, например осьминоги и кальмары, и ракообразные, например креветки и крабы. Пептиды могут быть очищены из указанных источников путем экстракции с использованием гонад-стимулирующей активности в качестве маркера. Очистка может проводиться любыми способами, могут использоваться как жидкостная хроматография, так и двухфазовый водный метод разделения. Однако для высокой очистки предпочтительно используется высокоэффективная жидкостная хроматография. Гель-фильтрация, ионообменная хроматография или обращенно-фазовая хроматография могут использоваться в качестве колонок. Таким образом, фракции, разделенные методом очистки, подвергаются отбору с использованием в качестве маркера гонад-стимулирующей активности. Гонад-стимулирующая активность может быть измерена следующими методами: Те особи, яйцеклетки которых очень чувствительны к гормону, стимулирующему созревание ооцита, выбираются из зрелых женских особей беспозвоночных и отделяются как контрольные особи. У контрольных особей извлекаются яичники в морскую воду и разрезаются на маленькие части. Аналитический планшет для большого числа исследуемых проб заполняют 200 мкл морской воды. Два мкл пробы, исследуемой на гонад-стимулирующую активность, разбавляют в 200 раз, смешивая с 398 мкл морской воды. Половину пробы (200 мкл) разбавляют в два раза, смешивая с 200 мкл морской воды на аналитическом планшете. Половину пробы (200 мкл) далее разбавляют в два раза, смешивая с 200 мкл морской воды на аналитическом планшете. Повторение процедуры приводит к получению серий двукратного разведения от 200-кратного разведения до 102,400-кратного разведения. Маленькую часть яичника добавляют к каждой разведенной пробе и оставляют при температуре 25°C. Гонад-стимулирующую активность оценивают по образованию зрелых яйцеклеток из отрезанной маленькой части яичника после 1 часа. Относительную гонад-стимулирующую активность оценивают по степени разбавления. Гонадотропный гормон имеет много функций, а именно длительные и короткие периоды (долгосрочный: развитие гонады, краткосрочный: индукция выброса яйцеклеток). Так как анализ зависит от краткосрочной функции, контрольные особи должны нести зрелые ооциты со способностью к продукции яйцеклеток. Предпочтительно отбирать контрольные особи в их ранний период нереста и поддерживать их в лабораторном аквариуме. Не всегда все собранные особи настолько зрелые, чтобы производить икру. Кроме того, если особи когда-либо являются способными к производству икры, степень их зрелости едва отличается, и обычно чувствительность к гормону варьируется в зависимости от особи. Поэтому для оценки гонад-стимулирующей активности необходимо отбирать зрелые особи (те, которые несут ооциты со способностью к производству икры) и, кроме того, готовить несколько особей с одинаковой чувствительностью к гормону. Гонадотропный гормон стимулирует овариальные клетки фолликула, при этом клетки фолликула выделяют другой гормон – “гормон, стимулирующий созревание ооцита”, который стимулирует ооциты к производству икры. Так как гормон, стимулирующий созревание ооцита, непосредственно влияет на ооциты, одна из краткосрочных функций гонадотропного гормона заключается в индукции образования гормона созревания ооцита. Для оценки чувствительности может использоваться коммерчески доступный гормон, стимулирующий созревание ооцита. Вышеупомянутая очистка может быть повторена несколько раз. В качестве результата может быть получен уникальный пептид с молекулярной массой 4500-4900. Как показано в следующих Примерах, пептид включает две субъединицы (GSS-A и GSS-B), где молекулярная масса субъединиц – 2000-2400 и 2400-2600. Эти субъединицы могут быть получены при восстановлении вышеупомянутого пептида с высокой молекулярной массой. Различные восстановители используются для восстановительной реакции, при этом предпочтительно используются такие умеренные восстановители, как дитиотреитол, 2-меркаптоэтанол, тиогликолевая кислота, тиофенол, паратиокрезол и другие. Напротив, смешивание и окисление этих двух пептидов приводят к образованию пептида (т.е. гонадотропного гормона настоящего изобретения) с более высокой молекулярной массой, чем вышеописанные пептиды. Так как каждая из этих двух субъединиц содержит цистеины, окисление дает SS-мостики между SH-группами цистеинов, содержащихся в субъединицах, чтобы связать эти две субъединицы. Вышеупомянутое окисление может быть выполнено следующими окислителями кроме способа, показанного в Примерах: – o-Иодобензойная кислота, иод – Диоксид марганца, перманганат калия – Перекись водорода – Свободный кислород (это реагент, используемый в следующих Примерах, реакция окисления свободным кислородом может быть ускорена в присутствии следовых количеств иона и ионов меди). Кроме того, возможно увеличить выход образования правильных SS-мостиков между пептидами в присутствии следующих реактивов, которые используются для рефолдинга SS-мостиков белка во время реакции окисления синтетических пептидов: – тиоредоксин (белок, используемый для окислительно-восстановительного взаимодействия в живой системе), – протеин дисульфид изомераза (фермент обмена дисульфидов белка в живых организмах), – BMC, (±)-транс-1,2-бис(2-меркаптоацетамидо)циклогексан, – 4-меркаптофенилацетат. Пептид согласно настоящему изобретению может также быть получен генной рекомбинацией. Например, вектор, содержащий ДНК (т.е. основания 88-144 и 277-348 из SEQ ID NO: 3), кодирующую аминокислотные последовательности из SEQ ID NOs: 1 и 2 или аминокислотную последовательность, в которой одна или несколько (то есть 2-3) аминокислот кроме Cys делетированы, заменены или добавлены, или вектор, содержащий ДНК, в которой нуклеотидная последовательность более чем на 70% гомологична нуклеотидной последовательности SEQ ID NO: 3, создают, вводят в организмы-хозяева с целью трансформации. Затем трансформированные организмы-хозяева культивируют и выращивают. Целевые пептиды выделяют из организмов-хозяев или их культуральной среды. Иногда полученные пептиды представляют собой пептиды со связью между двумя субъединицами (GSS-A и GSS-B). Однако окисление полученных пептидов (две субъединицы) в присутствии указанных окислителей приводит к пептидам с гонад-стимулирующей активностью. Способы стимуляции ускорения созревания и овуляции при помощи полученных пептидов с гонад-стимулирующей активностью включают введение этих пептидов непосредственно во вторичную полость или яичник беспозвоночных путем добавления к морской воде в аквариуме или смешивания с кормом. Следующие Примеры приводятся для пояснения настоящего изобретения, но не предназначены для ограничения его объема. В следующих Примерах гонад-стимулирующая активность исследована следующим путем: Две-три особи, несущие ооцит с высокой чувствительностью к гонадотропному гормону (1-метиладенину), отобирают из нескольких десятков зрелых женских особей Asterina pectinifera (или Asterias amurensis) и отделяют как контрольные особи. Выбор был осуществлен следующим образом: Гормон, стимулирующий созревание ооцитов, был растворен в морской воде в 6 различных концентрациях, т.е. 10-6 M, 3×10-7 M, 10-7 M, 3×10-8 M, 10-8 M и 3×10-9 M, при этом маленький фрагмент яичника был добавлен к различным концентрациям вызывающего созревание гормона и выдержан 60 минут при комнатной температуре. Созревание ооцита было исследовано под микроскопом после 60 минут, чтобы увидеть, была ли структура ядер в ооците вырождена для подготовки к следующему оплодотворению. Была проверена концентрация гормона, стимулирующего созревание ооцита при созревании, и те особи, которые созревали при меньшей, чем 10-7 М концентрации, использовались как контрольные особи. Яичники (с пучкообразной морфологией) были извлечены у контрольных особей в морскую воду и разделены на фрагменты размером около 5 мм. Аналитический планшет для большого числа исследуемых проб заполняли 200 мкл морской воды. Два мкл пробы, исследуемой на гонад-стимулирующую активность, разбавляли в 200 раз, смешивая с 398 мкл морской воды. Половину пробы (200 мкл) разбавляли в два раза, смешивая с 200 мкл морской воды на аналитическом планшете. Половину пробы (200 мкл) далее разбавляли в два раза, смешивая с 200 мкл морской воды на аналитическом планшете. Повторение процедуры приводит к получению рядов двукратного разбавления от 200-кратного до 102,400-кратного разбавления. Маленькую часть яичника добавляют к каждой разведенной пробе и оставляют при температуре 25°C. Гонад-стимулирующую активность оценивают по образованию зрелых яйцеклеток из отрезанной маленькой части яичника после 1 часа. Относительную гонад-стимулирующую активность оценивают с учетом степени разбавления. Пример 1 Ткани радиального нерва были извлечены из Asterina pectinifera щипцами, заморожены на сухом льду и помещены на хранение. Нервные ткани влажным весом 126,3 г были отобраны у 5550 морских звезд. Жидким азотом заполнили пушку и ткани радиального нерва, сохраненные в замороженном состоянии, были раздроблены в порошок. Порошкообразные нервные ткани были добавлены к 600 мл 10 мМ водного раствора ацетата аммония (содержащего ингибиторы протеиназ, т.е. 1 После того, как полученный супернатант был заморожен и высушен, его растворили в 100 мл 0,15 М карбоната аммония. После того, как нерастворенные остатки удалили центрифугированием при 27,500xg при 4°C в течение 30 минут, раствор нанесли на обессоливающую колонку PD-10 (Amersham Biotech Co.), уравновешенную раствором карбоната аммония, и выделили фракцию элюата с высокой молекулярной массой (фракция PD-10), обладающую гонад-стимулирующей активностью. После того, как фракция элюата с высокой молекулярной массой (фракция PD-10) была заморожена и высушена, ее растворили в 150 мл 10 мМ фосфата натрия (pH 7,0). Раствор нанесли в три приема порциями по 50 мл на колонку Sephadex G-50 (500 см3), уравновешенную фосфатом натрия, и выделили фракцию (фракция без белков высокой молекулярной массы) с гонад-стимулирующей активностью (фракция G-50, выход составил приблизительно 600 мл). Приблизительно 415 мл фракции G-50 были обработаны высокоэффективной жидкостной хроматографией (Shimazu Corporation, Type LC-6AD) в 26 приемов. Затем активные фракции были элюированы линейным градиентом концентрации от 10 мМ фосфата натрия (pH 7,0) до 30% ацетонитрила/10 мМ фосфата натрия с колонки Девелосил RP-Aqueous AR5 (10Ч250 мм, Nomura Science). Фракции с гонад-стимулирующей активностью были элюированы в пределах концентрации ацетонитрила 18-19% (1-я фракция ВЭЖХ, фиг.2). 1-я фракция ВЭЖХ была сконцентрирована до 45 мл под уменьшенным давлением, и концентрат обработали высокоэффективной жидкостной хроматографией (Shimazu Corporation, Type LC-6AD) в 9 приемов. Затем, активные фракции были элюированы в линейном градиенте концентрации от 20% ацетонитрила/10 мМ триметиламинацетата (pH 4,0) до 25% ацетонитрила/10 мМ триметиламинацетата с колонки Девелосил RP-Aqueous AR5 (10×250 мм, Nomura Science). Фракции с гонад-стимулирующей активностью были элюированы в пределах концентрации ацетонитрила 21-22% (2-я фракция ВЭЖХ, фиг.3). 2-я фракция ВЭЖХ была сконцентрирована под пониженным давлением. Активные фракции были элюированы в линейном градиенте концентрации от 16,5% ацетонитрила/10 мМ фосфата натрия (pH 6,0) до 17,5% ацетонитрила/10 мМ фосфата натрия (pH 6,0) с колонки Девелосил RP-Aqueous AR5 (10×250 мм, Nomura Science). Фракции с гонад-стимулирующей активностью были элюированы в пределах концентрации ацетонитрила 17% (3-я SMART фракция, фиг.4). Активные фракции были элюированы в линейном градиенте концентраций от 15% ацетонитрила/10 мМ фосфата натрия (pH 6,0) до 30% ацетонитрила/10 мМ фосфата натрия (pH 6,0) с колонки Девелосил RP-Aqueous AR5 (10×250 мм, Nomura Science). Фракции с гонад-стимулирующей активностью были элюированы в пределах концентрации ацетонитрила 18% (4-я SMART фракция, фиг.5). Часть последней очищенной фракции (4-я SMART фракция) обессолили при помощи Ziptip (Milipore Co.) и проанализировали на масс-спектрометре типа MALDI-TOF (Bruker Daltonics Co., ReflexIII type). Результат показал наличие компонента с молекулярной массой 4737. Кроме того, последняя фракция была восстановлена 50 мМ дитиотреитола при комнатной температуре в течение 1 часа и снова была проанализирована масс-спектрометром. Результат показал наличие двух сигналов с массами 2236 и 2507 вместо 4737. Более того, последнюю фракцию обработали восстановителем и алкилирующим агентом, типа 0,2 М иодацетамида при комнатной температуре в течение 24 часов. Так как молекулярные массы пиков 2236 и 2507 были увеличены на 57 (в соответствии с 1 остатком SH) и 114 массовых единиц (в соответствии с 2-мя остатками SH) соответственно, остатки SH могли быть специфично алкилированы. Анализ 4-й SMART фракции секвенатором белка (ABI, Procise Type 494HT) дал два сигнала, которые указывают на смесь двух пептидов. Так как вышеупомянутые результаты показывают, что компонент с гонад-стимулирующей активностью, содержавшийся в последней фракции, был полипептидом с молекулярной массой 4737, и компоненты были разделены на два компонента восстановлением, то активный компонент имеет гетеродимерную структуру, включающую две пептидные субъединицы с молекулярной массой 2236 и 2507, где связь димерной структуры чувствительна к восстановителю и, как показано, является SS-мостиком между остатками цистеина. Аминокислотный анализ смеси пептидов молекулярной массы 2236 и 2507, полученный восстановлением полипептида с молекулярной массой 4737, был выполнен при помощи масс-спектрометрии на масс-спектрометре типа Q-TOF (Micromass Co.) (фиг.6 и фиг.7). Аминокислотные последовательности этих двух пептидов были: GSS-A: EKYCDDDFHMAVFRTCAVS (SEQ ID NO: 1) (19 аминокислотных остатков, молекулярная масса 2236); GSS-B: SEYSGIASYCCLHGCTPSELSVVC (SEQ ID NO: 2) (24 аминокислотных остатка, молекулярная масса 2507). Из результатов масс-спектрометра и анализа секвенатора белка было установлено, что эти пептиды освобождены от любой химической модификации (внутриклеточная пост-трансляционная модификация белка и пептида) в боковых цепях и концевых аминокислотах после биосинтеза в нервных клетках морской звезды. Ген GSS был клонирован, исходя из аминокислотных последовательностей GSS-A и GSS-B. Были синтезированы основанные на аминокислотных последовательностях GSS-A и GSS-B: 5′-праймер DF1 (SEQ ID NO: 5) и 5′-праймер DF2 (SEQ ID NO: 6), соответственно; 3′-праймер DR1 (SEQ ID NO: 7) и 3′-праймер DR2 (SEQ ID NO: 8), соответственно. Геномная ДНК была выделена и очищена из семенника Asterina pectinifera при помощи QIAGEN(R) Genomic-tip. Часть генной последовательности GSS была амплифицирована методом гнездовой ПЦР при помощи образца генома в качестве матрицы и синтезированных вырожденных праймеров, и в результате нуклеотидная последовательность была расшифрована ДНК-секвенатором. На основе вновь полученной последовательности были синтезированы 5′-праймер GR (SEQ ID NO: 9) и 3′-праймер GF (SEQ ID NO: 10). 5′-прямые и 3′-обратные последовательности ДНК были расшифрованы при помощи праймеров, геномной ДНК и наборов CLONTECH Genome Walker (ТМ). На основе полученных последовательностей были синтезированы 5′-праймер MF1 (SEQ ID NO: 11) и 5′-праймер MF2 (SEQ ID NO: 12); 3′-праймер MR1 (SEQ ID NO: 13) и 3′-праймер MR2 (SEQ ID NO: 14). Суммарная РНК была выделена и очищена из нервов, амбулакральных ног, гепатопанкреаса, семенников и яичников Asterina pectinifera при помощи NIPPON GENE ISOGEN или QUIAGEN (R) QIAzol (TM). cDNA синтезировали с очищенной суммарной РНК при помощи QIAGEN (R) Omniscript (ТМ) RT. Последовательность cDNA GSS была амплифицирована при помощи cDNA в качестве матрицы и синтезированных праймеров (5′-праймера MF1, 5′-праймера MF2, 3′-праймера MR1, 3′-праймера MR2) методом гнездовой ПЦР, и последовательность cDNA GSS была расшифрована ДНК-секвенатором. Таким образом, был получен ген GSS, показанный в SEQ ID NO: 3. Нуклеотидная последовательность была переведена в аминокислотную последовательность SEQ ID NO: 4. Последовательности GSS-A и GSS-B, проанализированные масс-спектрометрией и секвенатором белка, расположены в районах аминокислот 30-48 (GSS-A) и 93-116 (GSS-B) аминокислотной последовательности SEQ ID NO: 4. Аминокислоты 1-29 из аминокислотной последовательности SEQ ID NO: 4 являются характерной сигнальной последовательностью секреторных белков, а аминокислоты 49-92 из аминокислотной последовательности представляют собой состоящую из 116 аминокислот последовательность (обозначенную как GSS-C), вырезаемую после биосинтеза аминокислотной последовательности. Последовательность KR, которая специфически и энзиматически удаляется после биосинтеза, расположена на обоих концах последовательности GSS-C. Пример 2 На основе результата Примера 1 были синтезированы две пептидные цепи (GSS-A и GSS-B) (чистота > 99,5%). Два синтезированных пептида, растворенных в 20 мМ Tris-буфере в концентрациях 0,4 мМ или 1 мМ (каждый в эквимолярном количестве), реагировали при комнатной температуре в течение 3 или 20 дней в присутствии окислителей с постоянным перемешиванием. 99,999%-ный газообразный кислород или 0,1 М окисленная форма глутатиона использовались как окислители. После реакции реактанты были разделены микродозовой высокоэффективной жидкостной хроматографией, при этом была оценена гонад-стимулирующая активность каждой пиковой фракции. Гонад-стимулирующая активность была обнаружена в относительно маленькой пиковой фракции с площадью пика 4,2%. Кроме того, молекулярная масса 4737 (GSS-A/B), которая указывает на сложную структуру и является такой же, как молекулярная масса природного гормона, была обнаружена только в пиковой фракции с гонад-стимулирующей активностью согласно анализу каждого пика массовой спектрометрией. Другие пики без гонад-стимулирующей активности показали молекулярную массу 2236 или 2507 (фиг.8). Была оценена гонад-стимулирующая активность полученных пептидов. Результаты показаны в таблице.
Синтезированный пептид (GSS-A или GSS-B) по отдельности не проявлял гонад-стимулирующей активности, и только комплекс (GSS-A/B) с молекулярной массой 4737, синтезированный в реакции окисления, показал активность. Синтезируемый в реакции окисления гормон (GSS-A/B) показал гормональную функцию не только в Asterina pectinifera, но также и в таких родственных видах, как Asterias amurensis. Настоящее изобретение позволяет наладить массовое производство гонад-стимулирующего гормона беспозвоночного и увеличить объем производства водных беспозвоночных, типа крабов, креветок, морских ежей, морских огурцов и моллюсков, а также разрабатывать новые ценные виды. Кроме того, искусственный контроль количества морской звезды в морской области водного хозяйства за счет снабжения оплодотворенными яйцами Asterina pectinifera и Asterias amurensis или их искусственного быстрого размножения может улучшить качество воды, ускоряя биологическое разложение корма, накопленного на морском дне. Так как эти две разновидности морской звезды – доминирующие разновидности в морской области водного хозяйства, и их среда обитания покрывает целую область от Хоккайдо до Кюсю, может ожидаться обширное применение.
Формула изобретения
1. Гонадотропный гормон беспозвоночного, включающий следующие два пептида, в которых SS-мостики сформированы между 6 цистеинами этих двух пептидов: (a) пептид, имеющий аминокислотную последовательность SEQ ID NO:1, (b) пептид, имеющий аминокислотную последовательность SEQ ID NO:2. 2. Гонадотропный гормон беспозвоночного, полученный при смешивании и окислении следующих двух пептидов: (a) пептид, имеющий аминокислотную последовательность SEQ ID NO:1, (b) пептид, имеющий аминокислотную последовательность SEQ ID NO:2. 3. Гонадотропный гормон беспозвоночного по п.1 или 2, где указанные два пептида получены при очистке (а) нервного органа, выделенного из беспозвоночного или (b) определенного органа, о котором известно, что он обладает гонад-стимулирующей активностью, основанной на гонад-стимулирующей активности беспозвоночного как индикаторе. 4. Способ получения гонадотропного гормона, включающий (1) культивирование и выращивание организма-хозяина, трансформированного вектором, содержащим две ДНК, каждая из которых кодирует следующие пептиды: (a) пептид, имеющий аминокислотную последовательность SEQ ID NO:1, (b) пептид, имеющий аминокислотную последовательность SEQ ID NO:2, (2) смешивание и окисление пептидов, полученных из организма-хозяина или из культуральной среды культуры организма-хозяина, и (3) выделение продукта.
РИСУНКИ
MM4A – Досрочное прекращение действия патента СССР или патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе
Дата прекращения действия патента: 26.08.2009
Извещение опубликовано: 27.07.2010 БИ: 21/2010
|
||||||||||||||||||||||||||
 -субъединицу и
-субъединицу и  -субъединицу.
-субъединицу. M пепстатина, 0,5 мг/л лейпептина, 0,2 мМ 4-(2-аминоэтил) фенилсульфонилфторида) в три приема и были гомогенизированы далее до тонкого порошка электрогомогенизатором (Physcotron). Экстракты из гомогенизированных тканей центрифугировались при 22,500xg при 4°C в течение 30 мин с последующим отделением супернатанта. Осадок был гомогенизирован с 200 мл 10 мМ раствора ацетата аммония и отцентрифугирован при 22,500xg при 4°C в течение 30 мин. Супернатант был отделен и смешан с предыдущим супернатантом. Полученный при 22,500xg супернатант ультрацентрифугировали при 100,000xg при 4°C в течение 1 часа, и отделяли супернатант. Супернатант имел гонад-стимулирующую активность.
M пепстатина, 0,5 мг/л лейпептина, 0,2 мМ 4-(2-аминоэтил) фенилсульфонилфторида) в три приема и были гомогенизированы далее до тонкого порошка электрогомогенизатором (Physcotron). Экстракты из гомогенизированных тканей центрифугировались при 22,500xg при 4°C в течение 30 мин с последующим отделением супернатанта. Осадок был гомогенизирован с 200 мл 10 мМ раствора ацетата аммония и отцентрифугирован при 22,500xg при 4°C в течение 30 мин. Супернатант был отделен и смешан с предыдущим супернатантом. Полученный при 22,500xg супернатант ультрацентрифугировали при 100,000xg при 4°C в течение 1 часа, и отделяли супернатант. Супернатант имел гонад-стимулирующую активность.