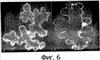
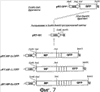
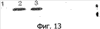Патент на изобретение №2344174
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) СПОСОБЫ И КОНСТРУКЦИИ ДЛЯ ПОВЫШЕНИЯ СОДЕРЖАНИЯ ОТДЕЛЬНЫХ АМИНОКИСЛОТ В СЕМЕНАХ
(57) Реферат:
Растения трансформируют конструкцией с нуклеотидной последовательностью, включающей ткане- или органоспецифичные регуляторные последовательности, контролирующие транскрипцию во время выбранных стадий морфогенеза. Регуляторные последовательности сцеплены с химерной нуклеотидной последовательностью, не имеющей своего собственного терминирующего кодона и содержащей на своем С-терминальном конце кодоны, кодирующие выбранное количество и комбинацию аминокислотных остатков. Химерная нуклеотидная последовательность обеспечивает направленную экспрессию специфического растительного белка-носителя, имеющего стабильное аминокислотное удлинение цепи. Стабильность конструкции определяют с помощью бесклеточной системы in vitro с применением кодоновых кассет, имеющих выбранную комбинацию и количество кодонов. Стабильная трансформация растений обеспечивает повышенное содержание аминокислот в тканях растений, в частности, в мембранах масляных телец и клеточных стенках семян, что обеспечивает пищевые и кормовые композиции. 10 н. и 39 з.п. ф-лы, 15 ил., 3 табл.
Область техники, к которой относится изобретение. Настоящее изобретение имеет отношение к биотехнологии растений. В частности, изобретение имеет отношение к способам и конструкциям, предназначенным для повышения содержания отдельных аминокислот в растениях или в тканях и органах растений, включая клеточные стенки, клеточные мембраны, масляные тельца (oil bodies) и, в особенности, семена. Повышение содержания аминокислот достигается путем введения конструкции с рекомбинантной нуклеотидной последовательностью, кодирующей белок-носитель, имеющий на 3′-терминальном конце протяженную полиаминокислотную последовательность. Подробно описаны способ получения и применение композиций, обогащенных вышеупомянутыми аминокислотами; а также применение растений, клеток растений и клеточных линий, трансформированных одной или несколькими указанными конструкциями. Уровень техники. Люди и сельскохозяйственные животные нуждаются в наличии в потребляемой пище восьми незаменимых аминокислот. Диеты, основанные на преимущественном употреблении только крупяных или бобовых продуктов, приводят к дефициту аминокислот, что может привести к негативному влиянию потребляемой пищи на жизнь людей и животных. Например, белки семян зерновых дефицитны по лизину и триптофану, тогда как семена бобовых содержат белки, дефицитные по серосодержащим аминокислотам, метионину и цистеину. Для поддержания животных в здоровом состоянии, для эффективного роста и хорошего качества мяса и молока при использовании белков семян в корме крупного рогатого скота необходимо, чтобы в пище содержались указанные аминокислоты. Таким образом, чтобы лучше адаптировать существующие ресурсы растительных белков к нуждам определенных животных, их выгодно модифицировать, в частности, для создания композиций незаменимых аминокислот. Были предприняты попытки получить композиции растительных аминокислот, соответствующих диетическим потребностям людей и животных, но достигнутые успехи носили ограниченный характер. Использование ценных в пищевом отношении мутантов растений и их тканей, однако, является компромиссным решением из-за наличия негативных плейотропных эффектов. Эти проблемы включают плохое прорастание семян, медленное высыхание, сниженный выход, повышенную чувствительность к микробам и насекомым и низкие мукомольные характеристики. Генная инженерия предоставляет альтернативные возможности изменения состава незаменимых или любых других аминокислот в растениях и тканях растений. Чтобы повысить содержание метионина или лизина, были разработаны подходы как изменения путей биосинтеза, так и экспрессии белков с высоким содержанием метионина/лизина в трансгенных семенах (WO 96/38574; WO 96/01905; WO 95/31554; WO 95/15392; WO 93/19190; ЕР 485970, WO 99/40209). В большинстве случаев использовались три метода: 1) изменение аминокислотной последовательности эндогенного белка (белков); 2) использование природных белков из других растений для гетерологической экспрессии; и 3) экспрессия синтетических генов, содержащих высокий уровень метионина/лизина. Метод использования белков с высоким содержанием метионина/лизина из других растений подробно раскрыт в патентах US 5633436, US 5580782 и WO 94/16078. С целью улучшения питательной ценности важнейших бобовых культур узколистный люпин (Lupinus angustifolius L.) был устойчиво трансформирован химерным геном, который определяет специфичную для семян подсолнечника экспрессию богатого серой альбумина. Ген альбумина семян подсолнечника, белка, который по расчетам составляет 5% от экстрагируемых белков семян, содержался в виде единичной тандемной вставки во вносимой ДНК. Трансгенные семена содержат меньшее количество сульфатов и большее количество общей серы, связанной с аминокислотами, по сравнению с нетрансгенной родительской линией; это было связано с увеличением на 94% содержания метионина и уменьшением на 12% содержания цистеина. При этом в семенах не происходило статистически значимого изменения содержания других аминокислот или общего азота, а также общего содержания серы. Комбинация первого и второго подходов (см. выше) описана в патенте US 5850016. Чтобы повысить содержание метионина в клубнях картофеля, клон кДНК, кодирующий ген 2S альбумина бразильского ореха, был подвергнут мутации для увеличения содержания метионина путем введения в него 2-7 дополнительных остатков метионина, а затем трансформирован в картофельное растение. Независимо от наличия мутации содержание белка в листьях было сниженным и составляло <0,01%-0,2% от общего белка. Использование синтетических белков подробно раскрыто в патентах FR 2744134, US 5559223 и WO 92/14822. Чтобы повысить содержание лизина и метионина в семенах, был сконструирован синтетический белок, в основе которого была двойная В патенте WO 99/15004 описана химерная конструкция для модификации состава запасающих органов растений. Кодируемый химерным геном обогащенный серой белок содержал на С-конце KDEL элемент, который обеспечивал направленное внедрение конструкции в эндоплазматический ретикулум и в аппарат Гольджи. Подходы, упомянутые выше, обладают определенными недостатками. Если в растении-хозяине экспрессируются значительные количества чужеродных белков, не имеющих функционального значения, это может привести ко многим вторичным проблемам, связанным с физиологическими аномалиями семян, как было отмечено для сходного типа мутантов, созданных с помощью традиционных методов селекции. Первым объектом настоящего изобретения является способ, включающий получение конструкции рекомбинантной нуклеотидной последовательности, создающий возможность отбора конструкций для эффективной трансформации любых желаемых видов растений с направленной экспрессией, обеспечивающей аккумуляцию стабильного белка, обогащенного одной или несколькими выбранными аминокислотами, закрепляющей эти белки в любых выбранных тканях растений. При использовании указанных конструкций можно исключить нежелательные эффекты экспрессии значительных количеств чужеродных белков, не имеющих функционального значения в растении-хозяине. Вторым объектом настоящего изобретения являются композиции, включающие стабильный обогащенный аминокислотами белок, который аккумулировался в определенных тканях растений в комбинации с совместимыми вспомогательными добавками. Предполагается применение указанной композиции как непосредственного источника пищи в виде определенных растений для человека, а также в качестве фуража, в особенности для кормления животных, и в качестве пищевых добавок. Сущность изобретения. Настоящее изобретение имеет отношение к улучшению качества растительных белков с помощью способа, который повышает содержание одной или нескольких выбранных аминокислот путем направленной экспрессии или аккумуляции белка, обогащенного аминокислотными последовательностями, удлиняющими с конца цепи этот растительный белок. Метод включает стадии трансформации растения, по меньшей мере, одной конструкцией рекомбинантной нуклеотидной последовательности. Конструкция включает в себя ткане- или органоспецифичные регуляторные последовательности, которые контролируют транскрипцию во время селективных стадий морфогенеза. Регуляторные последовательности присоединены к химерной нуклеотидной последовательности. Химерная нуклеотидная последовательность, которая используется для трансформации растений, в особенности сельскохозяйственных культур, включает два ключевых элемента – нуклеотидную последовательность, кодирующую белок-носитель с нормальными функциональными свойствами нативного белка, и нуклеотидную последовательность, включающую кодоны, кодирующие аминокислотную последовательность, имеющую от четырех до восьми аминокислотных остатков и включающую комбинацию одной или нескольких отобранных аминокислот, которая удлиняет с конца цепи функционально нормальный белок-носитель. Нуклеотидная последовательность, кодирующая белок-носитель, отбирается из нуклеотидных последовательностей, кодирующих специфичные белки растений, таким образом, чтобы она обеспечивала направленную экспрессию или аккумуляцию обогащенного аминокислотами белка в определенных тканях или органах растения. В нуклеотидной последовательности, кодирующей белок-носитель, отсутствует терминирующий кодон, она представляет собой соединение в рамке считывания 3′-терминального конца и нуклеотидной последовательности, включающей определенное число кодонов, кодирующих отобранную комбинацию одного или нескольких аминокислотных остатков. Указанная конструкция позволяет осуществлять стабильную направленную экспрессию или аккумуляцию белка-носителя, обогащенного аминокислотами и имеющего на конце цепи полиаминокислотное удлинение, в определенных тканях и органах растений. Описанная выше так называемая трансформирующая конструкция получена с помощью селекции конструкций, обеспечивающих стабильную трансляцию белка с полиаминокислотным удлинением цепи и его стабильную целевую экспрессию или аккумуляцию. Селекцию выполняют с помощью системы трансляции in vitro (IVT) и/или системы переменной экспрессии (transient expression) и в дальнейшем подтверждают с помощью других применяемых методов. Для определения оптимального количества кодонов, которые могут быть соединены в рамке считывания с нуклеотидной последовательностью, кодирующей белок-носитель, без компрессии эффективности трансляции обогащенного аминокислотами белка-носителя с удлиненной цепью, вначале используется система IVT. Система переменной (неустойчивой) экспрессии включает конструкцию, описанную выше, объединенную в рамке считывания с нуклеотидной последовательностью, кодирующей репортерный белок. Эта конструкция переменной экспрессии вводится в клетку растений, предпочтительно, с помощью метода микропроекционной бомбардировки (microprojective bombardment). После этого отбирают конструкции, которые обеспечивают стабильную направленную экспрессию репортерного гена в клетке указанного растения, и в отобранных конструкциях удаляют нуклеотидные последовательности, кодирующие репортерный белок. Указанные конструкции применяют в производственных целях, трансформируя растения конструкциями, не содержащими репортерный ген, как описано выше, предпочтительно используя системы трансформации, опосредованные Agrobacterium. Способ делает возможным продукцию белков, обогащенных аминокислотами, то есть продукцию белков-носителей с удлиненной на конце полиаминокислотной последовательностью, включающей выбранную комбинацию от четырех до восьмидесяти аминокислотных остатков. Аминокислоты представляют собой главным образом гистидин, цистеин, метионин, глицин, лизин, триптофан, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, тирозин, серии, треонин, аргинин, аспартат, глутамат, аспарагин, глутамин или любые другие их комбинации. Белок-носитель, позволяющий осуществлять стабильную направленную экспрессию или аккумуляцию в тканях или органах растения, может быть выбран из белков, функционирующих во внутриклеточных транспортных путях растения. Чаще всего белок-носитель является белком клеточной стенки или белком вирусов растений. Полезные белки-носители представляют собой, например, олеозин, калеозин, стеролеозин, круциферин, напин или транспортный белок вирусов растений. Регуляторная последовательность представляет собой подходящий промотор, экспрессирующий во время эмбриогенеза. Подходящие регуляторные последовательности включают промоторы напина (NAP), 35S, химерный гибридный промотор (HYB), промоторы 19S, нопалина, фазеолина, стеролеозина, калеозина, круциферина, промотор вируса альфальфа мозаики (AMV), промоторы белков теплового шока, альбумина 2S или олеозина. Репортерный белок представляет собой главным образом детектируемый белок. В особенности выгодными являются флуоресцентные белки, такие как зеленый флуоресцентный белок (GFP) и красный флуоресцентный белок (RFB), а также Способ, предложенный в настоящем изобретении, в особенности полезен для создания композиций, содержащих в растительном материале, в том числе и в обломках клеточной стенки, белок-носитель, обогащенный аминокислотами, имеющий на конце цепи полиаминокислотное удлинение; содержание определенных аминокислот в растительном материале, полученном предложенным способом, по сравнению с содержанием аминокислот в соответствующем не модифицированном растении дикого типа, находится, по меньшей мере, в соотношении 2:1. Указанная композиция применима для получения корма, обогащенного аминокислотами, из жмыха, полученного после извлечения масла из растений. Также в настоящем изобретении подробно раскрыты растения, растительные клетки и растительные клеточные линии, трансформированные с помощью конструкций переменной экспрессии или конструкций для трансформации. Типичные возможности настоящего изобретения описаны в разделе Формула изобретения. Краткое описание фигур. Фигура 1 показывает 18 экспрессирующих плазмид, где pRT (в названии) означает конструкцию, содержащую промотор 35S гена вируса мозаики цветной капусты в плазмидном векторе pRT100 (Topfer et al., Nucleic Acid Res. 15(14):5890, 1987). Восемнадцать плазмид были сконструированы с использованием генов любого из двух белков-носителей – олеозина (OLE) или транспортного белка вируса табачной мозаики (МР) – в сочетании с одним из трех промоторов транскрипции, указанных стрелками (NAP – промотор напина, 35S – промотор 35S гена вируса мозаики цветной капусты, HYB – гибридный промотор, в котором энхансерный участок промотора CAMV 35S соединен с промотором напина), и внутренних участков нуклеотидной последовательности, обогащенных кодонами гистидина (His) или цистеина-метионина (Cys-Met) (так называемые “кассеты”, подробно см. на фигуре 3) различной длины (2х – содержит две кассеты, 4х – содержит четыре кассеты, 6х – содержит шесть кассет), обозначенных серыми клетками. Участок, кодирующий зеленый флуоресцентный белок (GFP), был соединен в рамке считывания с последовательностью, обогащенной кодонами His или Cys-Met. Стрелки обозначают промоторы, небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 2 показывает четыре экспрессирующих плазмиды, которые были отобраны на основе результатов измерения переменной экспрессии при стабильной трансформации в растение Brassica campestris. NAP – промотор напина, HYB – гибридный промотор, в котором энхансерный участок промотора CaMV 35S соединен с промотором напина, OLE – олеозин, МР – ген транспортного белка 30К вируса табачной мозаики, GFP – ген зеленого флуоресцентного белка, 4х – четыре участка нуклеотидных последовательностей, кодирующих зону, обогащенную аминокислотами (кассеты). Стрелки обозначают промоторы, небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 3 показывает последовательность нуклеотидов участка, обогащенного кодонами гистидина (А), участка, обогащенного кодонами цистеина-метионина (В), участка, обогащенного кодонами глицина (С), и участка, обогащенного кодонами лизина (D). Последовательность транслируемого белка показана под нуклеотидной последовательностью. К – лизин; Н – гистидин; R – аргинин; V – валин; G – глицин; L -лейцин; С – цистин; S – серин; М – метионин. Звездочка указывает на кодон, терминирующий транскрипцию. Сайты рестрикции, использованные при клонировании, выделены полужирным шрифтом и указаны над последовательностью. Фигура 4 показывает схему конструирования плазмид, несущих последовательности ДНК различной длины, обогащенные гистидиновыми или цистеин-метиониновыми кодонами, соединенных с геном TMV 30К МР (не заштрихованный блок “МР”). Для создания конструкции нуклеотидной последовательности, обогащенной гистидиновыми кодонами, использовали экспрессирующий вектор pGEM-7Zf(+) (Promega Corporation, USA; номер по каталогу Р2251). Клон pGEM-His-24 содержит сегмент ДНК, кодирующий пептид из 19 аминокислот, в состав которого входит 14 остатков His (фигура 3А). Последовательность этого фрагмента ДНК фланкирована сайтами рестриктаз XhoI и BamHI и содержала также сайт рестрикции Sail, сконструированный для последующих стадий клонирования (фигура 3А). Последовательности ДНК, обогащенные гистидиновыми или цистеин-метиониновыми кодонами, обозначены серыми блоками различной протяженности. pGEM-His-24 является примером соединения последовательности ДНК, обогащенной гистидиновыми кодонами, с геном TMV 30К МР (пример 11). Такая же схема конструирования может быть использована для соединения последовательности ДНК, обогащенной цистеин-метиониновыми кодонами, с геном TMV 30К МР (пример 12). На фигуре указано положение нескольких сайтов рестрикции. Фигура 5А описывает конструирование экспрессирующих плазмид pNAP и pHYB, где RT означает, что конструкция содержит промотор 35S гена вируса мозаики цветной капусты (промотор CaMV 35S) из плазмидного вектора pRT100 (Topfer et al., Nucleic Acid Res. 15 (14): 5890, 1987). Экспрессирующие плазмиды содержат промотор напина (NAP) или химерный промотор (HYB), который состоит из собственно промотора напина, соединенного с энхансерным участком промотора 35S гена вируса мозаики цветной капусты (CaMV) (пример 16). Стрелки обозначают промоторы, а небольшой блок, помеченный значком “t”, означает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 5В описывает клонирование гена зеленого флуоресцентного белка (GFP) в плазмиды на основе промоторов 358 CaMV и напина, где RT означает, что конструкция содержит промотор 35S гена вируса мозаики цветной капусты (промотор CaMV 35S) из плазмидного вектора pRT100 (Topfer et al., Nucleic Acid Res. 15 (14): 5890, 1987). Экспрессирующие плазмиды содержат промотор напина (NAP) или химерный промотор (HYB), который состоит из собственно промотора напина, соединенного с энхансерным участком промотора 35S гена вируса мозаики цветной капусты (CaMV) (пример 16). Стрелки обозначают промоторы, а небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 6 показывает относительную экспрессию GFP, получаемую при использовании промоторов 35S CaMV (а) и HYB (b) в конструкциях pRT-OLE-4x-GFP и pHYB-OLE-4x-GFP, соответственно (см. фигуру 1), после трансформации этими конструкциями эпидермальных клеток Nicotiana benthamiana методом бомбардировки частицами (пример 8). Фигура 7 описывает конструирование экспрессирующих векторов растений, состоящих из гена TMV 30К МР (МР), кассет различной длины с последовательностями, обогащенными кодонами His и Cys-Met (серые клетки), соединенных с участком, кодирующим GFP, под контролем промотора 35S CaMV. pRT означает, что конструкция содержит промотор 35S гена вируса мозаики цветной капусты (промотор CaMV 35S) из плазмидного вектора pRT100 (Topfer et al., Nucleic Acid Res. 15 (14): 5890, 1987). Стрелки обозначают промоторы, а небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 8 описывает конструирование экспрессирующих векторов растений, состоящих из гена TMV ЗОК МР (МР), кассет различной длины с последовательностями, обогащенными кодонами His и Cys-Met (серые клетки), соединенных с участком, кодирующим GFP, под контролем промотора напина (NAP). pRT означает, что конструкция содержит промотор 35S гена вируса мозаики цветной капусты (промотор CaMV 35S) из плазмидного вектора pRT100 (Topfer et al., Nucleic Acid Res. 15 (14): 5890, 1987). PGEM – 30К содержит ген транспортного белка 30К из TMV в плазмиде pGEM-7Z(+)(Promega Corporation, USA) (пример 7). Стрелки обозначают промоторы, а небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 9 описывает конструирование экспрессирующих векторов растений, состоящих из гена TMV 30К МР (МР), кассет различной длины с последовательностями, обогащенными кодонами His и Cys-Met (серые клетки), соединенные с участком, кодирующим GFP, под контролем гибридного промотора (HYB). pRT означает, что конструкция содержит промотор 35S гена вируса мозаики цветной капусты (промотор CaMV 35S) из плазмидного вектора pRT100 (Topfer et ей., Nucleic Acid Res. 15 (14): 5890, 1987). pGEM-30К содержит ген транспортного белка 30К из TMV в плазмиде pGEM-7Z(+) (Promega Corporation, USA) (пример 7). Стрелки обозначают промоторы, а небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 10 описывает конструирование экспрессирующих векторов растений (pRT), состоящих из гена олеозина (OLE), кассет различной длины с последовательностями, обогащенными кодонами His и Cys-Met (серые клетки), соединенных с участком, кодирующим GFP, под контролем промотора 35S CaMV. pRT означает, что конструкция содержит промотор 35S гена вируса мозаики цветной капусты (промотор CaMV 35S) из плазмидного вектора pRT100 (Topfer et al., Nucleic Acid Res. 15 (14): 5890, 1987). Для подробной информации о pOLE4/11 см. пример 6. Стрелки обозначают промоторы, а небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 11 описывает конструирование экспрессирующих векторов растений (pRT), состоящих из гена олеозина (OLE), кассет различной длины с последовательностями, обогащенными кодонами His и Cys-Met (серые клетки), соединенных с участком, кодирующим GFP, под контролем промотора напина (NAP) Для подробной информации о pOLE4/11 см. пример 6. Стрелки обозначают промоторы, а небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 12 описывает конструирование экспрессирующих векторов плазмид (pRT), состоящих из гена олеозина (OLE), кассет различной длины с последовательностями, обогащенными кодонами His и Cys-Met (серые клетки), соединенных с участком, кодирующим GFP, под контролем гибридного промотора (HYB). Для подробной информации о pOLE4/11 см. пример 6. Стрелки обозначают промоторы, а небольшой блок, помеченный значком “t”, показывает последовательность, терминирующую транскрипцию. На фигуре указано положение нескольких сайтов рестрикции. Фигура 13 показывает вестерн-блоттинг продуктов экспрессии pNAP-MP-4xHis-GFP (см. фигуру 2) в растении Brassica campestris. Дорожки 1, 2 и 3 соответствуют трем независимым трансформантам pNAP-MP-4xHis-GFP (линии 5.1 А7, 5.1 А11 и 5.1 А18, соответственно). Маркеры молекулярной массы (точки в правой части) соответствуют 50 кДа (верхняя точка) и 40 кДа (нижняя точка). В качестве индикатора использовали антитела к His. Фигура 14 показывает вестерн-блоттинг продуктов экспрессии pHYB-OLE-4xHis-GFP (см. фигуру 2) в растении Brassica campestris. Дорожки 1, 2, 3, 5, 6 и 7 соответствуют шести независимым трансформантам pHYB-OLE-4xHis-GFP (линии 17.1237, 17.1238, 17.1240, 17.20с8, 17.20с11 и 17.20с20, соответственно); дорожка 4 соответствует контрольному (нетрансформированному) растению дикого типа. Маркеры молекулярной массы (дорожка 8) соответствуют 50 кДа (верх) и 40 кДа (низ). В качестве индикатора использовали антитела к His. Фигура 15 показывает вестерн-блоттинг продуктов экспрессии pNAP-OLE-4xHis-GFP (см. фигуру 2) в растении Brassica campestris. Дорожки 2, 3, 5, 6, 7 и 8 соответствуют шести независимым трансформантам pNAP-OLE-4xHis-GFP; дорожка 4 соответствует контрольному (нетрансформированному) растению дикого типа. Маркеры молекулярной массы (дорожки 1 и 9) соответствуют 50 кДа (верхняя точка) и 40 кДа (нижняя точка). В качестве индикатора использовали антитела к His. Раскрытие изобретения. Разъяснение используемых терминов. Используемые в настоящем изобретении термины имеют тот же самый смысл, что и аналогичные термины в области биотехнологии растений, химии белков и пищевых композиций. Некоторые термины в настоящем изобретении, однако, используются в более широком смысле или их смысл приобретает, до некоторой степени, несколько другое значение. Поэтому ниже некоторые термины определены более детально. “Конструкция рекомбинантной нуклеотидной последовательности” или просто “конструкция” означает конструкцию ДНК, переменно экспрессирующую кассету или вектор, или содержащую кассету или вектор для трансформации. Конструкция может включать линейные или кольцевые соединенные “конец в конец” нуклеотидные последовательности, дополнительно включенные в плазмиду. Для создания эффективных стабильно трансформирующих конструкций, предназначенных для производственных целей, разрабатываются, идентифицируются и отбираются стабильные наиболее удачно выполненные переходные или промежуточные конструкции для экспрессии репортерного белка. Затем, чтобы получить конструкцию для трансформации в определенные виды растений, которые в конечном итоге применяются в производственных целях, из указанных отобранных эффективных конструкций удаляют репортерную последовательность. В настоящем изобретении конструкция для трансформации включает регуляторные последовательности и нуклеотидную последовательность, кодирующую белок-носитель, соединенную (слитую, fused) в рамке считывания с кодонами выбранных аминокислотных остатков. В переходную конструкцию затем включают нуклеотидную последовательность, кодирующую детектируемый репортерный белок. В переходную или промежуточную конструкцию включена, по меньшей мере, одна кассета нуклеотидной последовательности, но более предпочтительно введение большего количества кассет, например, от двух до шести. Каждая кассета включает, по меньшей мере, два, предпочтительно пять и более кодонов, кодирующих выбранные аминокислотные остатки. Кассеты, включающие до десяти или пятнадцати кодонов выбранных аминокислот, могут быть вставлены между нуклеотидными последовательностями, кодирующими белок-носитель и репортерный белок. Следует отметить, что нуклеотидные последовательности, кодирующие репортерный белок, удаляют из тех конструкций, для которых было показано, что они стабильны и эффективны, то есть они работают так, как этого хотелось, демонстрируя направленную экспрессию. Эти “вставляемые” (inserted) конструкции с отсутствующим репортерным геном используют как трансформирующие конструкции. Обычно количество аминокислотных кодонов в стабильной конструкции составляет от четырех до восьмидесяти. “Кассета нуклеотидных последовательностей” или “кассета нуклеотидной последовательности” включает набор кодонов, которые кодируют последовательность аминокислот, так называемую “аминокислотную кассету”. В настоящем изобретении кассета означает вставку, включающую непрерывную нуклеотидную последовательность, имеющую, по меньшей мере, два, предпочтительно четыре или любое другое количество (до восьми) кодонов или триплетов. Кассеты особенно удобны при создании различных конструкций и для проверки их свойств, в особенности их участия в трансляции. Однако кассеты не являются необходимым условием настоящего изобретения. Конечной целью является предоставление конструкции, которая кодирует белок-носитель, имеющий полиаминокислотное удлинение на конце цепи, но все еще обладает теми же функционально интактными (нормальными, неповрежденными) свойствами, что и соответствующий нативный белок-носитель. Предпочтительно, чтобы оптимальное количество кодонов приводило к образованию стабильного белка-носителя, удлиненного с конца цепи полиаминокислотной последовательностью и, таким образом, обогащенного определенными аминокислотными остатками. Обычно количество аминокислотных остатков, которые можно стабильно присоединить для удлинения цепи, составляет от четырех до восьмидесяти аминокислотных остатков или любое другое промежуточное число. Число аминокислотных остатков, которые могут быть присоединены к белку-носителю, можно определить случайным образом после включения в конструкцию кассет с нуклеотидными последовательностями, содержащими различные количества аминокислотных кодонов, с последующим скринингом конструкций в бесклеточной системе трансляции для подтверждения правильности трансляции кодона. Отбор “стабильных конструкций” удобно осуществлять с помощью “бесклеточной системы трансляции”, которая представляет собой систему трансляции in vitro (IVT), в которой нормальная клеточная реакция воссоздается в отсутствие клеток и которая включает, например, системы IVT, способные синтезировать белок с мРНК, используя, например, лизат ретикулоцитов кролика или зародышей пшеницы. Особенно удобна бесклеточная система трансляции из зародышей пшеницы, выпускаемая, например, фирмой Promega (TNT®T7/SP6 Coupled Wheat Germ Extract System L5030, Promega corporation). “Направленная экспрессия или аккумуляция” означает, что белок, обогащенный аминокислотами, экспрессируется в специфических, селективных тканях или органах растения или транспортируется в указанные ткани или органы. Это может быть достигнуто с помощью селекции белков-носителей, которые принимают участие во “внутриклеточных транспортных путях”. Такие пути представляют собой системы транспорта, специфичные к компоненту мембраны, к органелле и т.д. Экспрессированный белок переносится по внутриклеточным транспортным путям растений в определенные органы и ткани, которые таким образом способны аккумулировать экспрессированный продукт, например, в клеточных стенках или клеточных мембранах семян. Если экспрессия белков-носителей, обогащенных аминокислотами, в семенах трансгенных видов Brassica нацелена на клеточные стенки семян, то это делает возможным закрепление белка, обогащенного аминокислотами, в жмыхе, остающемся после отжима масла. Жмых, наделенный улучшенными свойствами, т.е. повышенным содержанием аминокислот, является полезным ингредиентом в композициях для пищевой промышленности. Таким же образом белок, обогащенный аминокислотами, может быть экспрессирован в мезофильной ткани салата-латука и использоваться непосредственно в качестве источника питания для человека. “Выбранная (отобранная) аминокислота” может быть любой аминокислотой, но предпочтительней всего она представляет собой одну или более из восьми незаменимых аминокислот, необходимых человеку или крупному рогатому скоту. Выбранной аминокислотой может быть, например, гистидин, цистеин, метионин, глицин, триптофан, лизин, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, тирозин, серин, треонин, аргинин, аспартат, глутамат, аспарагин или глутамин. Любой белок, обогащенный аминокислотами, может быть приготовлен с помощью способа настоящего изобретения, в частности, таким образом, как это проиллюстрировано на примере получения белка, обогащенного гистидином. Белок, обогащенный аминокислотами, получают с помощью конструкции, включающей нуклеотидную последовательность, кодирующую белок-носитель, удлиненную на оптимальное количество кассет, включающих один и тот же или разные кодоны, кодирующих желаемые аминокислоты, например, His, Met-Cys, His-Met-Cys и т.д. В предпочтительной промежуточной или переходной конструкции настоящего изобретения аминокислотные кодоны расположены между нуклеотидной последовательностью, кодирующей носитель, и репортерным белком таким образом, что дополнительные аминокислотные остатки не мешают нормальным биологическим функциям белка-носителя, не препятствуя его участию в секреторных внутриклеточных транспортных путях. “Белок-носитель” в настоящем изобретении означает белок, который может быть стабильно удлинен с помощью полиаминокислотной последовательности или пептида. Нуклеотидная последовательность, кодирующая функционально интактный белок-носитель, соединена в рамке считывания с нуклеотидной кассетой, включающей один или более выбранных кодонов, кодирующих аминокислотные остатки. Вставленные кодоны, кодирующие желаемые аминокислотные остатки, должны не мешать нормальным биологическим функциям белка-носителя при сравнении их с функциями соответствующего нативного немодифицированного белка. Нуклеотидные последовательности, кодирующие белки-носители, применяемые в настоящем изобретении, выбраны из специфических белков растений. Эти специфические белки растений используют секреторные внутриклеточные транспортные пути, что делает возможным аккумуляцию белка, обогащенного аминокислотами, в клеточных стенках или мембранах. Особенно пригодными являются мембраны масляных телец, плазматические мембраны, вакуолярные мембраны, мембраны пластид определенных тканей и органов растений, включая семена, листья, корни, клубни и т.д. В настоящем изобретении белки-носители могут происходить из генов, кодирующих три основных белка семян. Указанные гены представляют собой полезные модельные белки для выполнения генно-инженерных манипуляций с белками семян, в особенности с белками семян растений семейства Criciferae. Кроме этого, похожие генно-инженерные подходы можно применять для любых других видов растений. Указанные выше в качестве примера три белка семян, которые используют в настоящем изобретении, являются круциферином (500 аминокислотных остатков), напином (165 аминокислотных остатков) и олеозином (165 аминокислотных остатков), но также можно использовать и другие белки, такие как калеозин и стеролеозин. Все упомянутые выше белки присутствуют в нативных семенах В. campestris, и содержание гистидина в них снижено. Круциферин содержит только 9 остатков His на 500 аминокислот. Напин содержит только 2 остатка His на 165 аминокислот, и олеозин вовсе не содержит His. Круциферин, олеозин, напин, калеозин и стеролеозин подходят для использования в семенах в прикладных задачах пищевой отрасли. В настоящем изобретении олеозин (OLE) использовали в качестве модельного белка-носителя, но таким же образом могут быть использованы и другие белки-носители. Например, предполагается, что два родственных белка, калеозин и стеролеозин, упомянутые выше, являются потенциально пригодными для этих целей, так как они аккумулируются сходным образом в масляных тельцах и клеточной стенке семян. Свойства олеозина рассмотрены в обзорах, например, в (i) Murphy 1996. TIBTECH 14. 206-213; (ii) Methods in Mol. Biol. vol.44: Agrobacterium protocol. Eds. K.M.A.Gartland и (iii) M.R.Daey, Humamana Press Inc. Totowa, N.J.; and Brassica Oilseeds: Production and Utilization. Eds D.S.Kimber and D.I. Mc.Gregor. Cab International. 1995. Показано, что олеозин является пригодным белком-носителем, так как это гидрофобный белок, имеющий относительно небольшие размеры, а также он является компонентом мембран, окружающих тельца, которые запасают масла в семенах В. campestris. При переносе нуклеотидной последовательности, извлеченной из одного выбранного растения, в другое выбранное растение существует потенциальный риск посттранскрипционного выключения гена, в особенности в том случае, если растения близкородственны или высоко гомологичны. Риск индукции посттранкрипционного выключения гена в трансгенном растении, возникающий из-за гомологии нуклеотидной последовательности между переносимым геном и эндогенным геном, можно избежать при использовании нуклеотидных последовательностей, кодирующих белки-носители, обладающих по возможности слабой гомологией. Например, чтобы избежать этой проблемы, при трансформации В. capestris в настоящем изобретении был успешно использован белок-носитель (кодируемый геном OLE) из Arabidopsis thaliana, но не из В. capestris. Преимущество использования гена, кодирующего транспортный белок (МР) состоит в том, что этот ген не имеет гомологии в своей последовательности с эндогенным белком из В. capestris. “Белки-носители” в настоящем изобретении включают изомеры, аминокислотные последовательности с минорными модификациями в некоторых аминокислотных остатках. Белок-носитель может быть более короткой формой нативного белка-носителя. Нуклеотидные последовательности, кодирующие белок-носитель, могут также до известной степени различаться. Они могут быть, например, укороченными. Обязательным условием является то, что ген, кодирующий белок-носитель, должен обладать интактными функциональными свойствами или биологическими функциями, которые в значительной степени должны быть такими же, как у нативного белка-носителя, включая и направленную экспрессию белка в определенных тканях и органах растения. “Интактные функциональные свойства” означают, что экспрессия указанного белка-носителя может быть направлена на определенные ткань, орган или компартмент растения. Другими словами, белок-носитель с удлиненной аминокислотной последовательностью должен аккумулироваться в выбранной ткани, органе или компартменте растения. “Регуляторная последовательность” означает нуклеотидные последовательности, которые регулируют транскрипцию и экспрессию структурных нуклеотидных последовательностей с помощью механизмов понижения и повышения уровня транскрипции и экспрессии. Регуляторные последовательности включают в себя промоторы, энхансеры, сигнальные последовательности, терминаторы и т.д. Наиболее предпочтительными являются относительно короткие, органоспецифичные или тканеспецифичные транскрипционные промоторы, способные контролировать транскрипцию химерных нуклеотидных последовательностей во время различных стадий морфогенеза и, в особенности, во время эмбриогенеза. Другие потенциально пригодные промоторы представляют собой промоторы, происходящие из вирусов растений: вируса альфальфа мозаики (AMV) и вируса мозаики цветной капусты (CaMV) 19S, которые могут контролироваться как с помощью внешних условий (например, нагреванием), так и с помощью специфических абиотических (например, в ответ на обработку салициловой кислотой) или биотических (патогены) воздействий. Наиболее предпочтительными и показательными примерами “транскрипционных промоторов” в настоящем изобретении являются промотор напина (NAP), в особенности промотор из Arabidopsis thalwna, промотор 35S CaMV и химерный “гибридный” (HYB) промотор, который включает собственно промотор напина, соединенный с энхансерной последовательностью промотора CaMV 35S. Как и следовало ожидать, промоторы могут включать любые другие работающие регуляторные последовательности, в особенности промоторы, способные осуществлять аккумуляцию белков, обогащенных аминокислотами, в выбранных тканях. Предпочтительными тканями являются масляные тельца и клеточные стенки семян. Промотор напина, контролирующий экспрессию гена напина (NAP), регулируется во время эмбриогенеза и включается после цветения. Опубликована нуклеотидная последовательность промотора NAP из A. thaliana (Rask et al. 1998, J. Plant Physiol. 152, 595-599). Важным преимуществом промотора напина (NAP) в настоящем изобретении является то, что он имеет относительно небольшие размеры (152 п.н.) по сравнению с промотором гена 01е, который составляет 2,1 тыс. п.н. Показано, что промотор 35S гена вируса мозаики цветной капусты (CaMV) активен в клетках В. campestris (Harper et al., 1988. Mol. Gen. Genet. 212, 182-190). Промотор 35S также активен в зародышах, и выявление трансгенных растений В. campestris должно облегчаться за счет определения продукта экспрессии, рекомбинантного белка, обогащенного аминокислотами (OLE или МР), в зрелых листьях перед цветением. Под “репортерной последовательностью” или “нуклеотидной последовательностью, кодирующей репортерный белок”, имеют в виду нуклеотидную последовательность, используемую при создании промежуточной или переходной конструкции, которая делает возможной селекцию такой конструкции, которая позволяет осуществлять стабильную направленную экспрессию или аккумуляцию стабильных белков, обогащенных аминокислотами, с нормальными биологическими функциями в селективных тканях или органах растений. В этом случае нуклеотидная последовательность, кодирующая репортерный белок, делает возможньм простую, точную и однозначную идентификацию подходящей конструкции. Репортерная последовательность позволяет продемонстрировать, в каких тканях и органах активируется промотор и в каких условиях промотор активен. В наиболее предпочтительных воплощениях настоящего изобретения репортерный белок представляет собой видимый или детектируемый белок, желательно флуоресцентный белок. Предпочтительно это зеленый флуоресцентный белок, Чтобы ускорить селекцию пригодных стабильных конструкций, в настоящем изобретении были использованы две различные системы. Для селекции подходящих трансформирующих конструкций, позволяющих осуществлять нормальную экспрессию белка-носителя и аминокислотной кассеты, использовали бесклеточную систему трансляции in vitro (IVT) и систему переменной экспрессии. Бесклеточная система трансляции in vitro дает возможность идентифицировать и, таким образом, отбирать оптимальное количество кодонов, которые могли бы транслироваться правильным образом без каких бы то ни было проблем. Чтобы убедиться в том, что белок экспрессировался и обладал интактной биологической функцией (функциями) по сравнению с нативным немодифицированным белком, а также в том, что белок экспрессировался и накапливался в намеченной для этих целей ткани и органе растения, был использован анализ переменной экспрессии. Это облегчалось с помощью визуального контроля за репортерным белком. Особенно полезной для этих целей оказалась конфокальная лазерная сканирующая микроскопия, так как этот метод позволял осуществить раннее детектирование в трансгенных растениях экспрессии слитых в рамке считывания нуклеотидной последовательности, кодирующей белки, обогащенные аминокислотами, и нуклеотидной последовательности, кодирующей репортерный белок. В качестве альтернативной системы выявления в трансгенных растениях экспрессии слитых в рамке считывания нуклеотидной последовательности, кодирующей белки, обогащенные аминокислотами, и нуклеотидной последовательности, кодирующей репортерный белок, можно было использовать анализ, основанный на применении антител (например, иммуноферментный анализ), и прямое определение аминокислот. Осуществление изобретения. Основной целью настоящего изобретения была детальная разработка способов, позволяющих увеличивать содержание определенных аминокислот в тканях растений, таких как масляные тельца и клеточные стенки, в особенности, в семенах растений семейства Cruciferae. С помощью способа, представленного в настоящем изобретении, в процессе эмбриогенеза трансгенных растений Brassica campestris был успешно экспрессирован стабильный белок, обогащенный аминокислотами, и экспрессирующийся белок, обогащенный аминокислотами, накапливался в семенах. С помощью этого способа и с помощью конструкций, предоставленных в настоящем изобретении, указанный рекомбинантный белок, обогащенный аминокислотами, оказался прочно связанным с мембранами или клеточными стенками (CW) клеток семян, и, таким образом, этот белок мог аккумулироваться в остатках семян, оставаясь в жмыхе после отжима масла. При дальнейшей экстракции это позволило дополнительно извлекать масло и использовать остатки, т.е. жмых, который с его улучшенными свойствами, т.е. с увеличенным содержанием аминокислот, становится пригоден для производства корма животных в качестве полезного ингредиента композиции, обогащенного аминокислотами. Если в корме крупного рогатого скота присутствует дополнительная аминокислота, например гистидин, и если она связана с нормальными нативными белками, как в случае настоящего изобретения, она выполняет функцию природного источника аминокислот и может нормально утилизироваться в реакциях обмена веществ крупного рогатого скота. Таким образом, белки, обогащенные аминокислотами, например белки, обогащенные гистидином, являются полезным ингредиентом в корме крупного рогатого скота, выполняющим функцию усилителя производства молока. Обогащение другими полезными аминокислотами, например, в съедобных листьях салата-латука, сделает возможным применение такого белка (белков), обогащенного аминокислотами, в качестве источника пищи для человека без дальнейшего обработки. Нуклеотидные последовательности, кодирующие репортерные белки, показанные на примере визуально детектируемого зеленого флуоресцентного белка (GFP), расположенные за последовательностью, обогащенной кодонами выбранных желаемых аминокислот, создают возможность для легкого обнаружения экспрессии гена и внутриклеточной локализации экспрессированных белков в трансгенных растениях. Экспрессия репортерного белка из различных конструкций, слитых с геном репортерного белка, показана на примере с GFP и проиллюстрирована на фигуре 1, а также продемонстрирована на фигуре 6, показывающей переходную экспрессию конструкции, слитой с выбранной репортерной меткой, в единичных эпидермальных клетках табака. Таким образом, настоящее изобретение относится к способам получения трансформированных растений, растительных клеток или клеточных линий, которые способны экспрессировать высокий уровень обогащенных аминокислотами белков-носителей, имеющих стабильное полиаминокислотное удлинение цепи, которые, в частности, локализованы в определенных тканях или органах растений, или в компартментах клеток растений, включая мембраны масляных телец или клеточные стенки семян. Способ, предложенный в настоящем изобретении, включает трансформацию растений или клеток растений конструкциями, способными осуществлять экспрессию белков-носителей, удлиненных с помощью полиаминокислотной последовательности, в намеченной ткани или органе растения. Конструкция для стабильной трансформации включает регуляторные последовательности, содержащие органо- и/или тканеспецифичные транскрипционные промоторы, контролирующие транскрипцию интересующего гена (генов), кодирующего белки-носители, во время различных стадий морфогенеза, в особенности во время эмбриогенеза. Указанные регуляторные последовательности эффективно присоединены к: (а) нуклеотидной последовательности, кодирующей белок-носитель и не содержащей терминирующего кодона; и к (б) нуклеотидной последовательности, включающей, по меньшей мере, одну кассету, содержащую, по меньшей мере, два кодона, кодирующих желаемые аминокислотные остатки, объединенные в рамку считывания с нуклеотидной последовательностью, кодирующей белок-носитель. Различные конструкции можно сделать с или без репортерного гена. Конструкции, кодирующие аминокислоты под контролем регуляторных последовательностей, включающих выбранные промоторы, можно проанализировать, используя системы трансляции in vitro (IVT), которые позволяют наблюдать за стабильностью плазмид, имеющих в своем составе различные аминокислотные кассеты. Репортерные конструкции могут быть также проанализированы с использованием переходных измерений (transient assays). Стабильные конструкции, включающие нуклеотидные последовательности, кодирующие белок-носитель, удлиненные с 3′-конца аминокислотными кассетами, были отобраны с использованием системы IVT путем тестирования конструкций с выбранными случайным образом аминокислотными кодонами, например, по меньшей мере, с двумя аминокислотными кодонами, включенными, по меньшей мере, в одну кассету, но предпочтительней было использование большего количества кассет, например, двух, четырех, шести или восьми. Результаты были проанализированы. Если кассета, например, содержит 14 аминокислотных кодонов (14х), это приводит к тому, что белок-носитель содержал 28 (2х), 56 (4х) и 112 (8х) аминокислотных кодонов, расположенных за геном, кодирующим функционально интактный белок-носитель (01е или МР). Предварительные тесты указывали на то, что 8х-клоны (112 аминокислот) были нестабильно продуцируемыми делеционными вариантами (мини-плазмидами). Некоторые из 6х-клонов (84 аминокислоты) также были нестабильны. Таким образом, для дальнейшей работы клоны 2х, 4х и 6х были слиты с 3′-проксимальным репортерным геном, таким как ген GFP с 3′-терминирующими последовательностями. Точно таким же путем с помощью вставки различного количества аминокислотных кодонов в конструкцию, включающую нуклеотидную последовательность, кодирующую белок-носитель, можно определить оптимальное количество любых аминокислотных кодонов и отобрать конструкции, обеспечивающие получение стабильных результатов. Анализ с помощью вестерн-блоттинга нескольких линий растений (таблица 1) показал, что для обнаружения белков с известными молекулярньми массами (MWs) могут быть использованы антитела ко всем видам белков-носителей, аминокислот и репортерных белков, проиллюстрированных на примере гистидиновых последовательностей, МР, олеозина или GFP (фигуры 13-15). Предсказанные молекулярные массы олеозина, МР, GFP и 4xH18-кассеты (с 14 His-кодонами в каждой кассете) составляют 18,5, 30, 30 и 2,5 кДа, соответственно. Из этих значений можно рассчитать соответствующие молекулярные массы слитых белков как общую сумму молекулярных масс каждого из слитых компонентов. При анализе методом гель-электрофореза в ПААГ в присутствии додецилсульфата натрия (SDS) и методом вестерн-блоттинга было показано, что эти слитые белки идентичны по размерам их предсказанным молекулярным массам. Стабильность трансгенно-кодируемых белковых продуктов (олеозин-His и МР-His) была исследована методом вестерн-блоттинга. Анализ (самоопыляемого) потомства растений (3-e поколение) после трансформации (таблица 1) ясно показал, что трансгенная экспрессия оставалась стабильной и в последующих генерациях. Более того, вестерн-блоттинг анализ тех трансформированных растений, которые исследовали и в дальнейшем, выявил, что размер белкового продукта оставался постоянным независимо от поколения растения. Эти исследования также показали, что по отношению к другим растительным белкам количество слитого белкового продукта было относительно постоянным. Трансформированные растения можно проанализировать, используя антитела к белку-носителю, репортерным белкам или аминокислотам, соответственно, любые из них применимы для получения желаемого результата. Некоторые семена можно непосредственно проанализировать на содержание в них выбранных аминокислот. Обычно последовательность событий такова, что сначала анализировали результаты предварительной серии экспериментов, а затем для использования в последующих стадиях отбирали только те конструкции, которые оказались функциональными и были пригодными для дальнейшей работы. Таким образом, перед заключительной трансформацией растений, пригодных для производственных целей, для получения желаемых результатов были проанализированы и проконтролированы многие аспекты используемых методов. С помощью способа, описанного здесь, получали конструкции с желаемыми свойствами, к которым относятся стабильная трансформация растений и экспрессия стабильного белка, обогащенного аминокислотами. Репортерный белок делает возможным простую и точную селекцию конструкций в бесклеточной системе трансляции in vitro (IVT), которые обеспечивают надежную экспрессию стабильных белков-носителей, обогащенных аминокислотами, и содержат детектируемый репортерный белок, слитый с переходной экспрессируемой системой. Указанная выше переходная экспрессируемая система делает возможной проверку того, что биологическая функция (функции) белка-носителя не нарушена. В случае олеозина под интактной биологической функцией подразумевают то, что репортерный белок экспрессируется в масляных тельцах семян, в частности, в мембранах и клеточных стенках клеток семян. Были отобраны те конструкции, которые демонстрируют нормальную биологическую функцию (функции) белка-носителя, что следует из визуального наблюдения за репортерным белком, а затем нуклеотидная последовательность, кодирующая репортерный белок, была из них удалена. Таким образом, нуклеотидная последовательность, кодирующая белок-носитель, и кодоны, кодирующие желаемые аминокислоты, оказываются слитыми в рамке считывания. Удовлетворительные конструкции были отобраны и перенесены в Agrobacterium, и с помощью соответствующих подходящих способов, например с помощью дот-блот анализа, блот-гибридизации по Саузерну, были отобраны положительные (содержащие конструкцию) клоны, которые использовали для трансформации растений, предпочтительно пищевых растений, в частности, пищевых растений семейства Cruciferae. Трансформированные пищевые растения экспрессировали стабильные белки, обогащенные аминокислотами, и аккумулировали указанные белки в намеченной для этого выбранной ткани (тканях) растений. Среди регуляторных конструкций, включающих ткане- или органоспецифичные транскрипционные промоторы, которые способны осуществлять направленный синтез мРНК в листьях, семенах или других желаемых или выбранных органах растения и которые, таким образом, обеспечивают направленную аккумуляцию белка (белков), обогащенного аминокислотами, в этих органах были отобраны регуляторные последовательности, включающие транскрипционные промоторы. Чтобы обеспечить возможность селекции удовлетворительных конструкций, последовательность, обогащенная аминокислотными кодонами, кодирующими выбранные, желаемые аминокислотные остатки, помещают в рамку считывания между геном белка-носителя и нуклеотидной последовательностью, кодирующей детектируемый репортерный белок, предпочтительно это флуоресцентный репортерный белок, например, зеленый флуоресцентный белок (GFP), Предпочтительные пищевые растения настоящего изобретения принадлежат к семейству Brassicaceae, в котором белки, обогащенные аминокислотами, в частности белки, обогащенные гистидином, способные после экспрессии закрепляться и располагаться в мембранах мясляных телец, что обеспечивает их целевую аккумуляцию. Основываясь на тех же самых принципах, можно получить другие конструкции, обеспечивающие возможность аккумуляции белков, обогащенных любыми другими аминокислотами, в любых других желаемых видах растений и их тканях. Указанные выше цели достигаются с помощью растений, трансформированных конструкциями, включающими нуклеотидные последовательности, кодирующие белки-носители, выбранные среди растительных белков относительно небольшого размера, функционирующих в селективных секреторных внутриклеточных транспортных путях, что дает им возможность аккумулироваться в определенных органах-мишенях растений, например, в мембранах листьев, мембранах семян, клеточных стенках клеток листьев или семян. В настоящем изобретении это проиллюстрировано на примере двух различных белков-носителей, белка олеозина и белка TMV МР, полиаминокислотное удлинение цепи которых слито в рамке считывания с указанным нативным белком-носителем; эти белки затем аккумулируются в определенных тканях трансгенных растений. В частности, предпочтительные нуклеотидные последовательности, кодирующие белки-носители, представляют собой олеозины, способные к аккумуляции в мембранах клеток листьев или семян, и транспортные белки, такие как TMV МР, которые могут аккумулироваться в клеточной стенке листьев или семян, если транскрипция контролируется соответствующим промотором. Нуклеотидные последовательности, кодирующие белки-носители, чаще всего получают из тех видов растений, которые не являются близкородственными для трансформируемого растения-хозяина. Нуклеотидная последовательность, кодирующая одну или более аминокислот, представлена в виде одной или более кассет, которые несут кодоны выбранных аминокислот и которые кодируют выбранный пептид, удлиняющий молекулу белка-носителя. Кассета, включающая нуклеотидные последовательности, кодирующие аминокислотные остатки, предпочтительно расположена таким образом, чтобы не нарушать нормальную биологическую функцию (функции) белка-носителя, т.е. на 3′-терминальном конце последовательности, кодирующей белок-носитель. Если дополнительные аминокислотные остатки помещают на N-терминальном конце или где-нибудь в середине белка-носителя, то такой белок, обогащенный аминокислотами, не может аккумулироваться в намеченном органе, как бы этого не хотелось. Нуклеотидная последовательность, кодирующая репортерный белок, объединена в той же самой рамке трансляции с удлиненной нуклеотидной последовательностью, обогащенной кодонами выбранных аминокислот, но не содержащей терминирующий кодон. Оптимальное количество аминокислотных кодонов составляет приблизительно от десяти до восьмидесяти и определяется после проверки правильности трансляции белка в бесклеточной системе трансляции. Таким образом, в одном из воплощений настоящего изобретения, проилюстрированном в качестве примера, нуклеотидная последовательность, кодирующая белок-носитель, расположена на 5′-проксимальном конце нуклеотидной последовательности, а репортерная нуклеотидная последовательность расположена на 3′-терминальном конце. Предпочтительными белками-носителями или интересующими нуклеотидными последовательностями в настоящем изобретении являются OLE или TMV 30КМР. OLE, использованный в примерах настоящего изобретения, представляет собой нуклеотидную последовательность, соответствующую гену хромосомы Arabidopsis thaliana, кодирующему белок семян олеозин (01е), который является компонентом мембраны масляных телец у представителей семейства Criciferae. TMV 30К МР является нуклеотидной последовательностью, происходящей из геномной РНК вируса табачной мозаики U1 (TMV U1). Ген кодирует неструктурный гидрофобный белок 30К, ответственный в инфицированных растениях за перенос вирусного генома из клетки в клетку (транспортный белок, МР, movement protein) через плазмодесму (PD). 30К МР нацелен и аккумулируется в клеточных стенках, а также в PD и экспрессируется в растениях, трансгенных по МР гену. Оптимальное количество аминокислотных кодонов определяют случайным образом, путем трансформации в растения конструкций с нуклеотидной последовательностью, кодирующей белок-носитель с удлинением цепи различного размера, включающим различное число аминокислотных кодонов. С этой целью отбирают трансгенные растения, экспрессирующие белки с ненарушенными по сравнению с нативным немодифицированным белком биологическими функциями, и оценивают их, используя методы анализа переходной экспрессии. Функциональные конструкции и успешные трансформации в трансгенных растениях можно легко продемонстрировать с помощью экспрессии и внутриклеточной локализации экспрессирующихся белков, в частности, репортерного белка. В настоящем изобретении используют методы генной инженерии, которые позволяют осуществить быструю идентификацию и селекцию химерных конструкций, имеющих оптимальное содержание кодонов, делающих возможной стабильную аккумуляцию белка, обогащенного аминокислотами, в намеченных тканях растений. В настоящем изобретении способ продукции в трансгенных растениях белков, обогащенных аминокислотами, включает приготовление одной или более конструкций, описанных выше. Предпочтительный способ селекции включает экспрессию флуоресцентного рекомбинантного белка-носителя, обогащенного аминокислотами, из конструкций, таких как Ole-полиаминокислоты-ОРР или TMV МР-полиаминокислоты-GFP, в бесклеточной системе трансляции. Бесклеточная система трансляции, использованная в этом изобретении, представляет собой способ, который однозначным образом идентифицирует правильные конструкции и экспрессию рекомбинантных белков, обогащенных аминокислотами. В дополнение к традиционным методам детекции можно выявлять флуоресцентный белок (предоставленный в виде GFP) или другие репортерные белки, используя конфокальную лазерную сканирующую микроскопию семян и/или листьев. Использование конфокальной лазерной сканирующей микроскопии обеспечивает удобный способ ранней детекции и селекции трансгенных растений, экспрессирующих вместе с репортерным белком белки, обогащенные аминокислотами. Причиной использования вставки с репортерной нуклеотидной последовательностью являлось то, что такой белок можно использовать в качестве маркера для подтверждения того, что конструкции и экспрессия из рекомбинантных последовательностей белка-носителя, обогащенного аминокислотами, например, олеозин-полиаминокислоты-GFP или TMV МР-полиаминокислоты-GFP, были правильными. За флуоресценцией белка, в частности GFP, можно следить in vivo с помощью конфокальной лазерной сканирующей микроскопии, позволяющей детектировать экспрессию олеозин-полиаминокислоты или TMV МР-полиаминокислоты, а также определяя уровень их экспрессии. Экспрессирующие векторы растений включают нуклеотидные последовательности, кодирующие белки-носители, такие как Ole и TMV MP, слитые с последовательностями различной длины, кодирующими полиаминокислоты, и несущие репортерный ген, например, ген GFP, расположенный на 3′-терминальном конце. Для конструирования экспрессионных векторов растений эти конструкции были соединены с последовательностями различной длины, обогащенными кодонами желаемых аминокислот, например, для гистидина – кодонами САС и CAU, для цистеина и метионина – кодонами TGT, TGC и ATG, для глицина – кодонами GGA, GGT, GGC и GGG и для лизина – кодонами ААА и AAG. Эти конструкции не содержали терминирующий кодон. Стабильные конструкции можно отбирать, тестируя конструкции со случайно выбранным числом аминокислотных кодонов, например, по меньшей мере, с двумя кодонами аминокислот, включенными, по меньшей мере, в одну кассету, но более предпочтительным является большее количество кассет, например, две, четыре, шесть или восемь кассет; с последующим анализом полученных результатов. Если кассета содержит, например, 14 кодонов гистидина (14х), это приводит к тому, что белок-носитель содержит 28 (2х), 56 (4х) и 112 (8х) кодонов гистидина, расположенных за геном, кодирующим функционально интактный белок-носитель (Ole или MP). Предварительные тесты показали, что кассеты 8х (112 аминокислот) были нестабильно продуцируемыми делеционными вариантами (мини-плазмидами). Некоторые из 6х-клонов (84 аминокислоты) также были нестабильны. Таким образом, для дальнейшей работы клоны 2х, 4х и 6х были слиты с 3′-проксимальным репортерным геном GFP с 3′-терминирующими последовательностями. Точно таким же путем можно определить оптимальное количество любых аминокислотных кодонов, вставляя различное количество аминокислотных кодонов в конструкцию, включающую нуклеотидную последовательность, кодирующую белок-носитель, и затем отобрать конструкции, обеспечивающие получение стабильных результатов. Конструкции, такие как изображены на фигуре 1, включающие регуляторные последовательности, в частности такие промоторы, как промотор напина NAP, промотор 35S CaMV и гибридный промотор HYB, были успешно присоединены к белку-носителю, в частности к Ole и/или TMV МР. Все конструкции включали нуклеотидную последовательность, кодирующую репортерный белок; в качестве маркера для селекции особенно предпочтительным является зеленый флуоресцентный белок (GFP). Ген GFP был слит в рамке считывания с аминокислотным “хвостом”, который имел различную длину и который включал одну или более кассет, состоящих, по меньшей мере, из двух аминокислотных кодонов, кодирующих желаемые аминокислоты. В частности, было использовано кратное увеличение числа кассет (2х, 4х, 6х). Для идентификации и отбора наиболее функциональных конструкций в предварительных экспериментах измеряли флуоресценцию репортерного белка, анализируя переходную экспрессию. Анализ переходной экспрессии включал микропроекционную бомбардировку зародышей пищевых растений, как это показано в таблице 1 на примере экспериментов, включающих бомбардировку частицами зародышей В. campestris и листьев Nicotiana benthamiana. Измерения показали, что промотор NAP был активен в зародышах, тогда как промотор HYB был эффективен как в зародышах, так и клетках эпидермиса листьев. Эти данные сформировали основу для селекции четырех конструкций (из указанных доступных восемнадцати конструкций, фигура 1) для последующей работы по трансформации (фигура 2). Предпочтительной системой трансформации была трансформация, опосредованная агробактериями. Каждая генная конструкция, описанная выше, была перенесена в Agrobacterium tumefaciens; с помощью блот-гибридизации по Саузерну были отобраны трансформированные клоны. Каждый из трансформированных штаммов Agrobacterium затем был использован для трансформации семян весеннего рапса (В. campestris). Последующую селекцию трансгенных растений, экспрессирующих белки-носители, обогащенные аминокислотами (Ole-полиаминокислоты или МР-полиаминокислоты), осуществляли с помощью: (а) метода ПЦР, с использованием праймеров, специфичных к химерным генам Ole и TMV МР, содержащим кодоны селективных аминокислот; (б) метода вестерн-блоттинга с антителами к олеозину и 30К TMV МР; (в) измерения флуоресценции GFP в листьях, цветах, зародышах. Если при использовании методов, перечисленных выше, были обнаружены клоны, экспрессирующие белки с ненарушенной функцией, то из соответствующей конструкции (конструкций) можно было удалить репортерные нуклеотидные последовательности и такой конструкцией (конструкциями) с доказанными функциональными свойствами можно трансформировать полезные пищевые растения. Целью изобретения является получение композиции в растительном материале, включающей белок-носитель, обогащенный аминокислотами и удлиненный с помощью стабильных аминокислотных кассет; в особенности белок, который связан с клеточными стенками и жмыхом, полученном после отжима масла из масличных растений. В воплощениях нашего изобретения можно было бы продемонстрировать стабильные обогащенные аминокислотами белки семян. Принципы, лежащие в основе изобретения, можно осуществлять с помощью многочисленных известных технологий рекомбинантных ДНК, доступных в настоящее время. Поэтому приведенное ниже краткое изложение демонстрирует лишь предпочтительные воплощения изобретения. Изобретение детально описано в следующих примерах. Продукция белка, обогащенного аминокислотами, описана на примере использования гистидина, метионина, цистеина, глицина и лизина, но принципы, раскрытые в изобретении, можно применить к любой другой желаемой аминокислоте (аминокислотам) с помощью вставки кассет, включающей селективное число кодонов желаемых аминокислот для любых предусматриваемых целей. Пример 1. Выделение хромосомной ДНК и ПЦР. Хромосомную ДНК выделяли следующим образом. 300 мкг листьев A. thaliana гомогенизировали в ступке с жидким азотом до получения тонкодисперсного порошка. Затем добавляли 3 мл среды, содержавшей 100 мМ трис-HCl; рН 8,0, 500 мМ Nad, 50 мМ ЭДТА, затем добавляли 600 мкл 10%-ного додецилсульфата натрия (SDS) и 500 мкл 20%-ного поливинилпирролидона (средняя мол. масса 360000) и тщательно измельчали. Смесь переносили в полипропиленовую пробирку и инкубировали при 65°С в течение 10 минут. Затем в раствор добавляли 450 мкл охлажденного до температуры льда ацетата калия и смесь аккуратно перемешивали, переворачивая пробирку, инкубировали на льду в течение 30 минут и центрифугировали в течение 10 минут при 8000 об/мин при 4°С. Супернатант экстрагировали смесью фенол/хлороформ (1:1) и затем ДНК осаждали изопропанолом. Осажденную ДНК промывали 70%-ным этанолом и перед растворением в 10 мМ трис-HCl; рН 8,0, высушивали на воздухе. Реакционная среда для ПЦР включала однократный (1х) буфер для ПЦР (10х-буфер: 500 мМ KCl, 100 мМ трис-HCl; рН 9,0 при 25°С и тритон Х-100), содержащий 1,5 мМ MgCl2, 1 мкл полимеразы Taq (5 ед/мл), 0,2 мМ раствор всех dNTP, праймеры в концентрации 0,4 мкМ и 3 мкг геномной ДНК A. thaliana в 25 мкл реакционного объема. Матрицу денатурировали нагреванием в течение 3 минут при 95°С и проводили 30 циклов ПЦР, используя термоциклер iCycler Пример 2. Синтез и клонирование последовательности ДНК, обогащенной гистидиновыми кодонами. Чтобы сконструировать фрагмент ДНК, обогащенный гистидиновыми кодонами, использовали три химически синтезированных сегмента одноцепочечной ДНК: H-P-l(SEQ ID NO:n: 5′-GCGCCTCGAGTTCACCATCACCATCACCATCACGGGCACCATCAC-3′ Н-Р-2 (SEQ ID NO:2): 5′-CATCACCATCACCATGG-3′ H-M(SEQ ID NO:3): 5′-CCGGATCCTAAAGTCGACCATGGTGATGGTGATGGTGATGGTGCC-3′. Чтобы получить фрагмент двухцепочечной ДНК, несущий последовательность, обогащенную His кодонами, олигонуклеотиды Н-М и Н-Р-2 были подвергнуты отжигу в 1x-реакционном буфере для обратной транскриптазы (RT) AMV (50 мМ трис-HCl; рН 8,3, 50 мМ KCl, 10 мМ MgCl2, 0,5 мМ спермидин, 10 мМ DTT) в течение 30 минут при комнатной температуре, и реакцию элонгации цепи проводили в присутствии 1x-AMV RT буфера, содержащего 1 мМ dNTP и 1 единицу AMV RT в течение 45 минут при 37°С. Продукт реакции был отделен от негибридизованных олигонуклеотидов с помощью электрофореза в агарозном геле, очищен и подвергнут отжигу с олигонуклеотидом Н-Р-1 в 1x-буфере для большого фрагмента ДНК-полимеразы I (фрагмент Кленова) (50 мМ трис-HCl; рН 7,2, 10 мМ MgSO4, 0,1 мМ DTT) в течение 30 минут при комнатной температуре. Реакцию элонгации цепи проводили с 5 единицами полимеразы Кленова в присутствии 25 мМ каждого из dNTP в 1x-буфере Кленова в течение 30 минут при 37°С. Фрагмент двухцепочечной ДНК ожидаемого размера (79 п.н.) вырезали из агарозного геля (после соответствующего электрофоретического разделения), очищали, расщепляли с помощью рестриктаз Xhol и Bamtil и клонировали в расщепленном этими же рестриктазами векторе для клонироания pGEM-7Zf(+) (Promega Corporation, USA; номер по каталогу Р2251). После рестрикционного анализа и секвенирования для дальнейших манипуляций отбирали клон pGEM-His-24. Этот клон содержал сегмент ДНК, который мог бы потенциально кодировать 19-членный пептид, содержащий 14 остатков гистидина (фигура ЗА). Последовательность этого фрагмента ДНК фланкирована сайгами рестрикции Xhol и BamHl и содержала также сайг для рестриктазы Sail, который был сконструирован для последующих стадий клонирования (фигура 3А). Пример 3. Синтез и клонирование последовательности ДНК, обогащенной цистеиновыми и метиониновыми кодонами. Чтобы сконструировать фрагмент ДНК, обогащенный цистеиновыми и метиониновыми кодонами, использовали два химически синтезированных сегмента одноцепочечной ДНК: С-М-Р-1 (SEQ ID NO:4): 5′-CACCTCGAGTATGTTGTTGCATGTGCATGTGCTGTTGCATGTCGACAAAC-3′ C-M-P-2 (SEQ ID NO:5): 5′-GTTTGTCGACATGCAACAGCACATGCACATGCAACAACATACTCGAGGTG-3′ Чтобы получить фрагмент двухцепочечной ДНК, несущий последовательность, обогащенную Cys/Met кодонами, олигонуклеотиды С-М-Р-1 и C-M-P-2 были подвергнуты отжигу в 1x-буфере для большого фрагмента ДНК-полимеразы I (фрагмент Кленова) (50 мМ трис-HCl; рН 7,2, 10 мМ MgSO4, 0,1 мМ DTT) в течение 30 минут при комнатной температуре. Фрагмент двухцепочечной ДНК ожидаемого размера (50 п.н.) вырезали из агарозного геля (после соответствующего электрофоретического разделения), очищали, расщепляли рестриктазами XhoI и BamHl и клонировали в расщепленный таким же образом вектор для клонирования pGEM-7Zf(+) (Promega Corporation, USA; номер по каталогу Р2251). После рестрикционного анализа и секвенирования для дальнейших манипуляций отобрали клон pGEM-Cys/Met-10. Этот клон содержал сегмент ДНК, который мог бы потенциально кодировать 16-членный пептид, содержащий 10 Cys-Met аминокислотных остатков (фигура 3B). Последовательность этого фрагмента ДНК фланкирована сайтами рестрикции XhoI и BamI и содержит также сайт для рестриктазы Sail, который был сконструирован для последующих стадий клонирования (фигура 3B). Пример 4. Синтез и клонирование последовательности ДНК, обогащенной глициновыми кодонами. Чтобы сконструировать фрагмент ДНК, обогащенный глициновыми кодонами, использовали два химически синтезированных сегмента одноцепочечной ДНК: GL-P-1(SEQ ID NO:6): 5′-GCGCCTCGAGTTGGTGGAGGTGGAGGCGGTGGAGGTGGCGTCGACAAATGG АТСССС-3′ GL-P-2 (SEQ ID NO:7): 5′-GGGGATCCATTTGTAGACGCCACCTCCTCCACCGCCTCCACCTCCACCAACT CGAGGCGC-3′ Чтобы получить фрагмент двухцепочечной ДНК, несущий последовательность, обогащенную глициновыми кодонами, олигонуклеотиды GL-P-1 и GL-P-2 были подвергнуты отжигу в 1x-буфере для большого фрагмента ДНК-полимеразы I (фрагмент Кленова) (50 мМ трис-HCl; рН 7,2, 10 мМ MgSO4, 0,1 мМ DTT) в течение 30 минут при комнатной температуре. Фрагмент двухцепочечной ДНК ожидаемого размера (60 п.н.) вырезали из агарозного геля (после соответствующего электрофоретического разделения), очищали, расщепляли с помощью рестриктаз XhoI и BamHI и клонировали в расщепленный таким же образом вектор для клонирования pGEM-7Zf(+) (Promega Corporation, USA; номер по каталогу Р2251). После рестрикционного анализа и секвенирования для дальнейших манипуляций отобрали клон pGEM-Gly-9. Этот клон содержал сегмент ДНК, который мог бы потенциально кодировать 19-членный пептид, содержащий 9 Gly аминокислотных остатков (фигура 3С). Последовательность этого фрагмента ДНК фланкирована сайтами рестрикции XhoI и BamHI и содержит также сайт для рестриктазы Sail, который был сконструирован для последующих стадий клонирования (фигура 3С). Пример 5. Синтез и клонирование последовательности ДНК, обогащенной лизиновыми кодонами. Чтобы сконструировать фрагмент ДНК, обогащенный лизиновыми кодонами, использовали два химически синтезированных сегмента одноцепочечной ДНК: L-P-1(SEQ ID NO:8): 5′-GCGCCTCGAGTTAAAAAGAAAAAGAAAAAGAAAAAGAAAAAGAAAAAGGT CGACAAATGGATCCCC-3′ L-P-2(SEQ ID NO:9): 5′-GGGGATCCATTTGTCGACCTTTTTCTTTTTCTTTTTCTTTTTCTTTTTCTTTTTA ACTCGAGGCGC-3′ Чтобы получить фрагмент двухцепочечной ДНК, несущий последовательность, обогащенную лизиновыми кодонами, олигонуклеотиды L-P-1 и L-P-2 были подвергнуты отжигу в 1x-буфере для большого фрагмента ДНК-полимеразы I (фрагмент Кленова) (50 мМ трис-HCl; рН 7,2, 10 мМ MgSO4, 0,1 мМ DTT) в течение 30 минут при комнатной температуре. Фрагмент двухцепочечной ДНК ожидаемого размера (66 п.н.) вырезали из агарозного геля (после соответствующего электрофоретического разделения), очищали, расщепляли с помощью рестриктаз XhoI и BamHI и клонировали в расщепленный таким же образом вектор для клонирования pGEM-7Zf(+) (Promega Corporation, USA; номер по каталогу Р2251). После рестрикционного анализа и секвенирования для дальнейших манипуляций отобрали клон pGEM-Lys-12. Этот клон содержал сегмент ДНК, который мог бы потенциально кодировать 22-членный пептид, содержащий 12 лизиновых аминокислотных остатков (фигура 3D). Последовательность этого фрагмента ДНК фланкирована сайтами рестрикции XhoI и BamHI и содержала также сайт для рестриктазы Sail, который был сконструирован для последующих стадий клонирования (фигура 3D). Пример 6. Выделение гена олеозина (ole) из хромосомной ДНК Arabidopsis thaliana с использованием ПЦР и последующего клонирования. Чтобы клонировать ген олеозина из хромосомной ДНК Arabidopsis thaliana, были химически синтезированы два олигонуклеотидных праймера: OLE-P(SEQ ID NO:10): 5′-AAAACCATGGCGGATACAGCTAGAGGAACCCATC-3′ OLE-M(SEQID NO:11): 5′-GGGGCCATGGGAGTAGTGTGCTGGCCACCACGAGTAC-3′ Олигонуклеотиды OLE-P и OLE-M использовали в качестве праймеров для ПЦР геномной ДНК из Arabidopsis thaliana (пример 1). Полученный фрагмент ПЦР имел тупые концы и был клонирован по сайту рестрикции Smal в вектор pGEM-3Zf(+) (Promega Corporation, USA; номер по каталогу Р2271). Рекомбинантные клоны были проверены с помощью рестрикционного анализа и секвенирования. Для дальнейшей работы был отобран клон pOLE4H. Если сравнивать с опубликованной последовательностью олеозина (Х62353), последовательность клона pOLE4H содержала одну нуклеотидную замену (без изменений в последовательности кодируемых аминокислот) в интроне гена – замену G на А в положении 513 последовательности pOLE4H. В качестве праймера для ПЦР для амплификации участка плазмиды pOLE4H, кодирующей олеозин, был химически синтезирован и использован олигонуклеотид OLE-3′ XhoI (SEQ ID NO: 12) 5′-GCGCCTCGAGAAGTAGTGTGCTGGCCACCAC-3′, продукт ПЦР был клонирован по сайту рестрикции Xhol в вектор pGEM-7Zf(+) (Promega Corporation, USA; номер по каталогу Р2251). После клонирования pOLE4H была секвенирована с использованием этого праймера для того, чтобы убедиться в том, что все кодирующие свойства гена олеозина в полученных клонах были сохранены. На основании этого анализа был отобран клон pOLE4/11. pOLE4/11 содержал ген олеозина дикого типа без терминирующего кодона, фланкированный сайтами рестрикции Ncol и Xhol, подходящими для последующих стадий клонирования. Пример 7. Выделение гена транспортного белка 30К из геномной РНК вируса табачной мозаики (TMV 30К МР) с использованием ПЦР и последующего клонирования. Чтобы клонировать ген транспортного белка 30К из геномной РНК TMV были химически синтезированы два специфичных олигонуклеотидных праймера: Олигонуклеотидный праймер N30K (SEQ ID NO: 13): 5′-GCGGAATTCCCATGGCTCTAGTTGTTAAAGG-3′ Олигонуклеотидный праймер С30К (SEQ ID NO: 14): 5′-AGACCTCGAGGAAACGAATCCGATTCGGCGAC-3′ Эти праймеры содержали рестрикционные сайты EcdSl и Xhol, соответственно. Первая цепь кДНК была синтезирована на геномной РНК TMV, используя 20 нМ праймер С 30К, 1 мМ dNTP и 1 единицу AMV в 1x-реакционном буфере обратной транскриптазы (RT) AMV (50 мМ трис-HCl; рН 8,3, 50 мМ KCl, 10 мМ MgCb 0,5 мМ спермидин, 10 мМ DTT) в течение 45 минут при 37°С. После этого 5 мкл реакционной смеси помещали в 1x-буфер для ПЦР (10х буфер: 500 мМ KCl, 100 мМ трис-HCl; рН 9,0 при 25°С и тритон Х-100), содержащий 1,5 мМ MgCl2, 2 мкл полимеразы Taq (5 ед/мл), 0,2 мМ dNTP, праймеры в концентрации 0,4 мкМ в 25 мкл реакционного объема. Матрицу денатурировали нагреванием в течение 3 минут при 95°С, и проводили 28 циклов ПЦР, используя термоциклер iCycler Пример 8. Микропроекционная бомбардировка. Микропроекционная бомбардировка (бомбардировка частицами) была проведена с помощью метода разрывного диска (rupture disk method) с использованием аппарата высокого давления на основе гелия PD S-1000 (Bio-Rad), как описано в работе Morozov et al. (1997). Вольфрамовые частицы получали на вортексе перемешиванием 60 мг частиц в 70%-ном этаноле в течение 3-5 минут с последующея инкубацией на столе в течение 15 минут. Смесь осаждали коротким центрифугированием и супернатант удаляли. На осадок наслаивали стерильную воду и перемешивали на вортексе в течение 1 минуты. Затем давали частицам осесть в течение 1 минуты и центрифугировали в течение 2 секунд. Супернатант отбрасывали. Эту процедуру повторяли три раза. Добавляли стерильный 50%-ный глицерин таким образом, чтобы довести концентрацию частиц до 60 мг/мл. Для каждой стадии бомбардировки на вольфрамовые частицы (М-20, 1,3 мк) с помощью хлорида кальция и этанола осаждали ДНК. Добавляли 5-10 мкг плазмиды ДНК, 50 мкл CaCl2 (2,5 М) и 20 мкл спермидина (0,1 М) и перемешивали на вортексе в течение 2-3 минут, затем давали смеси осесть и центрифугировали в течение 2 секунд. Супернатант отбрасывали и на поверхность осадка наносили 140 мкл 70%-ного этанола, затем удаляли его, наносили 100%-ный этанол и удаляли его, стараясь не нарушать осадок, процедуру повторяли. На макроноситель наносили 6 мкл суспензии и использовали для бомбардировки. Отделенный лист Nicotiana benthamiana (размером 15-30 мм) помещали в центр пластиковой чашки Петри и на твердой подложке бомбардировали частицами с расстояния от цели в 7 см. Бомбардировку проводили с помощью пульса гелия 1350 кПа в вакуумной камере. Инокулированные листья анализировали через 24 часа после бомбардировки частицами. С помощью конфокальной лазерной сканирующей видеосистемы MRC-1024 (Bio-Rad) измеряли флуоресценцию GFP. Пример 9. Конструирование штамма Е. coli, суперпродуцирующего С-терминальный гидрофильный конец олеозина для последующей иммунизации кроликов и получения олеозин-специфических антител. Большая часть N-концевого участка последовательности олеозина гидрофобна, в то время как аминокислотная последовательность С-терминального участка олеозина, кодируемая вторым экзоном гена олеозина, представлена гидрофильной последовательностью, подходящей для экспрессии в Е. coli и используемой в качестве антигена при иммунизации. Поэтому для дальнейшей работы был выбран С-концевой участок олеозина (аминокислоты 120-173). Второй экзон гена олеозина был амплифицирован в ПЦР с использованием плазмиды pOLE4/11 (пример 6) в качестве матрицы и олигонуклеотидных праймеров: OLE-EX-B (SEQ ID NO: 15) 5′-TGTGGGATCCTACGCAACGGGAGAGCACCCA-3′ OLE-3’XhoI (SEQ ID NO:12) 5′-GCGCCTCGAGAAGTAGTGTGCTGGCCACCAC-3′ содержащих сайты рестрикции BamHl и Xhol, соответственно. Полученный ПЦР продукт был гидролизован BamHl и Xhol и клонирован в вектор pGEM-7Zf(+) (Promega Corporation, USA; номер по каталогу Р2251), расщепленный рестриктазами XhoI и BamHI. После рестрикционного анализа и секвенирования полученных клонов для последующих стадий клонирования был отобран клон pGEM-OLE2EX. Чтобы получить экспрессирующиеся конструкции, после расщепления рестриктазами BamHI и XhoI второй экзон из pGEM-OLE2EX был удален и клонирован в экспрессирующий вектор pQE30 (QIAGEN), расщепленный BamHI и SaiI. В полученной конструкции pQE-OLE2EX 3′-терминальная часть гена олеозина была трансляционно слита с лидерной последовательностью, содержащей 6xHis-конец, присутствие которого делает воможным аффинную очистку экспрессированного продукта. Плазмида pQE-OLE2EX была трансформирована в Е. coli, штамм М15 [pREP4] (QIAGEN). Индукцию экспрессии рекомбинантного белка проводили в среде Luria broth, добавляя IPTG до конечной концентрации 1 мМ, с последующей инкубацией на вращающемся шейкере при 220 об/мин в течение 2-4 часов при 37°С. Клетки Е. coli из индуцированной и неиндуцированной культуры собирали и экспрессию рекомбинантного белка анализировали с помощью электрофореза в ПААГ в присутствии SDS. Для проведения электрофореза готовили 12,5%-ный акриламидный гель. В течение 45 минут-1 часа через гель пропускали электрический ток 100 В/500 мА и затем гель окрашивали с помощью СВВ (0,05% Coomassie Brilliant Blue, 50% метанола, 7% ледяной уксусной кислоты) и отмывали смесью 7% метанол – 5% уксусная кислота. Основная полоса, присутствующая в индуцированных культурах, отсутствовала в неиндуцированных культурах. Подвижность этой полосы соответствовала ожидаемой подвижности С-терминальной части гена олеозина с His-концом (8,3 кДа). Пример 10. Система трансляции in vitro. Бесклеточная система трансляции in vitro (IVT) создает возможность селекции/идентификации оптимального количества кодонов, которые можно было бы транслировать правильным образом. Система IVT включает экстракт зародышей пшеницы, содержащий все компоненты (например, рибосомы 70S или 80S, тРНК, аминоацил-тРНК-синтетазы, факторы инициации, элонгации и терминации), необходимые для трансляции экзогенной РНК даже в присутствии низких концентраций двуцепочечной РНК (dsRNA) или окисленных тиолов. Экстракт также содержит необходимые аминокислоты, источники энергии (АТФ, ГТФ), системы регенерации энергии (креатинфосфат и креатинфосфокиназа) и другие важные кофакторы (Mg2+ К+ и т.д.). Для IVT анализа были отобраны клоны с 2, 4 и 6 повторами последовательностей, обогащенных His-кодонами. Плазмиды pMP-1x, pMP-2x, рМР-4х и рМР-6х, содержавшие ген 30К МР, слитый с последовательностями различной длины, кодирующими His, помещали под контроль промотора Т7. Плазмиды линеаризовали с помощью BamHI и использовали в качестве матрицы в IVT в присутствии РНК-полимеразы Т7. Равные количества полученных транскриптов транслировали в бесклеточной системе зародышей пшеницы (TNT®T7/SP6 Coupled Wheat Germ Extract System L5030, Promega corporation). Было установлено, что транскрипты, соответствующие клонам с одним, двумя и четырьмя повторами последовательности, обогащенной His-кодонами, вызывали появление продукта с ожидаемой молекулярной массой, тогда как транскрипты, потенциально способные кодировать белок с 6 повторами последовательности, обогащенной His (МР-6х), транслировались неэффективно и вызывали появление продуктов с низкой молекулярной массой неопределенного размера. Этот подход для селекции/идентификации клонов проиллюстрирован на примерах 11-14. Пример 11. Слияние последовательностей ДНК, обогащенных гистидиновыми кодонами, и гена TMV 30К МР и контроль точности трансляции генов, обогащенных His-кодонами, с использованием системы трансляции in vitro. Слияния последовательностей ДНК, обогащенных гистидиновыми кодонами, и гена TMV 30К МР проводили с помощью соответствующих клонирования комбинаций pGEM-His-24 (пример 2), включающей последовательность, кодирующую 14 остатков His (одна кассета) (фигура 3А), и плазмиды pGEM-30К (пример 7), включающей ген TMV 30К МР, не содержащий свой природный терминирующий кодон. Чтобы получить последовательности различной длины, обогащенные кодонами His, слитые с геном 30К МР, проводили ступенчатую процедуру клонирования (фигура 4). На первой стадии ген 30К МР вырезали из вектора pGEM-ЗОК с помощью рестриктаз EcoRI и XhoI, a последовательность, обогащенную His-кодонами, вырезали из вектора pGEM-His-24 с помощью рестриктаз XhoI и BamHI. Оба фрагмента ДНК лигировали в вектор pGEM-3Zf(+) (Promega Corporation, USA; номер по каталогу Р2271), расщепленный с помощью рестриктаз EcoRl и BamHI (фигура 4). Полученный клон, обозначенный pMP-1x, содержал один набор гистидин-кодирующей последовательности (одну кассету) из вектора pGEM-His-24, слитой с геном TMV 30К МР. Дополнительные стадии ступенчатого клонирования, проведенные для увеличения длины His-кодирующего “хвоста” в слитом гене МР, были основаны на использовании плазмиды pMP-1x. В этих процедурах клонирования были использованы сайты рестрикции XhoI и SalI, которые были сконструированы в последовательности pGEM-His-24, обогащенной His-кодонами, и примыкали к рамке считывания кодируемого полипептида. Наряду с тем, что расщепление с помощью рестриктаз XhoI и SalI приводило к образованию идентичных липких концов, на дальнейших стадиях клонирования, которые описаны ниже, это защищало рамку считывания в полученной дополнительно удлиненной последовательности, содержащей кодоны His. Чтобы получить плазмиду с двумя наборами гистидин-кодирующей последовательности рМР-2х из pGEM-His-24, EcoRI-SalI-фрагмент pMP-1x был клонирован в pMP-1x, расщепленный рестриктазами EcoRI и XhoI, что привело к удвоению набора гистидин-кодирующей последовательности. Полученная таким образом плазмида рМР-2х содержала в С-терминальном “хвосте” МР 24 остатка гистидина. Затем, чтобы получить рМР-4х, -EcoRI-SalI-фрагмент рМР-2х был клонирован в рМР-2х, расщепленный рестриктазами EcoRI и XhoI; чтобы получить рМР-6х – EcoRI- SalI-фрагмент рМР-4х был клонирован в рМР-2х, расщепленный рестриктазами EcoRI и XhoI; чтобы получить рМР-8х – EcoRI-SalI-фрагмент рМР-4х был клонирован в рМР-4х, расщепленный рестриктазами EcoRI. и XhoI. В частности аналогичные стадии клонирования повторяли для того, чтобы получить плазмиды, несущие 56, 84 и 112 гистидиновых остатков (4х, 6х и 8х кассет) в С-терминальном “хвосте” МР (фигура 4). Клоны с 112 His-кодонами в С-терминальном “хвосте” (серии рМР-8х) при выращивании в жидкой среде были не стабильны и приводили к образованию делеционных вариантов (мини-плазмид). Также наблюдали сниженную стабильность для некоторых клонов с 84 остатками гистидинами (серии рМР-6х) при многократном выращивании в жидкой среде. Таким образом, для дальнейших экспериментов были отобраны только клоны рМР-2х, рМР-4х и рМР-6х с 2, 4 и 6 повторами последовательности, обогащенной His-кодонами (фигура 4). В плазмидах pMP-1x, рМР-2х, рМР-4х и рМР-6х ген 30К МР, слитый с последовательностями различной длины, кодирующими His, был помещен под контроль Т7 промотора. Эти плазмиды перед транскрипцией с РНК-полимеразой Т7 in vitro были линеаризованы с помощью рестриктазы BamRI. Равные количества полученных транскриптов были транслированы в бесклеточной системе зародышей пшеницы (TNT®T7/SP6 Coupled Wheat Germ Extract System L5030, Promega corporation, описана в примере 10). Было обнаружено, что транскрипты клонов с одним, двумя и четырьмя повторами последовательности, обогащенной His-кодонами (trMP-1x, trMP-2x и trMP-4x), вызывали появление продуктов с ожидаемыми постепенно увеличивающимися молекулярными массами, тогда как транскрипт, потенциально способный кодировать белок с 6 повторами последовательности, обогащенной His (trMP-6х), транслировался неэффективно и вызывал появление продуктов с низкой молекулярной массой неопределенного размера. Этот эксперимент показал, что трансляция последовательностей, обогащенных His-кодонами, количество которых превышает 56 остатков, происходит с ошибками, например, His-кодоны могли представлять собой “бесплодные” (hungry) кодоны, на которых происходила остановка и “сбои” рибосомы. На основании этих результатов для дальнейшей работы т vivo были отобраны только клоны с 2 и 4 повторами. Пример 12. Слияние последовательностей ДНК, обогащенных цистеин-метиониновыми кодонами, и гена TMV 30К МР и контроль точности трансляции генов, обогащенных Cys/Met-кодонами, с использованием системы трансляции in vitro. Слияния последовательностей ДНК, обогащенных Cys/Met-кодонами, и гена TMV 30К МР проводили с помощью соответствующего клонирования комбинации pGEM – Cys/Met-10 (пример 3), включающей последовательность, кодирующую 10 Cys/Met-остатков (одна кассета) (фигура 3B), и плазмиды pGEM-30К (пример 7), включающей ген TMV 30К МР, не содержащий свой природный терминирующий кодон, как это описано в примере 10. Чтобы получить последовательности различной длины, обогащенные кодонами Cys/Met, слитые с геном 30К МР, проводили ступенчатую процедуру клонирования (фигура 4). На первой стадии ген 30К МР вырезали из вектора pGEM-ЗОК с помощью рестриктаз EcoRI и XhoI, а последовательность, обогащенную Cys/Met-кодонами, вырезали из вектора pGEM-Cys/Met-10 с помощью рестриктаз XhoI и BamHI. Оба фрагмента ДНК лигировали в вектор pGEM-3Zf(+) (Promega Corporation, USA; номер по каталогу Р2271), расщепленный с помощью рестриктаз EcoRI и BamHI (фигура 4). Полученный клон, обозначенный pMP-Cys/Met-1x, содержал один набор цистеин/метионин-кодирующей последовательности (одну кассету) из вектора pGEM-Cys/Met-10, слитой с геном TMV 30К МР. Чтобы получить плазмиду с двумя наборами цистеин/метионин-кодирующей последовательности pMP-Cys/Met-2x из pGEM-Cys/Met-10, XhoI-SalI-фрагмент pMP-Cys/Met-1x был клонирован в pMP-Cys/Met-1x, расщепленный рестриктазами EcoRI и XhoI, что привело к удвоению набора цистеин/метионин-кодирующей последовательности. Таким образом полученная плазмида pMP-Cys/Met-2x содержала 20 остатков цистеина/метионина в С-терминальном “хвосте” МР. Затем, чтобы получить pMP-Cys/Met-4x, EcoRI-SalI-фрагмент pMP-Cys/Met-2x был клонирован в pMP-Cys/Met-2x, расщепленный рестриктазами EcoRI и Xhol; чтобы получить pMP-Cys/Met-бх, EcoRI-SalI-фрагмент pMP-Cys/Met-4x был клонирован в pMP-Cys/Met-2x, расщепленный рестриктазами EcoRI и Xhol; и чтобы получить pMP-Cys/Met-8x, EcoRI-SalI-фрагмент pMP-Cys/Met-4x был клонирован в pMP-Cys/Met-4x, расщепленный рестриктазами EcoRI и XhoI. В плазмидах pMP-Cys/Met-1x, pMP-Cys/Met-2x, pMP-Cys/Met-4x и pMP-Cys/Met-6х ген 30К МР, слитый с последовательностями различной длины, кодирующими Cys/Met, был помещен под контроль Т7 промотора. Плазмиды были линеаризованы с помощью рестриктазы BamHI и подвергнуты транскрипции in vitro с РНК-полимеразой Т7. Равные количества полученных транскриптов были транслированы в бесклеточной системе зародышей пшеницы (TNT®T7/SP6 Coupled Wheat Germ Extract System L5030, Promega corporation). Было обнаружено, что транскрипты клонов с одним, двумя и четырьмя повторами последовательности, обогащенной Cys/Met-кодонами (MP-Cys/Met-1x, МР-Cys/Met-2x и MP-Cys/Met-4x), вызывали появление продуктов с ожидаемыми постепенно увеличивающимися молекулярными массами. Также как это было проиллюстрировано на примере 11, увеличение длины аминокислотной последовательности привело к уменьшению стабильности транскриптов. Для дальнейшей работы из последовательностей, обогащенных Cys/Met-кодонами, были отобраны только клоны с 2, 4 и 6 кассетами (MP-Cys/Met-2x, MP-Cys/Met-4x и MP-Cys/Met-6х). Пример 13. Слияние последовательностей ДНК, обогащенных глициновыми кодонами, и гена TMV 30К МР и контроль точности трансляции генов, обогащенных Gly-кодонами, с использованием системы трансляции in vitro. Слияния последовательностей ДНК, обогащенных глициновыми кодонами, и гена TMV 30К МР проводили с помощью соответствующего клонирования комбинации pGEM-Gly-9 (пример 4), включающей 9 остатков Gly (одна кассета) (фигура 3С), и плазмиды pGEM-30К, включающей ген TMV 30К МР, не содержащий свой природный терминирующий кодон, также как в примере 10. Чтобы получить последовательности различной длины, обогащенные кодонами Gly, слитые с геном 30К МР, проводили ступенчатую процедуру клонирования, такую как в примерах 11 и 12 и показанную на фигуре 4. Полученный первый клон, обозначенный pMP-Gly-1x, содержал один набор глицин-кодирующей последовательности (одну кассету) из вектора pGEM-Gly-9, слитой с геном TMV 30К МР. Затем эту кассету “размножали”, как в примерах 11 и 12; чтобы создать клоны pMP-Gly-2x, pMP-Gly-4x, pMP-Gly-бх и pMP-Gly-8x. Чтобы получить плазмиду с двумя наборами глицин-кодирующей последовательности pMP-Gly-2x из pGEM-Gly-9, EcoRI-SalI-фрагмент pMP-Gly-1x был клонирован в pMP-Gly-1x, расщепленный рестриктазами EcoRI и XhoI, что привело к удвоению набора глицин-кодирующей последовательности. Затем, чтобы получить pMP-Gly-4x,.EcoRI-SalI-фрагмент pMP-Gly-2x был клонирован в pMP-Gly-2x, расщепленный рестриктазами EcoRI и XhoI; чтобы получить pMP-Gly-6х, EcoRI-SalI-фрагмент pMP-Gly-4x был клонирован в pMP-Gly-2x, расщепленный рестриктазами EcoRI и XhoI; и чтобы получить pMP-Gly-8x, EcoRI-SalI-фрагмент pMP-Gly-4x был клонирован в pMP-Gly-4x, расщепленный рестриктазами EcoRI и XhoI. Плазмиды были линеаризованы с помощью рестриктазы BamHI и подвергнуты транскрипции in vitro с РНК-полимеразой Т7. Равные количества полученных транскриптов были транслированы в бесклеточной системе зародышей пшеницы (TNT®T7/SP6 Coupled Wheat Germ Extract System L5030, Promega corporation, описана в примере 10). Также как это было проиллюстрировано на примере 11, увеличение длины аминокислотной последовательности привело к уменьшению стабильности транскриптов. Для дальнейшей работы из последовательностей, обогащенных глициновыми кодонами, были отобраны только клоны с 2 и 4 кассетами (MP-Gly-2x и MP-Gly-4x). Пример 14. Слияние последовательностей ДНК, обогащенных лизиновыми кодонами, и гена TMV 30К МР и контроль точности трансляции генов, обогащенных Lys-кодонами, с использованием системы трансляции in vitro. Слияния последовательностей ДНК, обогащенных лизиновыми кодонами, и гена TMV 30К МР проводили с помощью соответствующего клонирования комбинации pGEM-Lys-12 (пример 5), включающей 12 остатков Lys (одна кассета) (фигура 3D), и плазмиды pGEM-30К, включающей ген TMV 30К МР, не содержащий свой природный терминирующий кодон, также как в примерах 11-13. Чтобы получить последовательности различной длины, обогащенные лизиновыми кодонами, слитые с геном 30К МР, проводили ступенчатую процедуру клонирования, такую как в примерах 11, 12 и 13 и показанную на фигуре 4. Полученный первый клон, обозначенный pMP-Lys-1x, содержал один набор лизин-кодирующей последовательности (одну кассету) из вектора pMP-Lys-12, слитой с геном TMV 30К МР. Затем эту кассету “размножали”, как в примерах 11,12 и 13; чтобы создать клоны pMP-Lys-2x, pMP-Lys-4x, pMP-Lys-6х и pMP-Lys-8x. Чтобы получить плазмиду с двумя наборами лизин-кодирующей последовательности pMP-Lys-2x из pMP-Lys-12 EcoRI-SalI-фрагмент pMP-Lys-1x был клонирован в pMP-Lys-1x, расщепленный рестриктазами EcoRI и XhoI, что привело к удвоению набора лизин-кодирующей последовательности. Затем, чтобы получить pMP-Lys-4x, EcoRI-SalI-фрагмент pMP-Lys-2x был клонирован в pMP-Lys-2x, расщепленный рестриктазами EcoRI и XhoI; чтобы получит pMP-Lys-6х, EcoRI-SalI-фрагмент pMP-Lys-4x был клонирован в pMP-Lys-2x, расщепленный рестриктазами EcoRI и XhoI; и чтобы получить pMP-Lys-Sx, EcoRI-SalI-фрагмент pMP-Lys-4x был клонирован в pMP-Lys-4x, расщепленный рестриктазами EcoRI и XhoI. Плазмиды были линеаризованы с помощью рестриктазы BamHI, подвергнуты транскрипции in vitro с РНК-полимеразой Т7 и транслированы в бесклеточной системе зародышей пшеницы (TNT®T7/SP6 Coupled Wheat Germ Extract System L5030, Promega corporation, описана в примере 10). Также как это было проиллюстрировано на примерах 10-12, увеличение длины аминокислотной последовательности привело к уменьшению стабильности транскриптов. Для дальнейшей работы из последовательностей, обогащенных лизиновыми кодонами, были отобраны только клоны с 2 и 4 кассетами (МР-Lys-2x и MP-Lys-4x). Пример 15. Выделение промотора напина из хромосомной ДНК Arabidopsis thaUana с использованием ПЦР и ступенчатого клонирования. Чтобы клонировать промотор напина из хромосомной ДНК Arabidopsis thaliana, были химически синтезированы два олигонуклеотидных праймера: NAP-P (SEQ ID NO:16): 5′-TCTTACTCGAGTGAAACCAAATTAAC-3′ NAP-M (SEQ ID NO:17): 5′-CTTGTTAGCCATGGTTTGCTATTTGTG-3′. Для облегчения ступенчатого клонирования последовательности праймеров содержали сайты рестрикции XhoI и NcoI. Хромосомную ДНК Arabidopsis thaliana выделяли, как описано для выделения (пример 1), и клонирования гена олеозина (пример 6). ПЦР проводили, как описано выше для гена олеозина (пример 6), за исключением того, что концентрация MgCl2 в реакционной среде составляла 1,5 мМ. Затем после электрофореза из 1%-ного агарозного геля выделяли продукт ПЦР ожидаемых размеров (369 п.н.). Продукт ПЦР очищали и обрабатывали ДНК-полимеразой Т4 в присутствии dNTP при 14°С, как в примере 1, и клонировали в вектор pGEM-3Zf(+) (Promega Corporation, USA; номер по каталогу Р2271). После секвенирования для дальнейшей работы отобрали клоны pGEM-NAP4 и pGEM-NAP9. Пример 16. Создание экспрессионных плазмид, содержащих промотор напина (NAP) или химерный промотор (HYB), включающий полный промотор напина и энхансерный участок промотора 35S гена вируса мозаики цветной капусты. Экспрессирующие векторы растений, содержащие промотор напина, были сконструированы с помощью плазмиды pRT100 (Topfer et al., 1987. A set of plant expression vectors for transcriptional and translational fusions. Nucleic Acid Res. 15(14): 5890). Основная стратегия для слияния генов МР и GFP с вектором 2x-Gly (т.е. с MP-Gly-2x-GFP) проиллюстрирована на фигуре 7. В этой стратегии использовали сайты рестрикции XhoI и SalI, которые различаются по месту расщепления, но имеют сходные 4-нуклеотидные выступающие концы ДНК. Ген GFP был вырезан из pRT-GFP в виде XhoI-BamHI-фрагмента, и чтобы получить конструкцию pRT-MP-2x-GFP, оба фрагмента ДНК были лигированы в плазмиду pRT101 (Topfer et al., Nucleic Acids Res. 15(14): 5890, 1987), расщепленную рестриктазами EcoRl и BamHI. PRT100 содержит промотор 35S гена вируса мозаики цветной капусты (CaMV 35S промотор). Для конструирования pNAP, в котором целиком удалили последовательность CaMV 35S промотора и заменили ее на последовательность промотора напина, с помощью рестриктаз Ecl136II и Ncol участок промотра напина вырезали из плазмиды pGEM-NAP4 (пример 15) и лигировали в pRT100, расщепленный рестриктазами HindII и NcoI (фигура 5). Чтобы сконструировать pHYB, в котором последовательность промотора напина заменила часть 35S промотора, сохранив энхансерный участок 35S промотора, расположенный до включенной последовательности промотора напина, pGEM-NAP9 (пример 15) расщепляли с помощью рестриктазы XhoI, с помощью фермента Кленова делали тупые концы и расщепляли с помощью рестриктазы NcoI. Полученный фрагмент лигировали в pRT100, расщепленный рестриктазами JcoRV и NcoI. Пример 17. Клонирование генов олеозина и TMV 30К МР, обогащенных His, в экспресионные плазмиды, содержащие промоторы CaMV 35S, напина и химерный промотор. Подробности процедуры конструирования описаны на фигурах 7-12. В начале был получен набор конструкций, в которых гены, представляющие слитые белки, включающие TMV MP, последовательности, обогащенные His, и GFP были помещены под контроль 35S CaMV промотора (фигура 7). Были отбраны последовательности, обогащенные His, имеющие три различные длины (т.е. те, которые содержали два, четыре или шесть повторяющихся наборов последовательности, кодирующей His; пример 11). Ген МР, слитый с двумя наборами последовательности, кодирующей His, был вырезан из рМР-2х с помощью -EcoRI и SalI (фигура 4), ген GFP был вырезан из pRT-GPP в виде XhoI-BamHI-фрагмента, и чтобы получить конструкцию pRT-MP-2x-GFP оба фрагмента ДНК были дотированы в плазмиду pRT101, расщепленную рестриктазами EcoRI и BamHI. Таким же образом, чтобы получить pRT-MP-4x-GFP, ген МР, слитый с четырьмя наборами последовательности, кодирующей His, был вырезан из рМР-4х с помощью EcoRI и SalI (фигура 4), ген GFP был вырезан из pRT-GFP (более подробно см. на фигуре 7) в виде XhoI-BamHI-фратмента, и оба фрагмента ДНК были лигированы в pRT101, расщепленный рестриктазами EcoRI и BamHI. Чтобы получить pRT-MP-6x-GFP, ген МР, слитый с шестью наборами последовательности, кодирующей His, был вырезан из рМР-6х с помощью EcoRI и SalI (фигура 4), ген GPP был вырезан из pRT-GFP в виде XhoI-BamHI-фрягмвта, и оба фрагмента ДНК были лигированы в pRT101, расщепленную рестриктазами EcoRI и BamHI (фигура 7). Чтобы получить эквивалентные конструкции под контролем промотора напина (заменяющего CaMV 35S промотор), были проведены следующие стадии клонирования. N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента; фрагмент, содержащий остаток гена МР, слитый с двумя наборами последовательности, кодирующей His, и ген GFP, вырезали из pRT-MP-2x-GFP с помощью HindIII и XbaI и, чтобы получить pNAP-MP-2x-GFP, оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI. Таким же образом, чтобы получить pNAP-MP-4x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с четырьмя наборами последовательности, кодирующей His, и ген GFP, вырезали из pRT-MP-4x-GFP с помощью HindIII и XbaI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI. И, наконец, чтобы получить pNAP-MP-6x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с шестью наборами последовательности, кодирующей His, и ген GFP, вырезали из pRT-MP-6x-GFP с помощью HindIII и XbaI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI (фигура 8). Аналогичную схему клонирования применяли при получении конструкций, в которых слитые гены MP-His-GFP были помещены под контроль гибридного промотора. N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента; фрагмент, содержащий остаток гена МР, слитый с двумя наборами последовательности, кодирующей His, и ген GFP, вырезали из pRT-MP-2x-GFP с помощью HindIII и XbaI и, чтобы получить pHYB-MP-2x-GFP, оба фрагмента лигировали в pHYB, расщепленный с помощью NcoI и BamHI. Таким же образом, чтобы получить pHYB-MP-4x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с четырьмя наборами последовательности, кодирующей His, и ген GFP, вырезали из pRT-MP-4x-GFP с помощью HindIII и XbaI и оба фрагмента лидировали в pHYB, расщепленный с помощью NcoI и BamHI. Чтобы получить pHYB-MP-6x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с шестью наборами последовательности, кодирующей His, и ген GFP, вырезали из pRT-MP-6x-GFP с помощью HindIII и XbaI и оба фрагмента лигировали в pHYB, расщепленный с помощью NcoI и BamHI. На основе гена олеозина из pOLE4/11 были сконструированы три набора экспрессионных векторов, которые имели промотры CaMV 35S и напина или гибридный промотор, каждый из наборов содержал участки, кодирующие His и имеющие три различные длины (фигура 9). Чтобы сконструировать экспрессионные векторы на основе CaMV 35S промотора, были выполнены следующие процедуры клонирования. Ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей His, слитый с геном GFP, вырезали из pHYB-MP-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pRT100, расщепленный с помощью XhoI и BamHI, в результате получали конструкцию pRT-OLE-2x-GFP. Чтобы получить pRT-OLE-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий четыре набора последовательности, кодирующей His, слитый с геном GFP, вырезали из плазмиды pHYB-MP-4x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pRT100, расщепленный с помощью NcoI и BamHI. Чтобы получить pRT-OLE-6x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий шесть наборов последовательности, кодирующей His, слитый с геном GFP, вырезали из плазмиды pHYB-MP-6x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лидировали в pRT100, расщепленный с помощью NcoI и BamHI (фигура 10). Аналогичную схему клонирования применяли при конструировании экспрессионных векторов на основе промотора напина. Ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей His, слитый с геном GFP, вырезали из pHYB-MP-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pNAP-OLE-2x-GFP. Чтобы получить pNAP-OLE-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий четыре набора последовательности, кодирующей His, слитый с геном GFP, вырезали из pHYB-MP-4x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI. Чтобы получить pNAP-OLE-6x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий шесть наборов последовательности, кодирующей His, слитый с геном GFP, вырезали из pHYB-МР-6х-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI (фигура 11). Сходным образом был сконструирован набор экспрессионных векторов на основе гибридного промотора. Ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей His, слитый с геном GFP, вырезали из pHYB-MP-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pHYB, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pHYB-OLE-2x-GFP. Чтобы получить pHYB-OLE-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий четыре набора последовательности, кодирующей His, слитый с геном GFP, вырезали из pHYB-MP-4x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pHYB, расщепленный с помощью NcoI и BamHI. Чтобы получить pNAP-OLE-6x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий шесть наборов последовательности, кодирующей His, слитый с геном GFP, вырезали из pHYB-MP-6х-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pHYB, расщепленный с помощью NcoI и BamHI (фигура 12). Пример 18. Клонирование генов олеозина и TMV 30К МР, обогащенных Cys/Met, в экспресионные плазмиды, содержащие промоторы CaMV 35S, напина и химерный промотор. Были отбраны последовательности, имеющие три различные длины и обогащенные Cys/Met (с двумя, четырьмя и шестью повторами наборов последовательности, кодирующей Cys/Met, Пример 12). Ген МР, слитый с двумя наборами последовательности, кодирующей Cys/Met, был вырезан из pMP-Cys/Met-2x с помощью EcoRI и SalI, ген GFP был вырезан из pRT-GFP в виде XhoI-BamHI-фрагмента, и чтобы получить конструкцию pRT-MP-Cys/Met-2x-GFP оба фрагмента ДНК были лигированы в pRT101, расщепленный рестриктазами EcoRI и BamHI. Чтобы получить pRT-MP-Cys/Met-4x-GFP, ген МР, слитый с четырьмя наборами последовательности, кодирующей Cys/Met, был вырезан из pMP-Cys/Met-4x с помощью EcoRI и SalI, ген GFP был вырезан из pRT-GFP в виде XhoI-BamHI-фрагмента, и оба фрагмента ДНК были лигированы в pRT101, расщепленный рестриктазами EcoRI и BamHI (фигура 7). Чтобы получить эквивалентные конструкции под контролем промотора напина (заменяющего CaMV 35S промотор), N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде XhoI-BamHI-фрагмента; фрагмент, содержащий остаток гена МР, слитый с двумя наборами последовательности, кодирующей Cys/Met, и ген GFP, вырезали из pRT-MP-2x-GFP с помощью HindIII и XbaI и, чтобы получить pNAP-MP-Cys/Met-2x-GFP, оба фрагмента лигировали в pNAP, расщепленный с помощью NcoI и BamHI. Таким же образом, чтобы получить pNAP-MP-Cys/Met-4x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с четырьмя наборами последовательности, кодирующей Cys/Met, и ген GFP, вырезали из pRT-MP-Cys/Met-2x-GFP с помощью HindIII и XbaI и оба фрагмента лигировали в pNAP, расщепленный с помощью NcoI и BamHI. И, наконец, чтобы получить pNAP-MP-Cys/Met-6x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с шестью наборами последовательности, кодирующей Cys/Met, и ген GFP, вырезали из pRT-MP-Cys/Met-6x-GFP с помощью HindIII и XbaI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI (фигура 8). Аналогичную схему клонирования применяли при получении конструкций, в которых слитые гены MP-Cys/Met-GFP были помещены под контроль гибридного промотора (HYB). N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента; фрагмент, содержащий остаток гена МР, слитый с двумя наборами последовательности, кодирующей Cys/Met, и ген GFP, вырезали из pRT-MP-Cys/Met-2x-GFP с помощью HindIII и XbaI и, чтобы получить pHYB-MP-Cys/Met-2x-GFP, оба фрагмента лигировали в pHYB, расщепленный с помощью NcoI и BamHI. Таким же образом, чтобы получить pHYB-MP-Cys/Met-4x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с четырьмя наборами последовательности, кодирующей Cys/Met, и ген GFP, вырезали из pRT-MP-Cys/Met-2x-GFP с помощью HindIII и XbaI и оба фрагмента лигировали в pHYB, расщепленный с помощью NcoI и BamHI. И наконец, чтобы получить pHYB-MP-Cys/Met-6x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с шестью наборами последовательности, кодирующей Cys/Met, и ген GFP, вырезали из pRT-MP-Cys/Met-6x-GFP с помощью HindIII и XbaI и оба фрагмента лигировали в pHYB, расщепленный с помощью NcoI и BamHI. На основе гена олеозина из pOLE4/11 были сконструированы три набора экспрессионных векторов, которые имели промотры CaMV 35S и напина, или гибридный промотор, каждый из наборов содержал участки, кодирующие Cys/Met, имеющие три различные длины (фигура 9). Чтобы сконструировать экспрессионные векторы на основе CaMV 35S промотора, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из pHYB-MP-Cys/Met-2x-GFP с помощью XhoI и BamI и оба фрагмента ДНК лигировали в pRT100, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pRT-OLE-Cys/Met-2x-GFP. Чтобы получить pRT-OLE-Cys/Met-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмета, участок, содержащий четыре набора последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из плазмиды pHYB-MP-Cys/Met-4x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pRT100, расщепленный с помощью NcoI и BamHI. Чтобы получить pRT-OLE-Cys/Met-6х-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий шесть наборов последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из плазмиды pHYB-MP-Cys/Met-6x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pRT100, расщепленный с помощью NcoI и BamHI (фигура 10). Аналогичным образом, для экспрессионных векторов на основе промотора напина ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из pHYB-MP-Cys/Met-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pNAP-OLE-Cys/Met-2x-GFP. Чтобы получить pNAP-OLE-Cys/Met-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий четыре набора последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из pHYB-MP-Cys/Met-4x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI. Чтобы получить pNAP-OLE-Cys/Met-6x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий шесть наборов последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из pHYB-MP-Cys/Met-6x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI (фигура 11). Сходным образом был сконструирован набор экспрессионных векторов на основе гибридного промотора. Ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из pHYB-MP-Cys/Met-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pHYB, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pHYB-OLE-Cys/Met-2x-GFP. Чтобы получить pHYB-OLE-Cys/Met-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий четыре набора последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из pHYB-MP-Cys/Met-4x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pHYB, расщепленный с помощью NcoI и BamHI. Чтобы получить pNAP-OLE-Cys/Met-6x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий шесть наборов последовательности, кодирующей Cys/Met, слитый с геном GFP, вырезали из pHYB-MP-Cys/Met-6x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лидировали в pHYB, расщепленный с помощью NcoI и BamHI (фигура 12). Пример 19. Клонирование генов TMV 30К МР и олеозина, обогащенных Gly, в экспресионные плазмиды, содержащие промоторы CaMV 35 S, напина и химерный промотор. Были отобраны последовательности, имеющие две различные длины и обогащенные Gly (с двумя и четырьмя повторами наборов последовательности, кодирующей Gly, Пример 13). Ген МР, слитый с двумя наборами последовательности, кодирующей Gly, был вырезан из pMP-Gly-2x с помощью EcoRI и SalI, ген GFP был вырезан из pRT-GFP (подробнее см. на Фигуре 7) в виде XhoI-BamHI-фрагмента, и чтобы получить конструкцию pRT-MP-Gly-2x-GFP оба фрагмента ДНК были лигированы в pRT101, расщепленный рестриктазами EcoRI и BamHI. Чтобы получить pRT-MP-Gly-4x-GFP, ген МР, слитый с четырьмя наборами последовательности, кодирующей Gly, был вырезан из pMP-Gly-4x с помощью EcoRI и SalI, ген GFP был вырезан из pRT-GFP в виде XhoI-BamHI-фрагмента, и оба фрагмента ДНК были лигированы в pRT101, расщепленный рестриктазами EcoRI и BamHI (фигура 7). Чтобы получить эквивалентные конструкции под контролем промотора напина (заменяющего CaMV 35S промотор), NcoI-HindIII-фрагмент гена TMV 30К МР вырезали из pGEM-30К фрагмент, содержащий остаток гена МР, слитый с двумя наборами последовательности, кодирующей Gly, и ген GFP, вырезали из pRT-MP-Gly-2x-GFP с помощью HindIII и XbaI и, чтобы получить pNAP-MP-Gly-2x-GFP, оба фрагмента дотировали в pNAP, расщепленный с помощью NcoI и BamHI. Чтобы получить pNAP-MP-Gly-4x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с четырьмя наборами последовательности, кодирующей Gly, и ген GFP, вырезали из pRT-MP-Gly-2x-GFP с помощью HindIII и XbaI и оба фрагмента лигировали в pNAP, расщепленный с помощью NcoI и BamHI. (фигура 8). Чтобы получить конструкцию гибридного промотора (HYB), N-терминальную часть гена TMV 30К МР вырезали из NcoII-HindIII-фрагмента pGEM-30К и ген GFP вырезали из pRT-MP-Gly-2x-GFP с помощью HindIII и XbaI и, чтобы получить pHYB-MP-Gly-2x-GFP, оба фрагмента лигировали в pHYB, расщепленный с помощью NcoI и BamHI. Чтобы получить pHYB-MP-Gly-4x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindIII-фргатлента, и фрагмент, содержащий остаток гена МР, слитый с двумя наборами последовательности, кодирующей Gly, и ген GFP, вырезали из pRT-MP-Gly-2x-GFP с помощью HindIII и XbaI и оба фрагмента лигировали в pHYB, расщепленный с помощью NcoI и BamHI (фигура 9). На основе гена олеозина из pOLE4/11 были сконструированы три набора экспрессионных векторов, которые имели промотры CaMV 35S и напина или гибридный промотор, каждый из наборов содержал участки, кодирующие Gly, имеющие две различные длины как в Примере 16. Чтобы сконструировать экспрессионные векторы на основе CaMV 35S промотора, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей Gly, слитый с геном GFP, вырезали из pHYB-MP-Gly-2x-GFP с помощью XhoI и BamHl и оба фрагмента ДНК лигировали в pRT100, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pRT-OLE-Gly-2x-GFP. Чтобы получить pRT-OLE-Gly-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий четыре набора последовательности, кодирующей Gly, слитый с геном GFP, вырезали из pHYB-MP-Gly-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК дотировали в pRT100, расщепленный с помощью NcoI и BamHI (фигура 10). Аналогичным образом, для экспрессионных векторов на основе промотора напина ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей Gly, слитый с геном GFP, вырезали из pHYB-MP-Gly-2x-GFP с помощью Xhol и BamHl и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pNAP-OLE-Gly-2x-GFP. Чтобы получить pNAP-OLE-Gly-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-partAGwa, участок, содержащий четыре набора последовательности, кодирующей Gly, слитый с геном GFP, вырезали из плазмиды pHYB-MP-Gly-4x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pNAP, расщепленный с помощью NcoI и BamHI (фигура 11). Сходным образом был сконструирован набор экспрессионных векторов на основе гибридного промотора. Ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей Gly, слитый с геном GFP, вырезали из pHYB-MP-Gly-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pHYB, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pHYB-OLE-Gly-2x-GFP. Чтобы получить pHYB-OLE-Gly-4x-GFP, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий четыре набора последовательности, кодирующей Gly, слитый с геном GFP, вырезали из pHYB-MP-Gly-4x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pHYB, расщепленный с помощью NcoI и BamHI (фигура 12). Пример 20. Клонирование генов TMV 30К МР и олеозина, обогащенных лизином, в экспресионные плазмиды, содержащие промоторы CaMV 35S, напина и химерный промотор. Были отбраны последовательности, имеющие две различные длины и обогащенные лизином (с двумя и четырьмя повторами наборов последовательности, кодирующей Lys, пример 13). Ген МР, слитый с двумя наборами последовательности, кодирующей Lys, был вырезан из pMP-Lys-2x с помощью EcoRI и SalI, ген GFP был вырезан из pRT-GFP в виде XhoI-BamHI-фрагмента, и чтобы получить конструкцию pRT-MP-Lys-2x-GFP оба фрагмента ДНК были лигированы в pRT101, расщепленный рестриктазами EcoRI и BamHI. Чтобы получить pRT-MP-Lys-4x-GPP, как в примерах 17-19 и на фигуре 7, ген МР, слитый с четырьмя наборами последовательности, кодирующей Lys, был вырезан из рМР-Lys-4x с помощью EcoRI и SalI, ген GFP был вырезан из pRT-GFP в виде XhoI-ВатШ-фрагмента и оба фрагмента ДНК были лигированы в pRT101, расщепленный рестриктазами EcoRI и BamHI. Чтобы получить эквивалентные конструкции под контролем промотора напина и гибридного промотора (HYB) (заменяющих CaMV 35S промотор), NcoI-HindIII-psruem гена TMV 30К МР вырезали из pGEM-30К фрагмент, содержащий остаток гена МР, слитый с двумя наборами последовательности, кодирующей Lys, и ген GFP вырезали из pRT-MP-Lys-2x-GFP с помощью HinaIII и XbaI и, чтобы получить pNAP-MP-Lys-2x-GFP и pHYB-MP-Lys-2x-GFP, оба фрагмента лигировали в pNAP и pHYB, расщепленные с помощью NcoI и BamHI. Чтобы получить pNAP-MP-Lys-4x-GFP и pHYB-MP-Lys-4x-GFP, N-терминальную часть гена TMV 30К МР вырезали из pGEM-30К в виде NcoI-HindHI-фрагмента, и фрагмент, содержащий остаток гена МР, слитый с четырьмя наборами последовательности, кодирующей Lys, и ген GFP, вырезали из pRT-MP-Lys-2x-GPP с помощью HindIII и XbaI и оба фрагмента лигировали в pNAP и pHYB, расщепленные с помощью NcoI и BamHI (фигуры 8 и 9). На основе гена олеозина из pOLE4/11 были сконструированы три набора экспрессионных векторов, которые имели промотры CaMV 35S и напина или гибридный промотор, каждый из наборов содержал участки, кодирующие Lys, имеющие две различные длины как в примере 19. Чтобы сконструировать экспрессионные векторы на основе CaMV 35S промотора, промотора напина и гибридного промотора, ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий два набора последовательности, кодирующей Lys, слитый с геном GFP, вырезали из pHYB-MP-Lys-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pRT100, расщепленный с помощью NcoI и BamHI, в результате получали конструкцию pRT-OLE-Lys-2x-GFP, pNAP-OLE-Lys-2x-GFP и pHYB-OLE-Lys-2x-GFP. Чтобы получить pRT-OLE-Lys-4x-GFP, pNAP-OLE-Lys-4x-GFP и pHYB-OLE-Lys-4x-GFP ген олеозина вырезали из pOLE4/11 в виде NcoI-XhoI-фрагмента, участок, содержащий четыре набора последовательности, кодирующей Lys, слитый с геном GFP, вырезали из pHYB-MP-Lys-2x-GFP с помощью XhoI и BamHI и оба фрагмента ДНК лигировали в pRT100, расщепленный с помощью NcoI и BamHI. Пример 21. Переходные экспрессионные системы. Чтобы убедиться в экспрессии белка, в том, что экспрессированные белки обладали той же интактной биологической функцией (функциями), что и нативные немодифицированные белки, и в том, что белки аккумулировались в органе-мишени, был использован анализ переходной экспрессии. Этот анализ был облегчен визуальным наблюдением за репортерным белком. В особенности полезной оказалась конфокальная лазерная сканирующая микроскопия, так как она позволяла на ранней стадии детектировать трансгенные растения, экспрессирующие белки, обогащенные аминокислотами, гены которых объединены в рамке считывания с нуклеотидной последовательностью, кодирующей репортерный белок. Пример 22. Тестирование активности напина и химерных промоторов с использованием переходной экспрессии гена GFP в клетках растений, трансформированных с помощью бомбардировки частицами. Чтобы проанализировать уровни относительной транскрипции, обеспеченной промотором напина в pNAP, гибридным промотором в HYB и промотором CaMV 35S в pRT100, в эти плазмиды был клонирован участок, кодирующий репортерный белок GFP. Для конструирования клонов, содержащих GFP, был использован ген мутанта GFP (S65T), обладающего сдвигом в красной области спектра. Участок, кодирующий GFP, был клонирован в виде NcoI-BamHI-фрагмента в расщепленные сходным образом pRT100, pNAP и pHYB. Эпидермальные клетки Nicotiana benthamiana, временно трансформированные бомбардировкой частицами (пример 8) с помощью pHYB-GFP (содержащей участок, кодирующий GFP под контролем гибридного промотора), отчетливо экспрессировали GFP. Наблюдаемые уровни экспрессии GFP были сравнимы с контролем pRT-GFP, в котором экспрессия GFP происходила под контролем промотора CaMV 35S. Эти эксперименты указывали на то, что экспрессионная кассета pHYB-GFP была полностью функциональной в живых клетках растений (фигура 6 показывает pRT-OLE-4x-GFP и pHYB-OLE-4x-GFP). pNAP-GFP экспрессировал в зародышах семян на ранних стадиях и было обнаружено, что такие зародыши флуоресцировали. Пример 23. Клонирование экспрессионных кассет в бинарные векторы и трансформация соответствующих бинарных векторов в Agrobacterium. Перед трансформацией в Agrobacterium из производных pRT в сайт HindIII Т-ДНК растительного трансформирующего бинарного вектора pBin19 были клонированы находящиеся под контролем промотора CaMV 35S, промотора напина и гибридного промотора кассеты генов, содержащие меченый GFP и обогащенные гистидином, цистеин/метионином, глицином и лизином гены олеозина (ole) и TMV МР. Agrobacterium tumefaciens AGLI, несущие гипервирулентную плазмиду pTi Во542 (Lazo et al., 1991. Biotechnology 3, 963-967), ослабленную в отношении индукции опухолей, были трансформированы с помощью модифицированного метода замораживания-оттаивания Holsters (Holsters et al., 1978. Mol. Gen. Genet. 163, 181-187). Agrobacteria культивировали на чашках с LB-агаром с добавлением 0,5 г/л MgSO4 в течение 2 дней при 26°С с последующим выращиванием до середины экспоненциальной фазы на жидкой среде 2xYT, содержавшей 50 мкг/мл рифампицина; бактерии затем промывали 150 мМ Nad и перед трансформацией ресуспендировали в 20 мМ CaCl2 при 1010 КОЕ/мл. К аликвотам клеточной суспензии добавляли глицерин до конечной концентрации 15% и клетки замораживали в жидком азоте. После добавления к клеткам 5 мкг ДНК клетки оттаивали и инкубировали на льду в течение 15 минут, а перед тепловым шоком – при 37°С в течение 5 минут. Перед трансформацией клетки разводили свежей средой 2xYT и культивировали 2 часа при 28°С при энергичной аэрации, затем переносили в чашки с 1%-ным агаром, содержащие 50 мкг/мл рифампицина и 100 мкг/мл канамицина. Колонии, устойчивые к канамицину, отбирали с помощью ПЦР-анализа на присутствие/отсутствие рекомбинантных конструкций. Пример 24. Трансформация Brassica campestris, опосредованная бактериями Agrobacterium. Трансформацию растений Brassica campestris проводили в полном соответствии с процедурой (Moloney et al., Plant Cell Reports 8, 238-272, 1989). Семядоли культивировали совместно с подходящими клеточными суспензиями Agrobacterium в течение 2-3 дней на 1%-ном агаре со Средой I (Murashige Minimal Organics (MMO) с 3% сахарозы, 4 мг/л ВАР, 0,7% фитоагара) при 22°С. Перед переносом в чашки Петри, содержащие Среду II (MMO с 4 мг/л ВАР, 3% сахарозы, 300 мг/л тикарциллина (DUCHEFA) и 0,7% фитоагара) семядоли ополаскивали три раза в дистиллированной воде. Через 7-10 дней экспланты переносили в Среду III (MMO с 4 мг/л ВАР, 3% сахарозы, 300 мг/л тикарциллина, 25 мг/л канамицина и 0,7% фитоагара). После дополнительной инкубации в течение 2-3 недель экспланты с зелеными наплывами или незрелые зеленые побеги переносили в свежую Среду III. Для образования побегов наплывы или незрелые зеленые побеги переносили в Среду IV (MMO с 3% сахарозы, 300 мг/л тикарциллина, 25 мг/л канамицина и 0,7% фитоагара). Полностью сформированные побеги переносили в Среду V (MMO с 0,2 мг/л IBA, 3% сахарозы, 300 мг/л тикарциллина и 0,7% фитоагара) для прорастания корней. После формирования корневой системы побеги с корнями вынимали из агара и переносили в горшок с влажной землей и растили с фотопериодом 16 часов на свету/8 часов в темноте при интенсивности света 12,1 мкмоль/м2·сек при 22°С. Пример 25. Предупреждение выключения генов в Brassica campestris. При использовании нуклеотидной последовательности, кодирующей Ole, которая произошла из В. campestris, существует риск посттранскрипционного выключения генов. В настоящем изобретении при трансформировании В. campestris геном Ole риск посттранскрипционного выключения генов в трансгенном растении, происходящего из-за гомологии нуклеотидной последовательности между трансгенным и эндогенным генами, можно успешно избежать, если использовать ген Ole из Arabidopsis thaliana, но не ген из В. campestris. Длина и степень гомологии одного гена другому гену влияет на выключение гена. Если вводимый ген на 100% гомологичен эндогенному гену, выключение гена вероятно. Сравнение олеозина из Brassica с олеозином из Arabidopsis (теперь в базе данных NCBI) показало, что гомология последовательностей на уровне ДНК колеблется в пределах 74-87%. Таким образом, степень гомологии на уровне ДНК отчетливо различается; это позволило получить трансгенные растения, у которых выключение генов не происходило. Было отобрано третье поколение растений, которые обладали стабильной экспрессией 4xHis-кассеты (исходя из данных детекции с помощью антител и дотблоттинга). Пример 26. Экспрессия конструкций с гистидиновыми, цистеин/метиониновыми, глициновыми и лизиновыми вставками различной длины в эпидермальных клетках табака. Было предположено, что по двум причинам, включающим стабильность и внутренние ограничения продукционной емкости клеток, длина спроектированных полиаминокислотных последовательностей имеет решающее значение для экспрессии His-, Cys/Met-, Gly- и Lys-конструкций. Длинные полиаминокислотные последовательности, например в SxHis-серии, были нестабильны в системе трансляции in vitro (TNT® T7/SP6 Coupled Wheat Germ Extract System L5030, Promega corporation), a 6xHis-серии ограничивали рост Е. coli в жидкой среде, как показано в примере 11. Также решающее значение для экспрессии имели внутренние ограничения возможности клеток продуцировать большие количества полиаминокислот без влияния на другие белки и на их синтез. Поэтому все конструкции, контролируемые промотором CaMV 35S, были протестированы in planta для проверки ограниченности применения больших полиаминокислотных последовательностей в клетках растений. Несмотря на ожидаемые ограничения, в эпидермальных клетках табака экспрессировались все конструкции (приготовленные, как описано в примерах 11-14). Однако конструкции, содержавшие более длинные полиаминокислотные последовательности, экспрессировались относительно хуже, чем более короткие полиаминокислотные последовательности. Поэтому для экспериментов по стабильной трансформации были выбраны конструкции с четырьмя His-, Cys/Met, Gly и Lys-кассетами. Вестерн-блоттинг анализ нескольких линий растений (таблица 1) показал, что антитела к гистидиновым последовательностям, МР, олеозину или GFP (фигуры 13-15) могли бы использоваться для обнаружения белков с известными молекулярными массами (MWs). Расчетные MWs олеозина, МР, GFP и 4xHis-кассеты (с 14 His-кодонами в каждой кассете) составляют 18,5, 30, 30 и 2,5 кДа, соответственно. Соответствующие MWs для слитых белков можно рассчитать из этих значений как общую сумму MWs каждого компонента слитого белка. Исследование методом электрофореза в ПААГ в присутствии SDS и методом вестерн-блоттинг анализа было показано, что эти слитые белки идентичны по размеру их расчетной молекулярной массе. Стабильность трансгенно-кодируемых белковых продуктов (олеозин-His и MP-His) была исследована методом вестерн-блоттинг анализа. Анализ генераций (самоопыляемых) растений (3-е поколение) после трансформации (таблица 1) четко показал, что трансгенная экспрессия была стабильна и в последующих поколениях. Эти анализы также показали, что количество слитого белкового продукта было относительно постоянным между различными поколениями растений и отражало уровень приобретенной стабильной экспрессии по отношению к другим белкам растений. Пример 27. Продукция /и planta слитых белков олеозин-гистидин и МР-гистидин. Были проанализированы четыре серии трансгенных линий Brassica campestris (коды: 5, 8, 14 и 17). Эти серии содержали следующие конструкции: 5-м серия: pNAP-MP-4x-His-GFP; 8-ая серия: pHYB-MP-4x-His-GFP; 14-м серия: pNAP-OLE-4x-His-GFP; 17-м серия: pHYB-OLE-4xHis-GFP. Вестерн-блоттинг анализ (Sambrook et al.. Molecular cloning: A laboratory manual, 2nd edn., Cold Spring Harbor, NY 1989) нескольких линий растений (таблица 1) показал, что антитела к последовательности His, MP, олеозину или GFP (фигуры 13-15) можно было использовать для обнаружения белков с известными молекулярными массами (MWs). Антитела к MP, олеозину и GFP были получены в лаборатории Атабекова. Антитела к полигистидину были коммерческим препаратом (Pierce, Rocford US). Ожидаемые MWs олеозина, MP, GFP и 4xHis-KacceTbi (с 14 His-кодонами в каждой кассете) были 18,5, 30, 30 и 2,5 кДа, соответственно. Соответствующие MWs для слитых белков можно рассчитать из этих значений как общую сумму MWs каждого компонента слитого белка, таким образом, MW OLE-4xHis-GFP составляет 62,5. При исследовании методом электрофореза в ПААГ в присутствии SDS и методом вестерн-блоттинг анализа было показано, что эти слитые белки OLE-4xHis-GFP и МР-4xHis-GFP идентичны по размеру их расчетной молекулярной массе. Стабильность трансгенно-кодируемых белковых продуктов (олеозин-His и MP-His) была исследована методом вестерн-блоттинг анализа. Анализ генераций (самоопыляемых) растений (3-е поколение) после трансформации (таблица 1) четко показал, что в последующих поколениях трансгенная экспрессия была стабильной. Более того, вестерн-блоттинг анализ исследованных в дальнейшем трансформированных растений обнаружил, что размер белкового продукта остается постоянным независимо от поколения. Эти анализы также показали, что количество слитого белкового продукта было относительно постоянным среди генераций растений и отражало уровень приобретенной стабильной экспрессии по отношению к другим белкам растений. При использовании антител к His, МР и OLE оба белка, МР или OLE, и GPP были обнаружены в соответствующих линиях растений. Это показывает, что экспрессия была стабильной и маловероятно, что, если носитель присутствует и экспрессируется, то последовательности His будут вырождаться. Таблица 1. Ответ трансгенных линий растения Brassica campestris на добавление специфических антител к белковым продуктам, кодируемых соответствующими трансгенами. Поликлональные антитела к OLE, выработанные к С-терминальной части белка, и поликлональные антитела к МР, продуцированному бактериями, были получены в лаборатории Атабекова. Антитела к His были коммерческим препаратом, доступным в фирме Pierce, Rocford US.
ND – не определяется; + – слабый ответ; ++ – хороший ответ; +++ – сильный ответ. Линии трансгенных растений с 5-ой по 14-ую серию содержали промотор NAP, а линии трансгенных растений 08-ой по 17-ую серию содержали промотор HYB. Пример 28. Аминокислотный анализ семян из линии трансгенных растений Brassica campestris, экспрессирующих четыре гистидиновые кассеты. Продублированные образцы семян, предназначенные для аминокислотного анализа, получали из трансгенных линий Brassica (T2), экспрессирующих четыре гистидиновые кассеты (линии 5-8 в качестве носителя имели МР, линии 14-17 в качестве носителя имели олеозин; см. фигуру 3 и пример 27). Аминокислотный анализ проводили в тотальном образце белка, полученного из семян (таблица 2). Анализ выполняли в Подразделении питания животных в Сельскохозяйственном исследовательском центре, Джокионен, Финляндия, в соответствии с Директивами европейской комиссии 98/64/ЕС (1998). Таблица 2. Семена, использованные для аминокислотного анализа.
WT – дикий тип. Линия трансгенных растений Brassica campestris 14.12Ф10 содержала на 30% гистидина больше, чем растения дикого типа (таблица 3). Повышенное содержание гистидина происходило не за счет других аминокислот, но суммарное содержание белка различалось. Таблица 3. Относительное увеличение содержания гистидина в трансгенных семенах В. campestris
WT – дикий тип. Увеличение содержания гистидина составляло 3-9%, при этом увеличение содержания других аминокислот было исключено.
Формула изобретения
1. Способ повышения содержания одной или более выбранных аминокислот в выбранной ткани или органе растения, включающий трансформацию растения с помощью, по меньшей мере, одной конструкции с рекомбинантной нуклеотидной последовательностью, которая включает ткане- или органоспецифичные регуляторные последовательности, контролирующие транскрипцию во время выбранных стадий морфогенеза; указанные регуляторные последовательности эффективно присоединены к химерной нуклеотидной последовательности, обеспечивающей направленную экспрессию растительного специфического белка-носителя, имеющего стабильное полиаминокислотное удлинение цепи, где указанная химерная нуклеотидная последовательность представляет собой последовательность нуклеотидов, кодирующую белок-носитель, которая не имеет своего собственного терминирующего кодона и содержит на своем С-терминальном конце стабильно присоединенные кодоны, кодирующие выбранное количество и комбинацию аминокислотных остатков, и где стабильность конструкции, кодирующей указанный белок-носитель, имеющий полиаминокислотное удлинение цепи, определяют с помощью бесклеточной системы трансляции in vitro (IVT) с применением кодоновых кассет, имеющих выбранную комбинацию и количество кодонов. 2. Способ по п.1, в котором стабильность, направленную экспрессию и интактные функции белка-носителя, имеющего полиаминокислотное удлинение цепи, определяют с помощью детектируемого репортерного гена. 3. Способ по п.1, в котором конструкцию с рекомбинантной нуклеотидной последовательностью, обеспечивающую направленную экспрессию белка-носителя, имеющего полиаминокислотное удлинение цепи, отбирают с помощью способа, включающего следующие стадии: (а) слияние конструкции с рекомбинантной нуклеотидной последовательностью, кодирующей репортерный белок, где указанная конструкция включает ткане- или органоспецифичные регуляторные последовательности, контролирующие транскрипцию во время выбранных стадий морфогенеза; указанные регуляторные последовательности эффективно присоединены к химерной нуклеотидной последовательности, обеспечивающей направленную экспрессию белка-носителя, имеющего полиаминокислотное удлинение цепи, где указанная химерная нуклеотидная последовательность представляет собой последовательность нуклеотидов, кодирующую белок-носитель, которая содержит на своем С-терминальном конце стабильно присоединенные кодоны, кодирующие выбранную комбинацию и количество аминокислотных остатков; (б) селекцию в бесклеточной системе трансляции (IVT) конструкции, включающей такое количество кодонов, которое обеспечивает стабильную трансляцию полиаминокислотного удлинения цепи; (в) введение конструкции, полученной на стадии (б) в клетку растения; (г) селекцию с помощью анализа транзитной экспрессии конструкций, обеспечивающих направленную экспрессию белка-носителя, имеющего полиаминокислотное удлинение цепи, с помощью детекции экспрессии репортерного белка в растительной клетке; (д) удаление нуклеотидной последовательности, кодирующей репортерный ген из конструкций, отобранных на стадии (г) для получения конструкции, не имеющей нуклеотидной последовательности, кодирующей репортерный белок; и (е) трансформацию растения с помощью конструкции, полученной на стадии (д). 4. Способ по п.3, в котором конструкцию, полученную на стадии (б) и включающую нуклеотидную последовательность, кодирующую репортерный белок, вводят в клетку растения с помощью метода микропроекционной бомбардировки. 5. Способ по п.3, в котором отбирают конструкцию, обеспечивающую стабильную направленную экспрессию репортерного белка, и которую в дальнейшем, после удаления нуклеотидной последовательности, кодирующей репортерный белок, вводят в растения с помощью трансформации, опосредованной Agrobacterium. 6. Способ по п.1, в котором кодоны, кодирующие полиаминокислотное удлинение цепи, включают в себя кодоны, кодирующие гистидин, цистеин, метионин, глицин, лизин, триптофан, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, тирозин, серин, треонин, аргинин, аспарагиновую кислоту, глутаминовую кислоту, аспарагин, глутамин или любые их комбинации. 7. Способ по п.1, в котором выбранное количество кодонов составляет от шести до восьмидесяти. 8. Способ по п.1, в котором выбранная ткань или орган растения являются семенами. 9. Способ по п.1, в котором выбранная ткань или орган растения являются клеточной стенкой или клеточной мембраной. 10. Способ по п.1, в котором выбранная ткань или орган растения являются масляным тельцем. 11. Способ по п.1, в котором специфическим белком-носителем растения является белок, функционирующий во внутриклеточном транспортном пути растения. 12. Способ по п.11, в котором белок-носитель является белком клеточной стенки или растительным вирусным белком. 13. Способ по п.12, в котором белок-носитель включает в себя олеозин, калеозин, стеролеозин, круциферин, напин или растительный вирусный транспортный белок. 14. Способ по п.13, в котором белок-носитель является транспортным белком вируса табачной мозаики (TMV МР). 15. Способ по п.1, в котором регуляторная последовательность является промотором, экспрессируемым во время эмбриогенеза. 16. Способ по п.1, в котором регуляторная последовательность включает в себя промоторы напина (NAP), 35S гена вируса мозаики цветной капусты (CaMV 35S), химерного гибрида (HYB), 198, нопалина, фазеолина, стеролеозина, калеозина, круциферина, альфа-альфа вируса мозаики (AMV), промотор теплового шока, промоторы 2S альбумина и олеозина. 17. Способ по п.16, в котором промотор напина (NAP) является промотором напина (NAP) из Arabidopsis thaliana. 18. Способ по п.16, в котором гибридный промотор (HYB) является химерным промотором, включающим энхансерную последовательность промотора (CaMV 35S) и собственно промотор напина (NAP). 19. Способ по п.1, в котором стабильную направленную экспрессию белка-носителя, имеющего полиаминокислотное удлинение цепи, обеспечивают с помощью экспрессии транспортного белка под контролем промотора напина (NAP). 20. Способ по п.3, в котором репортерный белок представляет собой нуклеотидную последовательность, кодирующую детектируемый белок. 21. Способ по п.3, в котором репортерный белок является флуоресцентным белком. 22. Способ по п.3, в котором репортерный белок является зеленым флуоресцентным белком (GFP), красным флуоресцентным белком, 23. Конструкция с рекомбинантной нуклеотидной последовательностью для повышения содержания одной или более выбранных аминокислот в выбранной ткани или органе растения, где конструкция, обеспечивающая направленную экспрессию в выбранной ткани или органе растения белка-носителя, имеющего стабильное полиаминокислотное удлинение цепи, включает в себя ткане- или органоспецифичные регуляторные последовательности, контролирующие транскрипцию во время выбранных стадий морфогенеза; указанные регуляторные последовательности эффективно присоединены к химерной нуклеотидной последовательности, обеспечивающей направленную экспрессию специфического растительного белка-носителя, имеющего полиаминокислотное удлинение цепи; указанная химерная нуклеотидная последовательность представляет собой последовательность нуклеотидов, кодирующую белок-носитель, которая не содержит терминирующий кодон и имеет на своем С-терминальном конце стабильно присоединенные кодоны, кодирующие выбранное количество и комбинацию аминокислотных остатков, и где стабильность конструкции, кодирующей указанный белок-носитель, имеющий полиаминокислотное удлинение цепи, определяют с помощью бесклеточной системы трансляции in vitro (IVT) с применением кодоновых кассет, имеющих выбранную комбинацию и количество кодонов. 24. Конструкция по п.23, в котором конструкция, обеспечивающая селекцию конструкции, обеспечивающей направленную экспрессию белка-носителя, имеющего стабильное полиаминокислотное удлинение в цепи и обладающего интактными биологическими функциями, представляет собой конструкцию, в которой нуклеотидная последовательность, кодирующая репортерный белок, присоединена к С-терминальному концу указанной химерной нуклеотидной последовательности. 25. Конструкция по п.23, в котором специфический растительный белок-носитель является белком, функционирующим в секреторном внутриклеточном транспортном пути. 26. Конструкция по п.23, в котором белок-носитель является белком растительной клеточной стенки или растительным вирусным белком. 27. Конструкция по п.23, в котором белок-носитель является олеозином, калеозином, стеролеозином, круциферином, напином или растительным вирусным транспортным белком. 28. Конструкция по п.23, в котором белок-носитель является транспортным белком вируса табачной мозаики (TMV МР). 29. Конструкция по п.23, в котором кодоны включают в себя кодоны, кодирующие гистидин, цистеин, метионин, глицин, лизин, триптофан, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, тирозин, серии, треонин, аргинин, аспарагиновую кислоту, глутаминовую кислоту, аспарагин, глутамин или любую их комбинацию. 30. Конструкция по п.23, в котором количество кодонов, обеспечивающих стабильную трансляцию и экспрессию желаемого белка, необязательно составляет число от шести до восьмидесяти. 31. Конструкция по п.23, в котором регуляторная последовательность представляет собой промотор, экспрессируемый во время эмбриогенеза. 32. Конструкция по п.23, в котором регуляторная последовательность включает в себя промоторы напина (NAP), вируса мозаики цветной капусты (CaMV 35S), химерного гибрида (HYB), 19S, нопалина, фазеолина, стеролеозина, калеозина, круциферина, альфа-альфа вируса мозаики (AMV), промотор теплового шока, промоторы 2S альбумина и олеозина. 33. Конструкция по п.32, в котором промотор напина (NAP) является промотором напина (NAP) из Arabidopsis thaliana. 34. Конструкция по п.32, в котором химерный гибридный промотор (HYB) включает энхансерную последовательность промотора CaMV 35S и собственно промотор напина (NAP). 35. Конструкция по п.24, в котором репортерный белок представляет собой детектируемый белок. 36. Конструкция по п.24, в котором репортерный белок является флуоресцентным белком. 37. Конструкция по п.24, в котором репортерный белок является зеленым флуоресцентным белком (GFP), красным флуоресцентным белком, 38. Конструкция по п.24, в котором конструкция включает в себя нуклеотидную последовательность, кодирующую белок-носитель, имеющий удлинение цепи, обогащенное гистидином, и зеленый флуоресцентный белок (GFP). 39. Способ по п.1 для создания композиции, включающей в растительный материал стабильный белок-носитель, обогащенный полиаминокислотами, имеющий полиаминокислотное удлинение цепи, где содержание аминокислот в растительном материале, полученном с помощью предложенного способа, по сравнению с содержанием аминокислот в соответствующем немодифицированном растении дикого типа составляет, по меньшей мере, 2:1. 40. Способ по п.39 для создания пищи, обогащенной аминокислотами, из жмыха, получаемого после отжима масла из растений. 41. Конструкция по п.23 для создания композиции, включающей в растительный материал белок-носитель, обогащенный аминокислотами, имеющий полиаминокислотное удлинение цепи, где содержание аминокислот в растительном материале, полученном с помощью указанного способа, по сравнению с содержанием аминокислот в соответствующем немодифицированном растении дикого типа составляет, по меньшей мере, 2:1. 42. Растение, трансформированное конструкцией по п.23, указанное растение обладает повышенным содержанием одной или более аминокислот в выбранной ткани или органе растения. 43. Растение, трансформированное конструкцией по п.2, указанное растение обладает повышенным содержанием одной или более аминокислот в выбранной ткани или органе растения. 44. Клетка растения, трансформированная конструкцией по п.23, указанная клетка обладает повышенным содержанием одной или более аминокислот в выбранной ткани или органе растения. 45. Клетка растения, трансформированная конструкцией по п.24, указанная клетка обладает повышенным содержанием одной или более аминокислот в выбранной ткани или органе растения. 46. Рекомбинантная линия клеток растения, полученная из растения, трансформированного конструкцией по п.23, указанная клеточная линия обладает повышенным содержанием одной или более аминокислот в выбранной ткани или органе растения. 47. Рекомбинантная линия клеток растения, полученная из растения, трансформированного конструкцией по п.24, указанная клеточная линия обладает повышенным содержанием одной или более аминокислот в выбранной ткани или органе растения. 48. Пищевая композиция, обогащенная аминокислотами, включающая растительный материал, включающий белок-носитель, обогащенный аминокислотами, имеющий стабильное полиаминокислотное удлинение цепи, полученный с помощью способа по п.1, в которой содержание аминокислот в указанном растительном материале, по сравнению с содержанием аминокислот в соответствующем немодифицированном растительном материале, полученном из растения дикого типа, составляет, по меньшей мере, 2:1. 49. Кормовая композиция, обогащенная аминокислотами, включающая растительный материал, включающий белок-носитель, обогащенный аминокислотами, имеющий стабильное полиаминокислотное удлинение цепи, полученный с помощью способа по п.1, в которой содержание аминокислот в указанном растительном материале, по сравнению с содержанием аминокислот в соответствующем немодифицированном растительном материале, полученном из растения дикого типа, составляет, по меньшей мере, 2:1.
РИСУНКИ
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 -спиральная структура, содержащая 31% лизина и 20% метионина (СР3-5). Под контролем промотора фазеолина или промотора
-спиральная структура, содержащая 31% лизина и 20% метионина (СР3-5). Под контролем промотора фазеолина или промотора  -конглицинина в семенах трансгенных растений табака накапливалось значительные количества синтетического белка.
-конглицинина в семенах трансгенных растений табака накапливалось значительные количества синтетического белка. (Bio-Rad), с денатурацией при 95°С в течение 1 минуты, отжигом праймера при 65°С в течение 1,5 минуты, с удлинением праймера при 72°С в течение 2 минут и заключительной стадией элонгации после 30 циклов при 72°С в течение 10 минут. Продукт ПЦР ожидаемых размеров (776 п.н.) изолировали из 1%-ного агарозного геля после электрофореза конечной реакционной ПЦР-смеси, очищали с помощью набора для извлечения ДНК из геля (Qiagen) и затем перед клонированием обрабатывали 1 единицей ДНК-полимеразы Т4 в 1x-реакционном буфере для ДНК-полимеразы Т4 (50 мМ NaCI, 10 мМ трис-HCl; рН 7,9, 10 мМ MgCl2, 1 мМ дитиотреитол), содержащем 100 мМ раствор всех dNTP, при 14°С в течение 15 минут.
(Bio-Rad), с денатурацией при 95°С в течение 1 минуты, отжигом праймера при 65°С в течение 1,5 минуты, с удлинением праймера при 72°С в течение 2 минут и заключительной стадией элонгации после 30 циклов при 72°С в течение 10 минут. Продукт ПЦР ожидаемых размеров (776 п.н.) изолировали из 1%-ного агарозного геля после электрофореза конечной реакционной ПЦР-смеси, очищали с помощью набора для извлечения ДНК из геля (Qiagen) и затем перед клонированием обрабатывали 1 единицей ДНК-полимеразы Т4 в 1x-реакционном буфере для ДНК-полимеразы Т4 (50 мМ NaCI, 10 мМ трис-HCl; рН 7,9, 10 мМ MgCl2, 1 мМ дитиотреитол), содержащем 100 мМ раствор всех dNTP, при 14°С в течение 15 минут.