Патент на изобретение №2305931
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) СПОСОБ РЕГЕНЕРАЦИИ РАСТЕНИЙ КЛЕВЕРА ЛУГОВОГО ПРИ ГЕНЕТИЧЕСКОЙ ТРАНСФОРМАЦИИ
(57) Реферат:
Изобретение относится к области сельского хозяйства и может быть использовано в селекции растений для направленного создания исходного материала клевера лугового. Проводят культивирование трансформированной морфогенной культуры клевера на питательной среде Гамборга В5 с добавлением канамицина и цефотаксима, причем морфогенную культуру получают без образования дедифференцированной ткани путем культивирования гипокотиля на питательной среде Гамборга В5 с 4,0 мг/л 6-бензиламинопурина, 1,0 мг/л кинетина и 0,05 мг/л
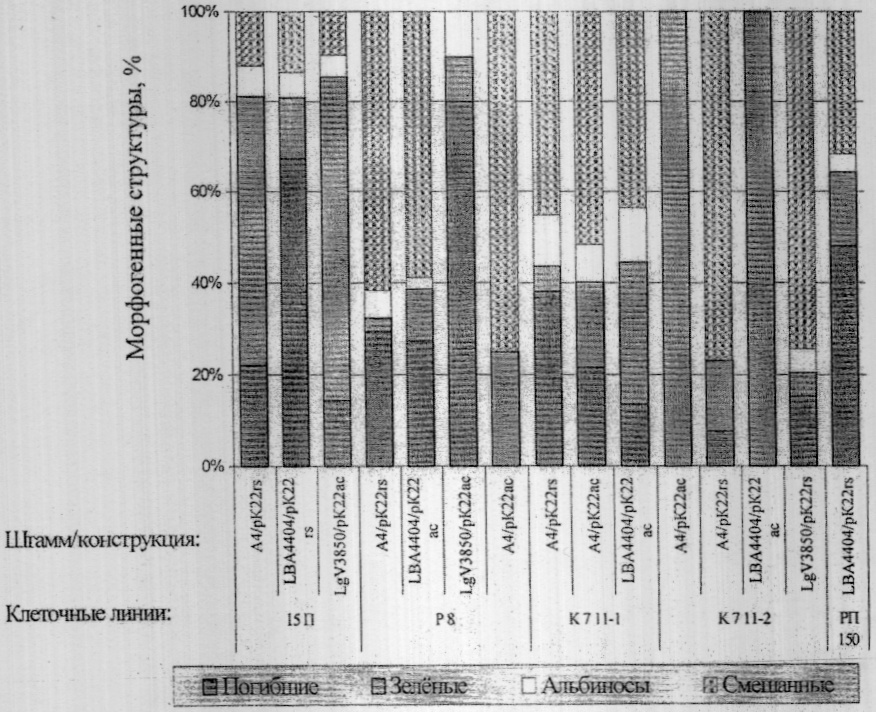
Изобретение относится к области сельского хозяйства и может быть использовано в селекции растений для направленного создания исходного селекционного материала клевера лугового с заданными признаками методами генетической трансформации, в исследованиях по физиологии, фитопатологии и генетике растений. В литературе имеются сведения о регенерации растений клевера лугового в культуре тканей in vitro. Регенерацию растений осуществляли из каллусной ткани, образующейся на поверхностях разрезов листовых эксплантов (1). К недостаткам данного способа можно отнести следующее. В этих экспериментах для получения морфогенных культур использовали каллусную ткань, индуцированную из листовых эксплантов на питательной среде Гамборга В5 с высоким содержанием фитогормонов (6 мг/л 2,4-дихлорфеноксиуксусной кислоты (2.4-Д) и 8 мг/л кинетина). Растения-регенеранты клевера лугового, полученные из морфогенных культур одной и той же клеточной линии (клеточные культуры, полученные из каллусной ткани одного экспланта) вышеназванным способом, различались между собой по ряду морфологических и физиолого-биохимических признаков. При этом морфогенные культуры к 5-8 пассажу теряли свою регенерационную способность. Известен способ регенерации растений клевера лугового в процессе генетической трансформации с использованием разобщенных доминирующих центров (2). В соответствии с этим способом регенерацию растений осуществляют посредством получения дедифференцированной ткани (каллуса) на поверхностях разрезанных на две части (в середине гипокотиля) 7-10-дневных асептических проростков. Для индукции каллусогенеза в среду Гамборга B5 добавляют 2 мг/л 2.4-Д и 0,2 мг/л кинетина. Индукцию морфогенеза из каллусов осуществляют, заменяя вышеназванные гормоны бензила минопурином (БАП) в концентрации 0,2 мг/л. Недостатком данного способа является то, что полученная таким образом морфогенная ткань и растения-регенеранты клевера лугового отличались высокой гетерогенностью и, как следствие, большой сомаклональной изменчивостью по ряду морфологических и физиологических признаков как в положительную, так и отрицательную сторону по сравнению с исходным генотипом. Кроме того, в связи с тем, что для получения каллусов используют как гипокотильную, так и эпикотильную части проростков, исходный генотип не сохраняется и становится невозможным контролировать сомаклональную изменчивость и в дальнейшем проводить полноценный сравнительный морфологический и цитологический анализ полученных в процессе генетической трансформации растений. В связи с этим данный способ регенерации растений оказался не пригоден для направленного создания методом генетической трансформации форм клевера лугового с одним из признаков (устойчивость к болезням, вредителям и т.д.) и сохраняющих при этом все ценные селекционные признаки исходного материала. Цель изобретения – разработка способа прямой регенерации растений клевера лугового, сохраняющих высокий морфогенетический потенциал и ценные селекционные признаки исходных генотипов при длительном культивировании на канамицинсодержащих питательных средах в процессе генетической трансформации. В предлагаемом способе поставленная цель достигается тем, что культивирование трансформированной морфогенной культуры клевера на питательной среде Гамборга В5 осуществляется с добавлением канамицина и цефотаксима, причем морфогенную культуру получают без образования дедифференцированной ткани путем культивирования гипокотиля на питательной среде Гамборга B5 с 4,0 мг/л 6-бензиламинопурина, 1,0 мг/л кинетина и 0,05 мг/л Способ осуществляется следующим образом: из сортообразцов клевера лугового проводили отбор генотипов с высокой регенерационной способностью. Для этого из семян клевера с ценными селекционными признаками получали 6-7-дневные асептические проростки. Эпикотили отделяли и культивировали in vitro для сохранения исходного растения на среде Гамборга B5 без гормонов или для микроразмножения на среде того же состава с 2 мг/л 6-бензиламинопурина (БАП). Гипокотили помещали на питательную среду Гамборга B5 с 4,0 мг/л БАП, 1 мг/л кинетина и 0,05 мг/л Пример 1. Оценка регенерационной способности различных линий клевера лугового после длительного культивирования на среде с канамицином (24 пассаж). Для генетической трансформации использовали только генотипы клевера лугового с высокой регенерационной способностью. Для чего морфогенные культуры с небольшими побегами, разрезанные на части величиной 3-5 мм, помещали срезом вверх на среду Гамборга В5 с 2,0 мг/л БАП, инокулировали штаммами агробактерий, несущих различные конструкции с маркерным (канамицинустойчивость) и целевым генами. Через 5 суток морфогенные культуры переносили на свежую питательную среду того же состава с добавлением 50 мг/л канамицина (для отбора клеточных культур клевера с встроенными генами канамицинустойчивости) и 500 мг/л цефотаксима для подавления роста бактерии. Субкультивирование на средах с цефотаксимом проводили через каждые 3-4 недели до полной элиминации агробактерий. Канамицин добавляли в среды в продолжение всего процесса трансформации и поддержания in vitro коллекции трансформированных морфогенных культур клевера лугового. На чертеже представлены результаты изучения регенерационной способности пяти генотипов клевера лугового сорта Ранний-2 (15П, Р8, К7 11-1, К7 11-2, РП-150) с ценными селекционными признаками (кислотоустойчивость, раннеспелость, высокая семенная продуктивность и т.д.), инокулированных шестью штаммами Agrobacterium tumefaciens и A.rhizogenes с конструкциями A4/pK22rs, A4/pK22ac, LBA 4404/pK22rs, LBA 4404/рК22ас, LGV 3850/pK22rs, LGV 3850/рК22ас. Через 24 пассажа на питательной среде Гамборга B5 без цефотаксима с 50 мг/л канамицина морфогенные культуры клевера лугового сохраняли способность к активной пролиферации, массовому образованию зеленых побегов и корней. В течение одного пассажа (1 месяц) из морфогенной ткани с одной чашки Петри получали в среднем более 100 зеленых побегов, и общее число их составляло 1000 и более штук и ограничивалось только потребностями практической селекции и площадью в фитотроне. Пример 2. Сравнительное изучение цитогенетических (табл.1) характеристик исходных растений и растений-регенерантов клевера лугового, полученных из каллусных культур и методом прямой регенерации из гипокотилей. Из таблицы 1 видно, что количество хромосомных и хроматидных перестроек в клетках растений-регенерантов клевера лугового сорта Ранний-2 клеточной линии РП 150, полученных методом прямой регенерации из гипокотилей, по всем показателям достоверно не отличались от контроля (исходные растения с ценными селекционными признаками), тогда как в варианте РП 116 число хроматидных перестроек с фрагментами и мостами в процентах возросло с 3,4±0,81 до 6,0±0,93 и 0,2±0,19 до 0,76±0,34 соответственно, а общий процент аберраций составил 8,15±1,07 против 3,8±0,85 в контроле. Кроме того, о значительных нарушениях в хромосомном аппарате клеток в процессе регенерации из каллусной культуры свидетельствовало появление хромосомных мостов (0,3±0,21), существенное снижение митотического индекса (Mi) до 4,6±0,66 и достоверное уменьшение числа диплоидных клеток (2n=14) с 98,0±0,99 до 91,0±2,02. Пример 3. Сравнительная оценка морфологических признаков растений исходных генотипов и растений-регенерантов, полученных методом прямой регенерации. Как следует из представленных результатов (табл.2), морфологические и биологические характеристики регенерантов полностью соответствуют исходным генотипам. По всем показателям представленных в таблице 2 признаков растения, полученные методом прямой регенерации, не отличаются от исходного генотипа и, при этом, разброс их характеристик укладывается в 5% ошибку опыта и соответствует величине вариации данных признаков у растений исходного сорта Ранний-2. Это свидетельствует об отсутствии сомаклональной вариабельности у растений-регенерантов РП 150 и их полной идентичности исходному генотипу. Таким образом, предлагаемый способ регенерации позволяет в процессе генетической трансформации на основе одного и того же генотипа, используя конструкции, несущие различные целевые гены, направленно создавать целый ряд генетических аналогов, полностью сохраняя при этом без изменения все ценные признаки исходного образца клевера лугового; получать и поддерживать in vitro морфогенные культуры, способные к длительной пролиферации на селективных средах и массовой регенерации канами-цинустойчивых растений клевера лугового; контролировать сомаклональную изменчивость и проводить сравнительный генетический анализ полученных в процессе генетической трансформации селекционных номеров. Источники информации 1. А.В.Мезенцев. Способ размножения клевера красного in vitro. / Авторское свидетельство СССР №781035, 1978.
Формула изобретения
1. Способ регенерации растений клевера лугового при генетической трансформации, включающий культивирование трансформированной морфогенной культуры клевера на питательной среде Гамборга В5 с добавлением канамицина и цефотаксима, причем морфогенную культуру получают без образования дедифференцированной ткани путем культивирования гипокотиля на питательной среде Гамборга В5 с 4,0 мг/л 6-бензиламинопурина, 1,0 мг/л кинетина и 0,05 мг/л 2. Способ по п.1, отличающийся тем, что морфогенные культуры в процессе генетической трансформации сохраняют способность к регенерации растений и ризогенезу до 24-го пассажа и более на питательной среде Гамборга B5 с 50 мг/л канамицина. 3. Способ по п.1, характеризующийся тем, что полученные из гипокотильных эксплантов морфогенные культуры по генетическим и морфологическим признакам не отличаются от исходных генотипов.
РИСУНКИ
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 -нафтилуксусной кислоты с дальнейшей пересадкой эксплантов на свежую среду того же состава с 2,0 мг/л 6-бензиламинопурина, а эпикотильную часть проростков сохраняют путем культивирования на среде Гамборга В5 без гормонов или путем микроразмножения на среде того же состава с добавлением 2 мг/л 6-бензиламинопурина. Изобретение позволяет провести прямую регенерацию клевера лугового с сохранением морфогенетического потенциала и ценных селекционных признаков исходных генотипов 2 з.п. ф-лы, 1ил., 2 табл.
-нафтилуксусной кислоты с дальнейшей пересадкой эксплантов на свежую среду того же состава с 2,0 мг/л 6-бензиламинопурина, а эпикотильную часть проростков сохраняют путем культивирования на среде Гамборга В5 без гормонов или путем микроразмножения на среде того же состава с добавлением 2 мг/л 6-бензиламинопурина. Изобретение позволяет провести прямую регенерацию клевера лугового с сохранением морфогенетического потенциала и ценных селекционных признаков исходных генотипов 2 з.п. ф-лы, 1ил., 2 табл.