Патент на изобретение №2324737
|
||||||||||||||||||||||||||||||||||||||||||||||
(54) СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА
(57) Реферат:
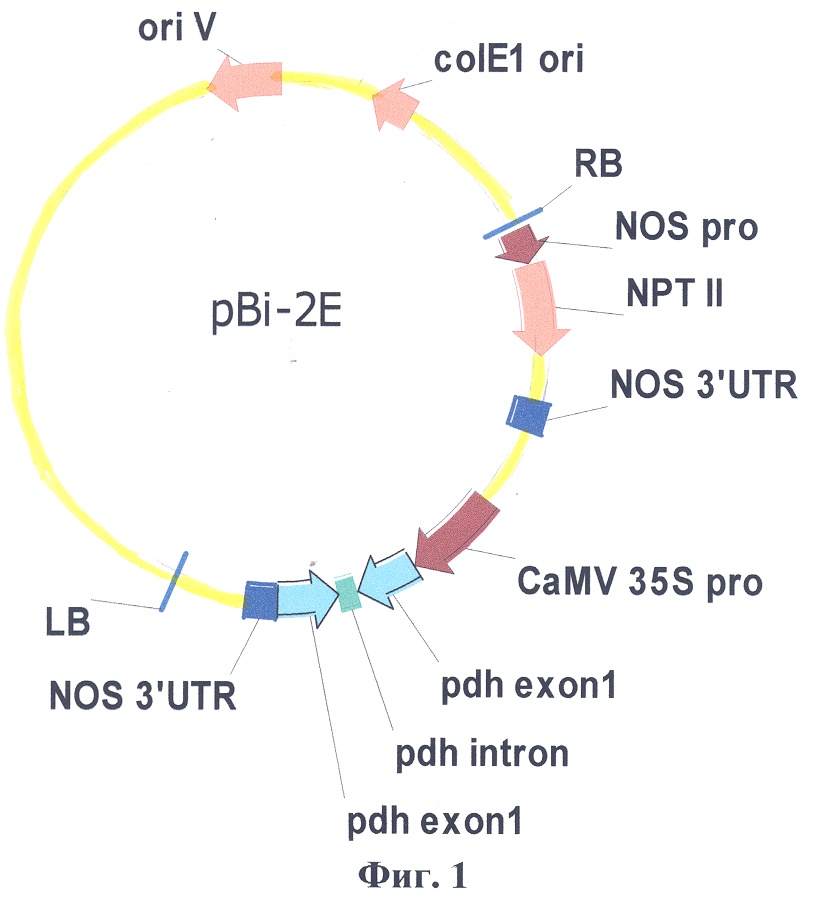
Изобретение относится к генетической инженерии растений. Конструируют рекомбинантную плазмидную ДНК pBi-2E, размером 14099 п.н., обеспечивающую перенос целевой последовательности ДНК в геном растений и экспрессию двуцепочечного супрессора гена PDH, состоящую из следующих элементов: – ДНК векторной плазмиды размером 14758 п.н. – фрагмента ДНК, содержащего первый экзон гена PDH в прямой ориентации размером 545 п.н. – фрагмента ДНК, содержащего первый экзон и фрагмент первого интрона гена PDH размером 691 п.н. в обратной ориентации; – 35S CaMV промотора из генома вируса мозаики цветной капусты; – 3’НТР области гена нопалинсинтазы. Плазмиду pBi-2E затем переносят в штамм Agrobacterium tumefaciens AGLO с последующей прямой трансформацией листовых дисков Nicotiana tabacum SRI сокультивацией с агробактерией. Отбор и генерацию производят непосредственно на средах, содержащих NaCl. Способ позволяет получать трансгенные растения табака без использования антибиотиков, обладающие увеличенной стрессоустойчивостью за счет повышенного содержания в них пролина. 3 ил., 1 табл.
Изобретение относится к области биотехнологии, в частности к генетической инженерии высших растений, и касается способа получения трансгенных растений табака с повышенным содержанием пролина. Известны два способа получения трансгенных растений с модифицированным метаболизмом пролина: с помощью повышения экспрессии гена Основными недостатком прототипа являются: 1. Необходимость введения в геном растений гена устойчивости к канамицину в составе Т-ДНК рекомбинантной плазмиды, который не играет никакой другой функциональной роли кроме возможности отбора. 2. Использование для отбора трансгенных растений селективной среды с добавлением антибиотика канамицина, что не позволяет классифицировать растения на повышенное содержание пролина, связанное со стрессоустойчивостью. Технической задачей настоящего изобретения является упрощение способа за счет оптимизации генетической конструкции для трансформации растений и повышение его экологической безопасности. Поставленная техническая задача достигается предлагаемым способом, заключающимся в следующем. Конструируют рекомбинантную плазмидную ДНК путем выделения первого экзона гена PDH из генома арабидопсиса (Arabidopsis thaliana) с помощью ПЦР со специфическими праймерами с последующим соединением фрагментов в прямой и обратной ориентациях в полилинкере Т-ДНК области бинарной векторной плазмиды рВi121. В результате получают рекомбинантную плазмидную ДНК pBi-2E, размером 14099 п.н., обеспечивающую перенос целевой последовательности ДНК в геном растений и экспрессию двуцепочечного супрессора гена PDH. Рекомбинантная плазмида состоит из следующих элементов: – ДНК векторной плазмиды размером 14758 п.н. – фрагмента ДНК, содержащего первый экзон гена PDH в прямой ориентации размером 545 п.н. – фрагмента ДНК, содержащего первый экзон и фрагмент первого интрона гена PDH размером 691 п.н. в обратной ориентации; – Промотора 35S РНК из генома вируса мозаики цветной капусты; – 3’НТР области гена нопалинсинтазы. Сконструированную рекомбинантную плазмиду pBi-2E, несущую последовательность гена PDH, переносят в штамм агробактерии (Agrobacterium tumefaciens) AGLO с последующей прямой трансформацией листовых дисков табака (Nicotiana tabacum) SR1 сокультивацией с агробактерией с последующим отбором и генерацией непосредственно на средах, содержащих NaCl. Физическая карта плазмиды pBi-2E с указанием генетических маркеров приведена на фиг.1, где pdh exoni – нуклеотидная последовательность первого экзона гена PDH арабидопсиса; pdh intron – фрагмент первого интрона гена PDH арабидопсиса; CaMV 35S pro – промотор гена 35S РНК вируса мозаики цветной капусты; NOS 3’UTR-3′-нетранслируемый район гена нопалинсинтазы из Ti-плазмиды агробактерии (Agrobacterium tumefaciens); NOS pro – промотор гена нопалинсинтазы; NPTII-ген неомицинфосфотрансферазы II; LB, RB – левая и правая границы Т-ДНК области. Определяющими отличительными признаками заявляемого способа по сравнению с прототипом, являются: 1. В качестве векторной плазмиды используют вектор рВi121, в котором ген
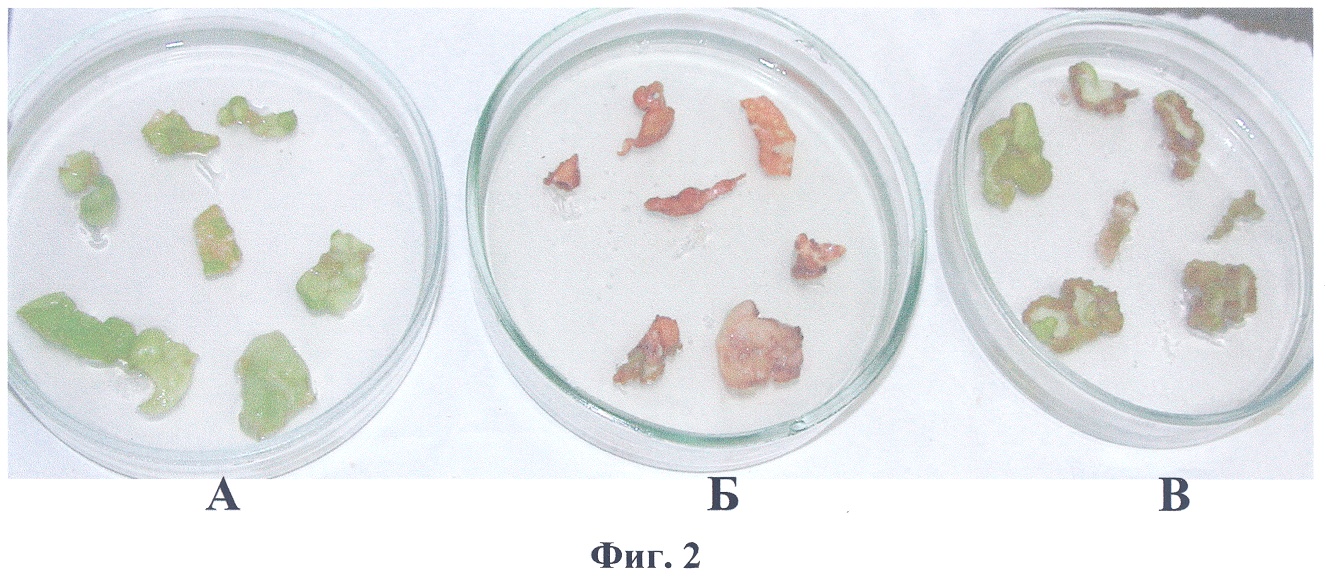
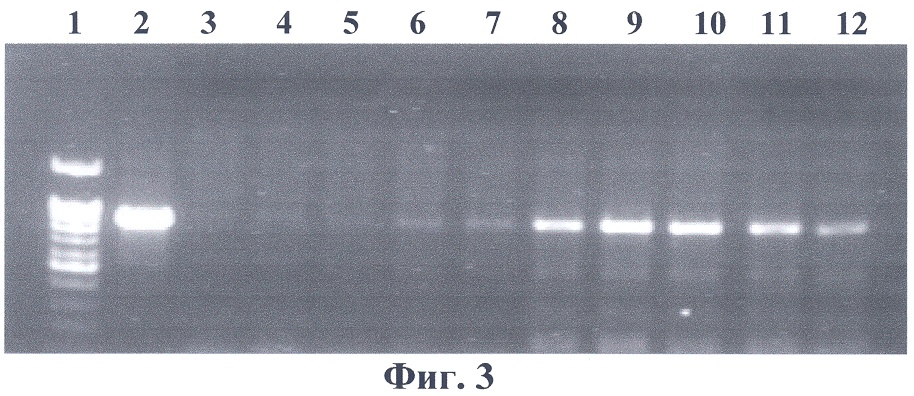
3. Отбор трансгенных растений осуществляют непосредственно на стрессовом фоне, а именно на среде, содержащей 250 mM NaCl. Изобретение поясняется следующими примерами конкретного выполнения способа. Пример 1 Конструирование рекомбинантной плазмиды pBi-2E Конструирование двуцепочечного супрессора в составе вектора pBi121 осуществляли посредством тройного лигирования. В конструкцию собирали три фрагмента: первый экзон гена пролиндегидрогеназы, первый экзон гена PDH с частью первого интрона и фрагмент плазмиды pBi121, содержащий в Т-области ген NPTII, промотор 35S РНК вируса мозаики цветной капусты и 3′-нетранслируемый район гена нопалинсинтазы, включающий сигнал полиаденилирования. Первый экзон гена PDH арабидопсиса был получен с помощью ПЦР (праймеры: 5′-aacaaactggatccggcgatcttac-3′; 5′-gagatgttggtctagatttggcagc-3′) и клонирован в плазмиде pBlueScript KSII по сайтам рестрикции BamHI и XbaI. Далее его выделяли с помощью рестриктаз BamHI и HindIII; ПЦР-ный фрагмент первого экзона гена PDH с частью первого интрона гидролизовали рестриктазой HindIII, плазмиду pBi121 гидролизовали рестриктазами BamHI и Есl136II. Продукты рестрикции разделяли в 1% агарозном геле, гель окрашивали раствором бромистого этидия в воде (1 мкг/мл) и методом сорбции ДНК на силикагеле выделяли необходимые фрагменты, которые далее обрабатывали ДНК-лигазой, и лигазной смесью трансформировали компетентные клетки Е. coli XL1 Blue. Трансформанты высевали на чашку с LB-агаром, содержащим 50 мкг/мл канамицина. Клоны, несущие рекомбинантные плазмиды со встроенным геном, отбирали методом ПЦР анализа ДНК отдельных колоний E.coli с праймерами на нуклеотидную последовательность первого экзона гена PDH арабидопсиса. Пример 2 Получение трансгенных растений, несущих конструкцию pBi-2E А – трансформация конструкцией pBi101, отбор на канамицине; Б – трансформация конструкцией pBi101, отбор на среде, содержащей 250 mM NaCl; В – трансформация pBi-2E, отбор на среде, содержащей 250 mM NaCl. На фиг.3 представлена электрофореграмма ПЦР-анализа геномной ДНК проростков трансгенных растений на наличие встройки 35S промотора вируса мозаики цветной капусты с фрагментом гена PDH в антисмысловой ориентации. Нанесения по дорожкам: 1 – маркерная ДНК; 2 – вектор pBi-2E; 3 – нетрансгенный табак SR1; 4-12 – трансгенные табаки SR1. Из фиг.3 видно, что ПЦР-анализ с праймерами на нуклеотидную последовательность первого экзона PDH подтверждает наличие нуклеотидной последовательности PDH арабидопсиса в геномах отобранных растений.
Из представленных в таблице данных видно, что полученные трансгенные растения табака, несущие рекомбинантную плазмиду pBi-2E, имеют повышенный в несколько раз уровень содержания пролина по сравнению с контрольными растениями. Таким образом, способ позволяет получать трансгенные растения табака, несущие последовательность первого экзона гена PDH в прямой и обратной ориентациях, с повышенным содержанием пролина.
Формула изобретения
Способ получения трансгенных растений табака с повышенным содержанием пролина, включающий конструирование рекомбинантной плазмидной ДНК, содержащей последовательность гена пролин дегидрогеназы (PDH) из Arabidopsis thaliana, передачу полученной плазмидной ДНК в штамм Agrobacterium tumefaciens, инфицирование последним растительного материала, отбор трансгенных растений на селективной питательной среде с последующим тестированием отобранных трансформантов на повышенное содержание пролина, отличающийся тем, что конструируют и передают в штамм указанной бактерии рекомбинантную плазмидную ДНК pBi-2E, состоящую из следующих элементов: ДНК векторной плазмиды рВI121, не содержащей ген бета-глюкуронидазы; фрагмента ДНК, содержащего первый экзон гена PDH в прямой ориентации размером 545 п.н; фрагмента ДНК, содержащего первый экзон и фрагмент первого интрона гена PDH размером 691 п.н. в обратной ориентации; 35S CaMV промотора из генома вируса мозаики цветной капусты; 3’НТР области гена нопалинсинтазы, при этом отбор трансгенных растений осуществляют на питательной среде, содержащей 250 mM NaCl и не содержащей антибиотики.
РИСУНКИ
|
||||||||||||||||||||||||||||||||||||||||||||||

 -глюкуронидазы был заменен на последовательности гена PDH, соединенных в прямой и обратной ориентациях, что позволило получить конструкцию меньшего размера, более оптимальную для трансформации.
-глюкуронидазы был заменен на последовательности гена PDH, соединенных в прямой и обратной ориентациях, что позволило получить конструкцию меньшего размера, более оптимальную для трансформации.