Патент на изобретение №2334793
|
||||||||||||||||||||||||||
(54) СПОСОБ ПОЛУЧЕНИЯ КУЛЬТУРЫ КЛЕТОК С ИНДУЦИРОВАННЫМ АНГИОГЕННЫМ ФЕНОТИПОМ, ПРЕДНАЗНАЧЕННОЙ ДЛЯ ТКАНЕВОЙ ИНЖЕНЕРИИ В ЗОНЕ ИШЕМИИ
(57) Реферат:
Изобретение относится к области биотехнологии, в частности к генной и клеточной инженерии, и может быть использовано в медицине. Предложен способ получения культуры клеток для тканевой инженерии в зоне ишемии, заключающийся в со-культивировании in vitro стромальных клеток из подкожной жировой клетчатки (СКЖТ) с клетками либо тотальной, либо обогащенной CD31-или NG2-положительными клетками кардиомиоцитарной фракции (КМЦ), которое осуществляют при соотношении СКЖТ/КМЦ от 1:4 до 4:1 до появления в культуре микроскопически определяемых капилляро-подобных структур. Получаемая предложенным способом культура клеток с индуцированным в процессе со-культивирования ангиогенным фенотипом обеспечивает эффективную локальную активацию ангиогенеза при введении в ишемизированную ткань. 3 з.п. ф-лы, 11 ил.
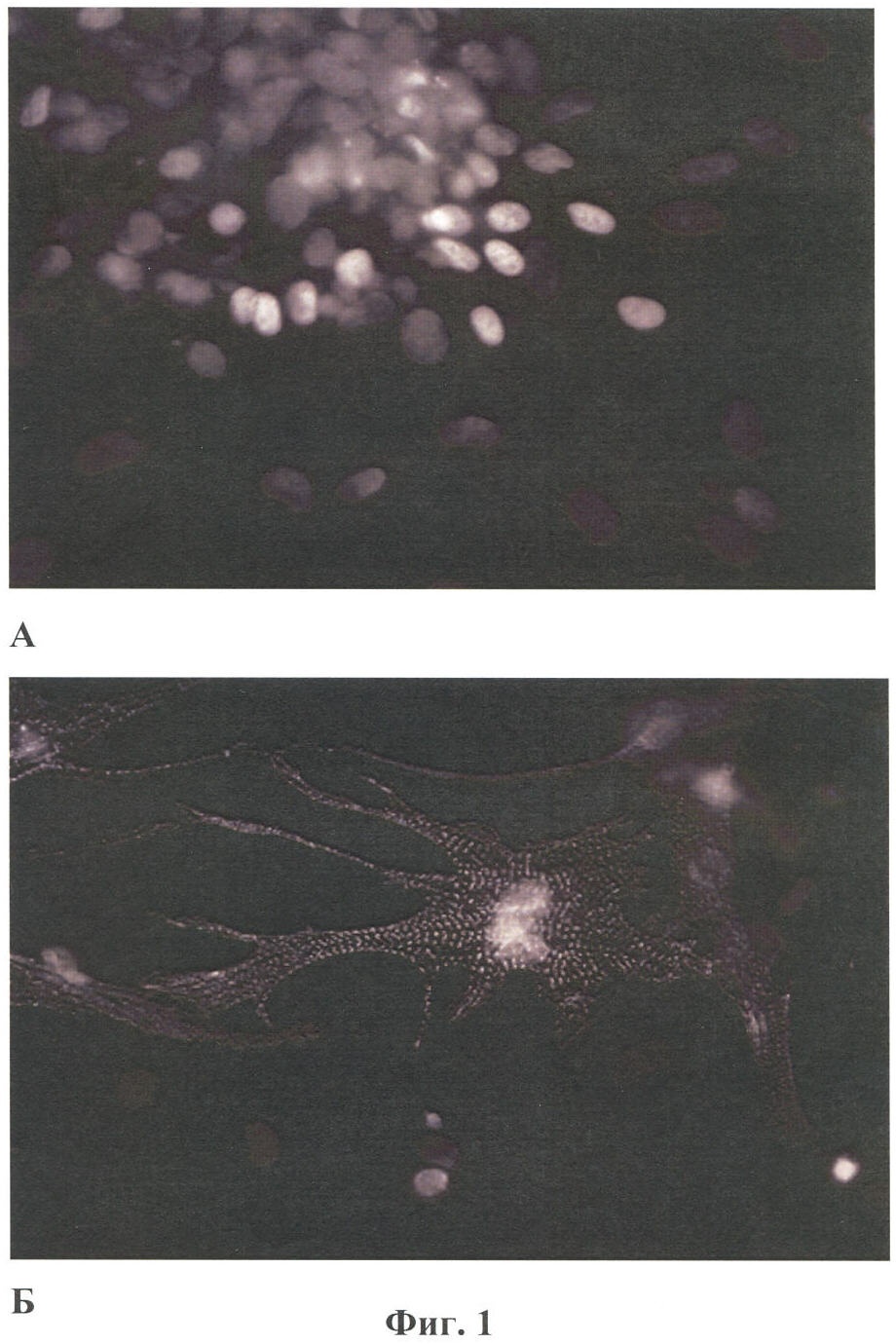
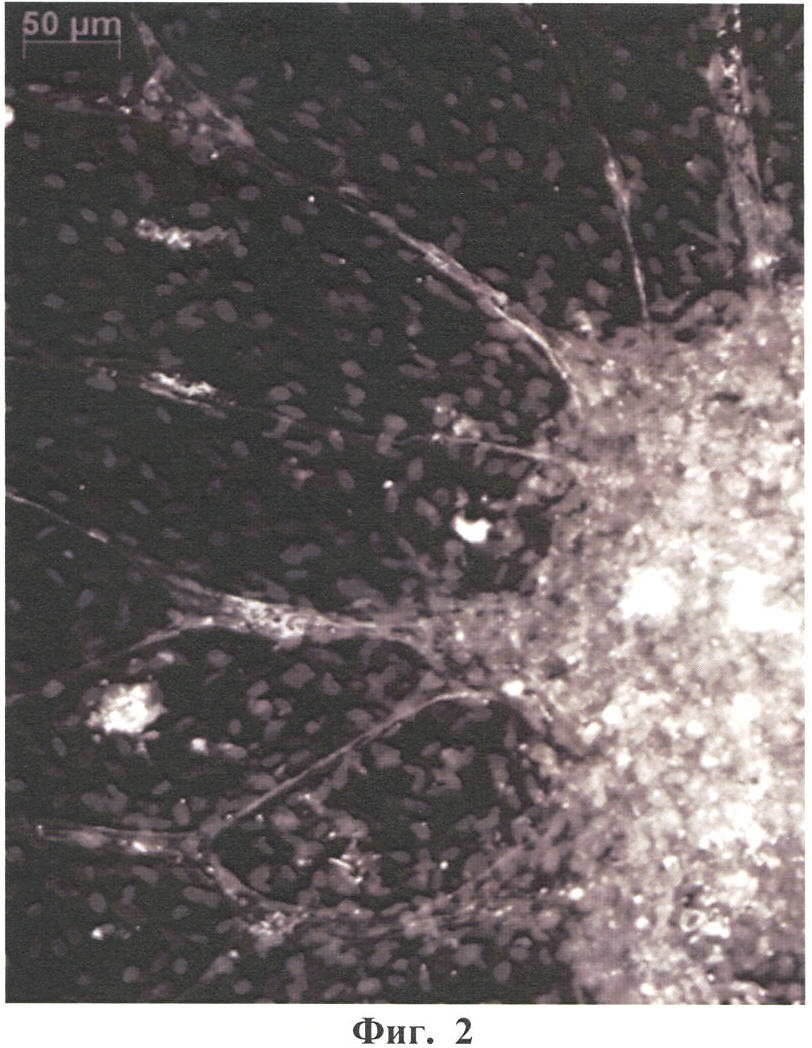
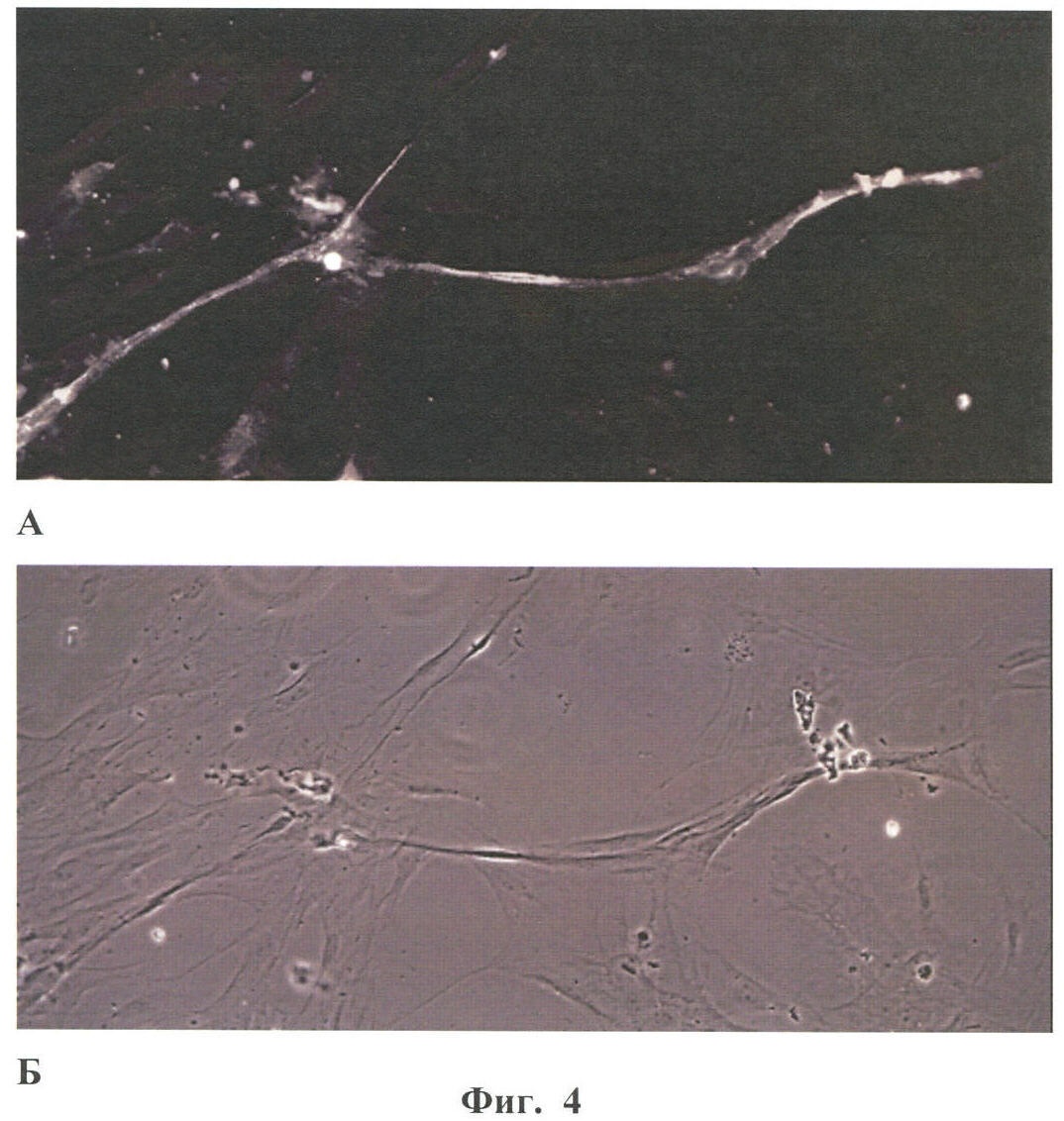
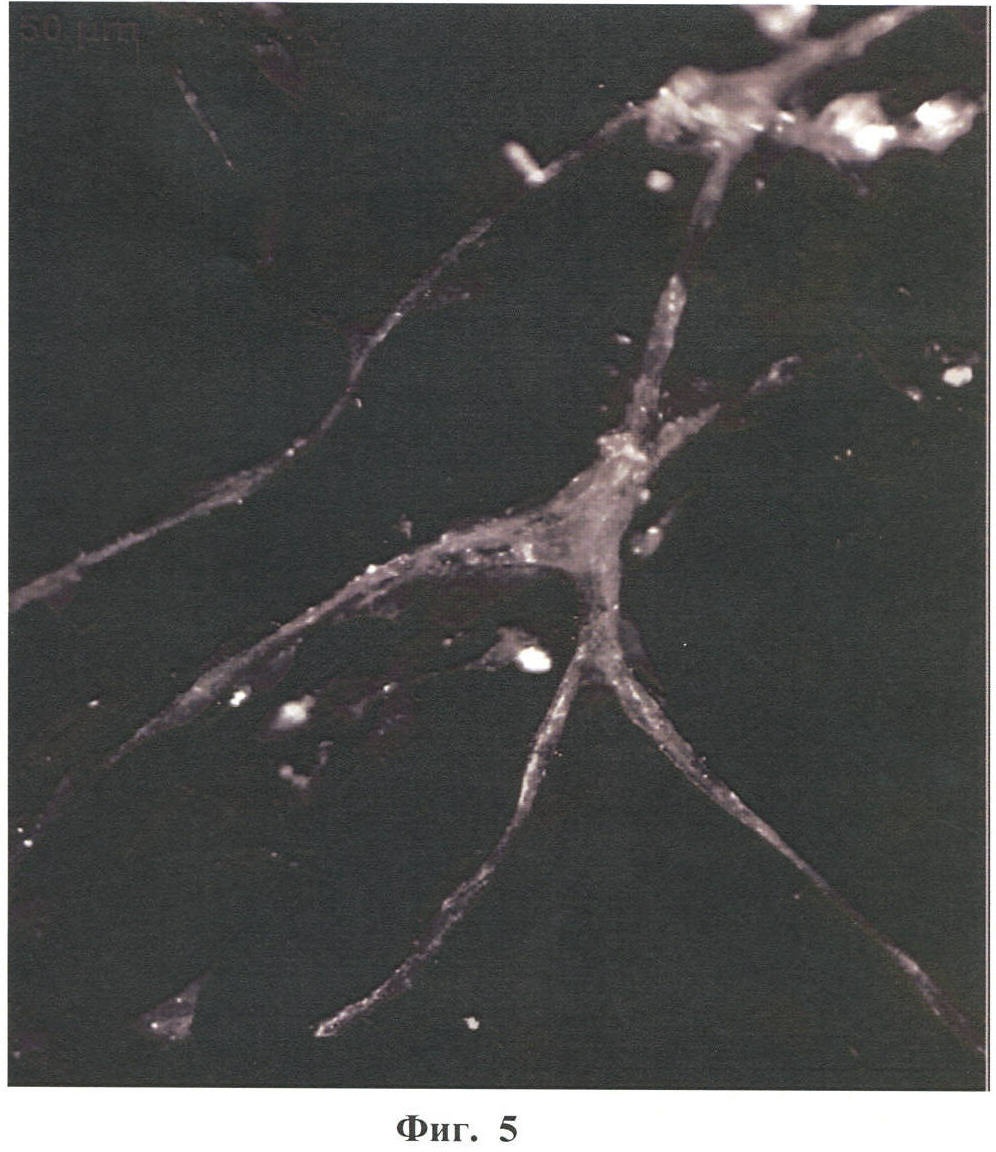
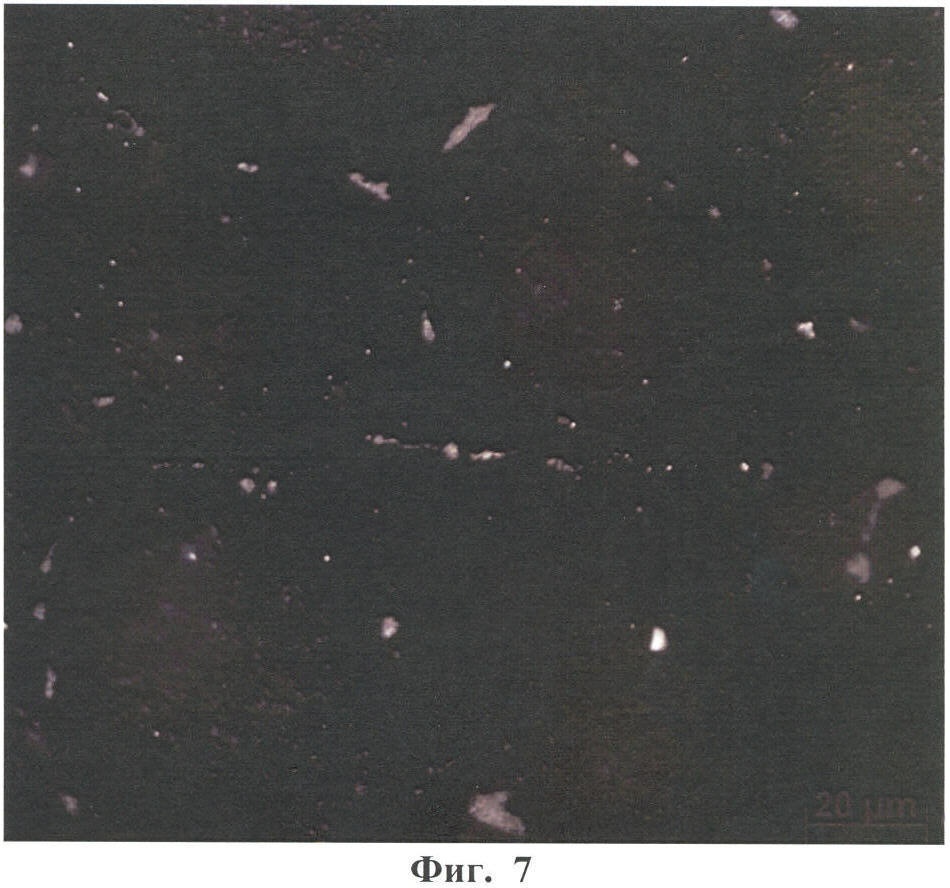
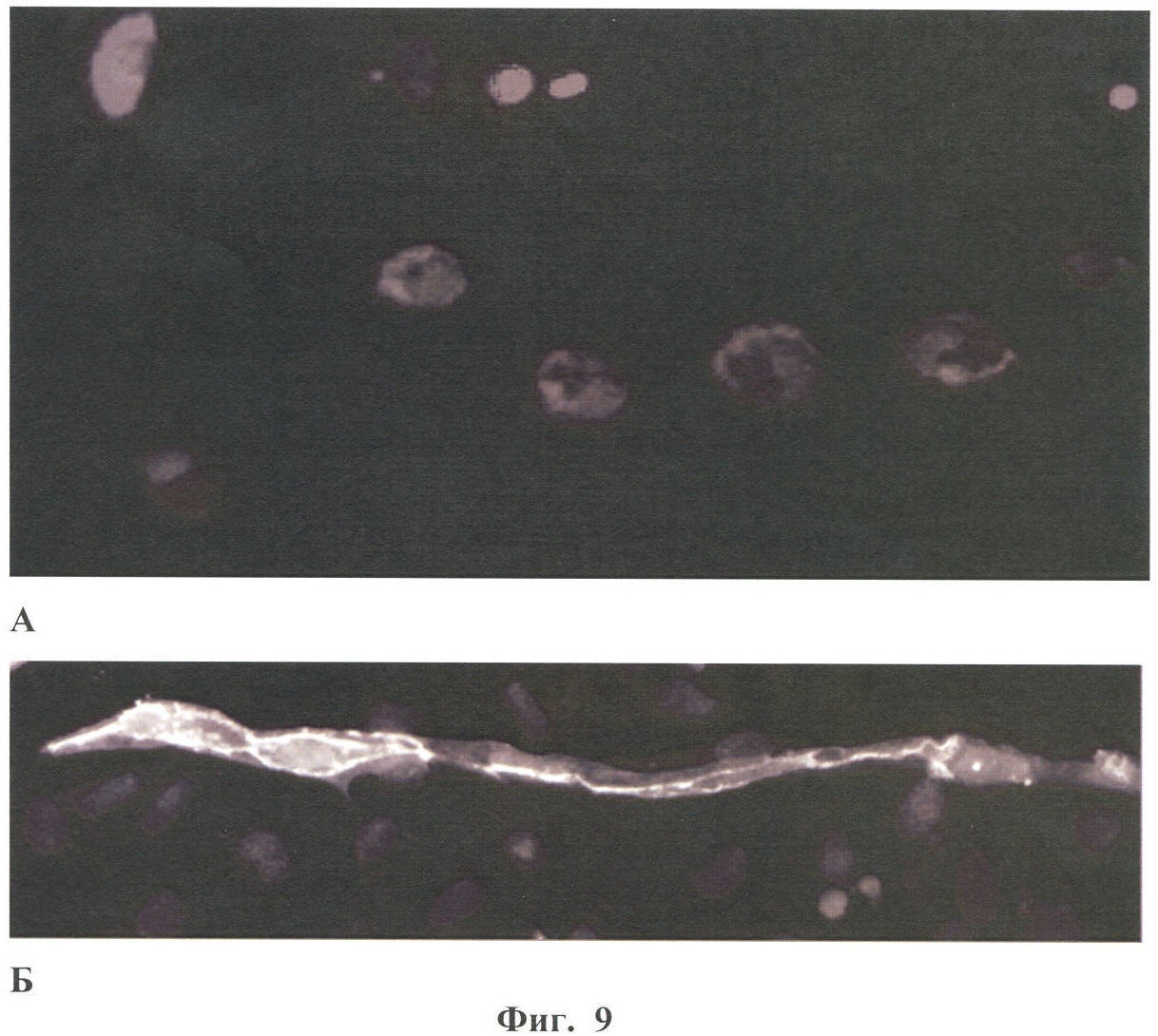
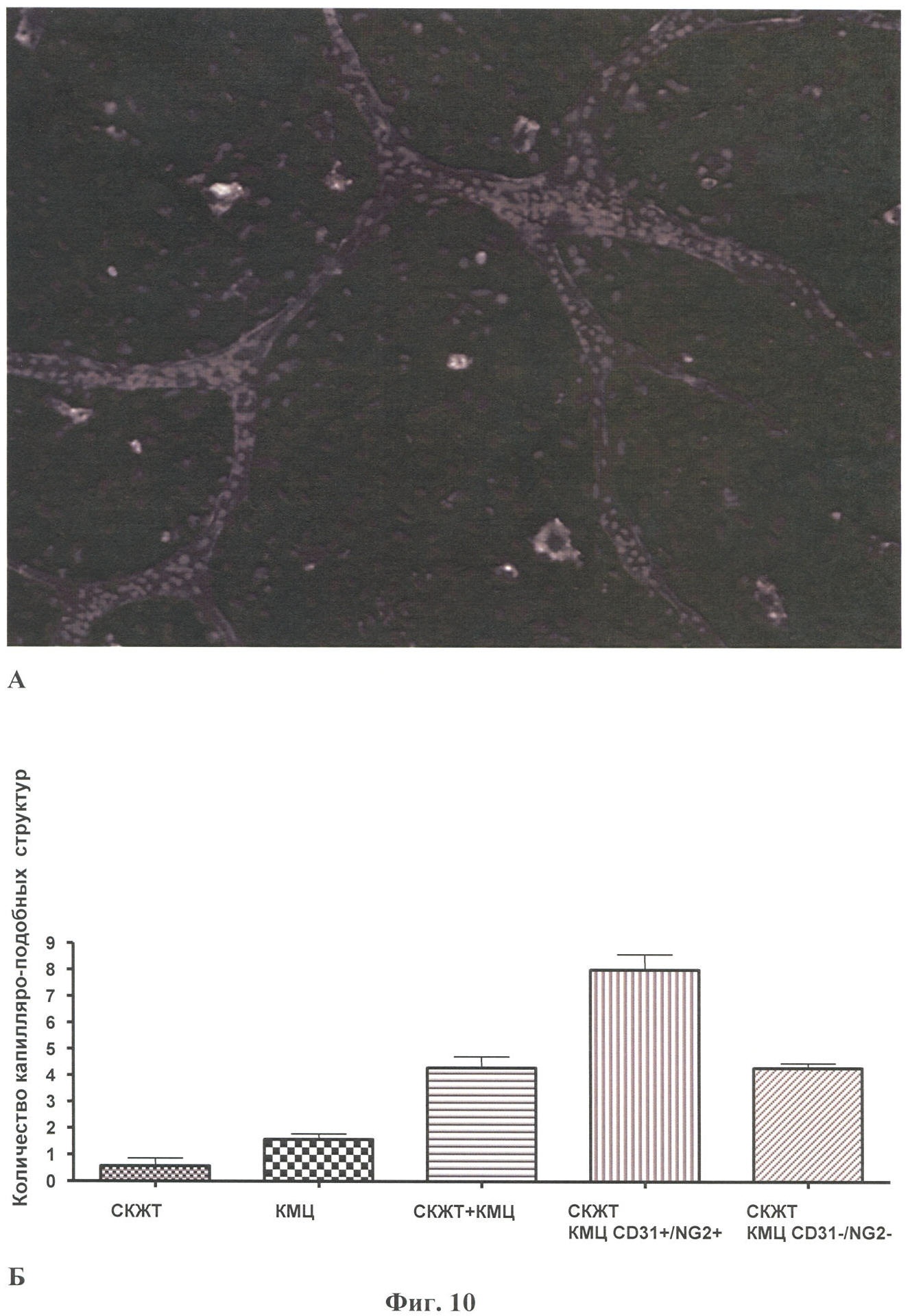
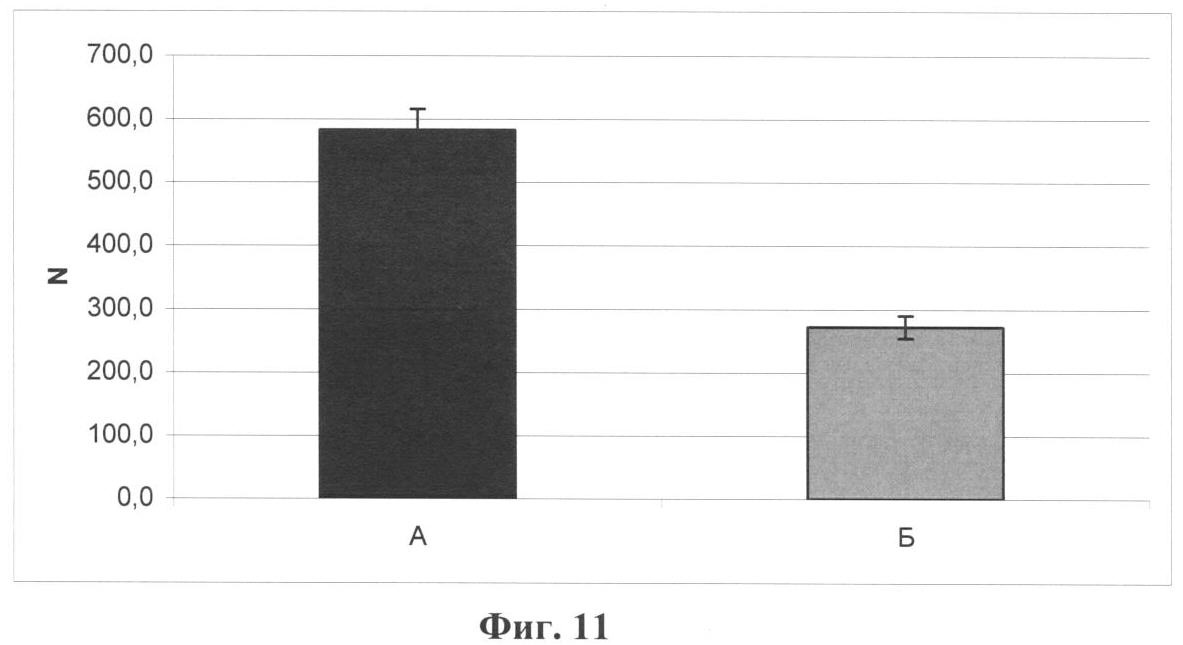
Настоящее изобретение относится к области биотехнологии, в частности к клеточной и тканевой инженерии, и может быть использовано в медицине и ветеринарии при разработке клинического метода локальной активации роста кровеносных сосудов (ангиогенеза) в ткани-мишени. Предлагается способ получения предназначенной для тканевой инженерии культуры клеток с ангиогенным фенотипом, заключающийся в со-культивировании клеток из постнатального сердца со стромальными клетками из подкожной жировой клетчатки (СКЖТ), приводящем к эффективной индукции образования и стабилизации сложных сосудистых структур in vitro. Ангиогенез – процесс образования новых кровеносных сосудов за счет ветвления и разрастания предсуществующего кровеносного русла. Ангиогенез сопровождает нормальное формирование органов и тканей человека в эмбриогенезе, а также наблюдается во взрослом организме в ряде физиологических и патологических процессов. В ходе образования новых кровеносных сосудов можно выделить несколько этапов: миграцию и пролиферацию эндотелиальных клеток и формирование ими новых капилляров; миграцию и пролиферацию субэндотелиальных клеток (перицитов) и их взаимодействие с эндотелиальными клетками. Взаимодействие перицитов с эндотелиальными клетками необходимо для формирования функционально зрелой и стабильной сосудистой сети, обеспечивающей нормальную трофику окружающих тканей. Все перечисленные этапы находятся под контролем целого ряда факторов, стимулирующих или подавляющих рост и/или стабилизацию сосудов. К таким факторам относятся растворимые биологически активные вещества (факторы роста, цитокины, гормоны и их метаболиты, модуляторы апоптоза), а также трансмембранные белки, экспрессирующиеся на поверхности клеток (кадгерины, интегрины и эфрины) (Brantley-Sieders et al., 2004, Ferrara et al., 2003). Возможность регулирования процесса формирования новых кровеносных сосудов открывает большие перспективы при разработке методов лечения и профилактики целого ряда тяжелых патологических состояний человека, в частности ишемических заболеваний сердца и нижних конечностей, которые сегодня являются одной из наиболее распространенных причин потери трудоспособности и смертности населения в развитых странах. Несмотря на внедрение в практику здравоохранения эффективных хирургических и медикаментозных методов лечения такого рода заболеваний, остается значительная часть больных, для которых эти методы либо непригодны, либо позволяют лишь частично восстановить кровоснабжение тканей. Для этой категории больных единственной альтернативой является терапия, направленная на стимуляцию роста новых сосудов в зоне ишемии. Раскрытие механизмов, регулирующих рост и ремоделирование сосудов, позволило разработать новую тактику лечения таких больных, основанную на введении в ишемизированные ткани ангиогенных факторов роста в виде рекомбинантных белков или генетических конструкций для стимуляции прорастания сосудов с целью улучшения их перфузии и функции. Сравнительно недавно с этой целью стали использовать стволовые или прогениторные клетки. С этой тактикой, получившей название терапевтического ангиогенеза, современная медицина связывает большие надежды, что определяет актуальность новых исследований и разработок в данной области. Уровень техники. Для клеточной терапии в фундаментальных и клинических исследованиях уже в течение ряда лет применяют такие типы прогениторных клеток, как эмбриональные стволовые клетки (Klug et al., 1996), скелетные миобласты (Menasche et al., 2003), культивированные предшественники эндотелиальных клеток (Kawamoto et al., 2001) и мезенхимальные клетки костного мозга (МККМ) (Perin et al., 2003). Несмотря на наличие позитивного терапевтического эффекта при введении многих из названных типов клеток в зону ишемии, сложность получения их в достаточном количестве (при использовании эндотелиальных клеток-предшественников), невозможность рутинного применения по этическим причинам (для эмбриональных стволовых клеток), наличие серьезных побочных эффектов (нарушения ритма сердца при введении скелетных миобластов), а также болезненность процедуры получения (для клеток МККМ) привели к необходимости поиска новых источников клеток для клеточной терапии. Одним из таких новых источников прогениторных клеток стала жировая ткань. Было обнаружено, что популяция клеток, выделяемых из жировой ткани специальными методами (стромальные клетки жировой ткани – СКЖТ), гетерогенна и обладает высокой пластичностью, а именно СКЖТ могут дифференцироваться не только в адипоциты, но и в другие типы клеток – хондробласты, остеобласты и миобласты (Zuk et al., 2001). Дальнейшие исследования показали, что вне зависимости от источника выделения СКЖТ характеризуются высокой пролиферативной способностью, наличием одного или нескольких маркеров стволовых клеток (CD34+ для человека и Sca-1+ для мыши) и отсутствием высокоспецифичных маркеров адипоцитов (Gronthos et al 2001; Beltrami et al., 2003). Более того, недавно было установлено, что подавляющее большинство маркеров, присутствующих на поверхности СКЖТ, схожи с маркерами, выявленными на поверхности МККМ (Трактуев и др., 2006). При этом по сравнению с клетками МККМ СКЖТ имеют ряд преимуществ, среди которых сравнительная легкость выделения и экспансии в культуре. СКЖТ могут быть получены хирургическим путем посредством взятия небольшого (примерно 15 г) образца подкожного жира из области живота и легко выращены в культуре in vitro до необходимого количества. С учетом указанных позитивных особенностей СКЖТ были предприняты первые попытки использования этих клеток для стимуляции ангиогенеза и восстановления кровотока в ишемизированных тканях. Так, на экспериментальной модели ишемии задней конечности у тимус-дефицитных мышей линии NOD/SCID были получены данные, свидетельствующие о том, что введение СКЖТ в хвостовую вену на следующий день после иссечения бедренной артерии предотвращает развитие некроза и восстанавливает кровоток в ишемизированной конечности у мыши (Парфенова и др., 2006). Аналогичные результаты по стимуляции ангиогенеза при ведении СКЖТ в зону ишемии были получены нами при использовании модели инфаркта миокарда с последующим развитием хронической ишемии миокарда и сердечной недостаточности у крыс линии Lewis (Zuk et al., 2001). Следует отметить, что во всех описанных до настоящего времени способах стимуляции ангиогенеза в ишемизированной ткани с помощью СКЖТ предлагается использовать введение индивидуальной популяции СКЖТ, либо свежевыделенных, либо пассированных известными методами (Zuk et al., 2001 – прототип). Однако при этом не учитывается тот факт, что вводимые в зону ишемии «живые» СКЖТ оказываются в пораженной ткани с частично или полностью нарушенной структурой и трофикой и сниженным функциональным потенциалом. Очевидно, что это осложняет реализацию стимулирующего и репаративного действия СКЖТ. Настоящее изобретение представляет собой разработку, в которой предпринята попытка устранить указанный недостаток и повысить потенциальные возможности вводимого клеточного материала в плане стимуляции ангиогенеза в ишемизированной области. Раскрытие изобретения Разработан способ получения клеточной культуры, предназначенной для стимуляции ангиогенеза в ишемизированной зоне, сущность которого заключается в предварительной индукции образования в этой культуре разветвленной и стабильной сосудистой сети, достигаемой в результате со-культивирования in vitro СКЖТ с клетками кардиомиоцитарной фракции (КМЦ) в определенных соотношениях и условиях. Предлагаемый способ основан на установленном нами экспериментально факте формирования in vitro стабильных капилляроподобных структур клетками, выделяемыми из ткани постнатального сердца, в присутствии СКЖТ и результатах исследования механизмов их образования, по поводу которых до настоящего времени не существовало единого мнения. Вкратце эти результаты сводятся к следующему. а) Кардиомиоцитарная фракция клеток содержит клетки-предшественники, способные дифференцироваться в различные типы клеток: кардиомиоциты, фибробласты и миофибробласты, а также эндотелиальные клетки. Последние, наряду с представленными в этой фракции дифференцированными эндотелиальными клетками, способны формировать капилляроподобные структуры в условиях in vitro. Однако эти спонтанно образующиеся в культуре структуры нестабильны и через некоторое время разбираются. б) Со-культивирование фракции клеток, выделяемых из постнатальных сердец, совместно с СКЖТ приводит к образованию значительно большего количества капилляроподобных структур, повышению степени их ветвления, а также к их существенной стабилизации. в) Механизм стимуляции ангиогенеза стромальными клетками из жировой ткани, наблюдаемой при со-культивировании кардиомиоцитарной фракции клеток с СКЖТ, реализуется, во-первых, за счет секреции клетками СКЖТ в среду культивирования специфических факторов, индуцирующих образование капилляроподобных структур как из эндотелиальных клеток, исходно представленных в кардиомиоцитарной фракции, так и в результате дифференцировки клеток-предшественников в эндотелиальные клетки; во-вторых, за счет клеточных взаимодействий между КМЦ и СКЖТ, стабилизирующих эти структуры. Раскрытый в настоящем изобретении способ подготовки клеточной культуры, включающей СКЖТ и предназначенной для тканевой инженерии в зоне ишемии, обладает очевидным преимуществом перед известными аналогами, которое заключается в расширении возможностей для повышения функционального потенциала вводимого клеточного материала. Во-первых, для индукции ангиогенеза в ишемизированной ткани предлагается использовать совместную культуру СКЖТ и КМЦ, содержащую не только клетки, стимулирующие ангиогенез (СКЖТ), но и полноценные клетки кардиомицитарного происхождения (например, прогениторные клетки) вместо частично поврежденных клеток, присутствующих в ишемизированной ткани сердца; во-вторых, со-культивирование in vitro открывает возможность, основываясь на результатах исследования механизма формирования капилляроподобных структур, оптимальным образом регулировать/изменять состав вводимой популяции клеток и соотношение СКЖТ и клеток кардиомиоцитарной фракции при подготовке культуры (см. пример 6). Прогениторные клетки, пригодные для использования в предлагаемом способе, могут быть и клетками костного мозга, и циркулирующими в крови клетками-предшественниками, и клетками, полученными из биопсийного материала (резидентные клетки-предшественники сердца). Предварительное со-культивирование клеток из жировой ткани с этими клетками позволяет еще до введения клеточного материала в организм стимулировать развитие в культуре ангиогенного фенотипа и обеспечить стабилизацию сосудов, образующихся затем в ткани-мишени. При этом клеточный материал для клеточной терапии может быть аутологическим, то есть СКЖТ и клетки-предшественники могут быть взяты у пациента и выращены в культуре для введения этому же пациенту. Благодаря этому, способ получения клеточной культуры по изобретению может быть предложен для непосредственного использования в клинике при лечении ишемии миокарда, а также легко адаптирован к применению для восстановления ишемизированных тканей других органов, например печени, поджелудочной железы, мочевого пузыря, соединительной ткани, дермы. В предлагаемом изобретении для со-культивирования с СКЖТ использовали кардиомиоцитарную фракцию клеток из ткани постнатального сердца; при этом наилучшие результаты были получены в случае применения свежевыделенных клеток из сердца, и особенно клеток, предварительно подвергнутых процедуре приплейтинга. Помимо тотальной фракции КМЦ применяли фракцию, обогащенную CD31-позитивными (эндотелиальными) клетками (пример 6а), что приводило к повышению эффективности формирования капилляроподобных структур, а также CD31+ фракцию, обогащенную NG2-позитивными клетками (перицитами), что дополнительно повышало скорость образования структур, их количество и стабильность (пример 6б). В случае СКЖТ предпочтительным оказалось использование пассированных клеток, в частности клеток второго пассажа для со-культивирования с КМЦ. Процедура со-культивирования при осуществлении изобретения проводится в стандартных средах и условиях, применяемых для культивирования клеток животного происхождения. В представленных ниже конкретных примерах осуществления изобретения нами использовалась среда следующего состава: DMEM/10% фетальная сыворотка быка/100 Ед/мл пенициллина/10 мкг/мл стрептомицина, однако для этой цели, кроме указанной выше, могут быть использованы и другие известные среды с добавлением фетальной сыворотки и антибиотиков, концентрации которых подбираются специалистом в типовом эксперименте. Культивирование происходит в стандартных условий в СО2-инкубаторе (5% СО2, 95% воздуха) при +37оС. Как было установлено, эффективность формирования капилляроподобных структур в культуре существенно зависела от соотношения со-культивируемых клеток. В частности, хотя образование их в той или иной степени наблюдалось при всех использованных соотношениях СКЖТ и КМЦ в интервале от 1:4 до 4:1, предпочтительным с точки зрения эффективности процесса было соотношение 1:4, которое и применялось нами в большинстве представленных примеров. Таким образом, объектом настоящего изобретения является способ получения предназначенной для тканевой инженерии культуры клеток с индуцированным ангиогенным фенотипом, предусматривающий со-культивирование пассированных стромальных клеток жировой ткани (СКЖТ) со свежевыделенными клетками либо тотальной кардиомиоцитарной фракции из ткани постнатального сердца (КМЦ), либо фракции КМЦ, обогащенной CD31-позитивными и/или NG2-позитивными клетками, при соотношении СКЖТ/КМЦ в интервале от 1:4 до 4:1, которое осуществляют в стандартных для культивирования клеток животного происхождения условиях до появления в культуре микроскопически определяемых капилляроподобных структур. Ниже приводятся конкретные примеры, которые подтверждают возможность осуществления изобретения, но при этом не ограничивают его объем. Краткое описание фигур Фиг.1. Культура клеток, выделяемых из постнатальных сердец. А – Иммунофлуоресцентное выявление экспрессии маркера кардиальных предшественников Nk×2,5; Б – Иммунофлуоресцентное выявление экспрессии маркера дифференцированных кардиомиоцитов Фиг.2. Кардиомиоцитарная колония, характеризующаяся синхронным биением в культуре, с образовавшимися в ней капилляроподобными структурами. Иммунофлуоресцентное окрашивание на маркер эндотелиальных клеток CD31. Фиг.3. Динамика изменения количества капилляроподобных CD31 позитивных структур, сформировавшихся в культуре клеток, выделяемых из постнатальных сердец. Столбец 1 – количество капилляроподобных структур через 1 день культивирования; столбец 2 – количество капилляроподобных структур через 3 дня культивирования; столбец 3 – количество капилляроподобных структур через 5 дней культивирования. Фиг.4. Культура клеток, получаемая при со-культивировании клеток, выделяемых из постнатальных сердец (КМЦ), со стромальными клетками из подкожной жировой клетчатки (СКЖТ) на 6-й день. А – Иммунофлуоресцентное окрашивание на маркер эндотелиальных клеток CD31. Б – Фазово-контрастное изображение этой же капилляроподобной структуры. Фиг.5. Со-культивирование КМЦ с СКЖТ. СКЖТ перед со-культивированием помечены флуоресцентным липофильным трейсером CM-DiI-Cell Tracker. Иммунофлуоресцентное окрашивание на маркер эндотелиальных клеток CD31 свидетельствует о смешанном происхождении капилляроподобных структур, где основу составляют клетки из КМЦ фракции, с которыми ко-локализуются СКЖТ. Фиг.6. Количество капилляроподобных структур, сформировавшихся в различных по составу культурах на 8-й день. Столбец 1 – количество капилляроподобных структур в культуре КМЦ; столбец 2 – количество капилляроподобных структур при со-культивировании КМЦ и СКЖТ (при соотношении 4:1); столбец 3 – количество капилляроподобных структур в культуре КМЦ при культивировании в присутствии среды культивирования СКЖТ; столбец 4 – количество капилляроподобных структур в культуре при со-культивировании КМЦ с СКЖТ, предварительно обработанными Митомицином С; р>0,05, U-критерий Манна-Уитни. Фиг.7. Культура клеток СКЖТ. Иммунофлуоресцентное окрашивание на маркер щелевых контактов коннексин 43. Фиг.8. Количество капилляроподобных структур, сформировавшихся в различных по составу культурах на 8-й день. Столбец 1 – количество капилляроподобных структур в культуре СКЖТ; столбец 2 – количество капилляроподобных структур в культуре КМЦ; столбец 3 – количество капилляроподобных структур при со-культивировании СКЖТ и КМЦ (при соотношении 1:4); столбец 4 – количество капилляроподобных структур при со-культивировании СКЖТ с кардиомиоцитарной фракцией CD31-позитивных клеток из постнатальных сердец (соотношение 1:4); столбец 5 – количество капилляроподобных структур при со-культивировании СКЖТ с кардиомиоцитарной фракцией CD31-негативных клеток из постнатальных сердец (соотношение 1:4); р>0,05, U-критерий Манна-Уитни. Фиг.9. А – Культура клеток СКЖТ. Иммунофлуоресцентное окрашивание на маркер перицитов NG2. Б – Со-культивирование КМС и СКЖТ при соотношении 4:1 (7-й день). Двойное иммунофлуоресцентное окрашивание на маркер эндотелиальных клеток CD31 и маркер перицитов NG2 свидетельствует о ко-локализации перицитов с капилляроподобными CD31-позитивными структурами. Фиг.10. А – Со-культивирование CD31+/NG2+ популяции КМЦ, выделяемых из постнатальных сердец после двойного иммуномагнитного сортинга, с СКЖТ на 6-й день в культуре (соотношение СКЖТ/КМЦ=1:4). Иммунофлуоресцентное окрашивание на маркер эндотелиальных клеток CD31. Б – Количество капилляроподобных структур, сформировавшихся в различных по составу культурах на 6-й день. Столбец 1 – количество капилляроподобных структур в культуре СКЖТ; столбец 2 – количество капилляроподобных структур в культуре КМЦ; столбец 3 – количество капилляроподобных структур при со-культивировании КМЦ с СКЖТ (при соотношении 4:1); столбец 4 – количество капилляроподобных структур при со-культивировании кардиомиоцитарной фракции CD31/NG2-позитивных клеток из постнатальных сердец с СКЖТ (соотношение 4:1); столбец 5 – количество капилляроподобных структур при со-культивировании СКЖТ с кардиомиоцитарной фракцией CD31/NG2-негативных клеток из постнатальных сердец (р>0,05, U-критерий Манна-Уитни). Фиг.11. Количество СD31-позитивных сосудов на срезах Матригеля через 2 недели после имплантации клеток мышам линии NUDE. Количество сосудов нормировали на площадь поля зрения (N). А – Введение совместной культуры КМЦ, выделяемых из постнатальных сердец, с клетками СКЖТ. Б – введение контрольных клеток индивидуальной культуры СКЖТ. Различия достоверны (р<0,0001, t-критерий Стьюдента). Осуществление изобретения. При осуществлении изобретения, помимо методов, подробно раскрытых в нижеследующих примерах, были использованы хорошо известные специалистам методики, описанные также в цитированных источниках. Эксперименты проводили на крысах линии Wistar, предоставленных биоклиникой РК НПК МЗ РФ. Пример 1. Получение первичной культуры СКЖТ крыс Выделение клеток проводили согласно ранее опубликованному протоколу (Zuk et al., 2001), с некоторыми модификациями. Для получения первичной культуры клеток СКЖТ использовали взрослых самцов крыс линии Wistar. Из подкожной жировой клетчатки, расположенной по бокам в нижней части туловища или паховой области, и висцерального внутрибрюшинного жира, расположенного непосредственно под широкой мышцей живота, изолировали участки ткани (в количестве примерно 2-5 грамм). Выделение жира проводили в асептических условиях, с использованием кетаминового наркоза из расчета 75 мг на кг веса животного. Выделение клеток из полученного в результате операции материала осуществляли в стерильных условиях ламинарного бокса. Ткань измельчали сосудистыми ножницами до консистенции суспензии мелких (размером не более нескольких кубических миллиметров) кусочков и смешивали с растворами ферментов коллагеназы I типа (200 ед/мл, Worthington Biochemical, США) и диспазы (40 ед/мл Invitrogen Corporation, Германия) при соотношении объема ткани (в мл) к объему ферментативного раствора (в мл) 1:2. Образец инкубировали при 37°С в течение 45 мин при постоянном покачивании; по окончании инкубации к нему добавляли равный объем среды DMEM (Gibco)/10% фетальная сыворотка быка (HyClone) и центрифугировали при 600 оборотах в течение 5 мин. Белесый поверхностный слой, представленный зрелыми адипоцитами и кусочками ферментативно необработанной ткани, удаляли, а осадок, состоящий из стромальных клеток жировой ткани (СКЖТ), а также остатков соединительной ткани и клеток крови суспендировали в лизирующем буфере для эритроцитов. Полученную смесь инкубировали 2-3 минуты в 37°С при перемешивании, после чего к образцу добавляли равный объем среды DMEM/10% ФСБ и фильтровали через нейлоновые мембраны (BD Falcon Cell Stainer # 352340) с размером пор 40 микрон. Для очищения популяции стромальных клеток от клеток крови и клеточного дебриса на конечном этапе суспензию центрифугировали при 800 оборотах 5 мин. Супернатант удаляли, а осадок ресуспендировали в среде MyeloCult (Stemcell, #M5300)/100 Ед/мл пенициллин/10 мкг/мл стрептомицин (GIBCO BRL). Затем высаживали клетки на чашки Петри (Corning) диаметром 60 мм в количестве 1×105 клеток СКЖТ на каждую чашку и культивировали в среде MyeloCult (8ml). На следующий день среду культивирования меняли на свежую среду состава DMEM/10% ФСБ/100 Ед/мл пенициллина/10 мкг/мл стрептомицина и выращивали их в СО2-инкубаторе (5% СО2; 95% воздуха) при 37°С. Смену среды проводили каждые 2-3 дня; при достижении монослоя клетки пассировали с использованием 0,25% раствора трипсин/ЭДТА. Аналогичным образом выделяли и использовали в экспериментах по со-культивированию СКЖТ из подкожного жира человека, полученного в результате удаления интактного жирового отложения при проведении пластических/косметических операций. Пример 2. Получение первичной культуры постнатальных КМЦ Для получения первичной культуры постнатальных кардиомиоцитов использовали крысят линии Wistar 2-5 дня после рождения. Сердца выделяли в асептических условиях и помещали в раствор Кребс-Рингера в присутствии Са2+ (50 mM). В стерильных условиях ламинарного бокса производили измельчение сердец в растворе коллагеназы А (Roche) (2mg/ml) на буфере Кребс-Рингер (при соотношении ткань:раствор примерно 1:2). Полученную суспензию кусочков в растворе фермента переносили в термошейкер при 37°С и инкубировали при покачивании 5-10 мин. По истечении времени инкубации смесь несколько раз пипетировали, супернатант отбирали, а к осажденным крупным фрагментам ткани добавляли новую порцию фермента. Процедуру повторяли 2 раза. После последней инкубации с ферментом отбирали осадок; к осадку добавляли равный объем среды DMEM/10% ФСБ и центрифугировали при 600 об/мин 4-8 мин. Супернатант сливали, а осадок, содержащий клетки, ресуспендировали в среде для приплейтинга DMEM/5% ФСБ /среда 199 (Gibco)/5% horse serum (Hyclone)/100 Ед/мл пенициллин/10 мкг/мл стрептомицин (соотношение среды DMEM:среда 199 – 4:1). Полученную суспензию клеток высаживали на чашку Петри в среде для приплейтинга и инкубировали в СО2-инкубаторе в течение 1 ч при 37°С. За это время фибробласты, обладающие повышенной адгезивной способностью в условиях приплейтинга, прикреплялись к чашке Петри. Не прикрепившиеся клетки собирали в отдельную пробирку, подсчитывали их концентрацию в камере Горяева и высаживали на чашки Петри в концентрации 500 клеток на 1 см2 (5×105 клеток на чашку Петри с диаметром 60 мм). В пробирку с оставшимися крупными фрагментами ткани добавляли новую порцию фермента. Процедуру повторяли 5 раз. Суммарную популяцию клеток, выделяемых после приплейтинга, центрифугировали при 600 об/мин 3-4 мин, ресуспендировали в среде для приплейтинга и высаживали на пластик. Первую смену среды осуществляли через 1 день, далее клетки культивировали в среде DMEM/5% ФСБ/100 Ед/мл пенициллин/10 мкг/мл стрептомицин в СО2-инкубаторе, а смену среды проводили каждые 2-3 дня. При достижении монослоя клетки пассировали с использованием 0,25% раствора трипсин/ЭДТА. Пример 3. Со-культивирование популяции клеток, выделяемых из постнатальных сердец, с СКЖТ Для экспериментов по со-культивированию использовали СКЖТ второго пассажа, достигшие конфлюэнтного состояния, и свежевыделенную кардиомиоцитарную фракцию клеток из постнатальных сердец (КМЦ). СКЖТ снимали с подложки с использованием 0,25% раствора трипсин/ЭДТА. Количество обоих типов клеток перед со-культивированием подсчитывали в камере Горяева, перемешивали в выбранном соотношении (СКЖТ/КМЦ: 1/4, 1/2, 2/1 или 4/1) и высаживали в пластиковые чашки Петри на стерильные покровные стекла. Максимальное количество капилляроподобных структур образуется при со-культивировании СКЖТ c КМЦ в соотношении 1:4, поэтому дальнейшие эксперименты проводили именно при этом соотношении клеток. СКЖТ со-культивировали с КМЦ в среде DMEM/10% ФСБ/100 Ед/мл пенициллина/10 мкг/мл стрептомицина в СО2-инкубаторе при 37°С течение 20 дней; при достижении монослоя совместную культуру пассировали. Наблюдения проводили с использованием инвертированного микроскопа Axiovert 200M (Zeiss) с фазовым контрастом и с системой для прижизненных наблюдений (столик с подогревом и камера с СО2-инкубатором). Пример 4. Иммунофлуоресцентный анализ Для фенотипической характеристики получаемых культур клеток проводили непрямое иммуноцитохимическое окрашивание клеток и флуоресцентный анализ изображений с использованием микроскопа Axiovert 200M (Zeiss) и программы AxioVision 3.1. Клетки на стеклах промывали теплым фосфатно-солевым буфером (ФСБ: 137 мМ NaCl, 2.7 мМ KCl, 4.3 мМ Na2HPO4*2H2O, 1.4 мМ KH2PO4), фиксировали 4% формалином на ФСБ в течение 15 мин и отмывали в ФСБ 3 раза по 10 мин. Для предотвращения неспецифического связывания стекла с клетками инкубировали в 10% растворе нормальной сыворотки донора вторых антител на 1% БСА (бычий сывороточный альбумин) в ФСБ в течение 40 мин. После этого наносили первые антитела в концентрации, рекомендованной производителем, разведенные на 1% БСА, и инкубировали во влажной камере в течение 1 часа при комнатной температуре. Клетки отмывали от первых антител в ФСБ 3 раза по 10 мин и наносили вторые антитела, меченные флуорохромом; инкубировали в течение 40-50 мин, отмывали в ФСБ 3 раза по 10 минут и докрашивали ядра DAPI (Molecular Probes, США) в концентрации 1 мкл/1 мл ФСБ в течение 20 минут. Затем стекла промывали 3 раза по 10 минут в ФСБ и заключали в водорастворимой специализированной среде для заключения иммунофлуоресцентных препаратов (Polysciences, Inc США). В качестве первых антител использовались следующие препараты: CD31(PECAM-1) (mouse antirat, Pharmingen 22711D), виметин (mouse Sigma, V-6630), GATA-4 (rabbit, Santa Cruz, 1404), Nk×2,5(rabbit, Santa Cruz, sc-14033), тропонин I (mouse, Chemicon, MAB3438), легкая цепь-1 миозина сердца (mouse, Abcam, ab680), коннексин 43 (mouse, Zymed, 40587905), Пример 5. Общие характеристики получаемых культур КМЦ, СКЖТ и совместной культуры КМЦ и СКЖТ а) По данным иммуноферментного анализа выделение популяции КМЦ по описанному протоколу приводило к получению разнородной культуры клеток, в которой на 2-4 день культивирования формировались колонии (Фиг.2), обладающие способностью к спонтанному синхронному сокращению на непокрытом пластике. В культуре КМЦ были обнаружены клетки-предшественники кардиомиоцитов, экспрессирующие Nk×2,5 (Фиг.1А) и GATA-4, дифференцированные кардиомиоциты, экспрессирующие б) Культура СКЖТ 2-го пассажа, которую использовали для со-культивирования в примере 3, представляет собой гетерогенную популяцию клеток, в которой присутствуют фибробласты (виментин и CD90-позитивные клетки), миофибробласты, экспрессирующие в) При со-культивировании СКЖТ и КМЦ использовались различные соотношения клеток: 1/4, 4/1, 2/1, 1/2. При этом во всех случаях, начиная примерно с 2 дня, наблюдалось появление капилляроподобных структур, хотя и с различной интенсивностью. Максимальное количество новообразованных структур в одни и те же сроки обнаруживалось при соотношении СКЖТ/КМЦ=1:4 (4×105 КМЦ и 1×105 СКЖТ на чашку Петри с диаметром 60 мм). Существенно, что в отличие от культуры КМЦ, в которой капилляроподобные структуры образуются, но спонтанно разбираются после 3-х дней культивирования (Фиг.3), в процессе со-культивирования КМЦ с СКЖТ капилляроподобные структуры, начинающие формироваться примерно на 2-3 день, остаются стабильными до 21 дня культивирования (Фиг.4). Из этих данных следует, что сосудоподобные структуры могут образоваться из клеток КМЦ фракции (предположительно за счет присутствующих там клеток предшественников или дифференцированных эндотелиальных клеток), однако стабилизируются эти структуры только в присутствии СКЖТ. Стабилизирующее действие СКЖТ на капилляроподобные структуры может осуществляться как за счет секреции специфических факторов, так и за счет межклеточных взаимодействий между СКЖТ и КМЦ. Для ответа на этот вопрос были проведены следующие эксперименты. Пример 6. Анализ клеточного состава капилляроподобных структур, образующихся при со-культивировании КМЦ и СКЖТ а) СКЖТ до начала со-культивирования метили липофильным флуоресцентным красителем CM-DiI-Cell Tracker (Molecular Probes, США), связывающимся с компонентами клеточных мембран. Для этого убирали среду культивирования и добавляли 4 мл теплого ФСБ, содержащего 6 мкл раствора CM-DiI-Cell Tracker, предварительно разведенного в 50 мкл этилового спирта, согласно рекомендациям производителя. Клетки инкубировали в СО2-инкубаторе при +37оС в течение 10 минут, а затем 15 минут при +4оС. Клетки отмывали в нескольких сменах ФСБ и среды и после этого со-культивировали с КМЦ, как описано в примере 3. Через 6 дней культуру КМЦ и СКЖТ фиксировали и окрашивали антителами к CD31, как описано в примере 4. С помощью флуоресцентного и фазово-контрастного анализа исследовали локализацию меченых СКЖТ и состав сформировавшихся капилляроподобных структур. При иммунофлуоресцентном анализе капилляроподобных структур в CM-DiI-Cell Tracker-меченные СКЖТ клетки флуоресцируют в красной области, а CD31-позитивные структуры – в зеленой. Соответственно, если меченые СКЖТ входят в состав CD31-позитивной структуры, то при наложении изображений спектр флуоресценции становится желтым. Если нет ко-локализации СКЖТ с CD31-позитивной структурой, то при наложении изображений регистрируется отдельно красная и зеленая флуоресценция. Оказалось, что CD31-позитивные капилляроподобные структуры имеют преимущественно кардиомиоцитарное происхождение (CM-DiI-Cell Tracker-меченные СКЖТ не входят в состав CD31-окрашенных капилляроподобных структур, а локализуются вдоль них). Однако в отдельных случаях капилляроподобные структуры демонстрируют смешанное происхождение, что, видимо, обуславливается плотным контактом КМЦ клеток с СКЖТ, локализованными в непосредственной близости от CD31-позитивных структур (Фиг.5). Отсюда можно заключить, что формирующиеся структуры собираются за счет эндотелиальных клеток и/или за счет дифференцирующихся клеток-предшественников, присутствующих в кардиомиоцитарной фракции постнатальных сердец. СКЖТ также могут входить в состав капилляроподобных структур, однако, как правило, они локализуются в непосредственной близости от этих структур и контактируют с ними. Поскольку, как описано ранее и показано на Фиг.3, в отсутствии СКЖТ формирующиеся в культуре структуры спонтанно разбираются, можно заключить, что именно наблюдаемое контактирование СКЖТ с капилляроподобными образованиями является основой их стабилизации. С высокой степенью вероятности стабилизирующее действие СКЖТ при формировании капилляроподобных может обеспечиваться межклеточными взаимодействиями с участием щелевых контактов, маркер которых коннектин 43 обнаруживается при иммунофлуоресцентном анализе культуры СКЖТ (Фиг.7). б) С целью оценки роли растворимых факторов, секретируемых СКЖТ, в образовании и/или стабилизации капилляроподобных структур были проведены эксперименты по культивированию КМЦ в присутствии среды культивирования СКЖТ и по со-культивированию КМЦ с СКЖТ, предварительно обработанными Митомицином С в течение 7 дней. Для этого среду для культивирования КМЦ готовили следующим образом: среду культивирования СКЖТ предварительно центрифугировли при 1000g в течение 10 минут, концентрировали в 100 раз с использованием фильтра Centricon YM-10 (Millipore) и далее разводили средой DMEM/10%/ФСБ/100 Ед/мл пенициллин/10 мкг/мл стрептомицин (как описано в примере 3) в соотношении 1:1. По окончании инкубации КМЦ в присутствии среды культивирования СКЖТ было отмечено достоверное увеличение количества капилляроподобных структур по сравнению с контролем (КМЦ), однако число их было достоверно меньшим (примерно в 2 раза), чем при со-культивировании с СКЖТ (Фиг.6). Таким образом, наличие в среде только растворимых факторов не компенсирует полного эффекта, наблюдаемого при со-культивировании КМЦ с СКЖТ. Этот вывод подтверждается и данными по со-культивированию КМЦ с СКЖТ, предварительно обработанными Митомицином С в концентрации 50 Из полученных в данном примере данных следует, что влияние СКЖТ реализуется как за счет выделения в среду культивирования факторов, стимулирующих образование и стабилизацию капилляроподобных структур, а также за счет клеточных взаимодействий между клетками кардиомиоцитарной фракции и СКЖТ, стабилизирующих сформированные структуры. Пример 7. Иммуномагнитный сортинг клеток Для определения типа клеток, вносящих основной вклад в формирование капилляроподобных структур в культуре in vitro, суммарную популяцию КМЦ разделяли на фракции методом иммуномагнитного сортинга клеток. Для иммуномагнитного сортинга клеток использовали магнитные частицы размером 40 нм, конъюгированные с антителами против мышиных IgG, (Dynabeads®Pan Mouse IgG, prod. No 110.41). а) Разделение КМЦ на CD31+ и CD31-фракции Магнитные частицы отмывали от азида натрия и инкубировали с мышиными антителами против CD31 (маркер эндотелиальных клеток) крысы в течение 40 минут на льду при постоянном помешивании. После окончания инкубации пробирку с магнитными частицами помещали в магнит и отмывали от не связавшихся антител. Все манипуляции проводили в стерильных условиях культурального бокса. Затем подготовленные магнитные частицы инкубировали с суспензией клеток, выделенных из постнатальных сердец, в течение 1 часа при постоянном покачивании на льду. После инкубации клетки помещали в магнит и разделяли на две фракции: связавшиеся с магнитными частицами CD31-позитивные клетки и оставшиеся в суспензии CD31-негативные клетки, после чего использовали их для со культивирования с СКЖТ, проводившегося, как описано в примере 3. При количественном анализе образовавшихся капилляроподобных структур при со культивировании CD31-негативных КМЦ с СКЖТ было обнаружено, что число их достоверно не отличалось от количества структур, образующихся при со культивировании СКЖТ с тотальной фракцией КМЦ (Фиг.8). Эти данные позволяют заключить, что при со культивировании СКЖТ с фракцией КМЦ, не содержащей CD31-позитивных клеток, формирование капилляроподобных структур реализуется за счет дифференцировки кардиальных клеток-предшественников в CD31-позитивные клетки и что данный механизм, соответственно, «работает» и при со культивировании СКЖТ с полноценной фракцией КМЦ. При обогащении КМЦ фракции эндотелиальными CD31-позитивными клетками и последующем культивировании их с СКЖТ количество капилляроподобных структур достоверно повышалось по сравнению с вариантами со-культивирования СКЖТ с тотальной фракцией КМЦ или со-культивирования СКЖТ с фракцией КМЦ после CD31 деплеции (Фиг.8). Это свидетельствует о наличии механизма формирования CD31-позитивных структур за счет организации эндотелиальных клеток, исходно присутствующих во фракции КМЦ. Таким образом, полученные данные позволяют сделать вывод о том, что при образовании капилляроподобных структур в условиях со-культивирования КМЦ с СКЖТ основную роль играют присутствующие в культуре эндотелиальные клетки, причем как зрелые CD31-позитивные клетки, исходно представленные во фракции КМЦ, так и появляющиеся в результате дифференцировки CD31-негативных клеток-предшественников. Это открывает дополнительную возможность повышения эффективности формирования капилляроподобных структур путем изменения состава предназначенной для со-культивирования с СКЖТ фракции КМЦ, а именно путем обогащения последней эндотелиальными клетками. б) Двойной иммуномагнитный сортинг Известно, что перициты, для которых характерна экспрессия NG2-маркера, играют важную роль в стабилизации и созревании кровеносных сосудов (Gerhardt and Betsholtz, 2003). Иммунофлуоресцентный анализ показал, что при со-культивировании NG2-позитивные клетки, которые исходно присутствуют как в культуре клеток из постнатальных сердец, так и в культуре СКЖТ (Фиг.9А), ко-локализуются с CD31-позитивными капилляроподобными структурами (Фиг.9Б). С помощью двойного иммуномагнитного сортинга популяция выделяемых КМЦ была разделена на две фракции: CD31+/NG2+ и CD31-/NG2-. Для этого было приготовлено два типа магнитных частиц: магнитные частицы, коньюгированные с антителами против мышиных IgG, и инкубированные с мышиными антителами против CD31 крысы, и магнитные частицы, коньюгированные с антителами против IgG кролика, инкубированные с поликлональными антителами кролика против NG2. Популяцию выделяемых КМЦ последовательно подвергали двойному иммуномагнитному сортингу (аналогично описанному в примере 6а) и со-культивировали с СКЖТ. При культивировании СКЖТ с CD31+/NG2+ популяцией клеток из постнатальных сердец образование капилляроподобных структур наблюдалось уже на 4-й день со-культивирования, а количество их было достоверно выше, чем при со-культивировании СКЖТ с фракцией CD31-/NG2- КМЦ или с тотальной фракцией КМЦ (Фиг.10б). Кроме того, структуры, образованные при со-культивировании СКЖТ с CD31+/NG2+ популяцией КМЦ, значительно отличались от структур, образованных при со-культивировании СКЖТ с тотальной фракцией КМЦ (Фиг.4 и Фиг.10А), в частности они характеризовались большей многослойностью, диаметром и разветвленностью (Фиг.10А). Соответственно, обогащение фракции КМЦ NG2-позитивными клетками при со-культивировании с СКЖТ играет дополнительную стабилизирующую роль при формировании капилляроподобных структур, образуемых в основном эндотелиальными клетками. Пример 7. Введение полученной совместной культуры СКЖТ+КМЦ мышам линии NUDE а) Подкожная имплантация клеток в Матригеле Совместную культуру клеток СКЖТ+КМЦ, полученную, как описано в примере 3, или индивидуальную культуру СКЖТ (контроль) вводили мышам линии NUDE подкожно в виде суспензии в Матригеле. Для этого клетки снимали с культуральных чашек 0,25% раствором трипсина на растворе Версена, центрифугировали при 1000g 5 минут и ресуспендировали в небольшом объеме среды; количество клеток считали в камере Горяева. Все манипуляции с клетками проводились в стерильных условиях. Далее готовили суспензию клеток в среде так, чтобы в 100 мкл среды находилось примерно 1,7 млн клеток. Далее 400 мкл холодного (+4°С) Матригеля смешивали со 100 мкл суспензии клеток и вводили подкожно мышам. Через 2 недели мышей умерщвляли изофлюрановым наркозом (Isofluran, Fso Flo б) Окрашивание срезов бляшек Матригеля методом иммунофлюоресценции и подсчет количества сосудов Матригели погружали в среду для заморозки тканей Tissue-Tek (Sacura Finetechnical Co., Ltd.), замораживали в жидком азоте и готовили срезы толщиной 7 мкм на криостате (НМ 505 Е, Microm, Германия). Срезы окрашивали антителами против CD31 – маркера эндотелиальных клеток, как описано в примере 4. Подсчет сосудов проводили на 4-5 срезах с каждого образца Матригеля в 4-5 полях зрения для каждого среза. Полученное количество сосудов в одном поле зрения нормировали на площадь поля зрения, оцененную по окрашиванию ядер с использованием DAPI. Статистический анализ данных проводился в программе Statistica 6.0. При подсчете числа сосудов использовали t-критерий Стьюдента. При введении совместной культуры КМЦ+СКЖТ в Матригеле мышам подкожно наблюдалось достоверное увеличение количества образующихся CD31-позитивыных сосудов по сравнению с контролем (СКЖТ) (Фиг.11). Эти данные свидетельствует о том, что совместная культура клеток, выделяемых из постнатальных сердец, с клетками СКЖТ более эффективна при стимуляции ангиогенеза по сравнению с индивидуальной культурой СКЖТ. Заключение Таким образом, полученные результаты свидетельствуют о том, что в кардиомиоцитарной фракции, выделяемой из постнатальных сердец крысы, присутствуют зрелые эндотелиальные клетки, способные организовываться в капилляроподобные структуры, а также клетки-предшественники (Фиг.1а), способные дифференцироваться в эндотелиальные клетки и участвовать в формировании капилляроподобных структур (Фиг.2). Однако эти структуры, образованные из КМЦ фракции клеток, нестабильны и к 3-му дню спонтанно разбираются (Фиг.3). Со-культивирование клеток КМЦ фракции с клетками СКЖТ приводит к образованию стабильных капилляроподобных структур (Фиг.4, Фиг.5). Стабилизирующая роль СКЖТ реализуется как за счет секреции в среду культивирования специфических факторов, так и благодаря образованию межклеточных щелевых контактов (Фиг.6, Фиг.7). При этом только секретируемых факторов для эффективной индукции и стабилизации структур недостаточно. Важную роль в стабилизации CD31-позитивных структур (предположительно за счет образования коннексин 43-содержащих щелевых контактов) играют присутствующие во фракции СКЖТ перициты. Разделение клеток, выделяемых из постнатальных сердец, на фракции, различающиеся по своей способности к формированию капилляроподобных структур при со-культивировании с СКЖТ, и, соответственно, использование при со-культивировании наиболее активных фракций позволяет повысить эффективность индукции образования и стабилизации сложных сосудистых структур in vitro. Это обстоятельство следует особо учитывать при реализации настоящего изобретения для стимуляции ангиогенеза в зоне ишемии in vivo. Список литературы 1. Трактуев Д.О., Парфенова Е.В., Ткачук В.А., Марч К.Л. Стромальные клетки жировой ткани – пластический тип клеток, обладающих высоким терапевтическим потенциалом. Цитология. 2006; том.4, №2, 83-94. 2. Парфенова Е.В., Цоколаева З.И., Трактуев Д.О., Джонсон Б., Талицкий К.А., Калинина Н.И., Ратрнер Е.И., Марч К., Ткачук В.А. Поиск новых «инструментов» для терапевтического ангиогенеза. Молекулярная медицина, 2006, №2, стр.10-23. 3. Beltrami AP, Barlucchi L, Torella D, Baker M, Limana F, Chimenti S, Kasahara H, Rota M, Musso E, Urbanek K, Leri A, Kajstura J, Nadal-Ginard B, Anversa P. (2003) Adult cardiac stem cells are multipotent and support myocardial regeneration. Cell. 114: 763-776. 4. Brantley-Sieders DM, Chen J. (2004) Eph receptor tyrosine kinases in angiogenesis: from development to disease. Angiogenesis. 7(1):17-28. 5. Ferrara N., Gerber HP., LeCouter J. The Biolgy of VEGF and its Receptors. (2003) Nat. Med. 9(6):669-676. 6. Gerhardt H., Betsholtz C. 2003. Endothelial-pericyte interactions in angiogenesis. Cell. Tissue Res. 314: 15-23. 7. Gronthos S, Franklin DM, Leddy HA, Robey PG, Storms RW, Gimble JM (2001) Surface protein characterization of human adipose tissue-derived stromal cells. J. Cell Physiol. 189:54-63. 8. Kawamoto A, Gwon HC, Iwaguro H, Yamaguchi JI, Uchida S, Masuda H, Silver M, Ma H, Kearney M, Isner JM, Asahara T (2001) Therapeutic potential of ex vivo expanded endothelial progenitor cells for myocardial ischemia. Circulation. 103:634-637. 9. Klug MG, Soonpaa MH, Koh GY, Field LJ (1996) Genetically selected cardiomyocytes from differentiating embronic stem cells form stable intracardiac grafts. J. Clin. Invest. 98:216-224. 10. Menasche P, Hagege AA, Vilquin JT, Desnos M, Abergel E, Pouzet B, Bel A, Sarateanu S, Scorsin M, Schwartz K, Bruneval P, Benbunan M, Marolleau JP, Duboc D (2003) Autologous skeletal myoblast transplantation for severe postinfarction left ventricular dysfunction. J. Am. Coll. Cardiol. 41:10781083. 11. Perin EC, Dohmann HF, Borojevic R, Silva SA, Sousa AL, Mesquita CT, Rossi MI, Carvalho AC, Dutra HS, Dohmann HJ, Silva GV, Belem L, Vivacqua R, Rangel FO, Esporcatte R, Geng YJ, Vaughn WK, Assad JA, Mesquita ET, Willerson JT (2003) Transendocardial, autologous bone marrow cell transplantation for severe, chronic ischemic heart failure. Circulation. 107: 2294-2302. 12. Tomasz M. (1987) Isolation and structure of a covalent cross-link adduct between mitomycin C and DNA. Science. 235:1204. 13. Zuk PA, Zhu M, Mizuno H, Huang J, Futrell JW, Katz AJ, Benhaim P, Lorenz HP, Hedrick MH. (2001) Mulitilineage Cells from Human Adipose Tissue: Implication for Cell-Based Therapies. Tissue Engineering, 7-2, 211-228.
Формула изобретения
1. Способ получения культуры клеток с индуцированным ангиогенным фенотипом для тканевой инженерии в зоне ишемии, предусматривающий сокультивирование in vitro пассированных стромальных клеток жировой ткани (СКЖТ) со свежевыделенными клетками либо тотальной кардиомицитарной фракции из ткани постнатального сердца (КМЦ), либо фракции КМЦ, обогащенной CD31-положительными и/или NG2-положительными клетками при соотношении СКЖТ/КМЦ от 1:4 до 4:1, которое осуществляют в стандартных для культивирования клеток животного происхождения среде и условиях до появления в культуре микроскопически определяемых капилляроподобных структур. 2. Способ по п.1, отличающийся тем, что для сокультивирования используют СКЖТ второго пассажа. 3. Способ по п.2, отличающийся тем, что клетки сокультивируют при соотношении СКЖТ/КМЦ, равном 1:4. 4. Способ по п.3, отличающийся тем, что сокультивирование проводят в течение 6-8 дней.
РИСУНКИ
|
||||||||||||||||||||||||||
 -актинина.
-актинина. g/ml. Известно, что обработка клеток Митомицином С приводит к блокированию синтеза ДНК (Tomasz, 1987) и, следовательно, к подавлению клеточной пролиферации. Установлено, что в этих условиях капилляроподобных структур также образуется достоверно больше, чем в контроле (КМЦ), однако их количество совпадает (различия недостоверны) с количеством структур, формирующихся при культивировании КМЦ в присутствии среды культивирования СКЖТ.
g/ml. Известно, что обработка клеток Митомицином С приводит к блокированию синтеза ДНК (Tomasz, 1987) и, следовательно, к подавлению клеточной пролиферации. Установлено, что в этих условиях капилляроподобных структур также образуется достоверно больше, чем в контроле (КМЦ), однако их количество совпадает (различия недостоверны) с количеством структур, формирующихся при культивировании КМЦ в присутствии среды культивирования СКЖТ. ), извлекали Матригель и замораживали в жидком азоте для последующего приготовления срезов, иммунофлюоресцентного окрашивания и подсчета сосудов.
), извлекали Матригель и замораживали в жидком азоте для последующего приготовления срезов, иммунофлюоресцентного окрашивания и подсчета сосудов.