Патент на изобретение №2344176
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) АНАЛИЗЫ ГОМОЦИСТЕИНА И ЦИСТАТИОНИНА ФЕРМЕНТАТИВНЫМ ЦИКЛИЧЕСКИМ ПРЕОБРАЗОВАНИЕМ
(57) Реферат:
Изобретение относится к биотехнологии и представляет собой способ оценки количества гомоцистеина и/или цистатионина в растворе ферментативным циклическим преобразованием. Анализ включает стадии контактирования содержащего гомоцистеин и/или цистатионин раствора с CBS, L-серином и CBL и восстановительным средством в период времени, достаточный для катализа циклического преобразования формы гомоцистеина в цистатионина и обратного преобразования цистатионина в гомоцистеин с образованием пирувата и аммиака; определения количества присутствующих в реакционной смеси пирувата и/или аммиака и определения количества присутствующего в растворе гомоцистеина и/или цистатионина, основываясь на количестве образующегося пирувата и/или аммиака. При этом первая стадия контактирования длится менее 15 минут. Изобретение охватывает также и набор для проведения такого анализа. Изобретение позволяет сократить срок проведения анализа и, следовательно, повысить пропускную способность для образцов. 9 н. и 14 з.п. ф-лы, 16 ил., 34 табл.
СВЯЗАННЫЕ ЗАЯВКИ Данная заявка представляет собой частичное продолжение заявки на выдачу патента США с серийным № 09/704036, поданную 1 ноября 2000 года, заявляющую приоритет на основании предварительных заявок с серийным № 60/163126, поданную 2 ноября 1999 года, и серийным № 60/203349, поданную 10 мая 2000 года, а содержание и спецификации каждой из данных предшествующих заявок включены сюда в качестве ссылки. СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ Содержащий 21 последовательность в форме компьютерного ASCII-файла список последовательностей по настоящему изобретению включен сюда в качестве ссылки и прилагается к данному документу в виде двух (2) оригинальных компакт-дисков в соответствии с 37 CFR 1.821(c), их идентичных копий в соответствии с 37 CFR 1.821(c) и одной (1) их идентичной копии в соответствии с CFR 1.52(e). ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ Высокие уровни гомоцистеина в человеческой плазме коррелируют с высокими рисками коронарной болезни, инсульта, атеросклероза и других заболеваний. Как результат в общей популяции желательно провести скрининг на увеличенные количества данной аминокислоты. Чтобы сделать широкомасштабное тестирование гомоцистеина осуществимым, необходимо разработать новые и менее дорогостоящие способы анализа. Обычно плазменный гомоцистеин измеряют посредством высокоэффективной жидкостной хроматографии (ВЭЖХ) и газовой хроматографией/масс-спектрометрией (ГХ/МС) при стоимости более $100 за анализ, что делает данные способы физического разделения слишком дорогими для охватывающего всю популяцию исследования. Например, образцы мочи и крови можно подготовить для хроматографии аминокислот, а содержание L-гомоцистеина измерить разделением и детекцией посредством ВЭЖХ. Fiskerstrand et al. (Clin. Chem., 39:263-271 (1993)) описывают способ анализа L-гомоцистеина с применением флуоресцентного мечения сывороточных тиолов с последующими разделением и детекцией производного L-гомоцистеина среди множества других содержащих серу соединений посредством ВЭЖХ. Однако такие способы обычно требуют времени, дороги и не так легко доступны для многих лабораторий. Также разработаны способы непрямого иммунологического анализа гомоцистеина, однако данные способы, основанные на применении антител, все еще являются относительно дорогостоящими при $24 за тест. В одном конкретном непрямом иммунологическом анализе гомоцистеин преобразуется в аденозилгомоцистеин, а количество аденозилгомоцистеина определяют посредством конкурентного ELISA (твердофазный иммуноферментный анализ) с применением антитела против аденозилгомоцистеина (например, см. патент США 5827645, содержание которого включено сюда в качестве ссылки). Для количественного анализа L-гомоцистеина разработаны способы непрямого ферментативного анализа. Например, в тестовый образец добавляют фермент S-аденозилгомоцистеингидролазу и аденозин. Получающуюся в результате концентрацию или изменение концентрации аденозина в реакционной смеси применяют как индикатор исходной концентрации гомоцистеина в образце. Также описаны способы прямого ферментативного анализа для измерения гомоцистеина. Обычно в данных протоколах необратимо преобразуют гомоцистеин в другие соединения, поддающиеся количественному определению. Например, применяют фермент гомоцистеиндегидратазу для удаления сульфгидрильной группы из гомоцистеина. Затем измеряют концентрацию удаленной сульфгидрильной группы. Главным препятствием данному и другим способам ферментативного анализа гомоцистеина служит то, что применяемые ферменты взаимодействуют с другими содержащими серу аминокислотами и родственными гомоцистеину соединениями, приводя к высокому и неустойчивому фону и неточным измерениям гомоцистеина в плазме. Способы анализа ферментативным (или ферментным) циклическим преобразованием описаны для очень малого числа анализируемых соединений. При анализе ферментативным циклическим преобразованием применяют две или более ферментативных активности, регенерирующие субстрат и обратимо преобразующие измеряемое соединение. Вместо этого “соединение” применяют каталитически для контроля скорости преобразования в количественно анализируемое при данном анализе соединение. Как результат интересующее анализируемое соединение остается в постоянной концентрации, являющейся достаточно низкой, чтобы создать скорость реакции псевдопервого порядка. Таким образом, постоянная концентрация анализируемого соединения линейно связана со скоростью общего анализа. Посредством измерения скоростей реакции легко определить количество анализируемого соединения. Способы анализа ферментативным циклическим преобразованием иногда называют “амплифицирующими” анализами, так как способы обычно увеличивают чувствительность измерения для анализируемого соединения в 100-1000 раз. Амплификация представляет собой прямой результат неуменьшающейся постоянной концентрации соединения. Не сообщалось ни об одном анализе ферментативным циклическим преобразованием для измерения гомоцистеина. Настоящее изобретение относится к анализу гомоцистеина и/или цистатионина ферментативным циклическим преобразованием, являющемуся менее дорогостоящим и предоставляющему более высокую пропускную способность для образцов, чем доступные в настоящее время способы диагностического анализа. Кроме того, изобретение относится к способам и векторам для рекомбинантной продукции ферментов, которые можно применять для получения реагентов для анализа и тестовых наборов для определения количества гомоцистеина и цистатионина в образце. СУЩНОСТЬ ИЗОБРЕТЕНИЯ Настоящее изобретение относится к способу анализа ферментативным циклическим преобразованием для определения количества гомоцистеина и/или цистатионина в растворе. Анализ использует преимущество реакции гомоцистеина и L-серина с формированием цистатионина при посредстве фермента цистатионин- В одном из осуществлений настоящее изобретение относится к способу оценки количества гомоцистеина и/или цистатионина в растворе, включающему стадии: (a) контакта раствора, содержащего гомоцистеин и/или цистатионин (до и/или после проведения стадии восстановления дисульфидных групп), с CBS или ее производным, L-серином и CBL или ее производным для формирования реакционной смеси на период времени, достаточный для катализа циклического преобразования гомоцистеина в цистатионин и обратного преобразования цистатионина в гомоцистеин с образованием пирувата и аммиака; (b) определения количества пирувата и/или аммиака, присутствующего в реакционной смеси, и (c) оценки количества гомоцистеина и/или цистатионина, присутствующего в растворе, основываясь на количестве образовавшегося пирувата и/или аммиака. Более конкретно способ предусматривает оценку количества гомоцистеина посредством добавления недорогой аминокислоты L-серина. Количество присутствующего в реакционной смеси пирувата можно измерить множеством способов. В одном конкретном осуществлении настоящего изобретения в реакционной смеси присутствуют лактатдегидрогеназа (LDH) или ее производное и NADH (восстановленный кофактор никотинамид) или его производное. В присутствии NADH LDH преобразует пируват в лактат с окислением NADH в NAD+ (окисленный кофактор никотинамид). Окисление NADH до NAD+ можно измерить множеством известных в данной области способов, включающих в себя наблюдение за реакционной смесью при 340 нм. Образование NAD+ также можно контролировать посредством окисления красителя с получением видимого изменения окраски. Предпочитаемые для применения по настоящему изобретению красители включают в себя в качестве неограничивающих примеров 5,5′-дитиобис(2-нитробензойную кислоту), 2,6-дихлорфенилиндофенол, соединения тетразола, феназинметосульфат, метилвиологен или производные любого из данных красителей. Количество присутствующего в растворе гомоцистеина и/или цистатионина определяют по интенсивности наблюдаемой окраски, сравниваемой со стандартной кривой, построенной для образцов с известной концентрацией анализируемого соединения. В альтернативном осуществлении для определения количества присутствующего в образце пирувата применяют пируватоксидазу совместно с пероксидазой хрена в присутствии пероксида водорода и хромогена. Для данной колориметрической реакции предпочтительными хромогенами являются N-этил-N-(2-гидрокси-3-сульфопропил)-m-толуидин натрия (TOOS) и другие производные N-этил-N-(2-гидрокси-3-сульфопропил)аланина. Как и выше, количество присутствующего в растворе гомоцистеина и/или цистатионина определяют по интенсивности наблюдаемой окраски, сравниваемой со стандартной кривой, построенной для образцов с известной концентрацией анализируемого соединения. Количество присутствующего в растворе гомоцистеина и/или цистатионина также можно измерить, основываясь на количестве присутствующего в реакционной смеси аммиака. Существует множество способов определения концентрации аммиака в растворе. В одном конкретном осуществлении количество аммиака измеряют с применением доступного в большинстве случаев стандартного датчика аммиака. В другом осуществлении настоящее изобретение относится к способу оценки количества гомоцистеина и/или цистатионина в растворе, включающему стадии контакта раствора с восстанавливающим средством на период времени, достаточный для восстановления в основном всех молекул гомоцистеина и других дисульфидов, присутствующих в растворе с гомоцистеином. Воздействие восстанавливающим средством также может действовать для высвобождения гомоцистеина, присоединенного посредством дисульфидной связи к белку и/или другим присутствующим в растворе молекулам. После восстановления раствор затем приводят в контакт с CBS или ее производным, L-серином и CBL или ее производным на период времени, достаточный для катализа циклического преобразования гомоцистеина в цистатионин и преобразования цистатионина в гомоцистеин с образованием пирувата и аммиака. Для оценки количества присутствующего в растворе гомоцистеина и/или цистатионина можно определить количество присутствующего в растворе пирувата и/или аммиака, как изложено выше. Предпочтительные восстанавливающие средства для применения по настоящему изобретению включают в себя восстанавливающие средства на основе борогидридных солей и тиолов. Типичные восстанавливающие средства на основе тиолов, подходящие для применения по настоящему осуществлению, включают в себя дитиоэритритол (DTE), дитиотреитол (DTT), В еще одном осуществлении для оценки количества присутствующего в растворе гомоцистеина и/или цистатионина раствор можно предварительно обрабатывать цистатионин- В еще одном осуществлении настоящее изобретение относится к способу оценки количества присутствующего в растворе гомоцистеина и/или цистатионина, включающему в себя реакцию раствора с L-серином, и CBS или ее производным, и CBL или ее производным, иммобилизованными на твердой поверхности. Твердая поверхность может представлять собой, например, бумагу, фильтровальную бумагу, нейлон, стекло, керамику, диоксид кремния, оксид алюминия, диатомовую землю, целлюлозу, полиметакрилат, полипропилен, полистирол, полиэтилен, поливинилхлорид и их производные. Твердая поверхность может представлять собой стороны и дно контейнера для тестирования или может представлять собой вещество, добавленное в контейнер для тестирования. В предпочтительном осуществлении твердая поверхность включает в себя гранулу, добавляемую в контейнер для тестирования. CBS или ее производное, CBL или ее производное и цистатионин- Настоящее изобретение в качестве способа оценки количества гомоцистеина в образце относится к формату анализа, который коррелирует с количеством комплекса гомоцистеин/транскрипционный фактор, связывающийся с консенсусной связывающей полинуклеотиды последовательностью. В конкретном осуществлении способ включает в себя контакт образца с восстанавливающим средством на период времени, достаточный для высвобождения гомоцистеина из любого ассоциированного белка; контакт восстановленного гомоцистеина с транскрипционным фактором, связывающим метаболит гомоцистеина, в условиях, подходящих для образования комплекса; смешивание образца с консенсусной полинуклеотидной последовательностью, специфически распознающейся комплексом гомоцистеин/транскрипционный фактор, и оценку количества присутствующего в образце гомоцистеина по количеству комплекса гомоцистеин/транскрипционный фактор, связанного с консенсусной полинуклеотидной последовательностью. Пригодные для применения в данном способе восстанавливающие средства содержат восстанавливающие средства на основе борогидридных солей и тиолов, включающие в себя дитиоэритритол (DTE), дитиотреитол (DTT), Еще одно осуществление настоящего изобретения относится к набору для тестирования, содержащему контейнер для хранения раствора, в котором оценивают количество гомоцистеина и/или цистатионина, L-серин, CBS или ее производные, CBL или ее производные и любые буферы, дополнительные вещества и растворители, необходимые для создания условий, способствующих высокой ферментативной активности. Набор для тестирования может дополнительно содержать лактатдегидрогеназу или ее производное и NADH или его производное. NADH можно измерять непосредственно при 340 нм или может содержаться краситель, способный изменять окраску при окислении. Количество гомоцистеина и/или цистатионина коррелирует с изменением оптической плотности, измеряемой в течение времени. В предпочтительном осуществлении ферменты предоставляют иммобилизованные на твердой основе. Твердая основа может включать поверхность контейнера, предоставленного для хранения образца для тестирования, или может представлять собой гранулу или другой предмет, добавляемый в контейнер. В дополнительном осуществлении настоящего изобретения для удаления любой формы цистатионина из раствора для тестирования до ферментативного циклического анализа как часть набора для тестирования может предоставлять цистатионин- Обнаружено, что предпочтительный анализ по настоящему изобретению можно проводить за относительно короткий промежуток времени и с относительно небольшими количествами фермента, предлагая анализ с существенными коммерческими преимуществами. Например, предпочтительный анализ включает в себя создание реакционной смеси, включающей в себя содержащий гомоцистеин образец, серин, CBS, CBL, лактатдегидрогеназу, NADH и восстановитель, такой как DTE или DTT, с соотношением CBS/CBL в смеси приблизительно от 1:1 до 25:1, более предпочтительно – приблизительно от 1:10, и еще более предпочтительно – приблизительно от 2:1 до 5:1. Было полезным обнаружение того, что восстановитель может находиться вместе с системой детекции (то есть лактатдегидрогеназа и NADH) без неблагоприятного влияния на анализ. Следовательно, для того чтобы уменьшить продолжительность анализа, анализ по настоящему изобретению можно проводить без отдельной стадии восстановления. Таким образом, когда необходимо оценить множество образцов, посредством последовательного создания в контейнере (например, кювета для спектрофотометрического анализа) реакционной смеси, состоящей из образца, серина, CBS, CBL, лактатдегидрогеназы, NADH и восстановителя, и оценки количества присутствующего в реакционной смеси пирувата посредством наблюдения за образованием NAD+ в течение времени, временной интервал между соответствующими стадиями реакции-создания может являться таким коротким, как 10-50 секунд, с общим временем реакции приблизительно не более 20 минут для данного образца. Более предпочтительно, чтобы общее время реакции для данного образца составляло не более 15 минут, а еще более предпочтительно – приблизительно не более 13 минут. Общее количество образцов, анализируемых за час, может составлять приблизительно 15-30 для более медленных устройств, но таким большим как 200-400 для более быстрых устройств. При данном предпочтительном анализе, подготавливают первую реакционную смесь, включающую в себя образец, серин, лактатдегидрогеназу, NADH и восстановитель, с подходящим периодом инкубации, чтобы обеспечить высвобождение преобладания (а предпочтительно – по существу всего) гомоцистеина (общий гомоцистеин), присутствующего в образце в связанном, окисленном и/или свободном состояниях. Далее, для составления полной реакционной смеси и инициации ферментативного циклического преобразования гомоцистеина и/или цистатионина добавляют CBS и CBL. В другом предпочтительном осуществлении цистатионин можно применять для создания калибратора(ов) для ферментативного анализа (так как анализ ферментативным циклическим преобразованием, преобразующим гомоцистеин и цистатионин друг в друга). Другими словами, изменяющиеся известные уровни цистатионина можно применять в системе анализа для “калибровки” или “стандартизации” анализа и/или устройства, позволяющего осуществить количественный анализ результатов образца. В данном осуществлении в биологический образец добавляют известную концентрацию цистатионина и затем подвергают анализу с применением одного из других осуществлений. Результаты, как правило, применяют для построения калибровочной прямой, которую затем применяют для установления прямой гомоцистеина ввиду высокой степени корреляции между данными двумя прямыми. Альтернативно известные уровни цистатионина можно применять в качестве меры контроля качества, чтобы убедиться, что анализ работает должным образом. В данном осуществлении контроля качества данные “известные” уровни цистатионина оценивают так, как будто они являлись неизвестными образцами, и результаты сравнивают с их известными (ожидаемыми) значениями, для того чтобы убедиться, что система анализа функционирует должным образом. Предпочтительные способы анализа проводят с применением выделенных (очищенных) ферментов CBL и CBS, по меньшей мере, приблизительно с 80% (предпочтительно – по меньшей мере, приблизительно 90%) идентичности последовательности с ферментами, выбираемыми из группы, состоящей из SEQ ID Nos. 19 и 20. Как применяют здесь, “идентичность последовательности”, как это известно в данной области, относится к родству между двумя или более белковыми или полипептидными последовательностями или двумя или более полинуклеотидными последовательностями, то есть эталонной последовательностью и данной последовательностью, сравниваемой с эталонной последовательностью. Идентичность последовательности определяют сравнением данной последовательности с эталонной последовательностью после оптимального сопоставления последовательностей для получения наибольшей степени подобия последовательности, определяемой посредством совпадения строк таких последовательностей. При таком сопоставлении идентичность последовательности устанавливают на основании принципа «позиция к позиции», например, последовательности “идентичны” в определенной позиции, если в данной позиции нуклеотиды или аминокислотные остатки идентичны. Общее число таких идентичных позиций затем делят на общее число нуклеотидов или остатков в эталонной последовательности для получения % идентичности последовательности. Идентичность последовательностей легко подсчитать посредством известных способов, включающих в качестве неограничивающих примеров примеры, описанные в Computational Molecular Biology, Lesk, A. N., ed., Oxford University Press, New York (1988), Biocomputing: Informatics and Genome Projects, Smith, D. W., ed., Academic Press, New York (1993); Computer Analysis of Sequence Data, Part I, Griffin, A. M., and Griffin, H. G., eds., Humana Press, New Jersey (1994); Sequence Analysis in Molecular Biology, von Heinge, G., Academic Press (1987); Sequence Analysis Primer, Gribskov, M. and Devereux, J., eds., M. Stockton Press, New York (1991) и Carillo, H., and Lipman, D., SIAM J. Applied Math., 48: 1073 (1988), руководства которых включены сюда в качестве ссылки. Предпочтительные способы определения идентичности последовательности предназначены для получения максимального совпадения между тестируемыми последовательностями. Способы определения идентичности последовательности закодированы в присутствующих в свободном доступе компьютерных программах, определяющих идентичность последовательности у данных последовательностей. Такие программы включают в качестве неограничивающих примеров программный пакет GCG (Devereux, J., et al., Nucleic Acids Research, 12(1):387 (1984)), BLASTP, BLASTN и FASTA (Altschul, S. F. et al., J. Molec. Biol., 215: 403-410 (1990). Программа BLASTX свободно распространяется из NCBI и других источников (BLAST Manual, Altschul, S. et al., NCVI NLM NIH Bethesda, MD 20894, Altschul, S. F. et al., J. Molec. Biol., 215:403-410 (1990), руководства которых включены сюда в качестве ссылки). Данные программы с применением принятых по умолчанию весовых коэффициентов для промежутков оптимально сопоставляют последовательности, чтобы получить наивысший уровень идентичности последовательности между данной и эталонной последовательностями. В качестве иллюстрации полинуклеотид с нуклеотидной последовательностью, например, по меньшей мере, с 95% “идентичности последовательности” с эталонной нуклеотидной последовательностью, означает, что нуклеотидная последовательность данного полинуклеотида идентична эталонной последовательности за исключением того, что данная полинуклеотидная последовательность может включать в себя до 5 точечных мутаций на каждые 100 нуклеотидов эталонной нуклеотидной последовательности. Другими словами, в полинуклеотиде с нуклеотидной последовательностью, по меньшей мере, с 95% идентичности относительно эталонной нуклеотидной последовательности до 5% нуклеотидов в эталонной последовательности могут являться удаленными или замещенными другим нуклеотидом или в эталонную последовательность могут быть встроены нуклеотиды в количестве до 5% от общего числа нуклеотидов в эталонной последовательности. Данные мутации эталонной последовательности могут происходить на 5′- или 3′-концевых участках эталонной нуклеотидной последовательности или где-либо между данными концевыми участками, расположенные случайным образом или индивидуально, между нуклеотидами в эталонной последовательности или одной или более непрерывными группами в эталонной последовательности. Аналогично полипептид с заданной аминокислотной последовательностью, например, по меньшей мере, с 95% идентичности последовательности с эталонной аминокислотной последовательностью означает, что заданная аминокислотная последовательность полипептида идентична эталонной последовательности за исключением того, что данная полипептидная последовательность может включать в себя до 5 изменений аминокислот на каждые 100 аминокислот в эталонной аминокислотной последовательности. Другими словами, для получения данной полипептидной последовательности, по меньшей мере, с 95% идентичностью последовательности с эталонной аминокислотной последовательностью до 5% аминокислотных остатков в эталонной последовательности могут являться удаленными или замещенными другими аминокислотами или в эталонную последовательность могут быть встроены аминокислоты в количестве до 5% от общего числа аминокислот в эталонной последовательности. Данные изменения эталонной последовательности могут происходить на амино- или карбоксиконцевых участках эталонной аминокислотной последовательности или где-либо между данными концевыми участками, расположенные случайным образом или индивидуально, между остатками в эталонной последовательности, или одной или более непрерывными группами в эталонной последовательности. Предпочтительно положения остатков, которые не идентичны, отличаются консервативными заменами аминокислот. Однако консервативные замены не включаются как совпадение, когда определяют идентичность последовательности. Пригодные для настоящего изобретения реагенты включают в себя CBS, CBL, L-серин, LDH, NADH, DTE В некоторых предпочтительных осуществлениях буфер реагента 1 представляет собой 250 мМ Tris при pH 8,4. Однако когда Tris применяют как буфер, pH может находиться в диапазоне от 7,0 до 9,0. Более предпочтительно, чтобы pH находился в диапазоне от 7,5 до 8,5 и даже более предпочтительно – приблизительно от 8,1 до 8,5. Применение увеличенной концентрации Tris улучшает стойкость анализа посредством помощи в поддержании более постоянного pH, т.к. он является более концентрированным буфером. Дополнительно, CBS и CBL являются существенно более активными в данном предпочтительном диапазоне pH, а особенно – при pH приблизительно от 8,1 до 8,5. Настоящее изобретение также относится к анализу, одинаково хорошо работающему с мутными образцами. Проблемы с мутностью уменьшают посредством добавления к R1 липазы и В другом изменении настоящего изобретения испытывали действия изменяющихся объемов реагентов. Такие изменения не оказывали существенного влияния на точность анализа. В настоящем изобретении также испытывали действия различных детергентов и их концентраций. Например, Genapol X-80 может присутствовать в реагенте 1 в концентрации приблизительно от 0,05 до 0,5%. Более предпочтительно, чтобы концентрация составляла от 0,1 до 0,5% и даже более предпочтительно – приблизительно от 0,2 до 0,4%. Из них испытывали три концентрации (0,1%, 0,3% и 0,5%) с концентрацией в размере 0,3%, обеспечивающей наиболее точные результаты. Также в реагенте 1 изменяли концентрацию Brij-35. Brij-35 можно применять в R1 по настоящему изобретению в концентрации приблизительно от 0,01 до 0,5%. Более предпочтительно, чтобы данная концентрация находилась в диапазоне приблизительно от 0,015 до 0,1% и даже более предпочтительно – приблизительно от 0,020 до 0,030%. Из испытываемых концентраций (0,025%, 0,05%, 0,1% и 0,5%) наиболее точные и непротиворечивые результаты получены с применением концентрации в размере 0,025%. В настоящем изобретении также испытывали замену восстанавливающего средства DTE на трис-(2-карбоксиэтилфосфин)гидрохлорид (TCEP), который стабилен в очищенной воде протяженный период времени. Такая замена уменьшала реакцию чистого реагента приблизительно от 12-15 мА/мин до приблизительно 4-5 мА/мин, что делало возможным большую степень точности для данного анализа. Когда по настоящему изобретению применяют TCEP, его концентрация может находиться в диапазоне приблизительно от 6 до 53 мМ. Более предпочтительно, чтобы данная концентрация находилась в диапазоне приблизительно от 10 до 45 мМ и даже более предпочтительно – приблизительно от 20 до 30 мМ. Из испытываемых в данной заявке концентраций концентрация 26,44 мМ обеспечивала наиболее точные результаты анализа. Состав, содержащей реагент 3 смеси, также изменяли. Например, буфер Tris заменяли фосфатным буфером, а к буферу добавляли глицерин. Фосфат являлся эффективным в концентрациях, изменяющихся приблизительно от 50 мМ до приблизительно 500 мМ, при pH, равном 7,6. Концентрации глицерина находились в диапазоне приблизительно от 0,5% до 15,0% (об./об.). Фосфат и глицерин применяли для обеспечения более длительного срока хранения ферментов, что невозможно при применении буфера Tris. Одно из преимуществ настоящего изобретения состоит в том, что его можно применять в любом из ряда устройств. Например, анализ можно адаптировать для тестирования в двух различных устройствах: Hitachi 911 и Beckman CX5. Примечательно, что точные и непротиворечивые результаты получены на любом из данных двух устройств. Кроме того, анализ можно проводить с применением только 2 реагентов посредством смешивания компонентов R1 и R2 вместе в правильном соотношении, основанном на технических требованиях устройств. В конечном счете настоящее изобретение можно адаптировать для применения с одной точкой калибровки. Существенно, что результаты калибровки с одной точкой являлись такими же точными, как и с кривой калибровки из нескольких точек. Краткое описание чертежей На фигуре 1 изображены графики зависимости поглощения от времени, полученные с применением водных растворов пирувата (5-100 мкМ), демонстрирующие количественный анализ пирувата. Компоненты реагента являлись такими, как описано в таблице 1, а настройки устройства являлись такими, как описано в таблице 2. На фигуре 2 изображена стандартная кривая из значений поглощения, нанесенных в зависимости от известных количеств пирувата (0-100 мкМ). Время анализа составляло 5 минут. На фигуре 3 изображены графики зависимости поглощения от времени, полученные с применением водных растворов цистатионина (0-100 мкМ). Цистатионин измеряли ферментативным преобразованием цистатионина в гомосерин, пируват и аммиак посредством CBL. На фигуре 4 изображены стандартные кривые анализа цистатионина, полученные с применением водных растворов цистатионина (от 0 до 100 мкМ). Время реакции составляло или 5, или 20 минут. На фигуре 5 изображены графики зависимости поглощения от времени, демонстрирующие систему циклического преобразования и передачи сигнала CBS/CBL/пируватоксидаза/пероксидаза. Цистатионин растворяли в воде для получения стандартов при концентрациях от 1 до 100 мкМ. На фигуре 6 изображены графики зависимости поглощения от времени, демонстрирующие систему циклического преобразования и передачи сигнала CBS/CBL/пируватоксидаза/пероксидаза. Гомоцистеин растворяли в воде для получения стандартов при концентрациях от 0 до 100 мкМ. На фигуре 7 изображены типичная кривая калибровки, полученная с применением водных растворов гомоцистеина (от 0 до 100 мкМ) с применением системы циклического преобразования и детекции CBS/CBL/PO/HRP. На фигуре 8 изображены типичные графики зависимости поглощения от времени, полученные с применением гомоцистеина (от 0 до 100 мкМ) в водном растворе с применением системы CBS/CBL/LDH без дитиотреитола. На фигуре 9 изображены типичные графики зависимости поглощения от времени, полученные с применением гомоцистеина (от 0 до 100 мкМ) в водном растворе с применением системы циклического преобразования CBS/CBL/LDH с дитиотреитолом. На фигуре 10 изображен график калибровки концентрации гомоцистеина (от 0 до 100 мкМ) в зависимости от поглощения с дитиотреитолом и без. Время реакции составляло 5 минут. На фигуре 11 изображена зависимость между результатами количественного анализа, полученная с применением системы ферментативного циклического преобразования CBS/CBL/LDH для измерения гомоцистеина в образцах человеческой плазмы с применением DTT в качестве восстанавливающего средства в сравнении с результатами количественного анализа, полученными посредством способа измерения гомоцистеина Abbott IMx. На фигуре 12 изображен график зависимости, иллюстрирующий зависимость между анализом по настоящему изобретению (y) и коммерчески доступным анализом IMx (x), как описано в примере 7. На фигуре 13 изображен график зависимости, иллюстрирующий зависимость между анализом по настоящему изобретению (y) и традиционным анализом ВЭЖХ (x), как описано в примере 7. На фигуре 14 изображен график зависимости, иллюстрирующий зависимость между анализом IMx (y) и анализом ВЭЖХ (x), как описано в примере 7. На фигуре 15 изображен график зависимости поглощения от времени для анализа с пируватом без DTT, описанного в примере 9 и На фигуре 16 изображен график зависимости поглощения от времени для содержащего DTT анализа с пируватом, описанного в примере 9. ОПИСАНИЕ КОНКРЕТНЫХ ОСУЩЕСТВЛЕНИЙ Настоящее изобретение относится к способу гомогенного анализа ферментативным циклическим преобразованием для оценки количества гомоцистеина в образце. Ферментативное циклическое преобразование поддерживает постоянный уровень гомоцистеина и дополнительно обеспечивает увеличение чувствительности оценки гомоцистеина. Дополнительно настоящее изобретение относится к генам и экспрессирующим векторам для рекомбинантного получения ферментов, применимых по настоящему изобретению. Также предоставлены наборы для тестирования для оценки количества гомоцистеина и/или цистатионина в растворе. Гомоцистеин присутствует в клетках как промежуточная аминокислота, синтезируемая тогда, когда метионин метаболизируется в цистеин. Как правило, гомоцистеин, синтезируемый в организме, быстро метаболизируется по одному из двух путей: (1) конденсация с серином с формированием цистатионина или (2) превращение в метионин, и его концентрация (а также концентрация его окисленной формы, гомоцистина) в организме в нормальных условиях низка. Средний плазматический уровень гомоцистеина у здорового взрослого составляет приблизительно 12 мкМ у мужчин, колеблясь приблизительно от 8,6 до приблизительно 17,1 мкМ, и 10 мкМ у женщин, колеблясь приблизительно от 3,9 до 16,8 мкМ (Vester et al., Eur. J. Clin. Chem. Biochem., 29:549-554 (1991); Jacobsen et al. Clin. Chem., 40:873-881 (1994)). Недавно определили, что для ряда состояний величина уровней гомоцистеина в биологическом образце представляет клиническую значимость. Увеличенные количества гомоцистеина в крови коррелируют с развитием атеросклероза (Clarke et al., New Eng. J. Med., 324:1149-1155 (1991)). В настоящее время умеренная гомоцистеинемия также рассматривается как фактор риска для сердечно-сосудистого заболевания. Точность обнаружения у нормальных взрослых нормального диапазона гомоцистеина очень важна, так как определено, что риск заболевания сердца заметно увеличивается всего при 30% превышении нормальной концентрации, увеличивая фактор риска в 3,4 раза (Stampfer et al., JAMA 268:877-881 (1992)). Как здесь подразумевается, термин “оценка” охватывает и количественное и качественное определение количества гомоцистеина и/или цистатионина, присутствующего в тестовом образце. Как применяется здесь, “раствор” или “тестовый раствор” относится к образцу клинической или биологической жидкости или тканевому экстракту и может включать в себя, например, кровь, производные крови, такие как плазма, или мочу и т.п. Содержащий гомоцистеин образец в присутствии фермента CBS подвергают взаимодействию с L-серином с образованием цистатионина. Цистатионин затем можно преобразовать в гомоцистеин, пируват и аммиак вторым ферментом, CBL. Результат применения данных анаболического и катаболического ферментов, действующих совместно, представляет собой “бесполезный цикл”, в котором (1) цистатионин и гомоцистеин постоянно превращаются друг в друга, (2) разрушается серин и (3) синтезируются пируват и аммиак. Применение относительно высоких концентраций серина по сравнению с гомоцистеином обеспечивает условия, благоприятствующие общей скорости реакции, соответствующей кинетике псевдопервого порядка, непосредственно зависящей от количества гомоцистеина и цистатионина в реакционной смеси. Если концентрация цистатионина является очень низкой по сравнению с концентрацией гомоцистеина, скорость всей реакции изменяется линейно в зависимости от количества гомоцистеина. Посредством измерения снижения серина или увеличения пирувата или аммиака можно определить количество гомоцистеина в неизвестном растворе или образце посредством сравнения контрольных реакций, содержащих известные концентрации гомоцистеина, подвергающихся идентичным ферментативным преобразованиям. Способ ферментативных циклических преобразований по настоящему изобретению обеспечивает процесс амплификации, образующий значительно большие количества конечных продуктов, чем количества гомоцистеина, которые, вероятно, существуют в тестовом растворе, т.е. крови. Следовательно, способы анализа для определения количества пирувата или аммиака, образующегося в реакции, не должны являться чрезвычайно чувствительными и измерения можно провести существующими способами, известными специалистам в данной области. Пируват обычно измеряют посредством ферментативного преобразования в лактат лактатдегидрогеназой или ее производным. Для данной реакции ферментативного преобразования необходим кофактор NADH (восстановленный кофактор никотинамид) или его производное, который преобразуется в NAD+ (окисленный кофактор никотинамид). Реакцию можно контролировать посредством измерения поглощения при 340 нм. Как применяют здесь, термин “производные” по отношению к кофакторам относится к солям, сольватам и другим химически модифицированным формам, сохраняющим активность кофактора, но могут обладать увеличенной стабильностью в водных растворах. Производные могут включать в себя в качестве неограничивающих примеров ацетильные производные NADH, включающие в себя 3-пиридинальдегид-NADH, 3-ацетилпиридин-NADH, 3-тионикотинамид-NADH и т.п. (например, см. патент США 5801006). Другие удобные способы анализа для определения окисления NADH или его производных включают в себя, например, измерение флуоресценции, как описано у Passoneau et al., в Enzymatic Analysis, A Practical Guide, страницы 219-222 (1993), (включенного здесь в качестве ссылки). В другом осуществлении NADH или его производные подвергают взаимодействию с пируватом в присутствии лактатдегидрогеназы с образованием NAD+, который далее подвергают взаимодействию с красителем, способным при окислении изменять окраску, предпочтительно в видимом диапазоне. Изменение окраски красителя можно применять для определения общей степени окисления NADH в NAD+, что соответствует количеству пирувата, образующемуся в реакции ферментативного циклического превращения. Примеры красителей, которые можно применять по настоящему изобретению, включают в себя в качестве неограничивающих примеров 5,5′-дитиобис(2-нитробензойную кислоту), 2,6-дихлорофенолиндофенол, соединения тетразола, феназинметосульфат, метилвиологен или производные каждого из них. Также количество пирувата можно измерить в реакции с пируватоксидазой или ее производными в присутствии пероксидазы, т.е. пероксидазы хрена и т.п. Количество пирувата, преобразованного в пероксид водорода, определяют окислительной конденсацией, например, водорастворимого донора водорода, образующего цветное соединение для фотометрического определения концентрации пероксида. Особенно предпочтительные доноры водорода, образующие стабильный продукт цветной реакции, включают в себя водорастворимые производные N-алкил-N-сульфопропилаланиновых производных, т.е. натриевую соль N-этил-N-(2-гидрокси-3-сульфопропил)-m-толуидина (TOOS) (Tamaoku et al., Chem. Pharm. Bull. 30:2492-2497 (1982)). Количество H2О2 также можно измерить электрометрически. Это включает в себя амперометрические и потенциометрические измерения. Определение количества гомоцистеина и/или цистатионина проводят исходя из корреляции с количеством пероксида, образуемого за определенный период времени при преобразовании пирувата пируватоксидазой. Также посредством различных способов, включающих применения датчика аммиака (Willems et al., Clin. Chem. 34:2372 (1988)), можно определить содержание аммиака. Аммиак также можно легко обнаружить, применяя известные способы колориметрии. Например, образующийся в образце аммиак может взаимодействовать с формированием окрашенного продукта, формирование которого можно детектировать спектрофотометрически. Один такой способ, описанный в Methods of Enzymatic Analysis (Bergmeyer) 1:1049-1056 (1970), включенный сюда в качестве ссылки, основывается на реакции аммиака с фенолом в присутствии гипохлорита в щелочных условиях с формированием окрашенного красителя индофенола. В качестве катализатора можно применять нитропруссид натрия. Также можно применять модификации данного способа, где, например, применяют различные производные. Окрашенный конечный продукт образуется в количествах, прямо пропорциональных концентрации аммиака и, следовательно, гомоцистеина. Другие способы могут включать в себя, например, анализ микродиффузии (Seligson et al., J. Lab. Clin. Med., 49:962-974 (1957)), анионообменную технологию (Forman, Clin. Chem., 10:497-508 (1964)) и различные ферментативные способы (например, см. Mondzac et al., J. Lab. Clin. Med., 66:526-531 (1979)). Способ ферментативного циклического преобразования для оценки количества гомоцистеина и/или цистатионина уменьшает трудности, встречающиеся в других предшествующих способах анализа, где фоновые соединения и фоновые реакции образовывали “шум”. В анализе по настоящему изобретению для обеспечения полного преобразования L-серина в пируват и аммиак можно добавить большие количества недорогой аминокислоты, например L-серина. В предпочтительном осуществлении можно применять, а затем преобразовывать в пируват и аммиак концентрации серина в размере 250 мкМ или более. Обычно плазма содержит приблизительно 100 мкМ серина. Данное количество эндогенного серина несущественно для настоящего анализа, так как анализ измеряет скорость преобразования, которая ограничена количеством гомоцистеина, а не количеством серина. Считается, что концентрации L-серина приблизительно от 0,1 мкМ до 10 мМ не будут воздействовать на чувствительность или точность описанного анализа. Дополнительно реакция ферментативного циклического преобразования по настоящему изобретению обеспечивает скорость преобразования L-серина, которая зависит от концентрации гомоцистеина как в реакции псевдопервого порядка. Нормальные уровни содержания пирувата в крови обычно составляют приблизительно от 0,4 до 2,5 мг/100 мл (Lehninger, AL. Principles of Biochemistry. 1982. Worthington Publishers, Inc. p. 707, включенный сюда в качестве ссылки), что соответствует приблизительно от 45 до 284 мкМ. Данная концентрация пирувата может обеспечить незначительный размер фона, но настоящий анализ измеряет изменение в течение времени, и его можно легко настроить по начальному уровню пирувата в растворе. Кроме того, пируват можно удалить предварительной инкубацией с лактатдегидрогеназой или другими ферментами. Более важно, что цистеин и другие содержащие серу соединение, например, в крови, не вносят значительного вклада в реакцию ферментативного циклического преобразования и, следовательно, рассматриваются как незначительный шум в системе. В альтернативном осуществлении настоящего изобретения образец можно обработать для удаления цистатионина для увеличения точности измерения гомоцистеина. В качестве конкретного примера воздействие включает в себя контакт с раствором цистатионин- Данное конкретное осуществление изобретения также можно применять для оценки количества цистатионина посредством сравнения различий в скорости образования пирувата и/или аммиака в образце, обработанном цистатионин- В определенных биологических образцах, таких как кровь, гомоцистеин часто посредством дисульфидных связей связан с белками, тиолами и другими клеточными компонентами. Обычно образец можно обработать восстанавливающим средством, таким как борогидрид натрия или калия; тиолом, таким как дитиоэритритол, В типичном осуществлении образец, содержащий кровь, фракции крови, сыворотку, плазму или мочу, обрабатывают восстанавливающим средством и непосредственно анализируют на предмет содержания гомоцистеина. Также предпочтительным может являться нагревание образца до анализа для ускорения высвобождения гомоцистеина и для инактивации деградируемых ферментов и других факторов, которые могут мешать анализу. Специалистам в данной области известен общий способ обработки образцов крови для способов анализа гомоцистеина в других способах, таких как тесты ВЭЖХ. CBS и CBL представляют собой ферменты, обычно встречающиеся в природе. Люди и дрожжи используют CBS для синтеза метионина; а человеческую кДНК можно заменить геном CBS пивных дрожжей (Kruger et al., Hum. Molec. Genet., 4:1155-1161 (1995)). CBL обнаружена у многих бактерий, включая E. coli (Laber et al., FEBS Lett., 379:94-96 (1996)). Полные последовательности ДНК генов CBS и CBL в данных организмах и других видах известны, предоставляя квалифицированным молекулярным биологам источник для легкого клонирования или синтеза данных генов или их производных. Как используют здесь, “производное” по отношению к ферментам относится к мутантам, получаемым посредством добавления, удаления, замещения и/или модификации аминокислот; мутантам, получаемым посредством рекомбинации и/или перестановки ДНК и солям, сольватам и другим химически модифицированным формам ферментов, которые сохраняют ферментативную активность выделенного природного фермента. Способ создания производного фермента посредством перестановки ДНК, применимый в настоящем изобретении, описан в патенте США 5605793 (включенном сюда в качестве ссылки). Данные два фермента были очищены из ряда различных организмов даже без клонирования. См., например, Ono et al., Yeast 10:333-339 (1994); Dwivedi et al., Biochemistry 21:3064-3069 (1982), включенные сюда в качестве ссылки. В предпочтительном осуществлении настоящего изобретения для получения CBS или ее производного и для получения CBL или ее производного, которые можно применять в анализе ферментативным циклическим преобразованием, предоставлены генетически измененные организмы для получения реагентов с более низкой стоимостью и для упрощения очистки ферментов. Подобным образом термин “производное” по отношению к нуклеотидным последовательностям относится к вариантам и мутантам, получаемым посредством добавления, удаления, замещения и/или модификации нуклеотидов и/или посредством комбинаторной химии; мутантам, получаемым посредством рекомбинации и/или перестановки ДНК, и солям, сольватам и другим химически модифицированным формам последовательности, которые сохраняют желательную активность последовательности, такую как характеристики связывания MetR. Когда применяют модифицированные производные последовательности, последовательности предпочтительно обладают, по меньшей мере, приблизительно 60% идентичности последовательности с исходной последовательностью. Более предпочтительно, чтобы данная идентичность представляла собой, по меньшей мере, приблизительно 70% и более предпочтительно – по меньшей мере 85%. Наиболее предпочтительно, чтобы такие последовательности обладали по меньшей мере приблизительно 95% идентичности последовательности с исходной последовательностью. Секвенирован ген CBS и весь геном Saccharomyces cerevisiae. Также известны ген CBL и оставшиеся гены E. coli. Следовательно, с применением общих для области молекулярной биологии способов гены, кодирующие CBS и CBL, можно клонировать и экспрессировать в обоих организмах. Известны опубликованные способы очистки данных двух ферментов, в которых применяют стандартные способы очистки белков. Однако каждый из данных ферментов можно сконструировать в виде гибридного белка, включающего в себя, например, часть другого белка, содействующего солюбилизации и/или повторной укладке активного белка, или аминокислотные последовательности, которые можно добавлять для облегчения очистки. Пример аминокислотных последовательностей, добавляемых для очистки, включает в себя аффинные метки, такие как поли-His или глутатион-S-трансферазу (GST), которые можно добавлять, чтобы обеспечить более быструю очистку белков на одной аффинной колонке. Хотя для диагностического теста по настоящему изобретению ферменты не требуют очистки до гомогенности, посредством очистки можно уменьшить до незначительных уровней содержание протеаз и других активных соединений, преобразующих серин в пируват. Кроме того, концентрация ферментов, как правило, приводит к более высокой стабильности ферментативной активности во время реакции и к более длительному сроку хранения. Ожидается, что CBS и CBL из других организмов, отличных от S. cerevisiae и E. coli, особенно термофилов, являются источниками ферментов даже с большей стабильностью. Выделенные ферменты можно выбрать по причине более высокого сродства к гомоцистеину. Кроме того, белковая инженерия дрожжевой CBS и бактериальной CBL может привести к получению ферментов с улучшенными коммерческими свойствами, включающими в себя повышенные аффинности, более высокие скорости реакции, более легкую очистку и больший период полужизни при хранении. Подобным образом у многих видов подробно описана цистатионин- Рекомбинантная экспрессия генов CBS, CBL и CGL является типичной. Данные способы могут обеспечить большие количества очищенного белка, а гены можно модифицировать, например, для добавления к экспрессирующимся ферментам заданной пептидной последовательности (аффинной метки), которую можно применять для аффинной очистки. Посредством применения аффинных меток в экспрессии рекомбинантных ферментов, очистку и иммобилизацию данных ферментов можно значительно упростить, что ведет к меньшим стоимостям. Аффинные метки известны в данной области и включают в себя, в качестве неограничивающих примеров, полигистидин, GST (глутатион-S-трансферазу), стреп-метку, “flag”, последовательности c-myc и т.п. Метки широко применяют для аффинной очистки рекомбинантно экспрессированных белков, а многие доступны коммерчески. В предпочтительном осуществлении CBS и CBL иммобилизуют на твердой поверхности таким образом, чтобы сохранить активности ферментов, и таким образом, чтобы оставить гомоцистеин свободным для прямого взаимодействия с белками. Посредством иммобилизации ферментов при высокой плотности уменьшают дистанции диффузии между различными белками по сравнению с ферментами в растворе. Как результат общая скорость реакции анализа значительно увеличивается, так как на продукт одного фермента можно воздействовать другим ферментом и наоборот. Увеличение скоростей реакции происходит вследствие локального увеличения концентраций промежуточных соединений, присутствующих в непосредственной близости со следующим ферментом, как результат иммобилизации. Увеличение скорости анализа гомоцистеина может представлять собой важный фактор для коммерческого успеха диагностического средства, так как Km CBS и CBL находятся в миллимолярном диапазоне, а концентрации гомоцистеина в крови составляют приблизительно 10 мкМ. Известно, что ограниченные диффузией реакции для высокой скорости реакции требуют высокой концентрации ферментов. Иммобилизация ферментов при анализе циклическим преобразованием преодолевает трудности, которые могут существовать из-за низких аффинностей данных двух ферментов к их соответствующим субстратам. В еще одном предпочтительном осуществлении CBS и CBL иммобилизуют как гибридный белок. Дистанцию и ориентацию данных двух различных белков относительно друг друга можно контролировать во время конструирования экспрессирующего гибридный ген вектора. Посредством правильного позиционирования двух активностей в гибридном белке взаимопревращение гомоцистеина и цистатионина может проходить предельно быстро, практически в белке, так как дистанции диффузии между двумя активными центрами можно минимизировать. Следовательно, гомоцистеин в общей реакции ферментативного циклического преобразования, продуцирующей пируват и аммоний из L-серина, работает подобно кофактору. Анализ ферментативным циклическим преобразованием по настоящему изобретению можно проводить в виде ряда способов анализа. Простейший способ анализа предоставляет раствор реагентов, содержащий L-серин, CBS или ее производные, CBL или ее производные и различные буферы и вспомогательные вещества, необходимые для оптимизации ферментативных реакций. В особенно предпочтительном осуществлении настоящего изобретения CBS или ее производные и CBL или ее производные являются иммобилизованными на твердой поверхности. Иммобилизация ферментов на твердой поверхности, при которой сохраняется по существу вся активность фермента, хорошо известна в данной области. Для иммобилизации используют несколько способов, включающих в себя ковалентное связывание, физическую абсорбцию, захват и электрохимическое осаждение. В качестве примера для иммобилизации креатининдезаминазы и глутаматоксидазы на дериватизированные пропиламином стеклянные гранулы с регулируемыми порами (Rui et al., Ann. NY Acad. Sci., 672:264-271 (1992)) применяли глутаровый альдегид. Другие неограничивающие примеры включают в себя применение карбодиимидов для ковалентного прикрепления ферментов к дериватизированным сукцинатом стеклянным гранулам (Rui et al., выше) и применение гетеро- или гомобифункциональных перекрестно-сшивающих реагентов (например, SPDP, N-сукцинимидил-3-(2-пиридилдитио)пропионат; SMPT, сукцинимидилоксикарбонил- Как применяют здесь, “твердая поверхность” может включать в себя пористое или непористое не растворимое в воде вещество, которое может обладать любой из ряда форм, таких как полоска, стержневая частица, включая гранулы или подобное. Подходящие вещества хорошо известны в данной области и могут включать в себя, например, бумагу, фильтровальную бумагу, нейлон, стекло, керамику, оксид кремния, оксид алюминия, диатомовую землю, целлюлозу, полиметакрилат, полипропилен, полистирол, полиэтилен, поливинилхлорид и производные каждого из них. Твердая поверхность может представлять собой стороны и дно контейнера для тестирования или может представлять собой вещество, добавленное в контейнер для тестирования. В одном предпочтительном осуществлении твердая поверхность содержит гранулу, добавляемую в контейнер для тестирования. Дрожжи и бактерии обладают ферментами, преобразующими L-серин в пируват, активные вещества, на которые иногда ссылаются как на сериндезаминазу или сериндегидратазу. Для значительного уменьшения или удаления данной активности, которая может вносить значительный вклад в фон при анализе гомоцистеина циклическим преобразованием, можно применять общепризнанные способы мутагенеза. В частности, можно генетически манипулировать Saccharomyces cerevisiae с применением простого способа, известного как “разрыв гена”, для удаления активности сериндезаминазы, которая необходима для (1) синтеза изолейцина и (2) роста на серине как на источнике углерода и азота. При делеции гена cha1 дрожжей удаляется основная активность сериндезаминазы, таким образом, снимается одна из потенциальных фаговых проблем анализа циклическим преобразованием. Кроме того, делеция cha1 может являться пригодной для отбора гибридных белков CBS-CBL и для генетического конструирования данных гибридных белков. Штамм с делецией cha1 не способен к росту на серине как единственном источнике углерода или азота. Гибридный ген CBS-CBL способен к комплементации делеции потому, что система циклического преобразования приводит к такому же конечному результату, как и сериндезаминаза, поскольку гомоцистеин доступен как “кофактор”. Таким образом, для отбора и поддержания усовершенствованных гибридов CBS-CBL можно применять делецию cha1, особенно если популяция гибридных генов мутирована и помещена в однокопийные векторы или в хромосомы дрожжей. Клетки, растущие быстрее на среде с серином или обладающие сниженной потребностью в гомоцистеине, могут нести мутации, кодирующие более быстрые ферменты или белки с большей аффинностью к гомоцистеину и цистатионину. Настоящее изобретение в качестве альтернативного способа оценки количества гомоцистеина в образце относится к применению транскрипционного фактора, связывающего метаболиты. Когда связывающий метаболиты транскрипционный фактор связывается с соответствующим ему метаболитом, комплекс с фактором связывается со специфической рецепторной полинуклеотидной последовательностью. Связывание комплекса с транскрипционным фактором с его рецепторной консенсусной последовательностью можно контролировать, и оно коррелирует с количеством присутствующего в образце метаболита. В данной области известен ряд транскрипционных факторов, связывающихся со специфическими сайтами связывания на ДНК при ассоциации с низкомолекулярными соединениями или метаболитами. Например, E. coli обладает регулирующим оперон элементом, транскрипционным фактором MetR, регулирующим оперон Met в ответ на связывание с гомоцистеином (Cai et al., Biochem. Biophys. Res. Commun., 163:79-83(1989)). MetR или его производные регулируют такие гены, как MetE (кобаламиннезависимая метионинсинтаза), MetH (кобаламинзависимая метионинсинтаза) и GlyA (серингидрогеназа). MetR и его производные предпочтительно связываются с их консенсусными последовательностями ДНК в присутствии гомоцистеина и находятся в цитоплазме клетки в свободном виде при его отсутствии. Связывание MetR в присутствии гомоцистеина можно использовать для оценки количества гомоцистеина в образце, в котором предполагают наличие гомоцистеина. В конкретном осуществлении изобретения анализ включает в себя полинуклеотидную последовательность, кодирующую консенсусную последовательность рецепторного полинуклеотида для MetR, иммобилизованную на твердой подложке, таким образом, который обеспечивает доступность консенсусной рецепторной последовательности для связывания с MetR. Нуклеотидная последовательность может представлять собой, например: GTTAATGTTGAACAAATCTCATGTTGCGTG (SEQ ID No.: 11) Образец, такой как плазма, кровь или моча, обрабатывают для высвобождения любого связанного с белками гомоцистеина до смешивания образца с раствором реагента, содержащим MetR в буферном средстве, в присутствии иммобилизованной консенсусной рецепторной последовательности в условиях, способствующих формированию комплекса. После периода инкубации твердую подложку отмывают для удаления несвязавшихся реагентов и оценивают количество оставшегося связанным с твердой подложкой MetR. Определение количества оставшегося связанным с твердой подложкой MetR обычно может представлять собой ELISA, поверхностный плазмонный резонанс, т.е. BIACORE или просто стандартный анализ белка, и данное количество коррелирует с количеством присутствующего в образце гомоцистеина. Обработку образца для высвобождения гомоцистеина можно выполнить при помощи восстанавливающего средства. Как описано выше, в данной области хорошо известен ряд средств. Обычно образец смешивают с буферным раствором, содержащим количество восстанавливающего средства, достаточное для высвобождения по существу всего гомоцистеина из любых ассоциированных с ним белков, присутствующих в образце. Применяемые в большинстве случаев средства включают в себя в качестве неограничивающих примеров борогидрид натрия и калия, Белок MetR также можно модифицировать, чтобы он включал детектируемую метку, такую как хромофор, флуорофор и т.п., для увеличения скорости и чувствительности анализа. Добавление детектируемой метки может уменьшить период времени, необходимый для проведения анализа, посредством элиминации определенных стадий манипуляции с образцом, например стадий дополнительной отмывки. Введение флуорофора, испускающего свет при длине волны, поглощаемой олигонуклеотидами в консенсусной связывающей последовательности, позволяет проводить измерение переноса энергии флуоресценции для определения количества связавшегося с консенсусной последовательностью белка. Способы, которые можно применять для измерения передачи флуоресценцией, включают в себя, например, резонансный перенос энергии флуоресценции (FRET) или анализ сцинтилляций при сближении. Специфичность анализа также можно усилить дополнительными модификациями связывающей MetR консенсусной последовательности посредством добавления дополнительной детектируемой метки в связывающий домен. Дополнительно к белку MetR можно добавить определенную пептидную последовательность (или аффинную метку) для содействия очистке и детекции. Аффинные метки известны в данной области и включают в себя в качестве неограничивающих примеров полигистидин, стреп-метку, “flag”, последовательности c-myc и т.п. Метки широко применяют для аффинной очистки рекомбинантно экспрессированных белков, и многие доступны коммерчески. Кроме того, мутационные изменения с целью улучшения аффинности связывания рекомбинантно экспрессируемого MetR можно применять для предоставления дополнительных реагентов для анализа. В отдельном осуществлении настоящего изобретения количество гомоцистеина в образце можно определить посредством ферментативной утилизации гомоцистеина в образце. Преобразование гомоцистеина при клеточном метаболизме обычно высвобождает протон из сульфгидрильного остатка гомоцистеина. Высвобождение данного протона применяли для измерения аффинности связывания гомоцистеина с независимой от кобаламина метионинситазой E. coli (Jerret et al., Biochemistry, 36:15739-15748 (1997)). Катализируемые белками, такими как метионинсинтаза, ферментативные реакции, как правило, нуждаются в кофакторах или дополнительных субстратах для завершения реакции. Если данные кофакторы и субстраты не присутствуют, гомоцистеин останется связанным с ферментом. В настоящем способе измерение высвобождаемого протона удаляет протон, высвобождаемый при ферментативной реакции, направляя реакцию в сторону поглощения присутствующего в образце гомоцистеина. Измерения изменения pH, ассоциированного с высвобождаемыми протонами, обеспечивает определение количества гомоцистеина в образце. В предпочтительном осуществлении для анализа можно применять, например, независимую от кобаламина метионинсинтазу. Реагенты для применения по настоящему изобретению для удобства можно предоставлять в наборе для тестирования для применения в способе анализа. Типичный набор включает в себя упакованную комбинацию контейнеров для тестируемого раствора, L-серин, CBS или ее производное и CBL или ее производное и дополнительные буферы и вещества, необходимые для оптимальных условий реакций. Важно, что вместе с любым/всеми реагентами набора и/или ферментами также можно включить ряд консервантов и/или стабилизаторов для продления срока хранения, так же как срок службы во “включенном состоянии” при тестировании образца. Набор может дополнительно содержать восстанавливающие средства или CGL для предварительной обработки образца до анализа циклическим преобразованием. Кроме того, набор может включать в себя лактатдегидрогеназу, NADH и краситель, способный к видимому изменению окраски при окислении. В соответствующих обстоятельствах один или более реагентов в наборе можно предоставить в растворе (с консервантами и/или стабилизаторами или без них) или в качестве сухого порошка, обычно лиофилизированного, включающего наполнители, консерванты и/или стабилизаторы, который при растворении может обеспечить раствор реагентов с соответствующими концентрациями для проведения анализа по настоящему изобретению. Для увеличения универсальности данного изобретения реагенты можно предоставлять в упакованном наборе, в одном и том же или раздельных контейнерах, так чтобы соотношение реагентов обеспечивало существенную оптимизацию способа и анализа. Каждый из реагентов может находиться в отдельном контейнере, или разные реагенты можно объединять в одном или более контейнерах в зависимости от стабильности реагентов. Для удобства применяемые в способе анализа по настоящему изобретению реагенты можно предоставлять в заранее установленных количествах. Набор для тестирования также может содержать письменные инструкции по применению реагентов и/или по проведению конкретного анализа, например, в форме вложенного вкладыша. Следующие примеры предложены для иллюстрации, но не для ограничения. Пример 1 Клонирование и экспрессия гена цистатионин- В данном примере для клонирования гена CBL E. coli применяли стандартные способы ПЦР. Выделенный ген встраивали в экспрессирующий вектор с последовательностью аффинной метки (6HIS), а экспрессированный белок очищали на аффинной колонке. Из клеточных лизатов очищали существенное количество активного вещества фермента CBL клетки-хозяина, трансформированной конструкцией экспрессирующего вектора. Клонирование гена цистатионин- Для амплификации гена CBL E. coli применяли стандартные способы ПЦР с применением системы EXPAND HIGH FIDELITY PCR (Roche Biochemicals) и с применением компетентных клеток E. coli ONE SHOT TOP10 (Invitrogen) в качестве источника геномной ДНК. Последовательности праймеров для ПЦР для клонирования CBL являлись следующими: (N-концевой праймер для CBL: 5′-CGACGGATCCGATGGCGGACAAAAAGCTTG-3′; SEQ ID No.: 1) и (C-концевой праймер для CBL: 5′-CGCAGCAGCTGTTATACAATTCGCGCAAA-3′; SEQ ID No.: 2). Затем амплифицированный в ПЦР продукт гена CBL очищали посредством электрофореза в агарозном геле, расщепляли рестрикционными эндонуклеазами BamHI и PvuII и снова очищали посредством электрофореза в агарозном геле. Очищенный ген CBL E. coli затем лигировали в очищенный в геле обработанный BamHI/PvuII вектор для экспрессии белков pRSETB (Invitrogen). Полученная конструкция предоставляла ген CBL под контролем транскрипционного промотора T7, клонированный в рамку с лидерной последовательностью из шести аминоконцевых остатков гистидина (6HIS) (обозначаемая здесь далее как 6HISCBL), которая обеспечивала удобную экспрессию, аффинную очистку и анализ иммунным окрашиванием рекомбинантного белка 6HISCBL из клеток E. coli. Последовательность ДНК гена цистатионин- Экспрессия и очистка белка цистатионин- Плазмидную конструкцию pRSETB (6HISCBL) трансформировали в штамм E. coli BL21 (DE3)pLysS (Invitrogen), распределяли на планшетах со средой LB/Agar, содержащих (100 мкг/мл ампициллина, 35 мкг/мл хлорамфеникола) и затем инкубировали в течение ночи при 37°C. Единичные бактериальные колонии затем переносили и в течение ночи растили в колбах, содержащих 100 мл среды для выращивания LB, содержащей ампициллин и хлорамфеникол. 100 мл ночной культуры затем добавляли во множество колб, содержащих 1 л свежей среды для выращивания LB, содержащей ампициллин, и позволяли расти до достижения A600 в размере 0,6 (приблизительно 2,5 часов). Затем для индукции экспрессии белка CBL (регулируемой промотором T7) к среде для выращивания добавляли изопропил- Обнаружено, что очищенный белок 6HISCBL элюировался преимущественно в элюирующих буферах 2 и 3 (EB2 и EB3). Содержащий CBL элюат затем концентрировали с применением центробежной фильтровальной камеры (Amicon) и подвергали диализу в течение ночи при 4°C против буфера хранения (50 мМ TrisHCl, pH 8, 100 мМ NaCl, 5 мМ PLP), разделяли на аликвоты и хранили замороженными при -80°C. Затем вычисляли концентрацию очищенного гомогенного фермента CBL на основании коэффициента экстинкции при 280 нм и массы субъединицы 43312 Да (Clausen et al. Biochem. 36:12633-42 (1997)). Типичный выход очищенного 6HISCBL находился в диапазоне от 20 до 30 мг 6HISCBL на грамм клеточной пасты E. coli. Анализ активности CBL Анализ активности CBL аффинно очищенного рекомбинантного белка 6HisCBL, экспрессируемого с плазмидной конструкцией pRSETB (6HisCBL), проводили по существу, как ранее описано (Clausen et al., Biochem. 36:12633-42 (1997) включено сюда в качестве ссылки). Кратко, анализируемые смеси содержали 100 мМ Tris/HCl (pH 8,5), 5 мМ L-цистатионина, 1 мМ 5,5′-дитиобис-(2-нитробензойную кислоту) (DTNB; реагент Эллмана) и фермент 6HisCBL в конечном объеме реакционной смеси величиной 1 мл. За реакцией следили посредством записи увеличения поглощения при 412 нм с течением времени. Такую же ферментативную реакцию проводили на образцах с известными концентрациями субстрата для получения стандартной кривой. В основном получали всю активность CBL, обнаруженную в целом клеточном экстракте, и ее обнаруживали в экспериментально очищенных образцах рекомбинантного фермента 6HisCBL, полученного с плазмидной конструкцией pRSETB (6HisCBL). Пример 2 Клонирование и экспрессия гена дрожжевой цистатионин- В данном примере для клонирования гена CBS из дрожжей применяли способы ПЦР. Фрагмент ПЦР, содержащий ген CBS, встраивали в экспрессирующий вектор с аффинной меткой, которую добавляли для обеспечения очистки экспрессируемого продукта гена, и без нее. В качестве источника гена CBS применяли лабораторный штамм Saccharomyces cerevisiae (5153-11-1 (his3 met2)). Клетки растили на среде YEPD при 32°C, осаждали центрифугированием, отмывали водой и переносили в 200 мкл буфера для плазмид дрожжей (100 мМ NaCl, 10 мМ Tris (pH 8), 1 мМ ЭДТА, 0,2% SDS). Клеточную суспензию смешивали с равным объемом 0,45 мм стеклянных гранул и обрабатывали на центрифуге типа “вортекс” при высокой скорости 3 × 30 секунд. Жидкость удаляли пипеткой и центрифугировали две минуты в микроцентрифуге для удаления клеточного дебриса. Супернатант экстрагировали PCI (фенол:хлороформ:изоамиловый спирт (25:24:1)) и хлороформом. Добавляли хлорид натрия (5 М) (приблизительно 20 мкл) и ДНК осаждали равным объемом изопропанола. Данную геномную ДНК дрожжей подвергали амплификации посредством ПЦР с праймерами CI-001 (CGGGGATCCTGATGCATGCATGATAAGGA; SEQ ID No.: 3) и CI-012 (CGGCATTACCGTTTCACTAATTTATTG; SEQ ID No.: 4), которые являются опубликованными нуклеотидными последовательностями, фланкирующими структурный ген дрожжевой CBS. Праймер CI-001 (SEQ ID No: 3) содержит сайт BamHI, обнаруженный в нетранслируемом 3′-конце гена CBS. Для добавления последовательности BamHI на 5′-конец фрагмента ПЦР ген дополнительно амплифицировали посредством ПЦР с CI-001 (SEQ ID No.: 3) и CI-002 (GTTGGATCCGGCATTACCGTTTCAC; SEQ ID No.: 5). Конечный продукт ПЦР представлял собой последовательность ДНК приблизительно из 1937 пар оснований, включающих 1521 пару оснований, кодирующих белок CBS. Добавочная ДНК, фланкирующая кодирующую последовательность, содержит последовательности природного промотора и терминации транскрипции гена CBS дрожжей. Выше от гена CBS встраивали более сильный промотор для обеспечения более высоких уровней экспрессии белка. В стандартной реакции ПЦР с применением праймеров CI-009 (GGTGGATCCATCAATAGATACGTACGTCCT; SEQ ID No.: 6) и CI-010 (TTTTAGTTTATGTATGTGTTTTTTG; SEQ ID No.: 7) из той же геномной ДНК дрожжей клонировали дрожжевой промотор TPI1. Для присоединения кодирующего участка CBS к промотору TPI1 применяли адаптерный праймер CI-011 (AACACATACATAAACTAAAAATGACTAAATCTGAGCAGCAA; SEQ ID No.: 8), включающий 20 оснований промотора TPI1 и первые 21 основания гена CBS. Полученную выше последовательность CBS амплифицировали посредством ПЦР с праймерами CI-001 (SEQ ID No.: 3) и CI-011 (SEQ ID No.: 8). Конечный продукт ПЦР смешивали с продуктом-промотором TPI1, как указано выше, и смесь амплифицировали посредством ПЦР с праймерами CI-001 (SEQ ID No.: 3) и CI-009 (SEQ ID No.: 6) для получения гибридного гена, обозначенного “TPICBS”, который включает промотор TPI1, кодирующую последовательность CBS и терминатор транскрипции CBS. Каждый из продуктов ПЦР CBS и TPICBS содержит сайты BamHI на обоих 5′- и 3′-концах кодирующей области. Данные продукты ПЦР очищали PCR WIZARD PREP (Promega), следуя рекомендациям производителя, и разрезали BamHI. Полученные нуклеотидные BamHI-фрагменты разделяли на 1% агарозном геле с TBE и очищали с применением бумаги NA45 (S&S). Для экспрессии гена CBS применяли челночный вектор дрожжей, E. coli C1-1, несущий уникальный сайт BamHI в гене устойчивости к тетрациклину. Вектор C1-1 разрезали BamHI и обрабатывали креветочной щелочной фосфатазой, следуя инструкциям производителя (Roche Biochemicals). BamHI-фрагменты CBS и TPICBS лигировали в линеаризованный вектор C1-1. Продукт реакции лигирования трансформировали в клетки TOP10 (Invitrogen), которые помещали на планшеты LB с ампициллином. Устойчивые к ампициллину трансформанты переносили на планшеты LB с ампициллином и LB с тетрациклином для определения клонов, чувствительных к тетрациклину. Выбирали несколько клонов, чувствительных к тетрациклину, выращивали в течение ночи на планшетах LB с ампициллином и далее обрабатывали посредством щелочного лизиса для получения плазмидной ДНК. ДНК разрезали BamHI и разделяли на 1% агарозном геле для определения того, какие плазмиды содержат вставки CBS или TPICBS. Несущие вставки плазмиды C1-1 трансформировали в дрожжевой штамм, INVSc-1, диплоидный штамм, коммерчески доступный в Invitrogen. INVSc-1 несет мутацию в обеих копиях гена LEU2 и нуждается в гене LEU2 плазмиды C1-1 для селективного роста на среде без лейцина. INVSc-1 трансформировали родственными C1-1 плазмидами с применением набора для трансформации дрожжей из Invitrogen, следуя инструкциям производителя. Данный набор включает в себя создание сферопластов клеток зимолиазой с последующей обработкой кальцием для трансформации ДНК. Трансформанты помещали на синтетическую среду, содержащую 1 М сорбит, дрожжевые азотистые основания и дрожжевые добавки за исключением лейцина. Трансформанты становились видимыми на агаре через 3 дня инкубации при 32°C. Присоединение полигистидина к цистатионин- К 3′-концу кодирующей области CBS и в векторной конструкции с CBS, и в векторной конструкции с TPICBS добавляли последовательности ДНК, кодирующие шесть остатков гистидина. К исходному клону CBS добавляли кодирующие полигистидин последовательности посредством амплификации полученного выше фрагмента ПЦР CBS с праймерами CI-017 (ATGATGATGATGATGATGACCTGCTAAGTAGCTCAGTAA; SEQ ID No.: 9) и CI-002 (SEQ ID No.: 5). Полученные продукты ПЦР содержали природную промоторную область, кодирующую область CBS, и дополнительную кодирующую последовательность для глицина и шести остатков гистидина. Остаток глицина добавляли для увеличения пространственной подвижности пептида из поли-his после трансляции. Продукт ПЦР поли-hisCBS очищали на геле от исходной ДНК CBS и после этого дважды последовательно амплифицировали посредством ПЦР с праймерами CI-002 (SEQ ID No.: 5) и CI-017 (SEQ ID No.: 9) для полного исключения исходной ДНК CBS. Подобным образом полученный выше продукт ПЦР TPICBS амплифицировали с праймерами CI-009 (SEQ ID No.: 6) и CI-017 (SEQ ID No.: 9). Продукт ПЦР очищали на геле и еще дважды амплифицировали с праймерами CI-009 (SEQ ID No.: 6) и CI-017 (SEQ ID No.: 9) до значительного уменьшения исходной последовательности ДНК TPICBS. Ген CBS также клонировали посредством ПЦР с праймерами CI-001 (SEQ ID No.: 3) и CI-018 (CATCATCATCATCATCATTAAATAAGAACCCACGCT; SEQ ID No.: 10) для получения полигистидиновой последовательности со стоп-кодоном TAA с последующей последовательностью терминации транскрипции. Данный короткий продукт ПЦР (“терминатор”) приблизительно 190 п.н. также очищали на геле и клонировали посредством ПЦР с теми же праймерами. Фрагменты поли-his CBS и “терминатора” объединяли в реакции ПЦР с праймерами CI-001 (SEQ ID No.: 3) и CI-002 (SEQ ID No.: 5). В окончательной реакции амплификации синтезировали молекулы нуклеиновой кислоты, обозначаемые “CBSH”, содержащие природный промотор CBS, полную кодирующую CBS область, один дополнительный остаток глицина, шесть остатков гистадила со стоп-кодоном и терминатор транскрипции CBS. Подобным образом фрагменты поли-his TPICBS и терминатора объединяли в реакции ПЦР с праймерами CI-001 (SEQ ID No.: 3) и CI-009 (SEQ ID No.: 6). Окончательный продукт нуклеиновой кислоты, обозначаемый “TPICBSH”, обладал той же последовательностью, что и CBSH, за исключением того, что промотор TPI1 заменял природный промотор CBS. CBSH и TPICBSH (каждый из которых содержал полигистидиновую последовательность аффинной метки) разрезали BamHI и лигировали в C1-1 так, как выполняли выше для последовательностей CBS и TPICBS. Смеси после лигирования трансформировали в INVSc-1. Чувствительные к тетрациклину клоны проверяли на наличие вставки соответствующих клонированных генов в C1-1 посредством анализа ферментами рестрикции. Пример 3 Клонирование, экспрессия и очистка гена дрожжевой цистатионин- В данном примере для клонирования гена цистатионин- Клонирование В качестве источника гена CBS применяли лабораторный штамм Saccharomyces cerevisiae (INVSc1) (Invitrogen). Единичную колонию дрожжей INVSc1 5 мин кипятили в 100 мкл деионизированной воды и 2 мин быстро центрифугировали на центрифуге типа “вортекс” в присутствии равного объема 0,45 мм стеклянных гранул. Один микролитр лизата данных клеток затем применяли в качестве источника ДНК для амплификации ПЦР дрожжевого гена CBS с применением праймеров CI-024 (GCGGGTCGACTATGACTAAATCTGAGCAGCAAGCC; SEQ ID No.: 12) и CI-025 (GCGTGCGGCCGCGTTATGCTAAGTAGCTCAG; SEQ ID No.: 13), содержащих фланкирующие последовательности для опубликованного дрожжевого гена CBS и сайты рестрикции для клонирования в экспрессионный вектор pGEX6P-2 (Amersham Pharmacia). Конечный продукт ПЦР (приблизительно 1548 п.н.) затем выделяли и очищали посредством экстракции ДНК из агарозного геля с применением коммерческого набора (Roche). Очищенный продукт ПЦР, содержащий ген CBS, фланкированный сайтами рестрикционных ферментов SalI(5′-конец) и NotI (3′-конец) затем расщепляли (посредством SalI и NotI), очищали в геле и лигировали в экспрессионный вектор pGEX6P-2 (Amersham Pharmacia). Для подтверждения идентичности амплифицированной ПЦР ДНК дрожжевому гену CBS применяли картирование рестрикционными ферментами. Последовательность ДНК клонированного гена цистатионин- Экспрессия и очистка Экспрессионный вектор pGEX6P-2, содержащий клонированный ген CBS (описанный выше), трансформировали в клетки TOP10 E. coli (Invitrogen). Отбирали единичные колонии и клеточные культуры растили на среде LB, содержащей 100 мкг/мл ампициллина при 30°C до достижения поглощения при 600 нм приблизительно 0,5 ОП. Для индукции экспрессии гибридного белка GST-CBS (регулируемой промотором tac) к среде для выращивания добавляли изопропил- Затем проводили аффинную очистку белка GST-CBS с применением аффинной смолы Glutathione Sepharose 4B по существу так, как описано в протоколе производителя (Amersham Pharmacia). Кратко, клетки суспендировали в забуференном фосфатом солевом растворе (PBS) и разрушали ультразвуком на льду. Затем добавляли Triton X-100 до конечной концентрации 0,1% (об./об.) и обработанный ультразвуком раствор перемешивали встряхиванием при комнатной температуре 30-45 минут. Клеточный лизат затем очищали от нерастворимого дебриса посредством центрифугирования, обрабатывали ДНКазой I и помещали на колонку с аффинной смолой Glutathione Sepharose 4B. Неспецифически связанные PBS белки отмывали 10 колоночными объемами, а желаемый белок GST-CBS затем элюировали из аффинной смолы добавлением 5 колоночных объемов элюирующего буфера (50 мМ Tris, pH 8, 1,4 мМ Анализ активности CBS Активность CBS очищенного дрожжевого гибридного белка GST-CBS осуществляли, как описано у Kashiwamata и Greenberg (Biochim. Biophy. Acta 212:488-500 (1970)). Данный анализ основывается на способности CBS к синтезу цистатионина из серина и гомоцистеина. Количество цистатионина, образуемое CBS, затем определяют спектрофотометрически посредством детекции хромогенной реакции с нингидрином при 455 нм и сравнением со стандартной кривой, полученной с применением образцов с известной концентрацией цистатионина. Активность CBS также определяли посредством способов ферментативного анализа, описанных ниже, или посредством других способов анализа, хорошо известных специалистам в данной области. Пример 4 Конструирование гибридной конструкции ADH2CYS4 Экспрессия и очистка дрожжевой цистатионин- В данном примере сильный промотор ADH2 встраивали выше гена CYS4 и конструировали в виде гибрида с дрожжевым геном CBS для стимулирования более высокой экспрессии белка. Промотор ADH2 регулируется посредством Adr1p и репрессируется тогда, когда дрожжевые клетки растут в содержащей глюкозу среде, а когда клетки растут на неподдающемся ферментации источнике углерода, репрессия снимается. Это позволяет регулировать экспрессию, если перепроизводство белка летально для клетки. Конструирование гена ADH2CYS4 Дрожжевую геномную ДНК амплифицировали с праймерами CI-029 (CTATATCGTAATAATGACTAAATCTGAGCAGCAAGCCGATTCA; SEQ ID No.: 15) и CI-017 (ATGATGATGATGATGATGACCTCGTAAGTAGCTCAGTAA; SEQ ID No.: 9) для получения целого гена CYS4 без стоп-кодона и терминатора транскрипции. Праймер CI-029 (SEQ ID No.: 15) содержит последние 13 оснований промоторной области ADH2, клонированной в дополнение к первым десяти кодонам гена CYS4. Праймер CI-017 (SEQ ID No.: 9) несет последовательность последних шести кодонов гена CYS4 за исключением стоп-кодона, плюс остаток глицила и шесть остатков гистадила. Конечный продукт ПЦР содержит кодирующую область CYS4 и дополнительную кодирующую последовательность глицина и шести остатков гистидина. Глицин добавляли для увеличения пространственной подвижности пептида поли-his после трансляции. Фрагмент “терминатора” получали посредством амплификации той же геномной ДНК с праймерами CI-018 (CATCATCATCATCATCATTAAATAAGAACCCACGCT; SEQ ID No.: 10) и CI-040 (AGGGCTCGAGGATCCCGGGGGTGCTATTATGAATGCACGG; SEQ ID No.: 16) для получения полигистидиновой последовательности со стоп-кодоном TAA с последующим терминатором транскрипции. В реакции ПЦР с CI-029/CI-017 получали фрагмент длиной приблизительно 1530 п.н., тогда как в реакции с CI-018/CI-040 получали фрагмент длиной приблизительно 331 п.н. Данные два фрагмента объединяли смешиванием их продуктов ПЦР и амплификацией с праймерами CI-029/CI-040. Конечный продукт обозначали как CYS4H, и он составлял приблизительно 1861 п.н. в длину. Для объединения промотора ADH2 с фрагментом гена CYS4H сначала получали продукт ПЦР с праймерами CI-027 (CGGGGATCCGACGTTTTGCCCGCAGGCGGGAAACC; SEQ ID No.: 17) и CI-028 (AGATTTAGTCATTATTACGATATAGTTAATAGTTGATAG; SEQ ID No.: 18) из той же дрожжевой геномной ДНК для получения последовательности промотора ADH2. Праймер CI-028 (SEQ ID No.: 18) содержит первые 12 нуклеотидов кодирующей области гена CYS4. Таким образом, между праймерами CI-028 (SEQ ID No.: 18) и CI-029 (SEQ ID No.: 15) в их соответствующих продуктах находились 25 нуклеотидов комплементарной ДНК. Продукт ПЦР ADH2 смешивали с фрагментом CYS4H и амплифицировали с праймерами CI- 027 (SEQ ID No.: 17) и CI-040 (SEQ ID No.: 16) для получения продукта в виде фрагмента длиной приблизительно 2201 п.н., обозначенного ADH2CYS4H. Каждый из данных продуктов ПЦР затем подтверждали анализом с рестрикционными эндонуклеазами. Продукт ADH2CYS4H клонировали в YEp24 посредством разрезания обеих ДНК BamHI и лигирования с применением ДНК-лигазы T4. Лигированные продукты трансформировали в клетки E. coli (TOP 10; Invitrogen) и помещали на планшеты со средой LB-агар, содержащей ампициллин. Плазмиды, полученные с применением набора Plasmid Preparation kit (Roche) согласно рекомендациям производителя из предполагаемых рекомбинантов, анализировали посредством рестрикционной эндонуклеазы. Те плазмиды из клонов, у которых обнаружили правильную вставку, трансформировали в S. cerevisiae (BJ5460) и помещали на синтетическую полную среду без урацила (SC-ura), дополненную 2% глюкозой. После двух суток предполагаемые рекомбинанты еще раз штрихами наносили на такой же тип планшетов для выделения колоний. Клетки индуцировали посредством выращивания в SC-ura с 2% этанола и 1% пептона, как правило, дополненной 0,1% глюкозой для предоставления клеткам возможности расти в течение 24 часов. После выращивания в течение ночи клетки собирали посредством центрифугирования, а клеточный осадок хранили при -70°C до лизиса. Клетки лизировали посредством ресуспендирования клеточного осадка в лизирующем буфере в объеме 0,05 от исходного объема. Затем добавляли стеклянные гранулы до мениска буфера. Всю суспензию центрифугировали на центрифуге типа “вортекс” четыре раза, каждый раз по 1 минуте. В промежутках между центрифугированием клетки охлаждали на льду в течение 1 минуты. Лизат переносили в новую пробирку и центрифугировали 10 минут при 10000 × g. Очищенный лизат наносили на колонку с His-Resin (Roche), а белок очищали согласно рекомендациям производителя. Частично очищенный белок извлекали в третей элюции. Пример 5 Примеры нециклической части форматов анализа гомоцистеина В данном примере демонстрируют нециклическую часть анализа пирувата и цистатионина. Анализ пирувата Пируват: Настоящее изобретение относится к измерению гомоцистеина, основанному на применении ферментов CBS и CBL для цикла гомоцистеина с получением пирувата и аммиака. Предсказано, что скорость образования пирувата и аммиака является пропорциональной исходной концентрации гомоцистеина в образце. Измерение данной скорости с применением соответствующего(их) фермента(ов) представляет собой возможность для осуществления способа выявления гомогенного гомоцистеина. В одном из аспектов настоящего изобретения разработан высокочувствительный способ количественного анализа пирувата. В данном способе пируватоксидаза преобразует пируват в ацетилфосфат, диоксид углерода и пероксид. Далее пероксидаза преобразует пероксид, TOOS и 4-аминоантипурин в хромоген с максимумом поглощения около 550 нм. Молярный коэффициент поглощения данного хромогена является довольно высоким и составляет приблизительно 36 л моль-1см-1. С применением компонентов реагентов, представленных в таблице 1, и параметров устройства Cobas FARA (Roche, Basel, Switzerland), перечисленных в таблице 2, пируват легко измеряли в мкМ диапазоне посредством раннего чтения реакции чистой и конечной точки приблизительно через 5-10 минут. Графики зависимости поглощения от времени для водных калибровочных растворов показаны на фигуре 1. Фактическая кривая калибровки показана на фигуре 2.
Анализ цистатионина Цистатионин измеряли посредством добавления CBL к описанному выше реагенту для пирувата. CBL преобразует цистатионин в гомоцистеин, пируват и аммиак. Затем пируват измеряли, как описано выше. Состав реагента, применявшегося для измерения цистатионина, приведен в таблице 3. Измерение можно проводить любым подходящим устройством. В настоящем примере применяли Cobas FARA (Roche, Basel, Switzerland). Типичные параметры реакции приведены в таблице 4. На фигуре 3 изображены графики зависимости поглощения от времени, полученные с применением водных растворов цистатионина. Реакция достигала конечной точки менее чем за десять минут.
На фигуре 4 изображены стандартные кривые, полученные с применением водных растворов цистатионина. Данные кривые представляют собой прямые, по меньшей мере, при диапазоне концентрации цистатионина от 0 до 100 мкМ. Образующийся хромофор оставался стабильным приблизительно 15-20 минут. Однако если оценивали образцы, содержащие концентрации цистатионина, большие в 10-20 раз, была выражена некоторая нестабильность хромофора. В некоторых экспериментах в реакционную смесь добавляли серин. Серин при уровне 250 мкМ не проявлял значимого эффекта. Более высокие уровни серина также допускались в данном анализе. Неучастие серина является важным наблюдением, так как присутствие серина необходимо для анализа гомоцистеина. Пример 6 Способы анализа гомоцистеина ферментативным циклическим преобразованием Данный пример относится к системам ферментативного циклического преобразования, применяющим для определения концентрации гомоцистеина или сигнальную систему на основе пируватоксидазы/пероксидазы, или сигнальную систему на основе лактатдегидрогеназы. Демонстрация циклического преобразования: система ферментативного циклического преобразования CBS/CBL/PO/HRP Демонстрировали систему циклического преобразования, где реакцию инициировали с применением гомоцистеина или цистатионина. Применяемые в данных экспериментах компоненты реагентов приведены в таблице 5. CBS и серин добавляли отдельно с последующим смешиванием образца с реагентом 1, содержащим все другие компоненты. Подходящие параметры Cobas FARA приведены в таблице 6. Цистатионин или гомоцистеин растворяли в воде для получения стандартов в соответствующих концентрациях. Графики зависимости поглощения от времени показаны на фигурах 5 и 6. После временной задержки длительностью от трех до шести минут графики представляют собой прямую линию. Типичную калибровочную кривую для гомоцистеина (фигура 7), хотя и не полностью прямую, можно применять для количественного определения концентраций гомоцистеина в подходящих образцах с применением соответствующего способа подбора кривой.
Восстановление дисульфидных связей В человеческой плазме присутствует очень малое количество гомоцистеина в восстановленном, не связанном дисульфидными связями состоянии. Большое количество гомоцистеина связано с белками или низкомолекулярными соединениями посредством дисульфидных связей. Восстановление дисульфидных связей в данных соединениях является необходимым для высвобождения гомоцистеина для измерения любым способом определения общего гомоцистеина. Данный пример демонстрирует то, что применение восстанавливающего средства сочетается с системой, применяющей лактатдегидрогеназу и NADH. Возможные кандидаты на роль восстанавливающих средств включают в себя DTE, DTT, n-ацетилцистеин, тиогликолевую кислоту, TCEP и т.п. Для применения данных соединений в способе по настоящему изобретению необходима точная регулировка концентрации восстанавливающего средства и параметров устройства. Начальные признаки того, что некоторые восстанавливающие средства могут препятствовать действию пируватоксидазы или пероксидазы хрена, обусловили поиск альтернативной системы детекции основанной на пирувате, описанной ниже. Демонстрация циклического преобразования: система CBS/CBL/LDH: Пируват подвергают взаимодействию с NADH в присутствии фермента лактатдегидрогеназы с образованием молочной кислоты и NAD. Количество пирувата можно определить посредством измерения уменьшения поглощения при 340 нм. Молярный коэффициент поглощения NADH приблизительно в шесть раз меньше, чем молярный коэффициент поглощения образуемого пероксидазой хромогена. Однако это не влияет на систему по настоящему изобретению, так как циклическое преобразование гомоцистеина посредством CBL/CBS позволяет образовывать относительно большие количества пирувата. Применяемая для получения данных система реагентов, представленная на фигурах 8, 9, 10 и 11, содержит серин, CBL и лактатдегидрогеназу в буфере HEPES (pH 7,2) в качестве первого реагента. Затем последовательно добавляли NADH в буфере Tris (pH 8,5) и CBS. Однако все компоненты можно объединять иначе для формирования системы измерения количества гомоцистеина из двух или трех реагентов. Концентрации реагентов приведены в таблице 7, а параметры Cobas FARA приведены в таблице 8. Концентрации NADH регулировали для обеспечения достаточной линейности, при этом обеспечивая поглощение образца плазмы при 340 нм.
Анализ водных растворов Система хорошо подходила для детекции гомоцистеина в водном растворе. Типичные графики зависимости поглощения от времени показаны на фигурах 8 и 9. Данные графики являлись линейными с небольшой или неявной фазой задержки. Скорости исчезновения NADH являлись относительно высокими. Более того, калибровочный график концентрации гомоцистеина в зависимости от поглощения являлся линейным при диапазоне концентраций в образце от 0 до 100 мкМ (фигура 10). Таким образом, для калибровки необходим только один калибратор, например, 25 мкМ, вместе с чистым реагентом. Для демонстрации эффективности способа при диапазоне концентраций гомоцистеина от 2 до 20 мкМ готовили растворы гомоцистеина в различных концентрациях. Результаты приведены в таблице 9 как для реакций в присутствии и в отсутствие DTT. Как указано выше, DTT служит для восстановления дисульфидных связей в образцах (например, сыворотка, плазма и т.д.). DTT не служил значительным препятствием катализируемой лактатдегидрогеназой реакции.
Анализ основанных на человеческой плазме контрольных препаратов Правильное функционирование системы циклического преобразования CBS/CBL/LDH демонстрировали здесь с применением основанных на человеческой плазме контрольных препаратов гомоцистеина, произведенных Bio-Rad Co. Для достижения оптимальной регенерации гомоцистеина применяли различные концентрации DTT, а также варьировали длительности времени инкубации. Длительности времени инкубации от 5 до 15 мин с концентрациями DTT от 1,6 до 3,2 мМ обеспечивали хорошие измерения количества гомоцистеина в пределах диапазонов, установленных для различных других способов. Образцы человеческой плазмы Анализ гомоцистеина циклическим преобразованием демонстрировал хорошие эксплуатационные качества с применением образцов человеческой плазмы с DTT в качестве восстанавливающего средства. Результаты хорошо согласовывались с результатами, полученными независимой лабораторией, применявшей способ оценки количества гомоцистеина Abbott IMx (фигура 11). Параметры Cobas FARA, применяемые в данном эксперименте, приведены в таблице 10, а компоненты реагентов приведены в таблице 7. В данном анализе применяли калибратор в виде водного гомоцистеина (25 мкМ). Графики зависимости поглощения от времени для водных образцов являлись линейными в отсутствие DTT и близкими к линейным в присутствии DTT. Однако графики, полученные с применением образцов плазмы, отличались, т.е. являлись нелинейными после первых трех минут. Оказалось, что в условиях, приведенных в таблице 7 и таблице 10, циклическое преобразование постепенно замедляется с течением времени. Тем не менее, представленное на фигуре 11 исследование корреляции демонстрирует относительно близкое совпадение между настоящим способом (приведенным в таблице 7 и таблице 10) с результатами, полученными посредством IMx при тестировании образцов человеческой плазмы.
В предпочтительном осуществлении измеряли скорости за первые три минуты после начала реакции. Вероятно, дополнительная оптимизация системы, включающая в себя изменение концентраций солей и детергентов, приведет к более согласующимся характеристикам водных образцов и образцов плазмы. Анализ являлся в равной степени точным, когда компоненты объединяли в две или три смеси реагентов. Пример 7 Сравнение способов анализа гомоцистеина В данном примере предпочтительный анализ гомоцистеина по данному изобретению тестировали и сравнивали с двумя существующими способами анализа, а конкретно – со способом ВЭЖХ и коммерчески доступным способом анализа IMx. Анализ по изобретению проводили с применением трех реагентов, как представлено в таблице 11. Первый реагент R1 изначально находился в форме порошка, и его растворяли до уровня 1,5 г порошка на 100 мл воды. Полный набор для анализа также содержал калибраторы, включающие в себя L-цистатионин с пиками при в различных уровнях в матриксе человеческой плазмы, а также два контрольных образца, приготовленные в обработанном матриксе человеческой плазмы.
При проведении данного анализа применяли коммерчески доступное устройство Cobas MIRA. Программное обеспечение Cobas MIRA допускало 50 программируемых 25-секундных циклов. В частности, устройство программировали согласно параметрам таблицы 12. В цикле 1 в кювету добавляли 30 мкл образца с последующими 25 мкл промывочной воды и 90 мкл жидкого реагента R1. В начале цикла 2 добавляли 25 мкл реагента SR1 с дополнительными 25 мкл промывочной воды. Затем смеси инкубировали 15 циклов (6,25 мин), чтобы позволить восстановителю высвободить любой связанный гомоцистеин и разрушить любой присутствующий эндогенный пируват. В данной точке добавляли 35 мкл реагента с ферментами CBS/CBL с дополнительными 15 мкл промывочной воды. Уменьшение поглощения при 340 нм измеряли в течение времени между циклами 18 и 32 как меру гомоцистеина в образце. Общее время анализа между добавлением образца и конечным считыванием данных поглощения составило 32 цикла, или 13 мин и 20 сек. Так как множество образцов запускали параллельно, дополнительные результаты анализа образцов посредством MIRA получали при скорости приблизительно один каждые 30-90 секунд в зависимости от длины единичного цикла и числа применяемых калибраторов устройства. С применением сходных программных параметров данный анализ можно адаптировать даже для более быстрых устройств (например, Roche INTEGRA, Hitachi, ACE и т.д.), которые могут выдавать даже большие скорости прохождения материала. Всего с применением анализа по данному изобретению и известных сравнительных способов анализа тестировали 24 образца. В таблице 13 ниже приведены концентрации гомоцистеина, полученные с применением данных трех способов. Фигуры 12-14 представляют собой соответствующие графики корреляции, графически представляющие результаты таблицы 13. Как можно видеть, способ по данному изобретению тесно коррелирует и с IMx, и с ВЭЖХ; с обычно наблюдаемым более близким согласием с IMx. Однако настоящий способ значительно менее дорогостоящий, его можно провести быстрее, и его можно провести с применением обычно доступных химических анализаторов. Изобретение демонстрирует очень хорошую точность между единичными циклами, как показано в таблице 14. Найдено, что обычные CV в пределах единичного цикла составляют менее чем 4% при концентрациях гомоцистеина приблизительно от 7 до 20 мкмоль/л.
Пример 8 Эффект при изменениях концентрации реагентов В данном примере проводили предпочтительный анализ по изобретению, описанный в примере 7, с изменениями некоторых компонентов реагентов, а именно LDH, NADH и серина. Это сделано для определения оптимальных количеств данных компонентов реагентов. Анализы проводили, как описано выше, с соответствующими изменениями в концентрациях соответствующих компонентов. В таблице 15 приведены результаты данных тестов.
Представленные в таблице 15 результаты давали возможность выбора конечных уровней компонентов реагентов, обеспечивающих достаточную чувствительность с минимальной интенсивностью скоростью чистого образца, при попытках также минимизировать стоимость реагентов. Пример 9 Действие изменения состава R1 на анализ циклическим изменением В данном примере определяли действие изменения pH посредством изменения буфера на Tris, 250 мМ при pH 8,4 и посредством добавления липазы и
Посредством изменения буфера реагента 1 на Tris, 250 мМ при pH 8,4 улучшали стабильность анализа. Это происходило потому, что CBS и CBL являются существенно более активными при pH 8,4 и скорость циклического преобразования увеличивается приблизительно в три раза. Для борьбы с проблемой, ассоциированной с замутненными (липемическими) образцами, которые иногда встречаются в клинических лабораториях, добавляли липазу и В добавление к изменениям, производимым в компонентах реагентов, также можно изменять объемы для оценки их действия на эффективность анализа. Данные изменения отражены в таблице 17, демонстрирующей типичные параметры циклического преобразования для Cobas MIRA. Следуя данным изменениям, проводили изучение корреляции на 86 образцах от людей с результатами, представленными в таблице 18.
Как показано в данной таблице сравнения, результаты данного анализа при измененном составе являлись очень точными при сравнении с коммерчески доступным анализом IMx. Пример 10 Применение других детергентов и изменение их концентрации В данном примере применяли различные детергенты в комбинации с ЭДТА в качестве компонентов R1. Известно, что ЭДТА стабилизирует липидные частицы. Целью снова являлось достижение точных результатов в том случае, когда образцы являлись мутными. В частности, изучали детергенты Brij-35 и Genapol X-80. Обнаружено, что для содействия минимизации неспецифического уменьшения поглощения из-за изменений мутности при анализе липемических образцов ЭДТА являлся чрезвычайно эффективным в концентрации 2,5 мМ. Концентрации детергента также изменяли, как показано в таблицах 19 и 20. Один из предпочтительных составов для реагента R1 приведен в таблице 21. Brij-35 присутствовал в на уровне 0,025% об./об., а ЭДТА присутствовал в концентрации 2,5 мМ. Липаза и
Когда Brij-35 и ЭДТА заменяли на Triton X-100 и липазу с Пример 11 Результаты, полученные при изменении восстанавливающего средства на TCEP и изменении концентрации TCEP. В данном примере восстанавливающее средство DTE заменяли на трис-(2-карбоксиэтилфосфин)гидрохлорид (TCEP). TCEP обладает тем преимуществом, что остается стабильным в очищенной воде длительный период времени. Замена DTE на TCEP уменьшает реакцию чистого реагента со значений приблизительно от 12 до 15 мА/мин до значений приблизительно от 4 до 5 мА/мин. Меньший уровень чистого реагента делает возможной большую точность анализа гомоцистеина. Чтобы найти оптимальную концентрацию TCEP для данного анализа гомоцистеина, тестировали диапазон концентраций. Результаты данных экспериментов приведены в таблице 23.
Концентрации TCEP: A = 6,3 мМ; B = 13,85 мМ; C = 26,44 мМ; D = 32,74 мМ; E = 39,03 мМ; F = 52,88 мМ. Пример 12 Определение эффекта изменения буфера R3 на фосфат и добавления в буфер глицерина Для получения смесей ферментов, которые могли бы обладать большей стабильностью и совместимостью с анализом, состав ферментов изменили на смесь, обладающую значительно большим сроком хранения, чем предыдущий состав. Значительные изменения включали в себя замену Tris-буфера на фосфат и добавление к буферу глицерина. Применяли фосфат калия в диапазоне концентраций от 50 мМ до 500 мМ при pH 7,6 и глицерин в диапазоне концентраций от 0,5% до 15% (об./об.). Как результат данных изменений срок хранения при 4°C значительно увеличился с нескольких недель до более чем года. В предпочтительных формах фосфат калия может находиться в диапазоне приблизительно от 50 мМ до приблизительно 250 мМ. Более предпочтительно, чтобы фосфат калия мог варьировать приблизительно от 75 мМ до приблизительно 150 мМ. Особенно эффективной концентрацией фосфата калия в данном эксперименте являлась концентрация 100 мМ. Глицерин может находиться в диапазоне приблизительно от 1 до 15%. Более предпочтительно, чтобы глицерин находился в диапазоне приблизительно от 5 до 13%. Особенно эффективной концентрацией глицерина в данном эксперименте являлась концентрация 10%. Пример 13 Результаты, полученные с применением различных устройств Для определения общего улучшения результатов при всех изменениях анализа оценки точности проводили на нескольких устройствах при записи специально предназначенного для данного устройства протокола. В таблице 24 приведены параметры и установки, применяемые по настоящему изобретению для Roche Hitachi 911, а в таблице 25 приведены те же параметры и установки для Beckman CX-5. Ниже приведены таблицы, демонстрирующие результаты этих тестов точности. Данные результаты значительно лучше, чем те, которые получены с предыдущими составами для анализа. На устройствах Hitachi 911 (таблица 26) и Beckman CX5 (таблица 27) для каждого показанного образца проводили 20 единичных циклов и во время единичного цикла определяли точность. Процент C.V. для образцов с такой низкой концентрацией, как 5,92 мкмоль/л, составлял 3,31%, для образцов с концентрацией в диапазоне от 23,01 мкмоль/л до 25,4 мколь/л – от 1,9% до 2,03%. На образцах со значениями гомоцистеина от 4,6 мколь/л до 27,0 мколь/л на устройстве Cobas MIRA Plus получена общая точность (таблица 28) от 7,3% до 2,6%. Каждый образец анализировали дважды, 2 раза в день, всего 20 дней.
Для данного эксперимента T1 получали посредством смешивания R1 и деионизированной воды в соотношении 9 к 8,1. Реагенты T2 и T4 соответствовали реагентам SR1 и SR2 соответственно.
Для данного примера реагент A получали посредством смешивания R1 и деионизированной воды в соотношении 9 к 8,1. Реагенты B и C соответствовали реагентам MIRA SR1 и SR2.
Пример 4 Адаптация анализа к системе с 2 реагентами Настоящее изобретение также можно адаптировать к системе, применяющей только два реагента. Это делает изобретение применимым в широком диапазоне устройств, включая устройства, в которых применяются только два реагента. В таблице 29 приведены составы реагентов системы с двумя реагентами, а в таблице 30 приведены параметры MIRA plus, отображающие то, как проводили анализ на данном химическом анализаторе. В таблице 31 приведены данные сравнения, полученные с применением системы из 3 реагентов, по сравнению с системой из 2 реагентов.
Пример 15 Адаптация настоящего изобретения к применению с одной точкой калибровки Анализ количества гомоцистеина по настоящему изобретению также адаптировали к использованию одной точки калибровки с нулевым калибратором, служащим как чистый. В таблице 32 ниже приведены параметры для проведения калибровки с одной точкой на устройстве Mira. Данные ниже (таблица 33) демонстрируют то, что применение одной точки калибровки в режиме среднего углового коэффициента позволяет получить результаты, которые по существу эквивалентны применению калибровочной кривой с несколькими точками.
Таблица 33 Калибровка с одной точкой по сравнению с калибровкой со множеством точек
Пример 16 Изучение анализа количества гомоцистеина, описанного в WO 00/28071 В публикации PCT WO 00/28071 подробно описаны два способа анализа гомоцистеина. В “первом осуществлении” анализ проводят без стадии восстановления. Для преобразования данного способа к способу, пригодному для измерений количества гомоцистеина в человеческой плазме, необходимо ввести отдельную независимую или гомогенную стадию восстановления. В публикации WO 00/28071 описана отдельная независимая стадия восстановления, возможно, потому, что применяют систему циклического преобразования с пируватоксидазой. Данная стадия приводит к инкубации в течение 20 мин, перед тем как можно будет анализировать восстановленные образцы, что значительно увеличивает общее время анализа и затраты на рабочую силу. Кроме того, если в клинической лаборатории образец необходимо анализировать более чем для одного анализируемого соединения, исходный образец сыворотки или плазмы необходимо разделить на две аликвоты, одну для анализа количества гомоцистеина и другую для любого другого необходимого лабораторного теста. Напротив, настоящее изобретение дает возможность гомогенного восстановления, так как применяют систему циклического преобразования с LDH (пример 7). Здесь, в примере 16 (таблица 34) показано, что DTT фактически мешает системе детекции с пируватоксидазой, описанной во “втором осуществлении” WO 00/28071. В частности, применяли составы реагента и DTT и проводили анализы с DTT и без по отношению к известным стандартам пирувата (12,5, 25 и 50 мкмоль/л) с применением устройства Cobas FARA. В случае теста с DTT 40 мкл каждого стандарта смешивали с 30 мкл раствора DTT с последующей инкубацией 100 сек. Затем добавляли 90 мкл реагента с пируватоксидазой и инкубировали 30 сек. Считывание данных поглощения проводили через 30, 60, 90, 120, 150, 180 и 210 сек. В случае теста “без DTT” проводили те же процедуры за исключением того, что не добавляли DTT. В таблице 34 приведены результаты данного эксперимента с применением считывания данных в течение 210 сек. Фигуры 15 и 16 дополнительно иллюстрируют результаты данного теста.
Эти данные отчетливо демонстрируют необходимость отдельной стадии восстановления, описанной во “втором осуществлении” WO00/28071. Присутствие восстанавливающего средства мешает системе детекции с пируватоксидазой, применяемой для анализа количества пирувата. Следовательно, представленные здесь данные подтверждают, что занимающая время отдельная независимая стадия восстановления является необходимой во “втором осуществлении”. Кроме того, стадия независимого восстановления, описанная в WO 00/28071, вероятно, будет значительно повышать сложность и цену анализа относительно настоящего изобретения. Хотя указанное выше изобретение подробно описано посредством иллюстраций и примера с целью ясности понимания, очевидно, что в рамках прилагаемой формулы изобретения можно осуществлять определенные изменения и модификации. Следовательно, объем данного изобретения необходимо определять, основываясь не на предыдущем описании, но вместо этого его необходимо определять, основываясь на прилагаемой формуле изобретения наряду с полным объемом ее эквивалентов. Все публикации и патенты, цитируемые в данной заявке, полностью включены в нее в качестве ссылки для всех целей в таких пределах, как если бы каждая индивидуальная публикация или патент являлись указанными отдельно.
Формула изобретения
1. Способ оценки количества гомоцистеина и/или цистатионина в растворе, включающий: (a) создание реакционной смеси путем контакта указанного раствора, содержащего гомоцистеин и/или цистатионин, с восстановительным средством, цистатионин- (b) определение количества пирувата и/или аммиака, присутствующего в реакционной смеси; и (c) оценку количества гомоцистеина и/или цистатионина, присутствующего в растворе, на основании количества образовавшегося пирувата и/или аммиака, где реакционная стадий (а) длится менее 15 мин. 2. Способ по п.1, отличающийся тем, что L-серин добавляют в реакционную смесь до конечной концентрации, составляющей по меньшей мере от приблизительно 1 мкМ до приблизительно 50 мМ. 3. Способ по п.2, отличающийся тем, что указанный L-серин добавляют в реакционную смесь до конечной концентрации, составляющей по меньшей мере от приблизительно 10 мкМ до приблизительно 40 мМ. 4. Способ по п.1, отличающийся тем, что количество пирувата определяют посредством ферментативного преобразования пирувата в лактат. 5. Способ по п.4, отличающийся тем, что преобразование пирувата в лактат проводят посредством добавления лактатдегидрогеназы, и NADH или солей, сольватов и других химически-модифицированных форм NADH. 6. Способ по п.5, отличающийся тем, что указанную лактатдегидрогеназу добавляют в реакционную смесь до конечной концентрации, составляющей по меньшей мере от приблизительно 30 Ед/л до приблизительно 5000 Ед/л. 7. Способ по п.6, отличающийся тем, что указанную лактатдегидрогеназу добавляют в реакционную смесь до конечной концентрации, составляющей по меньшей мере от приблизительно 30 Ед/л до приблизительно 3000 Ед/л. 8. Способ по п.5, отличающийся тем, что указанный NADH добавляют в реакционную смесь до конечной концентрации, составляющей по меньшей мере от приблизительно 0,1 мМ до приблизительно 2 мМ. 9. Способ по п.8, отличающийся тем, что указанный NADH добавляют в реакционную смесь до конечной концентрации, составляющей по меньшей мере от приблизительно 0,1 мМ до приблизительно 1,5 мМ. 10. Способ по п.5, отличающийся тем, что общую степень окисления NADH или солей, сольватов и других химически-модифицированных форм NADH в NAD+ определяют посредством взаимодействия NAD+ с красителем, способным изменять окраску при окислении. 11. Способ по п.10, отличающийся тем, что указанный краситель выбран из группы, состоящей из 5,5′-дитиобис(2-нитробензойной кислоты), 2,6-дихлорфенолиндофенола, соединения тетразола, феназинметосульфата, метилвиологена или производных каждого из них. 12. Способ по п.1, отличающийся тем, что количество пирувата определяют посредством ферментативного преобразования пирувата в пероксид водорода. 13. Способ по п.12, отличающийся тем, что указанный фермент представляет собой пируватоксидазу. 14. Способ по п.13, отличающийся тем, что анализ дополнительно включает использование пероксидазы и водорастворимого донора водорода, который после отдачи водорода формирует стабильно окрашенный продукт, который можно определять спектрофотометрически. 15. Способ по п.14, отличающийся тем, что водорастворимый донор водорода представляет собой производное N-алкил-N-сульфопропиланилина. 16. Набор для тестирования, используемый в способе оценки количества гомоцистеина в тестируемом растворе по п.1, содержащий цистатионин- 17. Способ оценки количества гомоцистеина в образце, включающий стадии: (a) контактирования образца с восстанавливающим средством в период времени, достаточный для высвобождения восстановленного гомоцистеина из связывающего гомоцистеин белка; (b) контактирования указанного восстановленного гомоцистеина со связывающим метаболиты гомоцистеина транскрипционным фактором в условиях, способствующих образованию комплекса между указанным восстановленным гомоцистеином и указанным транскрипционным фактором, связывающим метаболиты гомоцистеина; (c) смешивания указанного образца с консенсусной полинуклеотидной последовательностью, специфицески распознаваемой указанным комплексом гомоцистеин/транскрипционный фактор, для связывания указанного комплекса гомоцистеин/транскрипционный фактор с указанной консенсусной полинуклеотидной последовательностью; (d) определения количества комплекса гомоцистеин/транскрипционный фактор, связанного с консенсусной полинуклеотидной последовательностью; и (e) сопоставление количества присутствующего в указанном образце гомоцистеина с указанным определенным количеством, где стадии (а)-(с) длятся менее 15 мин. 18. Способ оценки количества гомоцистеина в образце, включающий стадии: (a) получения реакционной смеси, включающей указанный образец, серин, лактатдегидрогеназу, NADH или соли, сольваты и другие химически-модифицированные формы NADH, сохраняющие его активность, и восстановитель, способный высвобождать связанный с белками гомоцистеин в образце; (b) инкубирования указанной реакционной смеси в период времени, достаточный для восстановления преобладания связанного с белками гомоцистеина; (c) добавления смеси ферментов, включающей цистатионин- (d) измерения образования NAD+или исчезновения NADH в период времени измерения гомоцистеина в образце, где стадии (а)-(с) указанного способа осуществляют менее чем за 15 мин. 19. Способ оценки количества гомоцистеина в образце, включающий стадии: (a) получения реакционной смеси, содержащей указанный образец, серин, восстанавливающий агент, цистатионин- (b) оценки количества присутствующего в указанной реакционной смеси пирувата посредством добавления к указанной реакционной смеси лактатдегидрогеназы и NADH, и контроля образования NAD+ или исчезновения NADH с течением времени в указанной реакционной смеси, где стадия (а) длится менее 15 мин. 20. Способ последовательной оценки множества жидких содержащих гомоцистеин образцов, включающий стадии: (а) получения реакционной смеси посредством помещения в контейнер одного из указанных образцов вместе с серином, цистатионин- (b) циклического преобразования в указанной реакционной смеси гомоцистеина в цистатионин и обратного преобразования цистатионина в гомоцистеин, сопровождающегося образованием пирувата; (c) оценки количества присутствующего в реакционной смеси пирувата посредством контроля образования там NAD+или исчезновения NADH с течением времени; и (d) повторение стадий (а)-(с) для каждого из указанных образцов, при этом временной интервал между соответствующими стадиями создания реакционной смеси для каждого из указанных образцов занимает приблизительно до 15 мин. 21. Выделенный фермент цистатионин- 22. Выделенный фермент цистатионин- 23. Способ оценки количества гомоцистеина и/или цистатионина в растворе, включающий: (а) создание реакционной смеси путем контакта указанного раствора, содержащего гомоцистеин и/или цистатионин, с восстановительным средством, цистатионин- (b) определение количества пирувата и/или аммиака, присутствующего в реакционной смеси, и (c) оценку количества гомоцистеина и/или цистатионина, присутствующего в растворе, на основании количества образовавшегося пирувата и/или аммиака, где реакционная стадия (а) длится менее 15 мин.
РИСУНКИ
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
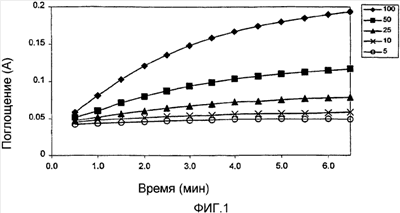
 -синтазы (CBS) или его производных и ферментативного преобразования цистатионина в гомоцистеин, пируват и аммиак при помощи цистатионин-
-синтазы (CBS) или его производных и ферментативного преобразования цистатионина в гомоцистеин, пируват и аммиак при помощи цистатионин- -лиазой (CGL) или ее производным на период времени, достаточный для удаления цистатионина из реакционной смеси посредством преобразования цистатионина в
-лиазой (CGL) или ее производным на период времени, достаточный для удаления цистатионина из реакционной смеси посредством преобразования цистатионина в  -кетоглутарат. После удаления цистатионина цистатионин-
-кетоглутарат. После удаления цистатионина цистатионин- 200 мЕд/л
200 мЕд/л