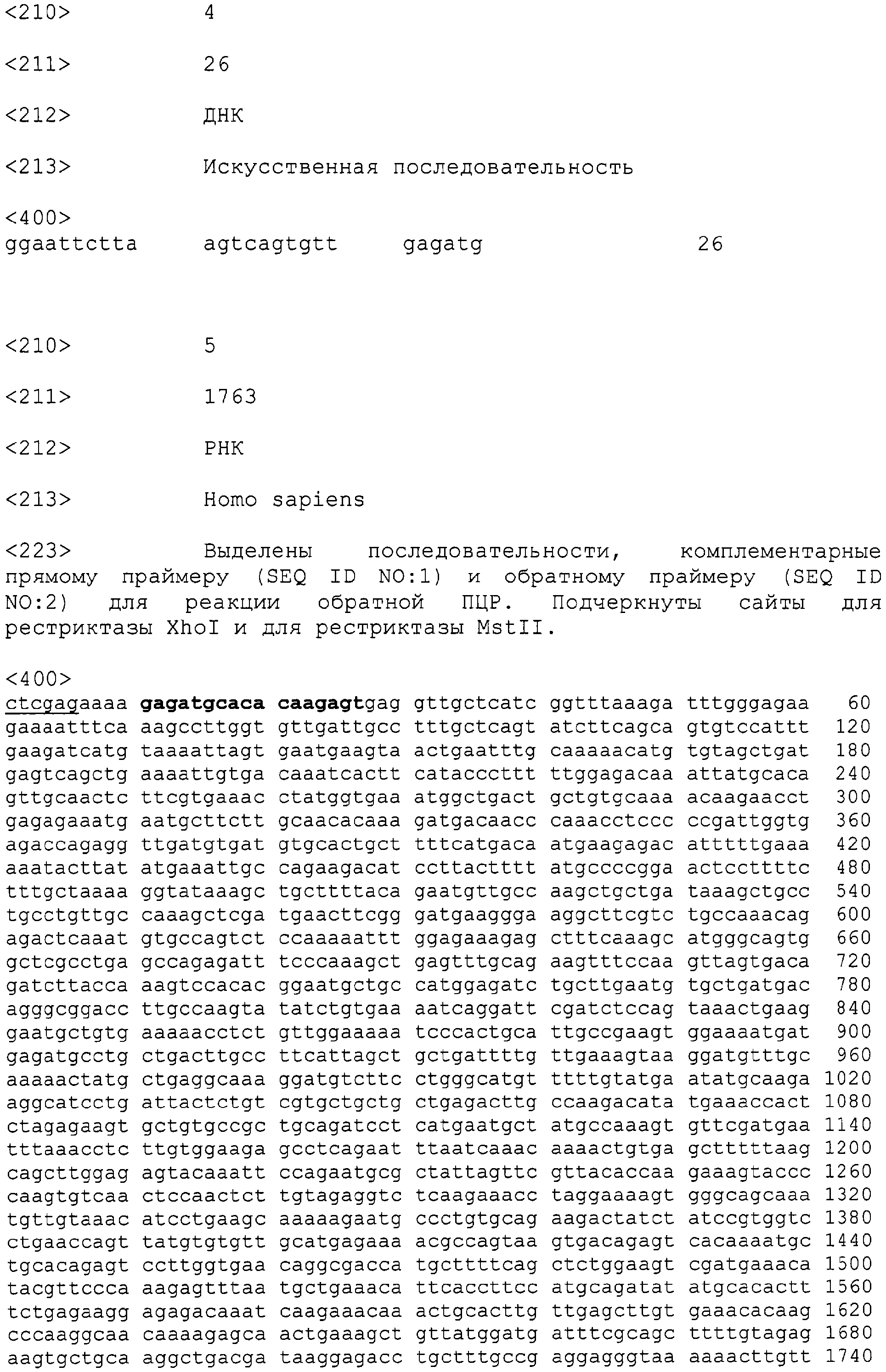
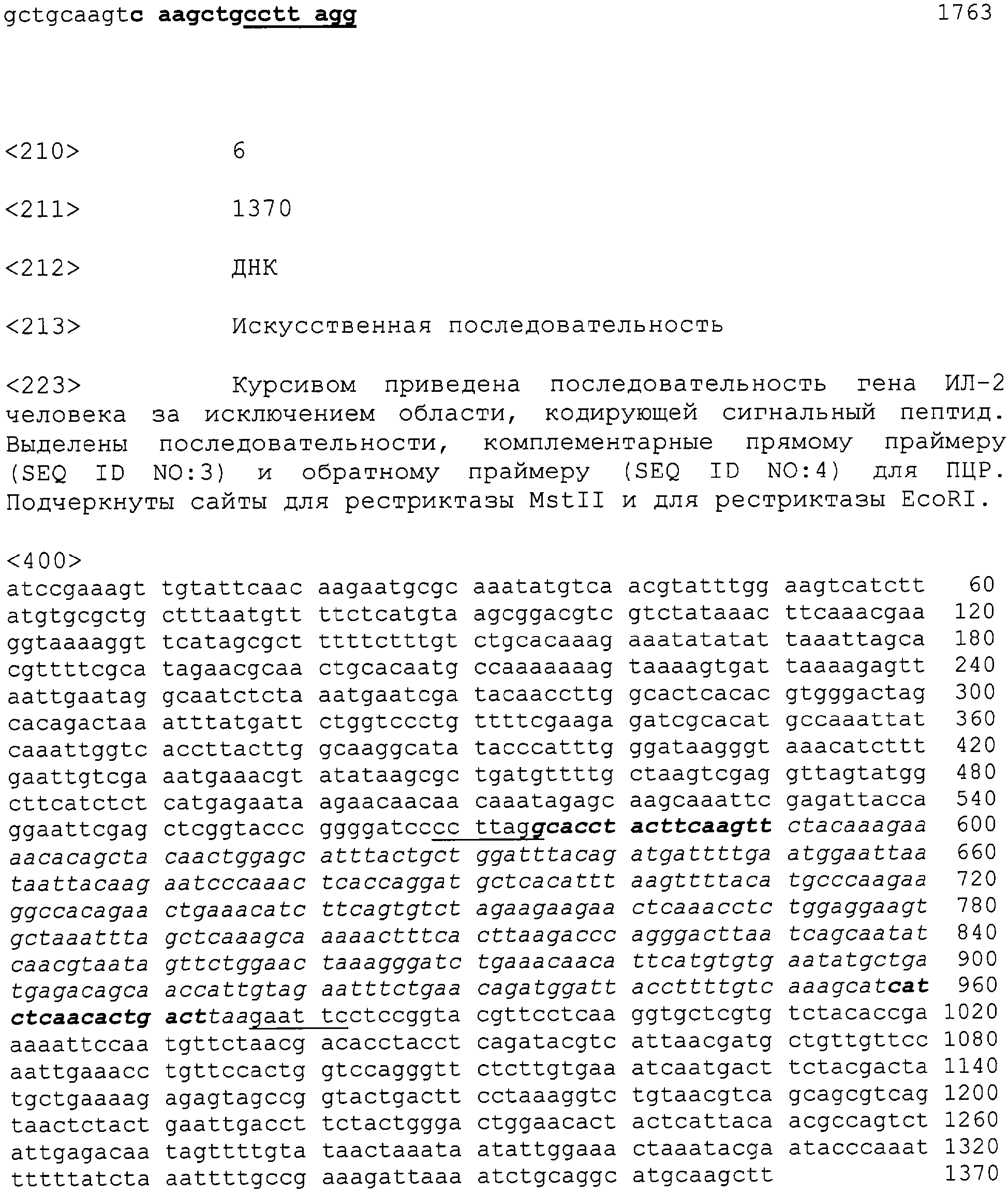
Патент на изобретение №2315105
|
||||||||||||||||||||||||||
(54) ШТАММ ДРОЖЖЕЙ PICHIA PASTORIS PS107(pPIC9HAbIL-2), ЯВЛЯЮЩИЙСЯ ПРОДУЦЕНТОМ ГИБРИДНОГО БЕЛКА, СОСТОЯЩЕГО ИЗ АЛЬБУМИНА ПЛАЗМЫ КРОВИ ЧЕЛОВЕКА И ИНТЕРЛЕЙКИНА-2 ЧЕЛОВЕКА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pPIC9HAbIL-2 И СПОСОБ ЕЕ КОНСТРУИРОВАНИЯ
(57) Реферат:
Изобретение относится к области биотехнологии и может быть использовано для получения препаратов гибридных белков. Штамм дрожжей Pichia pastoris PS99 (his4 pep4::PHO85) трансформируют сконструированной плазмидой pPIC9HAbIL-2 и получают штамм Pichia pastoris PS107 (pPIC9HAbIL-2) – продуцент гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека. Изобретение обеспечивает получение высокостабильного гибридного белка альбумина плазмы крови человека и ИЛ-2 человека по упрощенной технологии. 3 н.п. ф-лы, 2 ил.
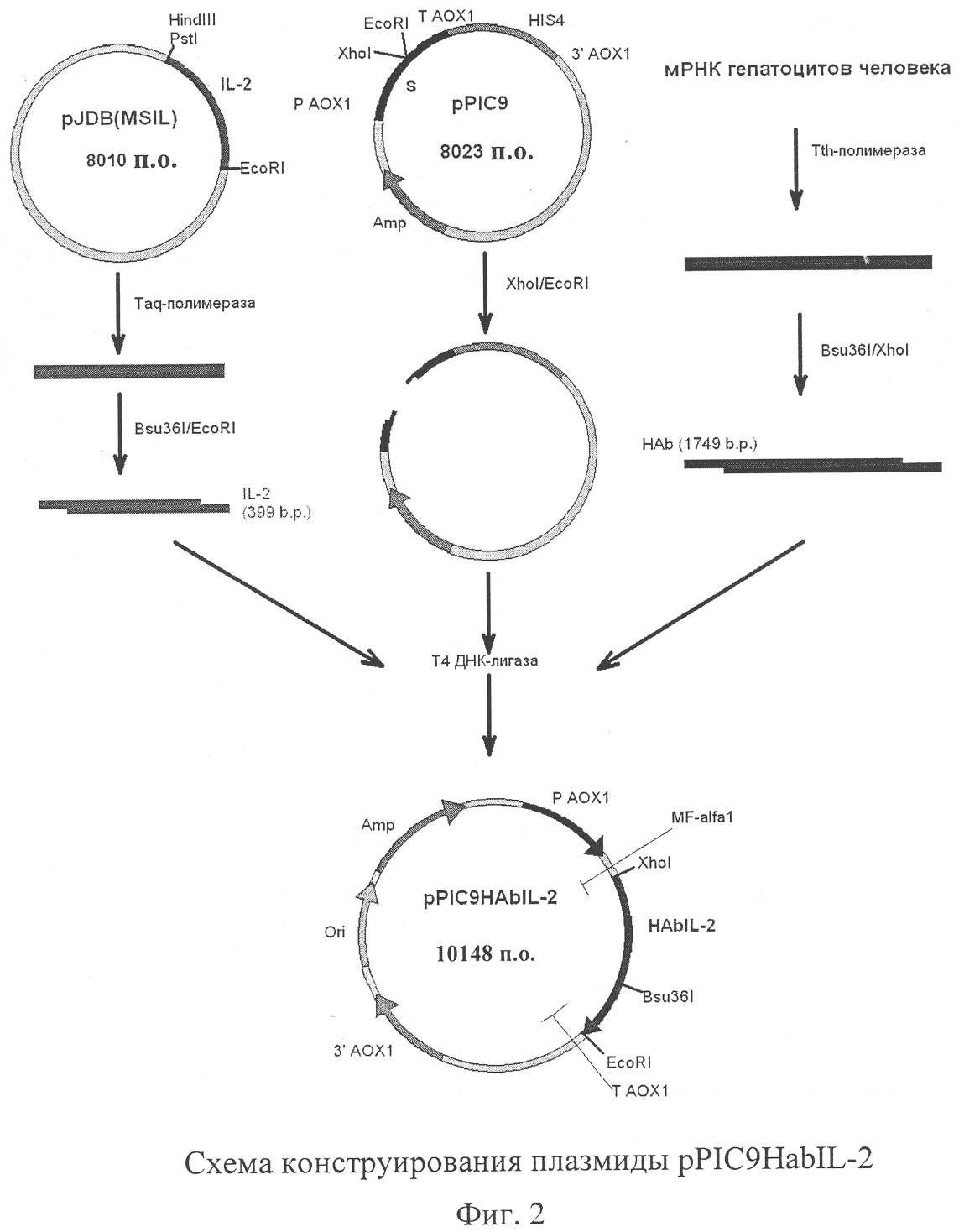
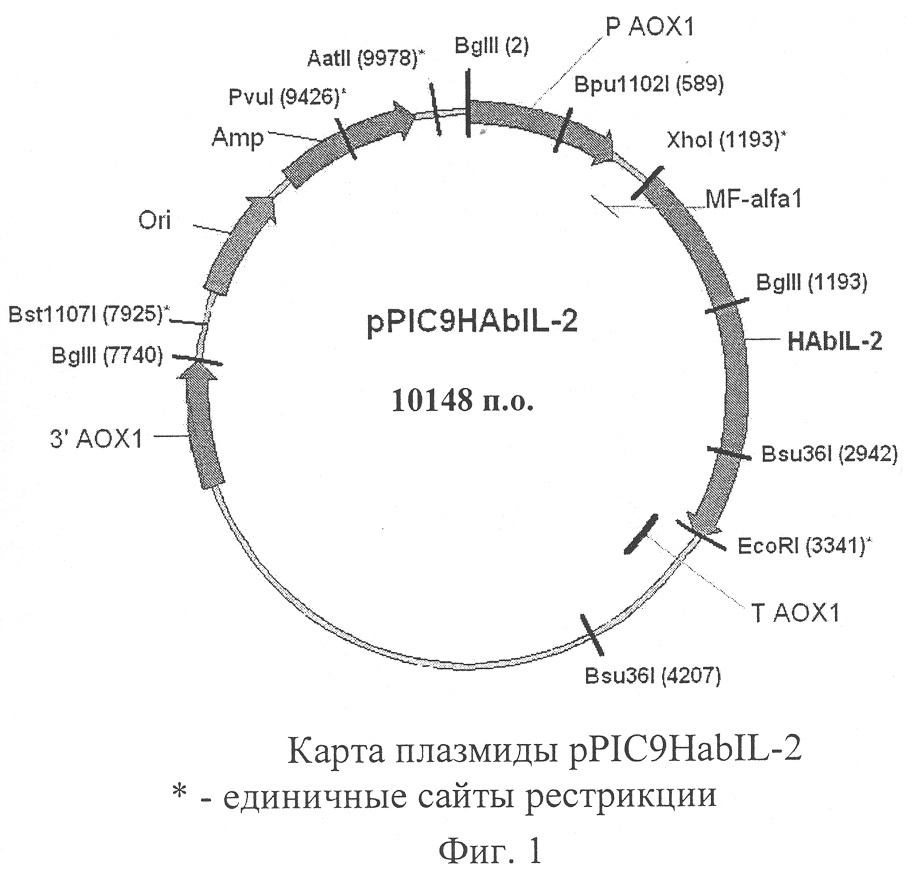
Изобретение относится к биотехнологии, к генной инженерии, микробиологической и медицинской промышленности и представляет собой дрожжевой штамм – продуцент гибридного белка, состоящего из альбумина плазмы крови человека и интерлейкина-2 (ИЛ-2) человека, содержащий сконструированную in vitro плазмиду, которая обеспечивает синтез и секрецию данного гибридного белка. Настоящее изобретение частично основано на обнаружении того факта, что гибридные белки, содержащие альбумин плазмы крови человека, присоединенный к целевому белку, обладают улучшенными фармакокинетическими свойствами, такими как более продолжительное время полужизни (Yeh P. et al. – Proc. Natl. Acad. Sci. – 1992. – V.89. – P.1904-1908; Smith В. et al. – Bioconjugate Chem. – 2001. – V.12. – P.750-756; Halpern W. et al. – Pharm. Res. – 2002. – V.19. – P.1720-1729). ИЛ-2 продуцируется Т-лимфоцитами в ответ на антигенную и митогенную стимуляцию или под влиянием ИЛ-1 (Gillis S. et al.- J. Immunol. – 1978. – V.120. – Р.2027-2032). Основная функция ИЛ-2 состоит в обеспечении клеточной составляющей адаптивного иммунитета. ИЛ-2 является фактором роста и дифференцировки Т-лимфоцитов, NK-клеток, обеспечивает дифференцировку Т-киллеров и способствует проявлению функциональной активности Т-хелперов. Он также воздействует на моноциты, экспрессирующие рецептор ИЛ-2, и усиливает синтез иммуноглобулинов предварительно активированными В-лимфоцитами (Smith К. – Science. – 1988. – V.240. – P.1169-1176). Широкий спектр биологических активностей ИЛ-2 явился мощным стимулом для использования его в качестве лекарственного препарата и для создания рекомбинантных штаммов микроорганизмов – продуцентов этого цитокина. В настоящее время в России зарегистрированы и применяются два рекомбинантных ИЛ-2 человека: «Пролейкин» (Chiron Corp., США) и «Ронколейкин» (Биотех, Россия). Действующим началом «Пролейкина» является ИЛ-2 человека, синтезированный в клетках Е.coli. Рекомбинантный белок не гликозилирован и является мутеином, у него отсутствует N-концевой аланин и цистеин в 125 положении замещен серином. Значительные побочные эффекты, сопровождающие клиническое применение «Пролейкина», а также высокая стоимость препарата ограничивают его широкое использование. «Ронколейкин» представляет собой ИЛ-2 человека, синтезированный в клетках дрожжей S.cerevisiae. Известными применяемыми штаммами являются ВКПМ Y-791 (Мясников А.Н. и др., SU 17703359) и ВКПМ Y-3079 (Смирнов М.Н. и др., SU 2230781). Рекомбинантный белок, как и в случае «Пролейкина», накапливается внутри клеток в восстановленном состоянии в виде «телец включения» и не содержит углеводного компонента. Но в отличие от «Пролейкина» в молекуле «Ронколейкина» отсутствуют аминокислотные замены, и он не является мутеином. Существенным недостатком ИЛ-2 является короткий период полужизни, поэтому в процессе лечения необходимы частые инъекции препарата. Это удорожает лечение, может сопровождаться побочными эффектами и отрицательно сказываться на качестве жизни пациентов (Winkelhake J., Gauny S. – Pharmacol. Rev. – 1990. – V.42. – P.1-28; Nadeau R. et al., Drug Metabol. Dispos. – 1995. – V.23. – P.904-909). Для увеличения периода полужизни рекомбинантных белков используют модификацию молекулы белка, которая заключается в ковалентном связывании активного белка с полиэтиленгликолем (ПЭГ) или альбумином. Пегилирование ИЛ-2 сопровождается увеличением периода полужизни препарата, замедлением выведения, понижением токсичности и иммуногенности (Katre N. et al. – Proc. Natl. Acad. Sci. – 1987. – V.84. – P.1487-1491; Knauf M. et al. – J. Biol. Chem. – 1988. – V.263. – P.15064-15070; Chapes S. et al. – J. Appl. Physiol. – 1999. – V.86. – P.:2065-2076). Однако ПЭГ-конъюгаты не лишены недостатков, среди которых следует отметить зависимость активности белка от структуры ПЭГ (Veronese F. – Biomaterials. – 2001. – V.22. – Р.405-417). Особый интерес представляет модификация рекомбинантных белков за счет их слияния с молекулой альбумина человека. Альбумин является основным белком плазмы крови человека. Период полужизни альбумина около 19 дней. Он обладает рядом ценных свойств, главным из которых является его способность связывать и транспортировать эндо- и экзогенные лиганды (в том числе и белки). Альбумин играет роль естественного стабилизатора и переносчика белков плазмы крови. Он широко используется в качестве стабилизирующего агента при формулировании лекарственных препаратов, созданных на основе белков (Peters Т. – All about albumin. – 1996. – Academic Press). Все вышеизложенное свидетельствует о потребности в создании штаммов микроорганизмов – продуцентов гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека. Использование для гетерологичной экспрессии метилотрофных дрожжей Pichia pastoris представляет особый интерес. Гликопротеины, секретируемые дрожжами Pichia pastoris, не содержат маннозных остатков, связанных Таким образом, в задачу настоящего изобретения входило создание штамма, желательно дрожжей Pichia pastoris, синтезирующего гибридный белок, состоящий из альбумина плазмы крови человека и ИЛ-2 человека, и секретирующего данный гибридный белок в культуральную жидкость, создание плазмиды, которая позволила бы синтезировать упомянутый гибридный белок в дрожжах Pichia pastoris, а также предложение способа конструирования такой плазмиды. Указанная задача решена предложением плазмиды pPIC9HAbIL-2, которая обеспечивает синтез и секрецию гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, трансформированными ею клетками дрожжей, состоящей из следующих элементов: – EcoRI-XhoI – фрагмента плазмидной ДНК бифункционального бактериально-дрожжевого вектора рРIС9 размером 8,00 т.п.о., включающего бактериальный ген устойчивости к ампициллину, бактериальную область инициации репликации, ген HIS4 дрожжей; фрагмент 5′-некодирующей области дрожжевого гена АОХ1 размером 0,95 т.п.о., содержащий область, обеспечивающую активацию транскрипции этого гена в присутствии метанола в качестве источника углерода в культуральной среде; препрообласть гена MF – XhoI-Bsu36I – фрагмента размером 1749. п.о., содержащего кодирующую часть гена альбумина плазмы крови человека за исключением области, кодирующей сигнальный пептид, и терминирующего кодона; – Bsu36I-EcoRI – фрагмента размером 399 п.о., содержащего кодирующую часть гена ИЛ-2 человека за исключением области, кодирующей сигнальный пептид; содержащая уникальные сайты распознавания следующих рестриктаз: XhoI – 1193 п.о.; EcoRI – 3341 п.о.; Bst1107I – 7925 п.о.; PvuI – 9426 п.о.; AatII – 9978 п.о. Схема плазмиды pPIC9HAbIL-2 с рестрикционной картой изображена на Фиг.1. Общий размер плазмиды 10148 п.о. Согласно изобретению также предложен способ конструирования плазмиды pPIC9HAbIL-2. Схема указанного способа приведена на Фиг.2. В предлагаемом способе ген альбумина плазмы крови человека за исключением области, кодирующей сигнальный пептид, и терминирующего кодона, получают при помощи обратной ПЦР с использованием матрицы мРНК, полученной из гепатоцитов человека, прямого праймера SEQ ID NO:1, содержащего сайт для рестриктазы XhoI, обратного праймера SEQ ID NO:2, содержащего сайт для рестриктазы MstII; ген ИЛ-2 человека за исключением области, кодирующей сигнальный пептид, получают при помощи ПЦР с использованием матрицы плазмиды pJDB(MSIL), прямого праймера SEQ ID NO:3, содержащего сайт для рестриктазы Bsu36I, обратного праймера SEQ ID NO:4, содержащего сайт для рестриктазы EcoRI; полученный ген альбумина плазмы крови человека обрабатывают рестриктазами XhoI и MstII, полученный ген ИЛ-2 человека обрабатывают рестриктазами Bsu36I и EcoRI и лигируют с плазмидой рРIС9, предварительно обработанной рестриктазами XhoI и EcoRI, затем лигазной смесью трансформируют клетки Escherichia coli и отбирают клоны, содержащие рекомбинантную плазмиду pPIC9HAbIL-2. Плазмиду pJDB(MSIL) (Мясников и др. А.с. SU 17703359 A1, МКИ С12N 15/26, 1/19. – 1989) используют в качестве матрицы для амплификации гена ИЛ-2 человека при помощи ПЦР. Последовательность фрагмента плазмиды pJDB(MSIL), использованная в качестве матрицы в ПЦР для получения гена ИЛ-2 человека за исключением области, кодирующей сигнальный пептид, представлена как SEQ ID NO:6. Курсивом приведена последовательность гена ИЛ-2 человека за исключением области, кодирующей сигнальный пептид. Выделены последовательности, комплементарные прямому праймеру (SEQ ID NO:3) и обратному праймеру (SEQ ID NO:4) для ПЦР. Подчеркнуты сайты для рестриктазы MstII и для рестриктазы EcoRI. В качестве прямого праймера служит олигонуклеотид 5 Для получения гена альбумина человека применяют метод обратной ПЦР, в качестве матрицы используют мРНК, выделенную из культуры гепатоцитов человека. Последовательность мРНК из гепатоцитов человека, использованная в качестве матрицы в реакции обратной ПЦР для получения гена альбумина плазмы крови человека за исключением области, кодирующей сигнальный пептид, и терминирующего кодона, представлена как SEQ ID NO:5. Выделены последовательности, комплементарные прямому праймеру (SEQ ID NO:1) и обратному праймеру (SEQ ID NO:2) для реакции обратной ПЦР. Подчеркнуты сайты для рестриктазы XhoI и для рестриктазы MstII. Прямым праймером является олигонуклеотид 5 Полученные фрагменты ДНК, содержащие ген ИЛ-2 человека и ген альбумина человека, лигируют с плазмидой pPIC9 («Invitrogen»), предварительно обработанной рестриктазами XhoI и EcoRI. Полученной лигазной смесью трансформируют клетки штамма DH5 Таким образом, плазмида pPIC9HAbIL-2 создана на основе челночного бактериально-дрожжевого интегративного вектора рРIС9 (имеется в продаже («Invitrogen»). В результате трансформации линеаризованной плазмидой и последующей гомологичной рекомбинации происходит встраивание экспрессионной кассеты в хромосому дрожжей Pichia pastoris, что обеспечивает стабильное поддержание клонированного гибридного гена, который состоит из находящихся в одной рамке считывания гена альбумина человека без терминирующего кодона и гена ИЛ-2 человека. Преимуществом такой интегративной системы экспрессии является также отсутствие в геноме дрожжей интегрантов фрагментов бактериальной ДНК и генов устойчивости к антибиотикам, что предотвращает опасность горизонтального переноса этих генов. В состав плазмиды входит ген HIS4 дрожжей, что позволяет селективно отбирать трансформантов при использовании в качестве реципиентов штаммы дрожжей с мутациями в этом гене. Экспрессия гибридного гена, который состоит из находящихся в одной рамке считывания гена альбумина человека без терминирующего кодона и гена ИЛ-2 человека, в составе плазмиды pPIC9HAbIL-2 находится под контролем промотора гена АОХ1, содержащим области, обеспечивающие активацию транскрипции в присутствии метанола в культуральной среде, а также область инициации транскрипции. Промотор гена АОХ1 относится к числу наиболее сильных дрожжевых промоторов. Уровень экспрессии генов, находящихся под контролем АОХ1 промотора, эффективно регулируется источниками углерода. Транскрипция гена АОХ1 полностью блокирована при выращивании дрожжей на среде с глюкозой, на среде с глицерином наблюдается только базальный уровень экспрессии гена. Использование метанола в качестве единственного источника углерода значительно усиливает экспрессию гена АОХ1 и, следовательно, генов, находящихся под контролем АОХ1 промотора. Это позволяет регулировать синтез гибридного гена, который состоит из находящихся в одной рамке считывания гена альбумина человека без терминирующего кодона и гена ИЛ-2 человека в клетках дрожжей. Регулируемая экспрессия клонированного гена позволяет существенно снизить метаболическую нагрузку на клетку дрожжей. Наличие в составе плазмиды препрообласти гена MF В качестве продуцента гибридного белка, который состоит из альбумина человека и ИЛ-2 человека, используют штамм PS107(pPIC9HAbIL-2). Штамм PS107(pPIC9HAbIL-2) получен трансформацией штамма дрожжей PS99 (his4 pep4::PHO85) плазмидой pPIC9HAbIL-2. Штамм PS99 несет мутацию в гене HIS4, что позволяет селективно отбирать трансформантов, несущих плазмиду pPIC9HAbIL-2. Мутация в гене РЕР4 приводит к отсутствию активности протеаз А и В, а также карбоксипептидазы Y в клетках дрожжей, что сопровождается повышением стабильности гетерологичных рекомбинантных белков (Hisch H.H. et al. In: Walton E.F., Yarranton G.T., Eds., Molecular and Cell Biology of Yeast. – 1989. – P. 134-200). Штамм дрожжей Pichia pastoris PS107(pPIC9HAbIL-2) характеризуется следующими признаками. Морфологические признаки. Клетки округлой, слегка овальной формы, размером 5-10 мкм, часть клеток имеет на поверхности почки или соединена с дочерними клетками. Культуральные признаки. Клетки хорошо растут на полной органической среде YEPD – 2% пептона, 1% дрожжевого экстракта, 2% глюкозы или 1% глицерина. Кроме того, клетки хорошо растут на минеральной среде SC: 1,34% Yeast Nitrogen Base («Difco», США), а также на других синтетических средах для дрожжей, содержащих в качестве источника углерода глюкозу (2%) или глицерин (1%). Клетки штамма отличаются слабым ростом на средах, содержащих в качестве источника углерода метанол (0,5%-1%). При росте на твердых средах клетки образуют гладкие, круглые колонии с матовой поверхностью, светло-кремового цвета, край неровный. При росте в жидких средах образуют интенсивную ровную суспензию. Культура имеет характерный запах метилотрофных дрожжей. Физиолого-биохимические признаки. Клетки растут в пределах от 4 до 37°С. Оптимальной температурой выращивания является 30°С. При росте в аэробных условиях клетки незначительно закисляют среду. Оптимум рН для роста составляет 4,5-6,5. В качестве источника углерода клетки могут использовать многие простые соединения, такие как: глюкоза, глицерин, метанол. В качестве источника азота клетки могут использовать минеральные соли в аммонийной форме, аминокислоты, мочевину. Клетки способны к аэробному и анаэробному росту. Существенными признаками штамма является отсутствие потребности в гистидине. Способ получения EcoRI-XhoI – фрагмента плазмидной ДНК рРIС9 проиллюстрирован следующим примером. ПРИМЕР 1. Клетки бактерий Escherichia coli, содержащие плазмиду рРIС9, выращивают при 37°с в течение ночи в 1 л питательной среды LB (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия), содержащей ампициллин в концентрации 50 мг/л. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут при 4°С, суспендируют в 20 мл 25 мМ трис-хлоридного буфера (рН 8,0), содержащего 10 мМ ЭДТА и 50 мМ глюкозы, добавляют 30 мг лизоцима и инкубируют 10 минут при комнатной температуре. Далее добавляют 40 мл 0,2 М гидроокиси натрия, содержащей 1% додецилсульфата натрия, осторожно перемешивают и инкубируют в течение 10 минут при 4°С. Раствор нейтрализуют добавлением 30 мл 3 М ацетата натрия (рН 5,0) и выдерживают в течение 10 минут при 4°С. После этого центрифугируют при 14000 об/мин в течение 40 минут при 4°С. К супернатанту добавляют 0,6 объема изо-пропилового спирта, выдерживают 20 минут при комнатной температуре и центрифугируют при 14000 об/мин в течение 20 минут при 20°С. Полученный осадок промывают 70% этиловым спиртом, высушивают в вакууме и растворяют в 4 мл дистиллированной воды. Далее добавляют 4,2 г хлористого цезия и 0,36 мл раствора бромистого этидия (10 мг/мл). Полученный раствор выдерживают в течение 1 часа при 4°С, затем центрифугируют при 15000 об/мин в течение 15 минут. Супернатант центрифугируют при 70000 об/мин в течение 16 часов в центрифуге TL100 («Beckman»). После центрифугирования отбирают полосу плазмидной ДНК (нижнюю из двух флюоресцирующих в ультрафиолетовом свете полос), дважды экстрагируют бромистый этидий равным объемом изо-амилового спирта, разбавляют в два раза дистиллированной водой и осаждают плазмидную ДНК двумя объемами этилового спирта и 1/15 объема 3 М ацетата натрия (рН 5,0). Осадок собирают центрифугированием при 10000 об/мин в течение 10 минут, промывают 70% этиловым спиртом и растворяют в 0,5-1 мл буфера ТЕ (10 мМ трис-хлоридный буфер, рН 8,0, содержащий 1 мМ ЭДТА). Концентрацию плазмидной ДНК определяют по поглощению раствора при длине волны 260 нм. Чистоту препарата контролируют при помощи электрофореза в 0,7% агарозном геле в буфере ТВЕ (0,1 М трис-боратный буфер, рН 8,3, содержащий 1 мМ ЭДТА). Гидролиз полученной плазмиды рРIС9 рестриктазами XhoI и EcoRI проводят в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 50 мМ хлористого натрия, 10 мМ хлористого магния и 1 мМ дитиотреитол. К 5 мкг плазмидной ДНК в объеме 20 мкл добавляют по 5 ед. каждой рестриктазы, после чего пробу инкубируют в течение 25 часов при 37°С. Реакционную смесь вносят в лунки в 0,7% агарозного геля в буфере ТВЕ и проводят разделение полученных фрагментов ДНК. По окончании разделения вырезают полоску геля, содержащую фрагмент ДНК размером 8 т.п.о., соответствующий линеаризованному XhoI-EcoRI фрагменту плазмиды рРIС9. Выделение ДНК из агарозного геля проводят по методике, разработанной фирмой QIAGEN. Полоску геля с фрагментом ДНК помещают в пробирку и добавляют раствор QX1 (300 мкл на 100 мг геля), добавляют реактив QIAEX (10 мкл на 5 мкг ДНК) и инкубируют при 50°С в течение 10 минут, периодически перемешивая. Далее центрифугируют 30 секунд при 15000 об/мин, супернатант отбрасывают, осадок дважды экстрагируют растворами QX2 и QX3, удаляют супернатант центрифугированием при 15000 об/мин в течение 30 секунд. Осадок высушивают на воздухе, растворяют в 20 мкл буфера ТЕ, центрифугируют 30 секунд при 15000 об/мин, супернатант переносят в новую пробирку. Способ получения гена ИЛ-2 человека проиллюстрирован следующим примером. ПРИМЕР 2. Ген ИЛ-2 человека амплифицируют при помощи ПЦР. В качестве матрицы используют плазмиду pJDB(MSIL). Выделение плазмиды pJDB(MSIL) проводят в условиях, аналогичных для плазмиды рРIС9. К 0,1 мкг плазмиды pJDB(MSIL), растворенной в 5 мкл буфера ТЕ, добавляют 1 мкл 0,5 М NaOH и нагревают при 85° в течение 3 минут. Затем пробу быстро переносят в лед, добавляют 1 мкл 0,5 М HCl и далее используют в ПЦР. В качестве прямого праймера служит олигонуклеотид 5′-caagctgccttaggcttagcacctacttcaagt, содержащий сайт для рестриктазы Bsu36I. В качестве обратного праймера используют олигонуклеотид 5′-ggaattcttaagtcagtgttgagatg, который содержит сайт для рестриктазы EcoRI. Проба для ПЦР содержит 5 мкл матрицы, 30 рМ каждого праймера, 10 мкл 10-кратного раствора дезоксинуклеотидтрифосфатов (дНТФ), содержащего 1,25 мМ каждого дНТФ (дАТФ, дТТФ, дГТФ, дЦТФ), 10 мкл 10-кратного буфера для ПЦР (100 mM KCl, 100 mM (NH4)2SO4, 200 mM Трис-HCl, pH 8,8, 20 mM MgSO4, 1% тритон Х-100). В пробу добавляется дистиллированная Н2O до конечного объема 100 мкл. Далее пробу прогревают 5 мин при 95°С, охлаждают, добавляют 2,5 ед. Вент-ДНК-полимеразы («BioLabs») и проводят 50 циклов ПЦР в следующих условиях: 1 мин при 95°С (плавление цепей ДНК), 1 мин при 46°С (отжиг праймеров), 1 мин при 72°С (полимеразная реакция). После окончания ПЦР пробу инкубируют при 72°С 5 мин. Реакционную смесь вносят в лунки в 0,7% агарозного геля в буфере ТВЕ и проводят разделение полученных фрагментов ДНК. По окончании разделения вырезают полоску геля, содержащую фрагмент ДНК размером 399 п.о., соответствующий гену ИЛ-2 человека. Выделение ДНК из агарозного геля проводят по описанной выше методике фирмы QIAGEN. Гидролиз фрагмента ДНК, содержащего амплифицированный ген ИЛ-2 человека, рестриктазами Bsu36I и EcoRI проводят в условиях, описанных для расщепления плазмиды рРIС9. Способ получения гена альбумина человека проиллюстрирован следующим примером. ПРИМЕР 3. Ген альбумина человека получают методом обратной полимеразной цепной реакции, используя в качестве матрицы мРНК, полученную из клеток печени человека. Клетки печени человека (банк клеточных культур Института цитологии РАН, Санкт-Петербург) культивируют в среде Игла, содержащей 2-глутамин и 10% эмбриональную сыворотку крупного рогатого скота, до плотности 0,5-1,0×105 клеток/мл. Снимают клетки, используя 0,02% версен, содержащий химотрипсина (0,1 мг/мл), дважды промывают клетки средой Игла, содержащей 2-глутамин, 10% эмбриональную сыворотку крупного рогатого скота и осаждают клетки центрифугированием (800 об/мин, 10 мин). Далее клетки дважды промывают изотоническим раствором, свободным от РНКаз и ресуспендируют в 100 мкл раствора, содержащего 4 М гуанидин тиоционат, 25 мМ цитрат натрия, pH 7.0, 0,5% саркозил натрия, 0,1 М 2-меркаптоэтанол и 0,3 М ацетата натрия (рН 4). Полученную смесь встряхивают на «Vortex» в течение 10 мин, центрифугируют 5 мин при 12000 об/мин, отбирают водную фазу, добавляют к ней равный объем смеси фенол:хлороформ (1:1), уравновешенной буфером, содержащим 25 мМ цитрат натрия, рН 7.0, 0,5% саркозил натрия, 0,1 М 2-меркаптоэтанол и 0,3 М ацетата натрия (рН 4), встряхивают на «Vortex» в течение 5 мин, центрифугируют 5 мин при 12000 об/мин. Отбирают водную фазу и повторяют экстракцию смесью фенол:хлороформ, центрифугируют 5 мин при 12000 об/мин. К водной фазе добавляют равный объем изо-пропилового спирта. Для осаждения помещают смесь на -20°С на ночь. После центрифугирования (12000 об/мин, 5 мин) осадок промывают холодным 70% этанолом, подсушивают и растворяют в 20 мкл деионизованной воды, свободной от РНКаз. Полученную РНК используют в качестве матрицы для синтеза кДНК гена альбумина человека при помощи обратной ПЦР с использованием Tth-полимеразы. Прямым праймером служит олигонуклеотид 5′-ccgctcgagaaaagagatgcacacaagagt, содержащий сайт для рестриктазы XhoI, обратным праймером служит олигонуклеотид 5′-tgaagtaggtgctaagcctaaggcagcttg, содержащий сайт для рестриктазы MstII. Проба для проведения ПЦР содержит 1 мкл матрицы (примерно 5 мкг), 30 рМ каждого праймера, 10 мкл 10-кратного раствора дезоксинуклеотидтрифосфатов (дНТФ), содержащего 1,25 мМ каждого дНТФ (дАТФ, дТТФ, дГТФ, дЦТФ), 10 мкл 10-кратного буфера (500 mM NaCl, 500 тМ Трис-HCl, рН 9,0, 100 mM MgCl2), 5 ед. Tth-полимеразы. В пробу добавляется дистиллированная Н2O до конечного объема 50 мкл. Стадию обратной ПЦР проводят при 70°С в течение 3 мин, затем 50 мин при 42°С и 2 мин при 94°С. Полученную кДНК гена альбумина человека используют как матрицу для амплификации этого гена при помощи ПЦР, которую проводят в следующем режиме: 30 с при 94°С, 30 с при 58°С и 30 с при 72°С. Повторяют этот цикл 45 раз, после чего инкубируют пробу при 72°С 5 мин. Синтезированный ген альбумина человека размером 1749 п.о. выделяют из агарозного геля по описанной выше методике фирмы QIAGEN. Гидролиз фрагмента ДНК, содержащего амплифицированный ген альбумина человека рестриктазами XhoI и MstII проводят в условиях, описанных для расщепления плазмиды рРIС9. Способ получения плазмиды pPIC9HAbIL-2 проиллюстрирован следующим примером. ПРИМЕР 4. Для получения плазмиды pPIC9HAbIL-2 проводят дотирование XhoI-EcoRI фрагмента плазмиды рРIС9 (получение которого описано в примере 1), Bsu36I-EcoRI фрагмента гена ИЛ-2 человека (получение которого описано в примере 2) и XhoI-MstII фрагмента гена альбумина человека (получение которого приведено в примере 3). Для этого смешивают эквимолярные количества ДНК плазмиды и амплифицированных генов в 10 мкл 40 мМ трис-хлоридного буфера (рН 7,8), содержащего 10 мМ дитиотреитола, 10 мМ хлористого магния, 0,5 мМ АТФ, добавляют 5 ед. ДНК-лигазы фага Т4 и инкубируют при 14°С в течение ночи. Полученной лигазной смесью трансформируют клетки штамма DH5 Из выросших отдельных клонов трансформантов выделяют плазмидную ДНК при помощи методики, использованной для получения плазмиды рРIС9, за исключением того, что клетки Escherichia coli выращивают в 10 мл LB, и, соответственно, объемы всех растворов уменьшают в 100 раз. Кроме того, вместо стадии центрифугирования в градиенте плотности хлористого цезия проводят обработку ДНК панкреатической РНКазой. Для этого нуклеиновые кислоты, осажденные изо-пропиловым спиртом, растворяют в 100 мкл буфера ТЕ, добавляют 10 мкл раствора РНКазы (1 мг/мл) и инкубируют 30 минут при 37°С. Далее проводят гидролиз полученной плазмидной ДНК рестриктазами EcoRI и XhoI или BglII. При рестрикции искомой плазмиды pPIC9HAbIL-2 и последующем электрофорезе в 0,7% агарозном геле в первом случае обнаруживаются фрагменты 2,148 п.о. и 8,0 т.п.о., во втором – фрагменты 1911 п.о., 2410 п.о. и 5827 п.о. Из выявленного таким образом клона препаративно выделяют плазмиду pPIC9HAbIL-2 так же, как описано для плазмиды рР1С9, и гидролизуют рестриктазами Bst1107I и AatII в условиях, описанных для расщепления плазмиды рРIС9. Bst1107I-AatII фрагмент плазмиды pPIC9HAbIL-2, размером 8,1 т.п.о., выделяют по описанной выше методике фирмы QIAGEN и используют его для трансформации клеток дрожжей, как описано в примере 5. ПРИМЕР 5. Для получения штамма дрожжей Pichia pastoris – продуцента гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, клетки дрожжей штамма PS99 трансформируют плазмидой pPIC9HAbIL-2. Клетки дрожжей выращивают в 100 мл среды YEPD при 30°С до достижения культурой оптической плотности, соответствующей 2-4 ед. поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 301°С в течение 30 минут. К 50 мкл полученной суспензии клеток добавляют 0,1-1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 минут при 100°) и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля 4000. Далее пробу инкубируют 30 минут при 30°С и 20 минут при 42°С, помещают на 15 секунд в ледяную баню и центрифугируют 10 секунд при 10000 об/мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую среду SC. Клоны трансформантов вырастают через 2-3 суток. Выросшие клоны пересевают на чашки со средой SC, содержащей 2% глюкозу, отдельными колониями, затем перепечатывают на среду ММ (1,34% Yeast Nitrogen Base («Difco», США), 0,5% метанола, 2% агара («Difco», США) для отбора трансформантов, отличающихся слабым ростом на среде с метанолом, что свидетельствует об интеграции гибридного гена, состоящего из гена ИЛ-2 человека и гена альбумина человека, в локус AOXI (фенотип Mets). Для анализа продукции гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, клетками трансформантов фенотипа Mets их выращивают при 30°С в 100 мл жидкой среды BMGY (2% пептона, 1% дрожжевого экстракта, 1% глицерина, 10 мл 1 М калий-фосфатного буфера, рН 6,0, 1,34% Yeast Nitrogen Base («Difco», США) до стационарной фазы роста в течение 2 суток. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут, супернатант сливают и переносят всю биомассу в 20 мл жидкой среды BMMY (2% пептона, 1% дрожжевого экстракта, 0,5% метанола, 10 мл 1 М калий-фосфатного буфера, рН 6,0, 1,34% Yeast Nitrogen Base («Difco», США) для индукции экспрессии гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека. Индукцию проводят при 30°С в течение 4 суток. По окончании индукции культуральную среду отделяют от клеточной биомассы центрифугированием при 5000 об/мин в течение 10 минут. В культуральной среде определяют содержание гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия и последующей гибридизации с антителами к ИЛ-2 человека. Разделение белков проводят в 15% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ трис, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). Параллельно проводят разделение белков контрольного штамма, выращенного в идентичных условиях. В качестве стандартов молекулярной массы используют Согласно полученным данным клетки дрожжей штамма PS107 (pPIC9HAbIL-2) синтезируют и секретируют около 5 мг гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, на литр культуры дрожжей. Гибридный белок обладает биологической активностью, характерной для ИЛ-2 человека. Суммируя вышесказанное можно заключить, что полученный штамм дрожжей Pichia pastoris PS107 (pPIC9HAbIL-2) синтезирует и секретирует гибридный белок, состоящий из альбумина плазмы крови человека и ИЛ-2 человека, в количестве, достаточном для его очистки в лабораторном масштабе. В результате такой очистки могут быть получены препараты гибридного белка, пригодные для исследования его биологических свойств и терапевтической ценности. Преимуществом данного продуцента является большая стабильность получаемого рекомбинантного белка, содержащего рИЛ-2 человека, по сравнению с продуктом штаммов-прототипов (ВКПМ Y-791, ВКПМ Y-3079), что связано с гликозилированием его молекулы и сшивкой с альбумином плазмы крови человека, а также упрощенная процедура очистки, обусловленная секрецией рекомбинантного белка.
Формула изобретения
1. Рекомбинантная плазмида pPIC9HAbIL-2, обеспечивающая биосинтез и секрецию гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, трансформированными ею клетками дрожжей, имеющая размер 10148 п.о. и состоящая из следующих элементов: EcoRl-Xho1 – фрагмента плазмидной ДНК бифункционального бактериально-дрожжевого вектора рР1С9 размером 8,00 т.п.о., ограниченного сайтами рестрикции EcoRl-Xho1 и включающего бактериальный ген устойчивости к ампициллину; бактериальную область инициации репликации; ген HIS4 дрожжей; фрагмент 5 Bsu36I-EcoR1 – фрагмента размером 399 п.о., содержащего кодирующую часть гена ИЛ-2 человека за исключением области, кодирующей сигнальный пептид. 2. Штамм дрожжей Pichia pastoris PS107(pPIC9HAbIL-2) – продуцент гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, представляющий собой штамм Pichia pastoris PS99 (his4 pep4::PHO85), трансформированный плазмидой pPIC9HAbIL-2 по п.1. 3. Способ конструирования рекомбинантной плазмиды pPIC9HAbIL-2 по п.1, при котором ген альбумина плазмы крови человека, за исключением области, кодирующей сигнальный пептид, и терминирующего кодона, получают при помощи обратной ПЦР с использованием матрицы мРНК, полученной из гепатоцитов человека, прямого праймера SEQ ID NO:1, содержащего сайт для рестриктазы Xho1, обратного праймера SEQ ID NO:2, содержащего сайт для рестриктазы MstII; ген ИЛ-2 человека, за исключением области, кодирующей сигнальный пептид, получают при помощи ПЦР с использованием матрицы плазмиды pJDB(MSIL), прямого праймера SEQ ID NO:3, содержащего сайт для рестриктазы Bsu36I, обратного праймера SEQ ID NO:4, содержащего сайт для рестриктазы EcoR1; полученный ген альбумина плазмы крови человека обрабатывают рестриктазами Xhol и MstII; полученный ген ИЛ-2 человека обрабатывают рестриктазами Bsu36I и EcoR1; лигируют гены альбумина плазмы крови человека и ИЛ-2 человека, обработанные как указано выше, с плазмидой рР1С9, предварительно обработанной рестриктазами Xho1 и EcoR1, и полученной лигазной смесью трансформируют клетки Escherichia coli и отбирают клоны, содержащие рекомбинантную плазмиду pPIC9HAbIL-2.
РИСУНКИ
PC4A – Регистрация договора об уступке патента СССР или патента Российской Федерации на изобретение
Прежний патентообладатель:
(73) Патентообладатель:
Договор № РД0032388 зарегистрирован 07.02.2008
Извещение опубликовано: 20.03.2008 БИ: 08/2008
|
||||||||||||||||||||||||||
 1,3-связями и являющихся сильными антигенными детерминантами, что позволяет получать наряду с внутриклеточными аутентичные секреторные формы рекомбинантных белков (Montesino R. et al. – Protein Expr. Purif. – 1998. – V.14. – P.197-207). Получение секреторного продуцента гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, было бы особенно актуально. Природный ИЛ-2 является секреторным белком и только в процессе секреции происходит правильное замыкание внутримолекулярной дисульфидной связи и гликозилирование белка. От корректного замыкания дисульфидной связи зависит биологическая активность ИЛ-2, а присоединение углеводных остатков повышает стабильность молекулы и увеличивает время ее циркуляции в кровотоке (Robb R. et al.- Proc. Natl. Acad. Sci. USA. – 1984. – V.81 – P.6486-6490). Накопление рекомбинантного белка в культуральной жидкости значительно упрощает процедуру его очистки.
1,3-связями и являющихся сильными антигенными детерминантами, что позволяет получать наряду с внутриклеточными аутентичные секреторные формы рекомбинантных белков (Montesino R. et al. – Protein Expr. Purif. – 1998. – V.14. – P.197-207). Получение секреторного продуцента гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека, было бы особенно актуально. Природный ИЛ-2 является секреторным белком и только в процессе секреции происходит правильное замыкание внутримолекулярной дисульфидной связи и гликозилирование белка. От корректного замыкания дисульфидной связи зависит биологическая активность ИЛ-2, а присоединение углеводных остатков повышает стабильность молекулы и увеличивает время ее циркуляции в кровотоке (Robb R. et al.- Proc. Natl. Acad. Sci. USA. – 1984. – V.81 – P.6486-6490). Накопление рекомбинантного белка в культуральной жидкости значительно упрощает процедуру его очистки. -caagctgccttaggcttagcacctacttcaagt (SEQ ID NO:3), содержащий сайт для рестриктазы Bsu36I. В качестве обратного праймера используют олигонуклеотид 5
-caagctgccttaggcttagcacctacttcaagt (SEQ ID NO:3), содержащий сайт для рестриктазы Bsu36I. В качестве обратного праймера используют олигонуклеотид 5 (lacZYA-argF)U169 deoR (
(lacZYA-argF)U169 deoR ( 80dlac
80dlac -галактозидазу (116,0 кДа), бычий сывороточный альбумин (66,2 кДа), овальбумин (45,0 кДа), лактатдегидрогеназу (35,0 кДа), эндонуклеазу рестрикции Bsp98I (25,0 кДа), р-лактоглобулин (18,4кДа), лизоцим (14,4 кДа). По окончании электрофореза белки ренатурируют, выдерживая гели 15 минут в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 4 М мочевину, 20 мМ ЭДТА, и переносят на нитроцеллюлозную мембрану в 25 мМ трис-192 мМ глициновом буфере (рН 8,3), содержащем 20% метилового спирта, при 30-40 В, в течение 1,5 часов. Далее мембрану выдерживают в буфере TBST (10 мМ трис-хлоридный буфер (рН 8,0), содержащего 150 мМ хлористого натрия, 0,05% твин-20, 1% бычьего сывороточного альбумина) в течение 2 часов при 37°С. Затем помещают мембрану в тот же буфер, содержащий разведенные в 500 раз кроличьи поликлональные антитела к ИЛ-2 человека, меченные биотином («NatuTec» Германия), и инкубируют 2 часа при 37°С. Далее трижды промывают мембрану буфером TBST и инкубируют 1 час при 37°С с разбавленным в 3000 раз конъюгатом стрептавидина со щелочной фосфатазой («Силекс М», Москва). После отмывки мембраны буфером PBST (58 мМ двузамещенного фосфата натрия, 17 мМ однозамещенного фосфата натрия, 68 мМ хлористого натрия, 0,1% твин-20) добавляют раствор субстратов для щелочной фосфатазы 0,56 мМ BCIP (5-бром -4-хлор-3-индолилфосфата р-толуидиновая соль), 0,48 мМ NBT (нитротетрозолиум синий) в 10 мМ трис-хлоридном буфере (рН 9,2), содержащем 59,3 мМ хлористого магния. Параллельно окрашивают гели 0,15% раствором кумасси G250 в 25% изо-пропаноле и 10% уксусной кислоте и отмывают в 10% уксусной кислоте. При сравнении спектра белков двух штаммов у штамма PS107(pPIC9HAbIL-2) обнаруживают появление дополнительной белковой полосы с молекулярной массой 82 кДа, дающей четкую положительную реакцию с антителами к ИЛ-2 человека. Молекулярная масса этого секретируемого белка совпадает с теоретически ожидаемой молекулярной массой гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека. Уровень синтеза гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека определяют, сравнивая интенсивность окрашивания полосы рекомбинантного белка с полосой стандартного белка.
-галактозидазу (116,0 кДа), бычий сывороточный альбумин (66,2 кДа), овальбумин (45,0 кДа), лактатдегидрогеназу (35,0 кДа), эндонуклеазу рестрикции Bsp98I (25,0 кДа), р-лактоглобулин (18,4кДа), лизоцим (14,4 кДа). По окончании электрофореза белки ренатурируют, выдерживая гели 15 минут в 10 мМ трис-хлоридном буфере (рН 7,5), содержащем 4 М мочевину, 20 мМ ЭДТА, и переносят на нитроцеллюлозную мембрану в 25 мМ трис-192 мМ глициновом буфере (рН 8,3), содержащем 20% метилового спирта, при 30-40 В, в течение 1,5 часов. Далее мембрану выдерживают в буфере TBST (10 мМ трис-хлоридный буфер (рН 8,0), содержащего 150 мМ хлористого натрия, 0,05% твин-20, 1% бычьего сывороточного альбумина) в течение 2 часов при 37°С. Затем помещают мембрану в тот же буфер, содержащий разведенные в 500 раз кроличьи поликлональные антитела к ИЛ-2 человека, меченные биотином («NatuTec» Германия), и инкубируют 2 часа при 37°С. Далее трижды промывают мембрану буфером TBST и инкубируют 1 час при 37°С с разбавленным в 3000 раз конъюгатом стрептавидина со щелочной фосфатазой («Силекс М», Москва). После отмывки мембраны буфером PBST (58 мМ двузамещенного фосфата натрия, 17 мМ однозамещенного фосфата натрия, 68 мМ хлористого натрия, 0,1% твин-20) добавляют раствор субстратов для щелочной фосфатазы 0,56 мМ BCIP (5-бром -4-хлор-3-индолилфосфата р-толуидиновая соль), 0,48 мМ NBT (нитротетрозолиум синий) в 10 мМ трис-хлоридном буфере (рН 9,2), содержащем 59,3 мМ хлористого магния. Параллельно окрашивают гели 0,15% раствором кумасси G250 в 25% изо-пропаноле и 10% уксусной кислоте и отмывают в 10% уксусной кислоте. При сравнении спектра белков двух штаммов у штамма PS107(pPIC9HAbIL-2) обнаруживают появление дополнительной белковой полосы с молекулярной массой 82 кДа, дающей четкую положительную реакцию с антителами к ИЛ-2 человека. Молекулярная масса этого секретируемого белка совпадает с теоретически ожидаемой молекулярной массой гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека. Уровень синтеза гибридного белка, состоящего из альбумина плазмы крови человека и ИЛ-2 человека определяют, сравнивая интенсивность окрашивания полосы рекомбинантного белка с полосой стандартного белка.