Патент на изобретение №2315807
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) СПОСОБ ПОЛУЧЕНИЯ НОРЛЕЙЦИНА И НОРВАЛИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ РОДА Escherichia, В КОТОРОЙ ВСЕ СИНТАЗЫ АЦЕТОГИДРОКСИКИСЛОТ ИНАКТИВИРОВАНЫ
(57) Реферат:
Изобретение относится к биотехнологии и представляет собой способ получения непротеиногенных аминокислот, таких как норлейцин и норвалин, с использованием бактерии, принадлежащей к роду Escherichia, у которой все синтазы ацетогидроксикислот (AHASы) инактивированы. Изобретение позволяет получать норлейцин и норвалин с высокой степенью эффективности. 2 н. и 2 з.п. ф-лы, 7 ил., 3 табл.
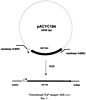
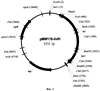
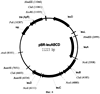
Область техники Настоящее изобретение относится к микробиологической промышленности, в частности к способу продукции непротеиногенных аминокислот, таких как норлейцин и норвалин, с использованием бактерии семейства Enterobacteriaceae, в которой все синтазы ацетогидроксикислот (AHASы) инактивированны. Предшествующий уровень техники Известно, что норлейцин в бактериальной клетке является аналогом метионина (Lawrence, J.Bacteriol., 109, 8-11 (1972)). Была показана его способность встраиваться в белки вместо метионина (Barker, D. and Bruton, С., J.Mol. Biol., 133, 217-31 (1979), Munier, R. and Cohen, G., Biochim. Biophys. Acta., 31, 378-90 (1959); Bogosyan, G. et al., J.Biol. Chem., 264,1, 531-539, (1989); патент США 5622845; патент США 5599690). Также известно, что конкурентное ингибирование внутриклеточной утилизации метионина за счет поступающего извне норлейцина останавливает рост Е.coli (Harris, J. S. and Kohn, H. I., J. Pharmacol., 73, 383-400 (1941)). При добавлении норлейцина наблюдалось повышение скорости роста метионинового ауксотрофа Е.coli в среде, содержащей метионин в концентрации, близкой к оптимальной (Lampen, J. О. and Jones, M. J., Arch. Biochem. Biophys., 13,47-53 (1947)). Описан способ получения норлейцина с использованием штамма Е.coli с высоким уровнем синтеза богатых лейцином белков, выращиваемого в ферментационной среде с низкой концентрацией аминокислот (патент США 5622845). Был получен мутантный штамм S. marcescens с дерепрессированными ферментами пути биосинтеза лейцина, на основе которого в дальнейшем был отобран изолейцинзависимый мутант, не имеющий активности треониндегидратазы. Далее, на основе этого двойного мутанта был отобран изолейцин-независимый (не содержащий активности треониндегидратазы) мутант с устойчивой к ингибированию по типу обратной связи альфа-изопропилмалатсинтазой. Полученный мутант был способен накапливать норлейцин. Было предложено, что норлейцин образуется из пирувата или 2-кетобутирата (вместо субстрата пути биосинтеза лейцина – 2-кетоизовалерата) (Фиг.1) в Serratia marcesens (Kisumi et al., J.Biochem., 1976, 80, 333-339) и Е.coli (Bogosyan, G., et al., J.Biol. Chern., 264,1,531-539 (1989)). Описан способ получения белков, содержащих непротеиногенные аминокислоты путем культивирования микроорганизма-продуцента белка, в частности Е.coli, в культуральной жидкости, содержащей непротеиногенные аминокислоты, включая норлейцин и норвалин (патент США 4879223). Включение норлейцина в продукты рекомбинантных генов может также осуществляться путем культивирования клеток, экспрессирующих ген, кодирующих целевой продукт в среде, содержащей норлейцин, или являющихся ауксотрофами по лейцину и/или метионину. Таким образом, эта методика может использоваться для получения аналогов норлейцина – IL-2, GCSF, гамма-интерферона и альфа-консенсус интерферона (патент США 5599690). Но в настоящее время нет данных о накоплении норлейцина и норвалина с использованием штаммов Е.coli, в которых отсутствуют активности AHAS. Краткое описание чертежей На Фиг.1 показана схема биосинтеза норлейцина и норвалина, включающая процесс удлинения углеродной цепи кетокислот. На Фиг.2 показана предполагаемая схема биосинтеза непротеиногенных аминокислот в штамме Е.coli, в котором отсутствует активность AHAS. KIV – 2-кетоизовалериат; PYR – пируат; KB – 2-кетобутират; KV – 2-кетовалериат; KIC – 2-кетоизокапроат; КС – 2-кетокапроат; IPMS – изопропилмалатсинтаза; IPMI – изопропилмалатизомераза; IPMD – изопропилмалатдегидрогеназа. На Фиг.3 показано относительное расположение праймеров ilvBN1 и ilvBN2 на плазмиде pACYC184. На Фиг.4 показана схема конструирования хромосомного фрагмента ДНК, содержащего инактивированный оперон ilvBN. На Фиг.5 показана плазмида pMW118-ilvIH. На Фиг.6 показана плазмида pBR-leuABCD. На Фиг.7 показано разделение на ВЖКХ: а) стандартной смеси аминокислот; b) стандартной смеси аминокислот, содержащей 0.06 мМ норвалина и норлейцина; с) ферментационный бульон. Описание изобретения Целью настоящего изобретения является предоставление штаммов-продуцентов непротеиногенных аминокислот и предоставление способа получения таких непротеиногенных аминокислот с использованием указанных штаммов. Данная цель была достигнута путем установления того факта, что инактивация AHAS может повысить продукцию непротеиногенных аминокислот, таких как норлейцин и норвалин. Далее, было показано, что усиленная экспрессия leuABCD оперона повышает продукцию непротеиногенных аминокислот штаммами с инактивированными АНАЗами. Таким образом было совершено настоящее изобретение. Настоящее изобретение представляет бактерию семейства Enterobacteriaceae, обладающую способностью к продукции непротеиногенных аминокислот, таких как норлейцин и норвалин. Целью настоящего изобретения является создание бактерии-продуцента непротеиногенных аминокислот семейства Enterobacteriaceae, при этом указанная бактерия модифицирована таким образом, что все синтазы ацетогидроксикислот (AHASы) инктивированы. Также целью настоящего изобретения является создание описанной выше бактерии, где непротеиногенными аминокислотами являются норлейцин и/или норвалин. Также целью настоящего изобретения является создание описанной выше бактерии, при этом указанная бактерия принадлежит к роду Escherichia. Также целью настоящего изобретения является создание описанной выше бактерии, в которой синтазы ацетогидроксикислот включают в себя AHAS I, кодируемую генами UvBN, AHAS II, кодируемую генами UvGM, и AHAS III, кодируемую генами UvIH. Также целью настоящего изобретения является создание описанной выше бактерии, при этом указанная бактерия модифицирована таким образом, что экспрессия оперона leuABCD усилена. Также целью настоящего изобретения является предоставление способа получения непротеиногенных аминокислот, при этом указанный метод включает: – выращивание указанной бактерии в питательной среде, которая не содержит L-лейцин, но содержит L-валин в концентрации, лимитирующей рост бактерии, с целью продукции и накопления непротеиногенных аминокислот в питательной среде, и – выделение полученной непротеиногенной аминокислоты из культуральной жидкости. Также целью настоящего изобретения является создание описанного выше способа, в котором непротеиногенной аминокислотой является норлейцин и/или норвалин. Детально настоящее изобретение будет описано ниже. Наилучший способ осуществления изобретения. 1. Бактерия согласно настоящему изобретению. Бактерией согласно настоящему изобретению является бактерия-продуцент непротеиногенной аминокислоты, принадлежащая к семейству Enterobacteriaceae, при этом указанная бактерия модифицирована таким образом, что все AHASы в клетке инактивированы. Термин “непротеиногенные аминокислоты” согласно настоящему изобретению означает аминокислоты, отличные от 20 основных протеиногенных аминокислот (L-аланин, L-аргинин, L-аспарагин, L-аспартат, L-цистеин, L-глутамат, L-глутамин, L-глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин). Такие непротеиногенные аминокислоты могут быть синтезированы из 2-кето-предшественника ферментами пути биосинтеза L-лейцина, кодируемого leuABCD опероном. Примером “непротеиногенной аминокислоты” согласно настоящему изобретению являются норвалин, синтезируемый из 2-кетовалериата, и норлейцин, синтезируемый из 2-кетовалериата через 2-кетокалроат. Другим примером непротеиногенной аминокислоты” согласно настоящему изобретению являются гомолейцин, синтезируемый из 2-кетоизокапроата и гомоизолейцин, синтезируемый из 2-кето-3-метилвалериата с помощью ферментов пути биосинтеза L-лейцина (смотри, например, Jakubowski, U. and Goldman, E., Microbiological reviews, v.56, No. 3, р.412-429 (1992)). Другие химические гомологи вышеупомянутых аминокислот также являются частью настоящего изобретения. Согласно настоящему изобретению термин “бактерия-продуцент непротеиногенной аминокислоты” означает бактерию, обладающую способностью к продукции и накоплению непротеиногенной аминокислоты в питательной среде, в условиях, когда бактерия согласно настоящему изобретению выращивается в указанной питательной среде. Способность к продукции непротеиногенной аминокислоты может быть придана или улучшена путем селекции. Используемый здесь термин «бактерия-продуцент непротеиногенной аминокислоты» также означает бактерию, которая способна к продукции полезного метаболита и накоплению метаболита в ферментационной среде в количествах больших, чем природный или родительский штамм E. coli, такой как штамм E. coli К-12, W3110 или MG1655. Предпочтительно, этот термин означает бактерию, которая способна к продукции и накоплению непротеиногенной аминокислоты в ферментационной среде в количествах не менее, чем 10 мг/л, более предпочтительно, не менее, чем 50 мг/л целевой аминокислоты. Семейство Enterobacteriaceae включает в себя бактерии, принадлежащие к родам Escherichia, Erwinia, Providencia и Serratia. Род Escherichia предпочтителен. Термин “бактерия, принадлежащая к роду Escherichia” означает, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, использованного в настоящем изобретении, может быть упомянута бактерия Escherichia coli (E.coli). Круг бактерий, принадлежащих к роду Escherichia, которые могут быть использованы в настоящем изобретении, не ограничен каким-либо образом, однако, например, бактерии, описанные в Neidhardt, F.C. et al. (Escherichia coli and Salmonella typhimurium, American Society for Microbiology, Washington D.C., 1208, Таблица 1) могут быть упомянуты. Термин “AHAS инактивирована” означает, что ген, кодирующий AHAS, модифицирован таким образом, что такой модифицированный ген кодирует мутантный белок со значительно сниженным уровнем активности, в частности с уровнем активности, не измеряемым традиционно используемыми для этого приборами, или такой ген кодирует полностью неактивный белок. Также возможно, что модифицированная область ДНК не способна к экспрессии гена естественным образом, из-за делеции части этого гена, сдвига рамки считывания этого гена, или модификации областей, прилегающих к оперону, включая последовательности, контролирующие экспрессию оперона, такие как промотор(ы), энхансер(ы), аттенуатор(ы) и т.д. AHASы согласно настоящему изобретению включают в себя AHAS I, II и III. AHAS I (ацетогидроксибутаноатсинтаза I / ацетолактатсинтаза I), AHAS II (ацетогидроксибутаноатсинтаза II / ацетолактатсинтаза II) и AHAS III (ацетолактатсинтаза III / ацетогидроксибутаноатсинтаза III) катализируют две реакции, продуктом которых являются 2-ацето-2-гидроксибутират, образующийся из пирувата и 2-оксобутаноата, и ацетолактат, образующийся из пирувата. AHAS I кодируется генами ilvBN (ilvBN оперон), AHAS II кодируется генами ilvGM (UvGM оперон) и AHAS III кодируется генами UvIH(ilvIH оперон). Ген UvB (номера нуклеотидов с 3850411 по 3848723 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1) и ген HvN (номера нуклеотидов с 3848719 по 3848429 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1) расположены между генами uhpA и ivbL на хромосоме штамма E. coli К-12 и кодируют каталитическую и регуляторную субъединицы AHASbi I соответственно. Ген ilvG (состоит из двух нуклеотидных последовательностей с 3948183 по 3949163 и с 3949162 по 3949827 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1) и ген ilvM (номера нуклеотидов с 3949824 по 3950087 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1) расположены между генами ilvL и ilvE на хромосоме штамма Е.coli К-12 и кодируют большую и малую субъединицы AHASы II соответственно. Ген ilvl (номера нуклеотидов с 85630 по 87354 в базе данных GenBank, последовательность с инвентарньм номером NC_000913.1) и ген ilvH (номера нуклеотидов с 87357 по 87848 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1) расположены между генами leuO и fruR на хромосоме штамма Е.coli К-12 и кодируют большую и малую субъединицы AHASы III соответственно. Инактивация указанного гена может быть осуществлена традиционными методами, такими как мутагенез с использованием УФ излучения или обработка нитрозогуанидином (N-метил-N’-нитро-N-нитрозогуанидин), сайт-направленный мутагенез, разрушение гена с использованием гомологичной рекомбинации и/или инсерционно-делеционный мутагенез (Datsenko K.A. and Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97:12: 6640-45), также известной как “Red-зависимая интеграция”. Мутантный белок, активность которого значительно снижена, или полностью неактивный белок также могут быть получены, например, с помощью сайт-специфичного мутагенеза нуклеотидной последовательности, кодирующей аминокислотные остатки, расположенные в консервативных областях AHAS. Выравнивание аминокислотных последовательностей AHAS нескольких видов и данные о структурно-функциональной корреляции представлены в обзоре Duggleby, R.G. и Pang, S.S. (J.Biochem. Mol. Biol., 33, No. 1,1-36 (2000)). Установлено, например, что в AHAS из Е.coli глутамат в 47 позиции играет каталитическую роль. Таким образом, мутация в этой позиции может оказаться критической для ферментативной активности AHAS из Е.coli. Бактерия согласно настоящему изобретению может быть в дальнейшем модифицирована таким образом, что экспрессия лейцинового оперона leuABCD усилена. Оперон leuABCD включает в себя гены leuA, leuB, leuC и leuD. В указанном опероне, ген leuA кодирует Ген leuA (номера нуклеотидов с 83529 по 81958 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1), ген leuB (номера нуклеотидов с 81958 по 80867 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1), ген leuC (номера нуклеотидов с 80864 по 79464 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1) и ген leuD (номера нуклеотидов с 79453 по 78848 в базе данных GenBank, последовательность с инвентарным номером NC_000913.1) расположены между геном leuL и открытой рамкой считывания yabMOW на хромосоме штамма Е.coli К-12. Примерами методик, используемых для усиления экспрессии генов, особенно методик, используемых для повышения количества белковых молекул в бактериальной клетке, являются методики изменения регуляторной последовательности ДНК, регулирующей экспрессию гена, кодирующего белок, и увеличения количества копий гена в клетке, но не ограничиваются ими. Изменение регуляторной последовательности ДНК, регулирующей экспрессию гена, может быть достигнуто путем помещения ДНК, кодирующей белок согласно настоящему изобретению под контроль сильного промотора. В качестве сильных промоторов известны, например, lac промотор, trp промотор, trc промотор, pr или pl промоторы фага лямбда. Кроме того, промотор может быть усилен, например, путем введения мутации в указанный промотор с целью повышения уровня транскрипции гена, расположенного после промотора. Далее, известно, что замена нескольких нуклеотидов в участке между местом связывания рибосомы (RBS) и старт кодоном, а в особенности, в последовательности непосредственно перед старт-кодоном, в значительной степени влияет на транслируемость мРНК. Например, было обнаружено 20-кратное изменение уровня экспрессии в зависимости от природы трех нуклеотидов, предшествующих старт кодону (Gold et al., Annu. Rev.Microbiol., 35,365-403,1981; Hui et al., EMBO J., 3,623-629, 1984). Кроме того, для повышения уровня транскрипции целевого гена в хромосому может быть введен энхансер. Введение ДНК, содержащей либо ген, либо промотор в хромосомную ДНК, описано, например в выложенной патентной заявке Японии 1-215280 (1989). С другой стороны, к методам увеличения числа копий гена относится клонирование целевого гена на многокопийном векторе с образованием рекомбинантной ДНК, с последующим введением полученной рекомбинантной ДНК в микроорганизм. Для этой цели предпочтительно использовать многокопийные векторы. Примеры многокопийных векторов включают в себя векторы pBR322, pUC19, pBluescript KS+, pACYC177, pACYC184, pAYC32, pMWl 19, pET22b и подобные им, но не ограничиваются ими. Используемый здесь термин «многокопийный вектор» используется для векторов, количество которых достигает 15-30 копий на клетку. В качестве метода трансформации может быть использован любой описанный к настоящему времени метод. Например, в настоящем изобретении может быть использован метод обработки клеток-реципиентов с помощью хлорида кальция для увеличения проницаемости ДНК через клеточную мембрану, описанный для Escherichia coli К-12 (Mandel, M. and Higa, A., J. Mol. Biol., 53, 159 (1970)). Усиление экспрессии гена может быть также достигнуто путем введения множественного числа копий указанного гена в хромосомы бактерии, например, методом гомологичной рекомбинации, Ми интеграции и подобными им методами. Например, один раунд Ми интеграции позволяет встроить в бактериальную хромосому до трех копий интегрируемого гена. Описанные выше методики, использующие сильный промотор или энхансер могут быть скомбинированы с методиками, основанными на увеличении количества копий или экспрессии целевого гена. Бактерия согласно настоящему изобретению может быть получена путем усиления экспрессии оперона leuABCD бактерии, в которой уже инактивированы все АНАSы. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем инактивации всех AHAS в бактерии, уже наследственно обладающей усиленной экспрессией оперона leuABCD. В частности, любой известный штамм-продуцент L-лейцина может быть использован в качестве родительского для придания ему способности к продукции непротеиногенных аминокислот, таких как норлейцин и норвалин. Примерами бактерий, принадлежащих к роду Escherichia и способных к продукции L-лейцина, являются штаммы Е.coli, такие как Н-9068 (АТСС 21530), Н-9070 (FERM ВР-4704) и Н-9072 (FERM ВР-4706), устойчивые к 4-азалейцину или 5,5,5-трифлуоролейцину (патент США 5744331), штаммы Е.coli, в которых отсутствует ингибирование изопропилмалатсинтазы L-лейцином по типу обратной связи (Европейский патент ЕР 1067191), штамм Е.coli АЛ 1478, устойчивый к Методы получения плазмидных ДНК, расщепления и дотирования ДНК, трансформации, подбора олигонуклеотидов в качестве праймеров и подобные им являются традиционными методами, хорошо известными специалисту в данной области. Эти методы описаны, например, в Sambrook, J., Fritsch, E.F., and Maniatis, Т., “Molecular Cloning A Laboratory Manual, Second Edition”, Cold Spring Harbor Laboratory Press (1989). 2. Способ согласно настоящему изобретению. Способом согласно настоящему изобретению является способ получения непротеиногенных аминокислот, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде, не содержащей L-лейцина, но содержащей L-валин в концентрации, лимитирующей рост бактерии, с целью продукции и накопления аминокислоты в питательной среде и выделения аминокислоты из культуральной жидкости. Более конкретно, способом согласно настоящему изобретению является способ получения норлейцина и/или норвалина, включающий этапы выращивания бактерии согласно настоящему изобретению в питательной среде, не содержащей L-лейцина, но содержащей L-валин в концентрации, лимитирующей рост бактерии, с целью продукции и накопления норлейцина и/или норвалина в питательной среде, и выделения норлейцина и/или норвалина из культуральной жидкости. Штаммы, дефицитные по AHASам, не способны расти в отсутствие валина и изолейцина в ферментационной среде (ауксотрофность по валину и изолейцину). Ферментационная среда не должна содержать L-лейцин, чтобы избежать репрессии лейцинового оперона (leuABCD оперон). Недостаток L-лейцина, необходимый для роста клеток, компенсируется присутствием L-валина, который частично деаминируется в 2-кетоизовалериат – предшественник L-лейцина. С другой стороны, для комплементации ауксотрофности по L-изолейцину, в ферментационную среду необходимо добавить значительное количество L-изолейцина. Термин “валин в концентрации, лимитирующей рост бактерии” означает, что концентрации валина, добавленного в ферментационную среду, недостаточно для полного покрытия потребности в валине у растущей биомассы в отсутствие биосинтеза валина. Таким образом, рост данного штамма будет ограничен доступностью валина. Согласно настоящему изобретению, этапы выращивания, выделения и очистки непротеиногенной аминокислоты из среды и подобные им могут быть произведены в соответствии с традиционными способами ферментации, в которых аминокислоту получают с использованием бактерии. Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, а также различные органические кислоты. В зависимости от характера ассимиляции используемого микроорганизма, могут использоваться спирты, такие как этанол и глицерин. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин и дрожжевой экстракт. Выращивание осуществляется предпочтительно в аэробных условиях, таких как перемешивание культуральной жидкости на качалке, взбалтывание с аэрацией, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 30 до 38°С. рН среды поддерживают в пределах от 5 до 9, предпочтительно от 6.5 до 7.2. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно, выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости. После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем L-аминокислоты могут быть выделены и очищены методами ионообменной хроматографии, концентрирования и кристаллизации. Наилучший способ осуществления изобретения. Более детально настоящее изобретение будет описано ниже со ссылками на Примеры. Пример 1. Конструирование штамма с инактивированными генами всех апетолактатсинтетаз. В отличие от других природных штаммов Е.coli, штамм Е.coli К-12 содержит мутацию с полярным эффектом – сдвиг рамки считывания в гене ilvG (Lawther, R. P. et al, Proc. Natl. Acad. Sci. USA 78:922-925,1981), Таким образом, для получения штамма, в котором все AHASы инактивированы, на основе штамма Е.coli К-12 необходимо инактивировать только два оперона, ilvBN и ilvIH. 1. Деления оперона ilvBN. Делеция оперона ilvBN была осуществлена с применением методики, впервые разработанной Datsenko и Wanner (Proc. Natl. Acad. Sci. USA, 2000, 97(12), 6640-6645), так называемой “Red-зависимой интеграцией”. В соответствии с указанной методикой, были синтезированы ПЦР праймеры ilvBNl (SEQ ID NO: 1) и ilvBN2 (SEQ ID NO: 2), гомологичные как областям хромосомы, примыкающим к оперону ilvBN, так и гену, сообщающему устойчивость к хлорамфениколу на плазмиде, используемой в качестве матрицы. Плазмида pACYC184 (NBL Gene Sciences Ltd., Великобритания) (GenBank/EMBL инвентарный номер Х06403) использовалась в качестве матрицы для ПЦР. Использовались следующие условия для ПЦР: денатурация в течение 3 мин при 95°С; температурный профиль для начальных двух циклов: 1 мин при 95°С, 30 сек при 34°С, 40 сек при 72°С; температурный профиль для последующих 30 циклов: 30 сек при 95°С, 30 сек при 50°С, 40 сек при 72°С; и заключительная полимеризация: 5 мин при 72°С. Полученный ПЦР продукт длиной в 935 п.о. (SEQ ID NO: 3) был очищен в агарозном геле и использовался для электропорации штамма Е.coli – Е.coli К 12 (ВКПМ В-7), содержащего плазмиду pKD46 с температурочувствительной репликацией. Плазмида pKD46 (Datsenko and Wanner, Proc. Natl. Acad. Sci. USA, 2000,97:12:6640-45) содержит 2,154 п.о. (31088-33241) фрагмент ДНК фага Х (GenBank инвентарный номер J02459), и содержит гены Электрокомпетентные клетки были получены следующим образом: ночная культура Е.coli K12 (ВКПМ В-7), выращенная при 30°С в LB среде с добавками ампициллина (100 мг/л), была разведена в 100 раз в 5 мл среды SOB (Sambrook et al, “Molecular Cloning A Laboratory Manual, Second Edition”, Cold Spring Harbor Laboratory Press (1989)) с ампициллином и L-арабинозой (1 мМ). Полученную культуру выращивали с аэрацией при 30°С до OD600 2. Подтверждение делений оперона ilvBNc помощью ПЦР. Мутанты, содержащие делецию оперона ilvBN, маркированного геном устойчивости к Cm, проверяли с помощью ПЦР. Локус-специфичные праймеры ilvBNC3 (SEQ ID NO: 4) и ilvBNC4 (SEQ ID NO: 5) использовались для проверочного ПЦР. Для ПЦР проверки использовались следующие условия: денатурация в течение 3 мин при 94 °С; температурный профиль для 30 циклов: 30 сек при 94°С, 30 сек при 53°С, 1 мин при 72°С; и заключительная полимеризация: 7 мин при 72°С. Длина ПЦР продукта, полученного в данной реакции с использованием хромосомной ДНК родительского штамма ilvBN+В-7 в качестве матрицы, составляла 2137 п.о. (Фиг. 4). Длина ПЦР продукта, полученного в данной реакции с использованием хромосомной ДНК мутантного штамма В-7 3. Деления оперона ilvIH. Делеция оперона ilvIH производилась таким же образом, как и делеция оперона ilvBN, описанная в Части 1 данного Примера. В соответствии с методикой, были синтезированы ПЦР праймеры ilvIHI1 (SEQ ID NO: 7) и ilvIHI2 (SEQ ID NO: 8), гомологичные как фрагментам хромосомы, прилегающим к оперону UvIH, так и гену, сообщающему устойчивость к канамицину из плазмиды-матрицы. Плазмида pACYC 177 (NBL Gene Sciences Ltd., Великобритания) (GenBank/EMBL инвентарный номер Х06402) использовали в качестве матрицы для ПЦР. Использовались следующие условия для ПЦР: денатурация в течение 3 мин при 95°С; температурный профиль для начальных двух циклов: 1 мин при 95°С, 30 сек при 34°С, 40 сек при 72°С; температурный профиль для последующих 30 циклов: 30 сек при 95°С, 30 сек при 50°С, 40 сек при 72°С; и заключительная полимеризация: 5 мин при 72°С. Полученный ПЦР продукт, длиной 1370 п.о. (SEQ ID NO: 9) был очищен в агарозном геле и использован для электропорации штамма Е.coli B-7 Пример 2. Продукция и экскреция норлейцина и норвалина у штамма Е.coli NS1118. Штаммы Е.coli К-12 (ВКПМ В-7) и NS118 выращивали в течение 18 часов при 37°С на чашках с L-агаром. Затем клетки, собранные примерно с 0.5 см2 поверхности чашки, переносили в ферментационную среду (2 мл) и выращивались в течение 72 часов при 32°С. Накопленные в среде количества норвалина и норлейцина измерялись с помощью ВЖКХ. Для подробного описания процесса измерений смотри Пример 6. Полученные результаты представлены в Таблице 1. Состав ферментационной среды (г/л):
Как видно из Таблицы 1, штамм Е.coli NS 1118, в котором все AHASы инактивированы, в отличие от природного родительского штамма В-7, продуцирует и выделяет в ферментационную среду норвалин и норлейцин. Добавление лейцина прекращает продукцию и выделение непротеиногенных аминокислот, преимущественно из-за ингибирования изопропилмалатсинтазы. Добавление валина в лимитирующих рост концентрациях необходимо для продукции непротеиногенных аминокислот. В данном случае, отсутствие ацетолактата приводит к увеличению пула 2-кетобутирата. Лимит по валину выражается в отсутствии 2-кетоизовалериата, что, в свою очередь, провоцирует ослабление синтеза лейцина. Лимит по лейцину дерепрессирует лейциновый оперон. 2-кетобутират используется изопропилмалатсинтазой как альтернативный субстрат (вместо 2-кетоизоалериата) и в дальнейшем используеся для синтеза норвалина и норлейцина с помощью других ферментов, кодируемых лейциновым опероном (Фиг.1). Повышение концентрации валина в исходной ферментационной среде также прекращает синтез норлейцина и норвалина за счет увеличения соотношения кетоизовалериат/2-кетобутират. Таким образом, для синтеза таких непротеиногенных аминокислот необходимы специфичные условия роста: отсутствие лейцина и присутствие валина в ферментационной среде. Также валин должен присутствовать в концентрациях, лимитирующих рост бактерии. Пример 3. Клонирование AHASIII. Оперон ilvIH клонировали в вектор pMW118 в виде ПЦР фрагмента длиной 2764 п.о. Хромосома штамма MG1655 использовалась как матрица для ПЦР. В качестве праймеров использовались синтетические олигонуклеотиды ilvIHL58 (SEQ ID NO: 13) и ilvIHR60 (SEQ ID NO: 14). Праймер ilvIHL58 содержит введенный в 5′-конец сайт рестрикции Sad, и праймер ilvIHR60 содержит введенный в 5′-конец сайт рестрикции Xbal. Использовались следующие условия для ПЦР: денатурация в течение 3 мин при 94°С; температурный профиль для 30 циклов: 30 сек при 94°С, 30 сек при 57°С, 2 мин при 72°С; и заключительная полимеризация: 7 мин при 72°С. Полученный ПЦР продукт длиной в 2778 п.о. был очищен в агарозном геле, обработан рестриктазами SacI и Xbal и клонирован в вектор pMW118, предварительно обработанный теми же рестриктазами. В качестве реципиента для клонирования использовался штамм NS1118. Полученная плазмида pMWl 18-ilvIH (Фиг.5) комплементирует фенотип AHAS” штамма NS 1118. Пример 4. Клонирование лейцинового оперона. Оперон leuABCD был клонирован в вектор pBR322 методом «shotgun». Для этой цели хромосома штамма MG1655 был обработана рестриктазой MfeI, затуплена фрагментом Кленова и лигирована с плазмидой pBR322, предварительно обработанной EcoRV. Штамм Е.coli leu::Тn10 (ВКПМ В-6038) был использован в качестве реципиента для клонирования. В результате была получена плазмида pBR-leuABCD, содержащая фрагмент хромосомы в 6.9 т.п.о. с опероном leuABCD (Фиг.6). Пример 5. Продукция и экскреция норлейцина и норвалина у производных штамма NS1118. Штамм Е.coli NS1118 и его производные выращивались в течение 18 часов при 37°С на чашках с L-агаром, в случае плазмидных штаммов, содержащих ампициллин (100 мкг/мл). Затем клетки, собранные примерно с 0.5 см2 поверхности чашки, переносили в ферментационную среду (2 мл. Пример 2) и выращивались в течение 72 часов при 32°С. Накопленные в среде количества норвалина и норлейцина измерялись с помощью ВЖКХ. Для подробного описания процесса измерений смотри Пример 6. Полученные результаты представлены в Таблице 2.
Как видно из Таблицы 2, восстановление активности AHAS путем введения плазмиды pMW118-ilvIH, содержащей гены UvIH (кодирующие AHAS III), блокирует продукцию и выделение норвалина и норлейцина штаммом NS1118, дефицитным по AHAS-активности. Амплификация природного лейцинового оперона, входящего в состав плазмиды pBR-leuABCD, повышает уровень накопления норлейцина в ферментационной среде. Пример 6. Условия хроматографического анализа норвалина и норлейцина в культуральной жидкости по окончанию ферментации. Для анализа использовался метод Waters AccQ-Tag®. Метод AccQ-Tag заключается в предколоночной модификации аминокислот для последующего их определения. Использовалась градиентная система ВЖКХ Alliance 2690 gradient system (Waters Corp.), оснащенная флуоресцентным детектором 1100 серии (Agilent Technologies) и соединенная с ПК, на котором установлено программное обеспечение для обработки хроматограмм “ChemStation A.08.04” (Agilent Technologies). Использовались следующие установки детектора: длина волны возбуждения атомов – 250 нм, длина волны излучения – 395нм. Для всех измерений использовалась петля инжектора объемом 5 мкл. Колонка для анализа аминокислот Waters AccQ-Tag Amino Acid Analysis Column оснащена предколонкой, разделяющей производные аминокислот, полученные в результате реакции дериватизации этих аминокислот с Accq-fluor. Колонку термостатировали в колоночной печи при 37°С. Скорость подвижной фазы составляла 1 мл/мин. Использовался следующий состав подвижной фазы: (В) водный буфер “Waters AccQ-Tag Eluent А”, (С) ацетонитрил HPLC-очистки и (D) вода Mili-Q. Градиентная программа для хроматографического анализа аминокислот приведена в Таблице 3. Профили хроматографического анализа аминокислот представлены на Фиг.7.
Формула изобретения
1. Бактерия, принадлежащая к роду Escherichia, – продуцент норлейцина и норвалина, при этом указанная бактерия модифицирована таким образом, что все синтазы ацетогидроксикислот (AHAS) инактивированы. 2. Бактерия по п.1, отличающаяся тем, что синтазы ацетогидроксикислот выбраны из группы, состоящей из AHAS I, кодируемой генами ilvBN, AHAS II, кодируемой генами ilvGM, и AHAS III, кодируемой генами ilvIH. 3. Бактерия по п.1, отличающаяся тем, что указанная бактерия дополнительно модифицирована таким образом, что экспрессия оперона leuABCD в указанной бактерии усилена. 4. Способ получения норлейцина и норвалина, включающий стадии выращивания бактерии по любому из пп.1-3 в среде, которая не содержит L-лейцин, но содержит L-валин в концентрации, лимитирующей рост бактерии, и выделения полученной непротеиногенной аминокислоты из культуральной жидкости.
РИСУНКИ
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 -изопропилмалатсинтазу, ген leuB кодирует
-изопропилмалатсинтазу, ген leuB кодирует  -изопропилмалатдегидрогеназу, гены leuC и leuD кодируют изопропилмалатизомеразы.
-изопропилмалатдегидрогеназу, гены leuC и leuD кодируют изопропилмалатизомеразы. Red системы гомологичной рекомбинации (гены
Red системы гомологичной рекомбинации (гены  0.6, после чего получали электрокомпетентные клетки путем 100-кратной концентрации и трехкратной отмывки ледяной деионизированной Н2О. Электропорацию осуществляли с использованием 70 мкл клеток и примерно «100 нг ПЦР продукта. После электропорации клетки инкубировались в 1 мл среды SOC (Sambrook et al, “Molecular Cloning A Laboratory Manual, Second Edition”, Cold Spring Harbor Laboratory Press (1989)) при 37°С в течение 2.5 часов, после чего высевались на чашки с L-агаром и выращивались при 37°С для отбора CmR рекомбинантов. Далее, для удаления плазмиды pKD46, производили 2 пассажа на L-агаре с Cm при 42°С и полученные колонии тестировались на чувствительность к ампициллину.
0.6, после чего получали электрокомпетентные клетки путем 100-кратной концентрации и трехкратной отмывки ледяной деионизированной Н2О. Электропорацию осуществляли с использованием 70 мкл клеток и примерно «100 нг ПЦР продукта. После электропорации клетки инкубировались в 1 мл среды SOC (Sambrook et al, “Molecular Cloning A Laboratory Manual, Second Edition”, Cold Spring Harbor Laboratory Press (1989)) при 37°С в течение 2.5 часов, после чего высевались на чашки с L-агаром и выращивались при 37°С для отбора CmR рекомбинантов. Далее, для удаления плазмиды pKD46, производили 2 пассажа на L-агаре с Cm при 42°С и полученные колонии тестировались на чувствительность к ампициллину. ilvBN::cat, составляла 1080 п.о. (Фиг.4, SEQ ID NO: 6).
ilvBN::cat, составляла 1080 п.о. (Фиг.4, SEQ ID NO: 6). ,
,