Патент на изобретение №2318019
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) ДНК-ВАКЦИНА ПРОТИВ ПРОЛИФЕРИРУЮЩИХ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК И СПОСОБЫ ЕЕ ПРИМЕНЕНИЯ
(57) Реферат:
ДНК-вакцина, эффективная для ингибирования пролиферации эндотелиальных клеток, включает ДНК-конструкцию, оперативно кодирующую рецепторный белок фактора роста сосудистого эндотелия (VEGF), выбранный из группы, состоящей из VEGFR-2 (KDR; SEQ ID NO: 2), VEGFR-1 (Flt-1; SEQ ID NO: 4) или Flk-1 (мышиный гомолог KDR, SEQ ID NO: 6) и их функционального эквивалента, гомологичного приблизительно на 80%. Кодирующие рецепторные белки ДНК-последовательности представлены в SEQ ID NO: 1, 3 и 5 в тексте описания. Описаны способы применения ДНК-вакцин для ингибирования пролиферации эндотелиальных клеток у млекопитающего путем введения ДНК-вакцины для стимуляции иммунного ответа в отношении пролиферирующих эндотелиальных клеток, в частности, для ингибирования ангиогенеза и опухолевого роста. Описана также готовая форма ДНК-вакцины с информацией, необходимой для индивидуального введения пациенту. В результате использования ДНК-вакцины по изобретению достигается эффект антиангиогенеза и последующее снижение роста и распространения опухоли. 5 н. и 9 з.п. ф-лы, 11 ил., 5 табл.
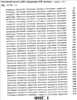
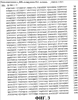
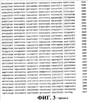
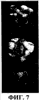
ПРАВИТЕЛЬСТВЕННЫЕ ПРАВА Настоящее изобретение было сделано при поддержке правительства, в соответствии с контрактом No. 5-70373-COLON, Национальными Институтами Здоровья. Правительство обладает определенными правами на данное изобретение. ОБЛАСТЬ ИЗОБРЕТЕНИЯ Данное изобретение связано с вакцинами дезоксирибонуклеиновой кислоты (ДНК), кодирующей соответствующие молекулы, эффективные для получения иммунного ответа против пролиферирующих эндотелиальных клеток. Более конкретно, настоящее изобретение связано с вакцинами ДНК, кодирующими рецептор фактора роста сосудистого эндотелия (VEGF). Данное изобретение связано также со способами применения ДНК-вакцин для ингибирования пролиферации эндотелиальных клеток сосудов, опухолевого роста и ангиогенеза. ОСНОВА СОЗДАНИЯ ИЗОБРЕТЕНИЯ Вакцины используются для обеспечения долговременной защиты против целого ряда болезненных состояний путем весьма ограниченного введения профилактического агента, который стимулирует иммунную систему организма к разрушению болезнетворных патогенов прежде, чем они смогут начать пролиферировать и вызвать патогенный эффект. Различные подходы к вакцинам и вакцинациям описаны у Bernard R. Glick и Jack J. Pasternak, Molecular Biotechnology, Principles and Applications of Recombinant ДНК, Second Edition, ASM Press pp. 253-276 (1998). Вакцинация является средством индуцирования собственной системы организма, в результате чего организм сможет распознать и разрушить инфицирующий агент прежде, чем тот вызовет патологическую реакцию. Обычно вакцины являются либо живыми, но аттенюированными инфекционными агентами (вирусами или бактериями), либо убитой формой этого агента. Вакцина, состоящая из живых бактерий или вирусов, должна быть непатогенной. Обычно бактериальную или вирусную культуру аттенюируют (ослабляют) путем физической или химической обработки. Полученный агент, хоть и не является вирулентным, однако все еще способен вызывать иммунную реакцию у субъекта, обработанного такой вакциной. Иммунный ответ вызывается антигенами, являющимися либо специфическими макромолекулами, либо инфекционными агентами. Такие антигены обычно являются либо белками, полисахаридами, липидами, либо гликолипидами, которые распознаются как “чужеродные” лимфоцитами, которые известны как B-клетки и T-клетки. Выдерживание (экспозиция) обоих типов лимфоцитов с антигенами вызывает быстрый ответ в виде стимуляции деления клеток и их дифференцировки, что приводит к образованию клонов экспонированных лимфоцитов. B-клетки продуцируют плазматические клетки, которые, в свою очередь, продуцируют белки, называемые антителами (Ab), которые выборочно связываются с антигенами, присутствующими на инфекционном агенте, таким образом нейтрализуя, или инактивируя, патоген (гуморальный иммунитет). В некоторых случаях для развития B-клеточного ответа требуется помощь со стороны CD4-хелперных Т-клеток. Специализированный Т-клеточный клон, который образуется в ответ на экспонирование с антигеном, представляет собой цитотоксические Т-лимфоциты (CTL), которые способны связываться с патогенами и тканями, на которых присутствует антиген, и вызывать их элиминацию (клеточно-опосредованный или клеточный иммунитет). В некоторых случаях антигенпредставляющие клетки (АПК), такие как дендритные клетки, обволакивают патоген или иную чужеродную клетку путем эндоцитоза. Затем АПК подвергают эти антигены процессингу и выставляют их на своей поверхности, представляя в виде комплекса из молекулы гистосовместимости и пептида T-клеточному рецептору (TCR) на поверхности цитотоксических лимфоцитов CTL, вызывая таким образом стимуляцию иммунного ответа. Гуморальный иммунитет, характеризующийся образованием специфических анител, обычно является более эффективным против острых бактериальных инфекций и повторных вирусных инфекций, тогда как клеточно-опосредованный иммунитет является наиболее эффективным против вирусной инфекции, хронической внутриклеточной бактериальной инфекции и грибковой инфекции. Клеточный иммунитет также известен в качестве защиты против рака, и он же ответственен за отторжение органных трансплантатов. Антитела против антигенов, связанных с предшествующими инфекциями, остаются детектируемыми в крови в течение долгого периода времени, что позволяет, таким образом, определить факт осуществления прежнего, более раннего, контакта с патогеном. При повторной встрече с тем же самым патогеном иммунная система эффективно предотвращает повторное инфицирование тем же самым патогеном путем элиминации патогенного агента еще до того, как он начнет пролиферировать и продуцировать патогенную реакцию. Тот же самый иммунный ответ, который мог бы возникнуть под действием патогена, может иногда быть вызван также и непатогенным агентом, который присутствует на том же самом антигене, что и патоген. Таким образом, субъект может быть защищен от дальнейшего контакта с патогеном без предварительной борьбы с ним. Однако не все инфекционные агенты могут быть успешно культивированы и инактивированы, как это требуется для образования вакцин. Современные технологии рекомбинантных ДНК создали возможность конструирования новых вакцин в попытке обойти это ограничение. Можно создать инфекционные агенты, в которых отсутствуют патогенные гены, что позволяет, таким образом, использовать невирулентные формы организмов в качестве вакцины. Возможно также сконструировать относительно непатогенный организм, такой как E. coli, для представления антигенов клеточной поверхности патогенного носителя. Иммунная система субъекта, обработанного таким трансформированным носителем, “обманным путем” начинает продуцировать антитела против указанного патогена. Антигенные белки патогенного агента могут быть сконструированы и экспрессированы у непатогенных видов, и эти антигенные белки могут быть выделены и очищены для получения “субъединичной вакцины”. Субъединичные вакцины имеют преимущества, связанные с их стабильностью, надежностью, а также с тем, что они хорошо охарактеризованы химически; однако их продукция может быть чрезвычайно дорогостоящей. Новый подход к вакцинам, в общем называемый генетической иммунизацией, появился в последние годы. В этом подходе ген, кодирующий антиген патогенного агента, оперативно встроен в клетку субъекта, который подлежит иммунизации. Обработанные клетки трансформированы и продуцируют антигенные белки патогена. Затем эти продуцируемые in vivo антигены запускают в организме хозяина требуемый иммунный ответ. Генетический материал, используемый в таких генетических вакцинах, может являться либо ДНК-, либо РНК-конструктом. Часто полинуклеотид, кодирующий антиген, вводят в комбинации с другими промоторными полинуклеотидными последовательностями для улучшения встраивания, репликации или экспрессии гена. ДНК-вакцины, кодирующие антигенные гены, могут быть введены в хозяйские клетки субъекта с помощью различных экспрессирующих систем. Такие экспрессирующие системы включают в себя экспрессирующие системы прокариот, млекопитающих и дрожжей. Например, одним из подходов является использование вирусного вектора, такого как вирус коровьей оспы, включающего в себя новый генетический материал, для инокуляции в хозяйские клетки. Альтернативно, генетический материал может быть встроен в вектор или может быть напрямую доставлен в хозяйские клетки в виде “голого” полинуклеотида, т.e. просто в виде очищенной ДНК. Кроме того, ДНК может быть стабильно трансфицирована в аттенюированные бактерии, такие как Salmonella typhimurium. Когда пациента перорально вакцинируют трансформированной Salmonella, эти бактерии транспортируются к Пейеровым бляшкам в пищеварительном канале (т.e. вторичным лимфоидным тканям), которые затем стимулируют иммунную систему. ДНК-вакцины предоставляют возможность иммунизировать против болезненных состояний, которые не вызываются традиционными патогенами, таких как генетические заболевания и рак. Обычно в генетической противораковой вакцине антигены к специфическому типу опухолевой клетки должны быть выделены, а затем введены в вакцину. Эффективная общая вакцина против ряда раковых заболеваний может, таким образом, послужить основой для развития целого ряда индивидуальных вакцин для каждого типа раковых клеток, против которых должна быть произведена иммунизация. Один из общих подходов для лечения опухолей включает в себя введение пациенту с растущими опухолями соединений, ингибирующих ангиогенез. Ангиогенезом называется процесс образования новых капилляров и кровеносных сосудов. Ангиогенез играет важную роль в период эмбрионального развития, роста тканей, репарации тканей и регенерации тканей. Помимо этих нормальных и основополагающих процессов, ангиогенез вовлечен также и во многие аномальные патологические процессы, такие как опухолевый рост, опухолевое метастазирование и заболевания сосудов глаза, такие как диабетическая ретинопатия. Ангиогенез вовлечен в целый ряд взаимозависимых процессов, включая (a) активацию сосудистых эндотелиальных клеток, (b) разрушение белков клеточного матрикса под действием эндотелиальных клеток, экспрессирующих протеиназную активность, (c) миграцию эндотелиальных клеток к сайтам потенциального роста, (d) пролиферацию эндотелиальных клеток, (e) образование протоков в результате дифференцировки эндотелиальных клеток. Каждый из этих процессов находится под влиянием различных промоторных веществ, таких как фактор роста фибробластов (FGF), фактор роста, полученный из тромбоцитов (PDGF), и факторы роста сосудистого эндотелия (VEGF). Факторы роста сосудистого эндотелия (коллективно обозначаемые VEGF) играют ключевую роль в процессах роста и дифференцировки. VEGF действует путем связывания с рецептором тирозиновых протеинкиназ, присутствующим в мембранах эндотелиальных клеток, который, в свою очередь, инициирует каскад реакций передачи сигнала, которые стимулируют клеточный рост. Ингибирование патологического ангиогенеза было предложено в качестве подхода для лечения опухолей. См., например, Folkman et al. Science, 221, 719 (1983). Основной концепцией такого лечения является то, что, поскольку опухоли, для того чтобы расти, нуждаются в прорастании кровеносных сосудов, ингибирование образования кровеносных сосудов путем введения соединений, ингибирующих ангиогенез, будет предотвращать опухолевый рост путем истощения опухоли, не снабжаемой кровью. Недостатком этого подхода является то, что ингибиторы ангиогенеза, для того чтобы предотвратить опухолевый рост, должны вводиться в относительно непрерывном режиме. Прекращение введения и доставки инибитора может привести к возобновлению опухолевого роста. Вакцина, эффективная для ингибирования ангиогенеза, могла бы быть весьма привлекательным превентивным средством против образования опухоли. Существует неослабевающая потребность в вакцине с широким спектром эффективности для иммунизации против ангиогенеза, которая могла бы также ингибировать рост различных опухолей, чтобы не было необходимости нацеливания ее на специфические опухолевые антигены. Настоящее изобретение удовлетворяет указанным требованиям. СУЩНОСТЬ ИЗОБРЕТЕНИЯ ДНК-вакцина, эффективная для ингибирования пролиферации эндотелиальных клеток, включает в себя ДНК-конструкцию, которая оперативно кодирует белок рецептора фактора роста сосудистого эндотелия (VEGF). ДНК-вакцина включает в себя полинуклеотид, который кодирует белок, осуществляющий рецепцию VEGF, такой как VEGFR-2 (KDR; последовательность SEQ ID NO: 2), VEGFR-1 (Flt-1; последовательность SEQ ID NO: 4) и Flk-1 (последовательность SEQ ID NO: 6; мышиный гомолог KDR), например ДНК-последовательности SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO 5 соответственно. Такая вакцина может включать в себя линейную нуклеиновую кислоту, такую как очищенный ДНК-конструкт или ДНК-конструкт, встроенный в плазмидный вектор. ДНК-вакцины согласно изобретению стимулируют образование цитотоксических лимфоцитов (CTL), активных против пролиферирующих эндотелиальных клеток, которые в повышенном количестве экспрессируют VEGFR-2. Эндотелиальные клетки образуют выстилку ткани сосудов у млекопитающих. Пролиферация эндотелиальных клеток является ключевым процессом ангиогенеза. Вакцины согласно изобретению обеспечивают способ обеспечения долговременного ингибирования ангиогенеза в организме, обработанном такой вакциной, путем стимуляции иммунного ответа против пролиферирующих эндотелиальных клеток. На непролиферирующих эндотелиальных клетках, таких как выстилка сформированных кровеносных сосудов, представлены незначительные количества антигенов рецептора VEGF, и, таким образом, эти клетки остаются незатронутыми цитотоксическими лимфоцитами, которые продуцируются в ответ на введение вакцины. В методическом аспекте согласно изобретению ДНК-вакцина используется для обеспечения долговременного ингибирования пролиферации эндотелиальных клеток у вакцинированного пациента. В одном из методических аспектов согласно изобретению ДНК-вакцину, включающую в себя полинуклеотидную конструкцию, оперативно кодирующую белок рецептора VEGF, вводят перорально пациенту, в случае необходимости ингибирования пролиферации эндотелиальных клеток, в количестве, достаточном для стимуляции иммунного ответа против пролиферирующих эндотелиальных клеток. Настоящее изобретение обеспечивает также способ ингибирования ангиогенеза у пациентов, вакцинированных ДНК-вакциной. При таком воплощении способа согласно изобретению пациенту, страдающему заболеванием, связанным с ангиогенезом, вводят количество вакцины, которое вызывает иммунный ответ, причем вакцина включает в себя ДНК-конструкцию, оперативно кодирующую белок рецептора VEGF. При другом воплощении способа согласно изобретению опухолевый рост ингибируется в результате вакцинирования пациента вакциной ДНК. При таком воплощении способа пациенту, у которого имеется растущая опухоль, вводят эффективное количество вакцины, которое вызывает иммунный ответ, причем эта вакцина включает в себя ДНК-конструкцию, оперативно кодирующую белок рецептора VEGF. Вакцинация приводит к остановке роста опухоли. Деструкция пролиферирующих эндотелиальных клеток под действием иммунных клеток пациента предотвращает васкуляризацию опухоли, что по существу приводит к истощению опухоли и ее гибели. В методических аспектах согласно изобретению, ДНК-вакцины могут быть введены энтерально, например, путем перорального введения или парентерально, например, путем инъекции или внутривенной инфузии. Вакцины согласно изобретению используются для лечения или предотвращения целого ряда болезненных состояний. Например, у пациентов, страдающих от рака, диабетической ретинопатии и тому подобного, может наступить улучшение под действием иммунизации вакцинами согласно изобретению. КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ На фиг. 1 приведена последовательность ДНК, кодирующая KDR человека, SEQ ID NO:1; на фиг. 2 приведена белковая последовательность KDR человека, SEQ ID NO:2; на фиг. 3 приведена последовательность ДНК, кодирующая Flt-1 человека, SEQ ID NO:3; на фиг. 4 приведена белковая последовательность Flt-1 человека, SEQ ID NO:4; на фиг. 5 приведена последовательность ДНК, кодирующая Flk-1 мыши, SEQ ID NO: 5; на фиг. 6 приведена белковая последовательность Flt-1 мыши, SEQ ID NO: 6; фиг. 7 представляет собой иллюстративное представление мышиных легких с различной степенью опухолевого поражения, составляющего от > 50% поражения (отмечено цифрой 3) до < 10% поражения (отмечено цифрой 1); фиг. 8 представляет собой графическое представление данных, показывающих, что у мышей, вакцинированных ДНК-вакциной согласно изобретению (сплошная, толстая черная линия), которым путем внутривенной инъекции переносили клетки карциномы кишечника CT-26, наблюдали значительное снижение смертности по сравнению с двумя контрольными группами мышей (необработанные вакциной (наивные) мыши: сплошная тонкая линия; мыши, обработанные контрольной вакциной: пунктирная линия); фиг. 9 представляет собой графическое представление данных, свидетельствующих о супрессии опухолевого роста карциномы легких Льюиса D121 у мышей, вакцинированных ДНК-вакциной согласно изобретению (pcDNA3.1-FLK-1), по сравнению с двумя контрольными группами мышей; фиг. 10 представляет собой графическое представление данных, свидетельствующих о супрессии опухолевого роста меланомы B16 у мышей, вакцинированных ДНК-вакциной согласно изобретению ( фиг. 11 представляет собой графическое представление данных, свидетельствующих о повышающей регуляции CD25-, CD69- и CD2-положительных CD8+-T-клеток у мышей, вакцинированных ДНК-вакциной согласно изобретению, по сравнению с контрольной группой мышей. ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВОПЛОЩЕНИЙ ДНК-вакцина, эффективная для ингибирования пролиферации эндотелиальных клеток, включает в себя ДНК-конструкцию, которая оперативно кодирует рецепторный белок фактора роста сосудистого эндотелия (VEGF). Используемый здесь в описании и в прилагаемой формуле изобретения термин “ДНК-конструкция”, или “ДНК-конструкт”, означает искусственную структуру ДНК, которая может быть транскрибирована в клетках-мишенях. Эта конструкция может включать в себя линейную нуклеиновую кислоту, такую как очищенная ДНК или, предпочтительно, ДНК, встроенная в плазмидный вектор. ДНК может также быть встроена в вирусный или бактериальный вектор, предпочтительно аттенюированный вирусный или бактериальный вектор, который не является патогенным. Подходящими являются такие ДНК, которые кодируют рецепторный белок VEGF, такой как VEGFR-2 (KDR; SEQ ID NO: 2), VEGFR-1 (Fit-1; SEQ ID NO: 4) и Flk-1 (SEQ ID NO: 6), например последовательности ДНК SEQ ID NO: 1, SEQ ID NO: 3 и SEQ ID NO 5 соответственно. Было идентифицировано пять подтипов VEGF, включая VEGF-1 (известный также как VEGF-A), VEGF-2 (известный также как VEGF-C), VEGF-B, VEGF-D и VEGF-E. См., например, патент США No. 6,235,713 выданный на имя Achen et al., и цитируемые здесь ссылки. Рецепторы VEGF представляют собой белковые тирозинкиназы, специфичные в отношении эндотелиальных клеток. Было идентифицировано несколько рецепторных белковых тирозинкиназ, специфичных в отношении эндотелиальных клеток, включая Flt-1 (рецептор 1 VEGF; VEGFR-1), KDR (VEGFR-2), Flk-1 (мышиный гомолог KDR), Flt-4 (VEGFR-3), Tie, Tie-2 и Tek, из которых несколько представляют собой рецепторы VEGF. ДНК-вакцины согласно изобретению стимулируют образование цитотоксических лимфоцитов, CTL, которые являются активными против пролиферирующих эндотелиальных клеток, которые экспрессируют повышенные количества VEGFR-2. Поскольку рецепторы VEGF в существенном количестве экспрессированы только на поверхности пролиферирующих эндотелиальных клеток, цитотоксические лимфоциты, которые образуются в ответ на введение вакцины, будут в основном нацелены только на те ткани, в которых происходит активный ангиогенез (например, васкуляризация). Непролиферирующие эндотелиальные клетки, такие как клетки, выстилающие сформированные кровеносные сосуды, являются в основном лишенными антигенов рецептора VEGF и являются, таким образом, не подверженными действию CTL, продуцируемых под действием вакцинации. В предпочтительном воплощении ДНК-вакцина включает в себя полинуклеотидную последовательность, которая оперативно кодирует белок рецептора VEGF. Эта вакцина может стимулировать активацию «наивных» T-клеток, как напрямую, так и опосредованно, через вмешательство дендритных клеток. Здесь под термином “иммунитет” подразумевается долговременная иммунологическая защита против вирулентной формы инфекционного агента или опухолевого антигена. Под термином “иммунизация” подразумевается профилактическое контактирование с антигеном патогенного агента, полученного из невирулентного источника, что приводит к развитию у субъекта, подвергнутого иммунизации, иммунитета против патогена. ДНК-конструкция согласно изобретению предпочтительно включает в себя нуклеотидную последовательность, которая кодирует белок рецептора VEGF, оперативно связанную с регуляторными элементами, необходимыми для экспрессии гена. Используемые ДНК-конструкции предпочтительно включают в себя регуляторные элементы, необходимые для экспрессии нуклеотидов. Такие элементы включают в себя, например, промотор, инициирующий кодон, стоп-кодон и сигнал полиаденилирования. Кроме того, для экспрессии последовательности, которая кодирует иммуногенный белок-мишень, часто требуются энхансеры. Как известно в данной области, предпочтительно, чтобы эти элементы были оперативно связаны с последовательностью, которая кодирует нужный белок. Предпочтительно выбирают такие регуляторные элементы, которые оперативны у видов, которым эти регуляторные элементы предполагается вводить. Инициирующие кодоны и стоп-кодоны предпочтительно включаются как часть нуклеотидной последовательности, которая кодирует белок рецептора VEGF в генетической вакцине согласно изобретению. Инициирующий и терминирующий кодоны должны быть в одной рамке считывания с кодирующей последовательностью. Промоторы и сигналы полиаденилирования, включенные в вакцину согласно изобретению, предпочтительно отбираются таким образом, чтобы они были функциональными в клетках субъекта, которого предполагается иммунизировать. Примеры промоторов, используемых в вакцинах согласно изобретению, особенно при получении генетических вакцин для человека, включают в себя, не ограничиваясь перечисленным, промоторы из обезьяньего вируса 40 (SV40), промотор вируса опухоли молочной железы мыши (MMTV), промотор вируса иммунодефицита человека (HIV), такой как промотор длинного концевого повтора (LTR) вируса иммунодефицита человека, вируса Молони, цитомегаловируса (CMV), такого как немедленный ранний промотор CMV, вируса Эпштейна-Барр (EBV), вируса саркомы Рауса (RSV), а также промоторы из других человеческих генов, таких как человеческий актин, человеческий миозин, человеческий гемоглобин, человеческий креатин мышц и человеческий металлотионеин. Примеры сигналов полиаденилирования, используемые в вакцинах согласно изобретению, особенно при получении генетических вакцин для человека, включают в себя, не ограничиваясь перечисленным, сигналы полиаденилирования SV40 и сигналы полиаденилирования LTR. Помимо регуляторных элементов, необходимых для экспрессии ДНК, в молекулу ДНК могут быть включены также и другие элементы. Такие дополнительные элементы включают в себя энхансеры. Энхансером может быть, например, человеческий актин, человеческий миозин, человеческий гемоглобин, человеческий креатин мышц и вирусные энхансеры, такие как энхансеры из CMV, RSV и EBV. Регуляторные последовательности и кодоны обычно зависят от вида, поэтому в целях максимального увеличения продукции белка, регуляторные последовательности и кодоны предпочтительно отбирают таким образом, чтобы они были максимально эффективными у видов, подлежащих иммунизации. Рядовой специалист в данной области может получить конструкции ДНК, которые будут функциональными у субъекта данного вида. ДНК-конструкции вакцин согласно изобретению могут представлять собой “голые” ДНК, как определено в публикации Restifo et al. Gene Therapy 7, 89-92 (2000), соответствующие фрагменты которой включены в данное описание в виде ссылки. Альтернативно ДНК могут быть оперативно встроены в вектор. Полезные векторы для доставки включают в себя биодеградируемые микрокапсулы, иммуностимулирующие комплексы (ISCOM) или липосомы и полученные генно-инженерным путем аттенюированные живые векторы, такие как вирусы или бактерии. Примеры соответствующих аттенюированных живых бактериальных векторов, как известно в данной области, включают в себя Salmonella typhimurium, Salmonella typhi, Shigella, Bacillus, Lactobacillus, Bacille Calmette-Guerin (BCG), Escherichia coli, Vibrio cholerae, Campylobacter или любой другой подходящий бактериальный вектор. Способы трансформации живых бактериальных векторов экзогенными ДНК-конструкциями хорошо известны в данной области. См., например, Joseph Sambrook and David W. Russell, Molecular Cloning, A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (2001). Предпочтительные вирусные векторы включают в себя бактериофаги, вирус герпеса, аденовирус, полиовирус, вирус коровьей оспы и Avipox-вирус. Способы трансформации вирусных векторов экзогенными ДНК-конструкциями также хорошо известны в данной области. См., например, Sambrook and Russell, выше. Липосомные векторы представляют собой униламеллярные или мультиламеллярные везикулы, имеющие образованную липофильным материалом мембранную часть и внутреннюю водную часть. В настоящем изобретении водная часть используется для того, чтобы в ней содержался полинуклеотидный материал, который должен быть доставлен к клеткам-мишеням. Обычно предпочтительно, чтобы образующие липосому материалы имели катионную группу, такую как группа четвертичного аммония, и одну или более липофильных групп, таких как насыщенные или ненасыщенные алкильные группы, имеющие приблизительно от 6 и приблизительно до 30 атомов углерода. Группа подходящих материалов описана в Европейской патентной публикации No. 0187702 и дополнительно обсуждается в патенте США No. 6228844, выданном на имя Wolff et al., описание соответствующей части которого включено здесь посредством ссылки. Многие другие подходящие образующие липосому катионные липидные соединения описаны в литературе. См., например, L. Stamatatos, et al., Biochemistry 27: 3917-3925 (1988); и H. Eibl, et al., Biophysical Chemistry 10: 261-271 (1979). Альтернативно можно использовать микросферы, такие как биодеградируемые полиактид-когликолидные микросферы. Конструкция нуклеиновой кислоты инкапсулируется в липосомы или микросферы или иным способом образует с ними комплекс для доставки нуклеиновой кислоты к ткани, как это практикуется в данной области. Методические аспекты согласно изобретению включают в себя стадию введения полинуклеотидов ДНК в ткань млекопитающего, такого как человек. В некоторых предпочтительных воплощениях полинуклеотиды ДНК вводят перорально, внутримышечно, интраназально, внутрибрюшинно, подкожно, внутрикожно или местно. В методических аспектах согласно изобретению ДНК-вакцина может быть использована для обеспечения долговременного ингибирования пролиферации эндотелиальных клеток у пациента, которому введена вакцина. В одном из предпочтительных воплощений данного способа ДНК-вакцина, содержащая полинуклеотидную конструкцию, оперативно кодирующую рецепторный белок VEGF, вводится млекопитающему в случае необходимости ингибирования пролиферации эндотелиальных клеток в количестве, которого достаточно для того, чтобы вызвать иммунный ответ против пролиферирующих эндотелиальных клеток. Настоящим изобретением обеспечивается также способ ингибирования ангиогенеза у млекопитающего, которому введена ДНК-вакцина. В таком воплощении способа вакцина, содержащая ДНК-конструкцию, оперативно кодирующую рецепторный белок VEGF, вводится млекопитающему, страдающему от связанного с ангиогенезом заболевания, в количестве, которого достаточно для того, чтобы вызвать иммунный ответ против пролиферирующих эндотелиальных клеток. В другом методическом аспекте согласно изобретению опухолевый рост ингибируется путем введения млекопитающему ДНК-вакцины. В таком методическом аспекте вызывающее иммунный ответ количество вакцины, содержащей конструкцию ДНК, оперативно кодирующую рецепторный белок VEGF, вводится млекопитающему, у которого наблюдается опухолевый рост. Обработка такой вакциной приводит к остановке опухолевого роста под действием иммунизации млекопитающего против пролиферирующих эндотелиальных клеток. Разрушение пролиферирующих эндотелиальных клеток под воздействием иммунной системы млекопитающего предотвращает или по меньшей мере минимизирует васкуляризацию опухоли. В методических воплощениях согласно изобретению вакцины могут быть введены энтерально, например, путем перорального введения или путем внутримышечной инъекции. В предпочтительном варианте осуществления млекопитающее, которому вводится вакцина согласно изобретению, является человеком. У пациента, страдающего от злокачественной опухоли, такой как карцинома легкого или кишечника, или же от опухолей предстательной железы, диабетической ретинопатии и тому подобного, может настать улучшение от иммунизации вакцинами согласно изобретению. Вакцины согласно изобретению предпочтительно составлены с фармацевтически приемлемыми носителями или эксципиентами, такими как вода, физиологический раствор, декстроза, глицерин и тому подобное, а также с их комбинациями. Вакцины могут содержать также и дополнительные вещества, такие как увлажняющие агенты, эмульсифицирующие агенты, буферы и тому подобное. Вакцины согласно изобретению предпочтительно вводят млекопитающему, такому как человек, перорально, в виде раствора или суспензии в фармацевтически приемлемом носителе, с концентрацией ДНК в области приблизительно от 1 до 10 микрограммов на миллилитр. Приблизительная дозировка будет зависеть от конкретного субъекта, которого предстоит вакцинировать, и отчасти – от практикующего медицинского работника, производящего вакцинацию или назначающего введение вакцины. Вакцины согласно изобретению могут быть запакованы в соответствующие стерилизованные контейнеры, такие как ампулы, бутылки или флаконы, либо во многократной, либо в единичной лекарственной формах. Предпочтительно контейнеры запечатываются герметично после их заполнения вакцинным препаратом. Предпочтительно, когда вакцины упакованы в контейнеры, которые снабжены надписью, позволяющей идентифицировать вакцину, а также несущей информацию в виде этикетки правительственной организации, такой как Управление по контролю за продуктами и лекарствами США, отражающую факт одобрения и защиты применения вакцины соответствующими законами, информацию о дозировке и т.д. Этикетка предпочтительно содержит информацию о вакцине, которая используется для профессионального введения вакцины для поддержания здоровья пациента. Предпочтительно также, чтобы на упаковке были напечатаны информационные материалы относительно введения вакцины, инструкции, указания и любые требуемые необходимые предостережения. Предпочтительно, чтобы вакцины согласно изобретению содержали конструкции ДНК, которые кодируют один или более рецепторных белков VEGF, таких как тирозинкиназа, специфичная в отношении эндотелиальных клеток, включая, например, Flt-1, KDR, Flk-1 и их функциональные гомологи. Функциональные гомологи предпочтительно имеют по меньшей мере 80% гомологию с вышеуказанными рецепторными белками VEGF. Аминокислотные последовательности рецепторных белков VEGF описаны в данной области так же, как и последовательности нуклеиновых кислот, кодирующие эти белки. Последовательность нуклеиновой кислоты, кодирующая KDR (фиг. 1, SEQ ID NO: 1), и соответствующая ей белковая последовательность (фиг. 2, SEQ ID NO: 2) были опубликованы Yu et al. в базе данных EMBL Европейского Института Биоинформатики, Wellcome Trust Genome Campus, Hinxton, Cambridge CB10 1SD, UK (регистационный номер EMBL следующий – EMBL: AF063658), описание которой включено в настоящее описание в виде ссылки. Последовательность нуклеиновой кислоты, кодирующая Flt-1 (фиг. 3, SEQ ID NO: 3), и соответствующая ей белковая последовательность (фиг. 4, SEQ ID NO: 4) были опубликованы Yu et al. в базе данных EMBL Европейского Института Биоинформатики, Wellcome Trust Genome Campus, Hinxton, Cambridge CB10 1SD, UK (регистационный номер EMBL следующий – EMBL: AF063657), описание которой включено в настоящее описание в виде ссылки. Последовательность нуклеиновой кислоты, кодирующая Flk-1, и соответствующая ей белковая последовательность были опубликованы Mathews et al., Proc. Natl. Acad. Sci. USA 1991, 88: 9026-9030, а структуры были откорректированы Quinn et al., Proc. Natl. Acad. Sci. USA 1991, 90: 7533-7537, релевантные описания которой включены в данное описание в виде ссылки. Правильная последовательность ДНК Flk-1 приведена на фиг. 5 как SEQ ID NO: 5, а откорректированная белковая последовательность Flk-1 приведена на фиг. 6 как SEQ ID NO: 6. Из-за вырожденности генетического кода другие последовательности ДНК, которые кодируют по существу такую же самую или функционально эквивалентную аминокислотную последовательность рецепторных белков VEGF, таких как KDR, Flk-1 и Flt-1, могут быть использованы при практиковании настоящего изобретения. Такие последовательности ДНК включают в себя такие последовательности, которые способны гибридизоваться также и с последовательностями рецептора VEGF. Предпочтительно функционально эквивалентные гомологи ДНК рецепторных белков VEGF имели по меньшей мере 80% гомологию с ДНК, кодирующей указанные выше рецепторные белки VEGF. Измененные последовательности ДНК, которые могут быть использованы в соответствии с настоящим изобретением, включают в себя делеции, добавки или замены различных нуклеотидных остатков, что в результате приводит к образованию последовательности, которая кодирует тот же самый или функционально эквивалентный продукт гена. Как таковой этот продукт гена может содержать внутри рецепторных последовательностей VEGF делеции, добавки или замены аминокислотных остатков, которые приводят к “молчащим” изменениям, и, таким образом, к продуцированию функционально эквивалентных рецепторных белков VEGF. Такие аминокислотные замены могут быть произведены на основе сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатической природы вовлеченных остатков. Например, отрицательно заряженные аминокислоты включают в себя аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают в себя лизин и аргинин; аминокислоты с незаряженными полярными боковыми группами, имеющие сходные величины гидрофильности, включают в себя следующие: лейцин, изолейцин, валин; глицин, аланин; аспарагин, глутамин; серин, треонин; фенилаланин, тирозин. Здесь термин “функционально эквивалентный рецептор VEGF” относится к рецептору, который связывается с VEGF или его фрагментом, но не обязательно с той же самой аффинностью связывания, что и с его нативными аналогами KDR, Flk-1 или Flt-1. Последовательности ДНК согласно изобретению могут быть получены генноинженерным путем, чтобы получить измененную последовательность, кодирующую рецептор VEGF, для различных целей, включая, но не ограничиваясь этим, изменения, которые модифицируют процессинг и экспрессию продукта гена. Например, мутации могут быть встроены с помощью технологий, которые хорошо известны в данной области, например, путем сайтнаправленного мутагенеза, для включения новых рестрикционных сайтов, для изменения характера гликозилирования, фосфорилирования и т.д. Мышиный Flk-1 (SEQ ID NO: 6) имеет приблизительно 85% гомологию с человеческим KDR (SEQ ID NO: 2) и играет роль в физиологии мыши, аналогичную таковой KDR у человека. Фактически VEGFR-2 часто относят к KDR/Flk-1, что отражает тесную аналогию между этими двумя гомологами рецептора VEGF. По этой причине обработку мышей ДНК-вакциной согласно изобретению, кодирующей Flk-1 (например, ДНК с последовательностью SEQ ID NO: 5), выбрали в качестве подходящей модели для человеческих ДНК-вакцин, кодирующих KDR. Следующие примеры приведены для дальнейшей иллюстрации признаков и воплощений настоящего изобретения, а не для ограничения ими данного изобретения. Материалы. Методы и примеры. Материалы. Мыши C57/BL/6J и Balb/C были получены из Научно-Исследовательского института Scripps по разведению животных. Для оценки использовали линии мышиных опухолевых клеток, включая линию клеток меланомы B16 и линию клеток карциномы кишечника CT26, все из которых были получены у доктора I. J. Fidler, MD Anderson Cancer Center, Houston, TX. Линия клеток D121 мышиной легочной карциномы Льюиса были получены у доктора Lea Eisenbach, Weizmann Institute, Rehovot, Israel. ДНК, кодирующая Flk-1, была любезно предоставлена доктором Lemischka (Princeton University, Princeton, NJ) и клонирована в эукариотический экспрессирующий вектор pcDNA3.1, предоставленный Invitrogen, Huntsville, Alabama, с использованием рестрикционных сайтов KpnI и Xbal. Аттенюированный штамм Salmonella typhimurium был предоставлен B. A. D. Stocker (Stanford University, Stanford, CA). Антитела были получены от компании BD Biosciences, Bedford, MA. Культуральная добавка T-STIM была получена от компании BD Biosciences, Bedford, MA. Флуоресцеин-изотиоцианат (FITC) и R-Фикоэритрин (PE) были получены от компании Molecular Probes, Eugene, OR. FITC-меченые и PE-меченые антитела были получены в соответствии с методиками, рекомендуемыми изготовителем. ПРИМЕР 1. Получение ДНК-вакцины, кодирующей Flk-1. Вектор pcDNA3.1, содержащий ДНК Flk-1 (SEQ ID NO: 5; приблизительно от 10 пг и приблизительно до 0,1 мкг рДНК), был электропорирован в свежепрепарированную аттенюированную Salmonella typhimurium, с использованием устройства Bio-Rad Pulser при 2,5 кВ, 25 мкФ и 200 Ом, в соответствии с процедурами, рекомендованными изготовителем. Salmonella, содержащие вектор, были отобраны и помещены в ампициллин-содержащие планшеты. Колонии собирали на следующий день, и культивировали их в течение ночи в бульоне LB (EM Science, Gibbstown, NJ) с добавками ампициллина. Бактерии выделяли и промывали в растворе фсфатного буфера (PBS). Отмытые бактерии затем суспендировали в среде PBS в концентрации приблизительно 1 x 109 рекомбинантных Salmonella на миллилитр PBS, с образованием вакцинного раствора для дальнейшего использования. Вакцину хранили в запаянных ампулах вплоть до использования. “Контрольная вакцина”, состоящая из Salmonella, трансформированных одним только вектором pcDNA3.1 (без ДНК Flk-1), была также получена в соответствии с той же самой методикой. Плазмидную ДНК до трансформации Salmonella хранили приблизительно при -80оC. ПРИМЕР 2. Вакцинация мышей ДНК-вакциной, кодирующей Flk-1. Мышей Balb/C (приблизительно по 6 мышей на опытную группу) вакцинировали ДНК-вакциной, полученной в соответствии с примером 1 (приблизительно 1 x 108 рекомбинантных Salmonella приблизительно на 100 микролитров PBS), путем перорального кормления с помощью зонда, три раза с двухнедельными интервалами. Другую группу мышей вакцинировали контрольной вакциной (состоящей из аттенюированной Salmonella, содержащей пустой вектор), в соответствии с той же схемой, что и мышей, вакцинированных вакциной согласно изобретению. ПРИМЕР 3. Оценка резистентности к опухоли у вакцинированных мышей. Приблизительно через две недели после третьей вакцинации мышам Balb/C из Примера 2 (приблизительно по 6 мышей на опытную группу) переносили либо приблизительно 1 x 105 клеток B16 меланомы (подкожно), либо приблизительно 1 x 105 клеток D121 легочной карциномы Льюиса (подкожно), либо приблизительно 7,5 x 104 клеток CT26 карциномы кишечника (внутривенно). Подкожные опухоли легких Льюиса хирургически удаляли приблизительно после двухнедельного роста, создавая условия для спонтанного диссеминирования в легкие. Подкожный опухолевый рост через день измеряли в двух проекциях, и объем опухоли рассчитывали для каждой опухоли в соответствии с формулой: объем =(ширина 2) (длина ÷ 2). Количество спонтанных метастазов D121 в легкие оценивали приблизительно через 30 дней после удаления первичной подкожной опухоли. Мышей забивали и делали вскрытие, и опухолевый нарост легких оценивали в соответствии с процентом легочной поверхности, которая была покрыта опухолью, причем за “0” принимали отсутствие опухоли, за “1” – состояние, при котором опухолью покрыто приблизительно менее 20% легочной поверхности, за “2” – состояние, при котором опухолью покрыто приблизительно от 20 и приблизительно до 30% легочной поверхности, а за “3” – приблизительно более чем 50% легочной поверхности. На фиг. 7 приведена картина легких трех мышей после переноса им клеток D121 легочной карциномы Льюиса. Самое нижнее легкое было классифицировано как 1, тогда как два верхних легких были классифицированы как 3, поскольку там большая часть легочной поверхности была покрыта опухолями. Животные, которые погибли до 30-го дня оценки развития опухолей, были отмечены как “+”. Результаты этих оценок приведены в Таблицах 1-4 и на фиг. 8-10, подробно обсуждаемых ниже.
У мышей Balb/C, которым путем внутривенной инъекции вводили клетки CT-26 карциномы кишечника, смертность оценивали в течение периода более 63 дня (7 недель). Информация о смертности этих мышей представлена в Таблице 2 ниже и графически проиллюстрирована на фиг. 8. На фиг. 8 процент выживших мышей, обработанных вакциной согласно изобретению из Примера 1, показан жирной сплошной линией, соответствующей 100% выживаемости. Процент выживших “наивных” (невакцинированных) мышей, которым вводили клетки C26, показан тонкой сплошной линией, тогда как процент выживших мышей, обработанных контрольной вакциной (не содержащей ДНК Flk-1), показан прерывистой пунктирной линией.
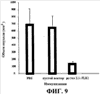
Супрессию роста первичной (подкожной) опухоли у мышей Balb/C, которым переносили клетки D121, оценивали путем определения объема первичной опухоли на день 14 после заражения. Результаты представлены в Таблице 3 ниже и графически проиллюстрированы на фиг. 9. На фиг. 9 первый столбец, отмеченный как “PBS”, относится к мышам, которые не были вакцинированы (“наивные” мыши), средний столбец, отмеченный как “пустой вектор”, относится к мышам, которые были обработаны контрольной вакциной, и третий столбец, отмеченный как “pcDNA3.1-FLK1”, относится к мышам, которые были иммунизированы вакциной согласно изобретению, из Примера 1.
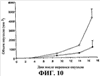
Супрессию опухолевого роста подкожной меланомы B16 оценивали путем мониторинга объема подкожной опухоли приблизительно в течение 17-дневного периода после переноса опухолевых клеток. Результаты представлены в Таблице 4 и графически проиллюстрированы на фиг. 10 ниже. На фиг. 10 приведены данные, соответствующие среднему размеру опухоли, причем (
ПРИМЕР 4. Положительная регуляция маркеров активации CD25, CD69 и CD2 в спленоцитах (CD8+ T-клетках), полученных из вакцинированных мышей. Мышей C5/7BL/6J (приблизительно по 4 мыши на опытную группу) вакцинировали ДНК-вакциной из Примера 1 и контрольной вакциной (без Flk-1), как описано в Примере 2. Спленоциты выделяли из иммунизированных мышей и из контрольной группы мышей приблизительно через шесть недель после третьей вакцинации. Клетки-спленоциты культивировали в течение 24 часов вместе с клетками линии B16 клеток меланомы, трансдуцированными для экспрессии Flk-1, и с нетрансформированными клетками B16 в T-клеточной среде (приблизительно 5 мл на культуру), содержащей приблизительно 4% по объему культуральной добавки T-STIM (Cat.# 354115, BD Biosciences, Bedford, MA). Затем клетки окрашивали FITC-конъюгированным CD8+-антителом и PE-конъюгированными антителами против CD25, CD69 и CD2. Клеточные суспензии оценивали с помощью установки Becton Dickenson FAC scan для определения процента CD8+-T-клеток, положительных в отношении CD25 и CD69, для каждой комбинации спленоцит/клетка B16 меланомы. Результаты представлены в Таблице 5 и графически проиллюстрированы на фиг. 11.
Результаты, приведенные в Таблицах 1-5 и на фиг. 8-11, показывают, что ДНК-вакцина из Примера 1, содержащая ДНК, кодирующую Flk-1, мышиный аналог KDR, может эффективно иммунизировать мышей против целого ряда образующих опухоль раковых клеток. Хоть авторы и не желают связывать себя определенной теорией, тем не менее очевидно, что вакцина действует таким образом, что ингибирует ангиогенез в опухоли, т.е. предотвращает образование новых кровеносных сосудов и способствует эффективному истощению опухоли. Данные, представленные в Таблице 1, показывают, что вакцина согласно изобретению из Примера 1 приводит к супрессии опухолевых метастазов в легкие мышей, зараженных клетками D121 легочной карциномы Льюиса. Ни одна из мышей, иммунизированных вакциной из Примера 1, не умерла, и у всех этих мышей наблюдали приблизительно менее чем 50% поражение легких опухолью (у 2 животных оно составляло < 20%). И наоборот, по 2 мыши умерли в каждой из контрольных групп, и у всех из оставшихся мышей область легких, пораженная опухолью, составляла приблизительно более 50%. Вакцина согласно изобретению из Примера 1 также способствовала значительному снижению смертности мышей Balb-C, которым внутривенно были введены клетки CT-26 карциномы кишечника, как показывают данные, приведенные в Таблице 2 и на фиг. 8. Все мыши, иммунизированные вакциной из Примера 1, выживали в течение всего 63-дневного периода наблюдения после заражения. В контрольных группах, однако, все мыши погибли к 36-му дню после заражения. Как показывают данные, приведенные в Таблице 3 и на фиг. 9, подкожный опухолевый рост клеток D121 легочной карциномы Льюиса в несколько раз тормозился под действием иммунизации вакциной согласно изобретению из Примера 1 – приблизительно от 4,3 до 4,5 раз по сравнению с контрольными группами мышей, не обработанных вакцинами или же обработанных контрольной вакциной. Сходным образом, как показано в Таблице 4 и на фиг. 10, подкожный опухолевый рост клеток B16 меланомы тормозился приблизительно в 4 раза у мышей, иммунизированных вакциной согласно изобретению из Примера 1, по сравнению с опухолевым ростом в контрольной группе. Данные, приведенные в Таблице 5 и на фиг. 11, показывают, что спленоциты, выделенные у мышей C57/BL/6J, вакцинированных ДНК-вакциной из Примера 1, вызывали положительную регуляцию маркеров активации CD2, CD25 и CD69, по сравнению с контрольной группой мышей, когда культивировались с клетками B16 меланомы, трансформированными для представления антигена Flk-1. Множество вариаций и модификаций описанных выше воплощений может быть осуществлено в рамках объема настоящего изобретения. Совершенно очевидно, что примеры специфических воплощений изобретения приведены для иллюстрации изобретения и никоим образом не подразумевают его ограничения. Подразумеваются, конечно, всевозможные модификации из тех, которые заключены в рамках объема, охватываемого прилагаемой формулой изобретения.
Формула изобретения
1. ДНК-вакцина, эффективная для ингибирования пролиферации эндотелиальных клеток, содержащая ДНК-конструкт, оперативно кодирующий рецепторный белок VEGF, встроенный в аттенюированный бактериальный вектор, где рецепторный белок VEGF, кодируемый ДНК-конструктом, выбран из группы, состоящей из VEGFR-2 (KDR; SEQ ID NO: 2), VEGFR-1 (Flt-1; SEQ ID NO: 4), Flk-1 (SEQ ID NO: 6) и их функционального эквивалента, который приблизительно на 80% им гомологичен. 2. ДНК-вакцина по п.1, где аттенуированный бактериальный вектор выбран из группы, состоящей из аттенуированных Salmonella typhimurium, Salmonella typhi, Shigella, Bacillus, Lactobacillus, BCG, Escherichia coli, Vibrio cholerae и Campylobacter. 3. ДНК-вакцина по п.1, где аттенуированный бактериальный вектор представляет собой аттенуированную Salmonella typhimurium. 4. ДНК-вакцина по любому из пп.1-3, где ДНК-конструкт содержит полинуклеотидную последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO: 1, SEQ ID NO: 3, SEQ ID NO: 5 и их функционального гомолога, который, по меньшей мере, приблизительно на 80% им гомологичен. 5. Способ ингибирования пролиферации эндотелиальных клеток у млекопитающего, включающий в себя стадию введения млекопитающему эффективного для стимуляции иммунного ответа количества ДНК-вакцины, охарактеризованной в любом из пп.1-4, при этом у указанного млекопитающего проявляется иммунный ответ, вызванный вакциной и специфичный в отношении пролиферирующих эндотелиальных клеток. 6. Способ по п.5, где млекопитающим является человек. 7. Способ по любому из пп.5 и 6, согласно которому вакцину вводят перорально. 8. Способ ингибирования ангиогенеза у млекопитающего, включающий введение указанному млекопитающему иммунологически эффективного количества ДНК-вакцины по любому из пп.1-4, при этом у указанного млекопитающего проявляется иммунный ответ, вызванный вакциной и специфичный в отношении пролиферирующих эндотелиальных клеток, приводящий к ингибированию образования кровеносных сосудов. 9. Способ по п.8, где млекопитающим является человек. 10. Способ по любому из пп.8 и 9, согласно которому вакцину вводят перорально. 11. Способ ингибирования опухолевого роста у млекопитающего, включающий в себя введение указанному млекопитающему иммунологически эффективного количества ДНК-вакцины по любому из пп.1-4, при этом у указанного млекопитающего проявляется иммунный ответ, вызванный вакциной и специфичный в отношении пролиферирующих эндотелиальных клеток, приводящий в результате к остановке опухолевого роста, уменьшению размеров опухоли или ингибированию распространения опухоли. 12. Способ по п.11, где млекопитающим является человек. 13. Способ по любому из пп.11 и 12, согласно которому вакцину вводят перорально. 14. Готовая форма ДНК-вакцины по любому из пп.1-4, запакованной в герметически запаянный стерильный контейнер, имеющий прикрепленную к нему этикетку, на которой напечатан материал, идентифицирующий вакцину и обеспечивающий информацию, используемую для индивидуального введения указанной вакцины пациенту.
РИСУНКИ
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 ), по сравнению с контрольной группой (o); и
), по сравнению с контрольной группой (o); и