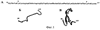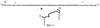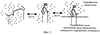Патент на изобретение №2304146
|
|||||||||||||||||||||||||||
(54) СПОСОБ СТАБИЛИЗАЦИИ ПЕТЛЕОБРАЗНОЙ СТРУКТУРЫ СИНТЕТИЧЕСКИХ ПЕПТИДОВ С ПОМОЩЬЮ N- И C-КОНЦЕВЫХ МОДИФИКАЦИЙ
(57) Реферат:
Изобретение относится к технологиям стабилизации петлеобразной структуры синтетических пептидов и может использоваться в фармакологии для получения препаратов нового поколения, предназначенных для диагностики, терапии и вакцинопрофилактики инфекционных заболеваний и патологических состояний человека и животных. Предлагается способ стабилизации петлеобразной структуры синтетических пептидов путем нековалентного взаимодействия гидрофобных углеводородов или их радикалов между собой, причем гидрофобные углеводороды или их радикалы располагают на противоположных N и С концах пептида, либо в непосредственной близости от них и названные гидрофобные углеводороды или их радикалы также нековалентно связывают с гидрофобным носителем. Изобретение решает задачу осуществления стабилизации петлеобразных структур пептидов, не затрагивая аминокислотную последовательность пептида, а также обеспечивает возможность нековалентного связывания стабилизированного синтетического пептидного антигена с носителем, при этом способ является универсальным и может использоваться для стабилизации различных структур. 8 з.п. ф-лы, 3 ил., 2 табл.
Изобретение относится к технологиям стабилизации изгибов и петлеобразных структур синтетических пептидов и может использоваться в фармакологии для получения препаратов нового поколения, предназначенных для диагностики, терапии и вакцинопрофилактики инфекционных заболеваний и патологических состояний человека и животных. Синтетические пептидные антигены все шире используются как ключевые компоненты качественно новых препаратов для диагностики, терапии и вакцинопрофилактики, так как они лишены недостатков, характерных для традиционно используемых рекомбинантных и природных антигенов, таких как патогенность, остаточная вирулентность, неполная инактивация, пирогенность, сопутствующие неспецифические антигены и т.д. Тем не менее синтетические пептиды имеют ряд специфических особенностей, затрудняющих их успешное использование в диагностике, терапии и вакцинопрофилактике, а именно: 1. Низкомолекулярные синтетические пептиды, неконъюгированные с носителем, как правило, не способны вызывать иммунный ответ [Hermanson GT, Mallia AK, Smith PK. «Immobilized Affinity Ligand Techniques» San Diego: Academic Press; 1992]. Этот способ стабилизации имеет следующие недостатки: существует высокая вероятность формирования нецелевых межмолекулярных дисульфидных связей; накладываются ограничения на аминокислотный состав целевого пептида, поскольку структура пептида не должна содержать внутренних цистеинов, иначе возможно формирование нецелевых внутримолекулярных дисульфидных связей; полученный таким образом пептид не является готовым прекурсором для связывания его с носителем. Известен также способ стабилизации бета-изгибов коротких пептидов, который основан на склонности аминокислотных последовательностей к формированию тех или иных вторичных структур пептидной цепи, стабилизированных внутримолекулярными нековалентными взаимодействиями (водородными связями, гидрофобными взаимодействиями, солевыми мостиками). Гидрофобные взаимодействия при нормальных условиях в водной среде самые сильные из всех нековалентных взаимодействий. Одним из примеров стабилизации бета-изгибов в коротких пептидах является триптофановый зиппер [A.G.Cochran, N.J.Skelton, M.A.Starovasnik “Triptophan zippers: stable, monomeric Прототип обладает следующими недостатками: – стабилизация изгиба пептидной цепи зачастую требует вмешательства в аминокислотную последовательность пептида, что может привести к потере аффиности комплексов антиген-антитело; – полученный пептид не является готовым прекурсором для связывания его с носителем; – способ пригоден только для стабилизации бета-изгибов. Изобретение решает задачу создания способа стабилизации изгибов и петлеобразных структур пептидов, позволяющего не затрагивать аминокислотную последовательность пептида, а также создавать пептиды, являющиеся готовым прекурсором для связывания их с носителями, при этом универсального, пригодного и для стабилизации изгибов и петлеобразных структур пептидов, имеющих произвольную аминокислотную последовательность. Поставленная задача решается тем, что предлагается способ стабилизации вторичной петлеобразной структуры синтетических пептидов путем нековалентного взаимодействия гидрофобных углеводородов или их радикалов между собой, в соответствии с которым гидрофобные углеводороды или их радикалы располагают на противоположных N и С концах пептида, либо в непосредственной близости от них, причем названные гидрофобные углеводороды или их радикалы также нековалентно связывают с гидрофобным носителем. При ковалентном связывании не менее чем двух сильно гидрофобных углеводородных радикалов, находящихся с противоположных N и С концов пептидной цепи, происходит изгибание пептидной цепи за счет сближения ее N и С концов и формирование нековалентного взаимодействия между N и С концевыми гидрофобными углеводородными радикалами, а также формирование нековалентного взаимодействия названых гидрофобных углеводородных радикалов с гидрофобньм носителем. Гидрофобные углеводородные радикалы могут быть ковалентно связаны с реакционноспособными группами пептида, содержащимися на его противоположных N и С концах или в непосредственной близости от них, причем реакционноспособными группами пептида могут являться карбоксильные группы, или амидные группы, или тиоловые группы, или иные реакционноспособные группы, способные обеспечить селективное ковалентное связывание гидрофобных углеводородных радикалов с молекулой пептида. Носитель может быть выполнен из полимера, им может быть высокомолекулярная структура из амфипатических молекул, а также возможны иные решения. Присоединяемые к концам пептидов гидрофобные углеводородные радикалы могут быть алифатическимим углеводородами или ароматическими углеводородами. На фиг.1А приведена аминокислотная последовательность пептида, обозначенного здесь как пептид №1, соответствующая фрагменту 89-119 белка Е вируса клещевого энцефалита (штамм 205). Звездочкой помечены аминокислоты Cys в положениях 92 и 116, по тиольным группам которых проводилось модифицирование пептида. На фиг.1Б приведено схематическое изображение пространственной структуры пептида №1. На фиг.2А приведена первичная структура пептида, обозначенного здесь №1-М – модифицированного пальмитиновыми кислотами пептида №1. Звездочкой помечены модифицированные пальмитиновой кислотой аминокислоты Cys в положениях 92 и 116. На фиг.2Б схематично изображена пространственная структура пептида №1-М. Синтетический пептид модифицируют путем ковалентного связывания производных гидрофобных углеводородов с карбоксильными, амидными либо тиоловыми группами, содержащимися на N и С концах пептида (пример 1). Обязательным условием такой модификации является формирование пары сильно гидрофобных углеводородных радикалов на противоположных концах пептидной цепи, как показано на фиг.2. Такая модификация, с одной стороны, приводит к формированию нековалентной внутримолекулярной связи между N- и С- концами пептида, а с другой стороны, позволяет осуществить нековалентное связывание полученного модифицированного пептида с гидрофобным носителем, как показано на фиг.3. В предлагаемом способе происходит стабилизация изгибов или петлеобразных структур пептида за счет внедрения дополнительных концевых молекул, способных осуществить внутримолекулярную связь, что подтверждается высокой аффинностью антипептид №1-М антител к нативному антигену (Пример 6), а также происходит связывание синтетического пептида с гидрофобными носителями за счет нековалентных взаимодействий (пример 3). Таким образом, предлагаемый способ имеет следующие преимущества: – возможность стабилизации как изгибов пептидной цепи, так и петлеобразных структур за счет нековалентных взаимодействий; – возможность связывания пептида с гидрофобными носителями и поверхностями; – способность образовывать высокомолекулярные структуры (мицелы) без участия дополнительных высокомолекулярных носителей. Ниже следуют примеры, раскрывающие сущность изобретения. Пример 1. Твердофазный синтез пептида №1.
Таким образом был изготовлен пептид, аминокислотная последовательность которого соответствует фрагменту 89-119 белка Е вируса клещевого энцефалита. Этот пептид является прекурсором для получения модифицированного пальмитиновыми аминокислотами пептида, способного формировать стабильную петлеобразную структуру. Пример 2. Синтез модифицированного пептида №1-М. Пептид №1-М, как уже упоминалось, это модифицированный пальмитиновыми кислотами пептид №1. Первичная структура пептида №1-М изображена на фиг.2А. Модифицирование пептида №1 проводилось по тиольным группам аминокислот Cys в положениях 92 и 116, эти положения на фиг.1А помечены звездочками. Для изготовления модифицированного пептида №1-М смесь 1,0 г (10 ммоль) малеинового ангидрида и 2,4 г (10 ммоль) цетиламина в 20 мл диоксана кипятили над 2 г молекулярных сит (4Å) 5 ч, охлаждали до комнатной температуры и отфильтровывали выпавший осадок N-гексадецилмалеамида. Выход 2,7 г (80%), т.пл. 58-59°С. Часть полученного продукта (1 г) нагревали в открытой пробирке в токе аргона 1 ч при 120°С, охлаждали, растворяли в 10 мл гексана и отфильтровывали от нерастворившегося остатка. Фильтрат упаривали и сушили в вакууме. Выход 0,8 г (85%). Найдено: С 74,02; Н 11,02; N 4,18. С20Н35NO2. Вычислено: С 74,12; Н 10,97; N 4,36. Растворяли 20 мг пептида №1 в смеси 4 мл этанола и 0,5 мл воды, добавляли раствор 10 мг N-гексадецилмалеимида в 0,2 мл этанола и выдерживали при 35°С до негативного теста на SH-группы с нитропруссидом натрия (5 ч). Реакционную массу упаривали в вакууме, промывали эфиром, сухим изопропанолом, лиофилизовали из воды с добавлением 10% уксусной кислоты. Выход модифицированного пептида №1-М 18 мг. Пример 3. Коньюгирование модифицированного пептида №1-М. Помимо того, что гидрофобные радикалы, входящие в состав пептида №1-М, обеспечивают стабилизацию изгиба пептидной цепочки, они же обеспечивают нековалентное связывание молекул модифицированного пептида с другими гидрофобными средами. Так при физиологических значениях рН через 1-2 суток в растворе пептида №1-М формируются высокомолекулярные структуры (эмульсия). Эти высокомолекулярные структуры представляют собой агрегировавшие молекулы пептида №1-М за счет нековалентного взаимодействия гидрофобных радикалов, входящих в состав соседних молекул пептида. Пептид №1-М обладал более высокой по сравнению с немодифицированным пальмитиновыми кислотами пептидом №1 способностью к адсорбции на поверхности гидрофобного полимера – полистирола. Кроме того, гидрофобные радикалы, входящие в состав пептида №1-М, способны фиксировать пептид на поверхности гидрофобных микрочастиц, как показано на фиг.3. Микрочастицы, представляющие собой мелкие катионные мицеллы, были изготовлены на основе коммерческого катионного амфифила DDAB – /Dimethyl-dioctadecyi – ammonium bromide (ICN)/ и неионного амфифила Tween 80. По данным спектротурбидиметрического анализа микрочастицы имели размер 200 нм. Коньюгирование модифицированного пептида №1-М с микрочастицами проводили инжектированном смеси пептида и микрочастиц через пористую мембрану с диаметром пор 0,5 мкм. Полноту коньюгирования пептида №1-М с микрочастицами подтверждали данными электрофореза на содержание свободного пептида. Схематическое изображение самопроизвольного процесса стабилизации петлеобразной структуры пептида №1-М, аналогичной нативной структуре фрагмента 89-119 белка Е вируса клещевого энцефалита, представлено на фиг.3. Таким образом было проведено коньюгирование пептида №1-М как с катионньми микрочастицами и гидрофобной поверхностью полистирола, так и молекул пептида №1-М между собой с образованием эмульсий. Способность пептида №1-М формировать высокомолекулярные структуры (эмульсии) и самопроизвольно связываться с гидрофобньми микрочастицами позволяет с легкостью изготавливать субстанции, содержащие пептидный антиген и способные вызывать иммунный ответ. Пример 4. Приготовление поликлональных антипептидных антител. Аффинность взаимодействия антиген-антитело очень чувствительна к структуре антигена. Это свойство антиген-антительного взаимодействия использовалось для подтверждения того, что предлагаемый способ стабилизации действительно позволяет стабилизировать изгиб или петлеобразную структур пептида, аналогичную нативной. Для этого были получены антипептидные антитела мышей. Мыши линии BALB/c в возрасте 5-6 недель иммунизировались внутрибрюшинно трехкратно изготовленными пептидами в составе катионных микрочастиц, разведенными в фосфатном буферном растворе. Мышам вводилось по 0,2 мл раствора пептидов в дозе 50 мкг/гол, вторая и третья иммунизации проводились через 2 и 4 недели, соответственно, после первой иммунизации. Контрольной группе животных вводили раствор пептида, мимикрирующего антигенный участок белка gp120 вируса иммунодифицита человека, по той же схеме и в тех же дозировках. Через 10 дней после последней иммунизации у животных из ретроорбитальной вены брали пробу крови для определения титра специфических антител в сыворотке. Для определения титра специфических антител в лунках микропланшетов для иммуноферментного анализа («Медполимер», г.Москва) иммобилизировали следующие антигены: 1) пептид №1 или 2) пептид №1-М. Пептидные антигены растворяли в 0,05 М карбонат-бикарбонатном буферном растворе, рН 9,6, до конечной концентрации 10 мкг/мл, вносили в каждую лунку микропланшета по 0,1 мл раствора и инкубировали в течение ночи при температуре 37°С. Лунки промывали 3 раза фосфатно-буферным раствором с твин 20 и блокировали неспецифическое связывание 0,5%-ным раствором бычьего сывороточного альбумина. Двукратные разведения проб сывороток крови мышей в трис-HCl буферном растворе наносили по 0,1 мл в лунки с иммобилизованным антигеном и инкубировали при 37°С в течение 30 мин, после чего лунки промывали 5 раз фосфатным буферным раствором. Образовавшиеся комплексы антиген-антитело выявляли путем нанесения в лунки микропланшета по 0,1 мл фосфатного буферного раствора, содержащего конъюгированные пероксидазой хрена антитела козы против иммуноглобулинов класса G мыши (Sigma), инкубирования при 37°С в течение 30 мин и добавления субстратной смеси, содержащей орто-фенилендиамин. Ферментную реакцию останавливали добавлением раствора серной кислоты, результат реакции учитывали по оптической плотности, измеренной с помощью планшетного ридера «Мультискан ЕХ» на длине волны 492 нм. Титром сыворотки считали максимальное разведение, при котором оптическая плотность в 2 раза превышала соответствующий сигнал в лунке с отрицательным контролем. В качестве контроля использовали сыворотки животных из контрольной группы. Результаты определения титра антипептидных антител в сыворотке крови мышей представлены в таблице 1.
Таким образом, трехкратная внутрибрюшинная иммунизация мышей пептидами, мимикрирующими фрагмент белка Е ВКЭ вызывает образование сывороточных антител, специфичных к пептидам (иммунизирующий антиген). Пример 5. Приготовление нативного антигена вируса клещевого энцефалита. Для получения нативного антигена вируса КЭ, штамм 205, осветленную вирусную суспензию очищали ультрацентрифугированием в комбинированном градиенте глицерина и тартрата калия, в котором верхнюю часть образует 30%-ный (вес/вес) раствор глицерина в STE-буфере (50 мМ трис-HCl, 1 мМ ЭДТА (рН 7,5)), а нижнюю (более плотную) 40%-ный (вес/вес) раствор тартрата калия в STE-буфере (рН 8,5). На приготовленный градиент наслаивали вирусную суспензию и центрифугировали при 100000 g в течение 3 часов при температуре +4°С на центрифуге L8-70 (Beckman). Фракцию, содержащую вирус («бэнд»), разводили в равном объеме STE буфера (рН 7,5) и центрифугировали в течение 2 часов при 100000 g при температуре +4°С. Осадок вируса растворяли в боратном буфере (0,15М NaCl, 50 mM Н3ВО3, рН 9,0) и хранили при -70°С. Концентрацию общего белка оценивали при помощи набора “Bio-Rad Protein Assay Kit”. Чистоту выделения вируса проверяли методом электрофореза в 12% полиакриламидном геле с 0,1% додецилсульфата натрия (Саминский, 1966). Окраску гелей проводили при помощи красителя Кумасси G-250. Две полосы на электрофореграмме соответствовали белкам Е (54 кД) и NS1 (41 kD) вируса КЭ. Для получения антигенов вируса КЭ очищенную вирусную суспензию обрабатывали 0,2% раствором додецилсульфата натрия. Пример 6. Определение аффинности антипептидных антител к нативному антигену. Аффинность взаимодействия антиген-антитело очень чувствительна к структуре антигена. Высокие значения аффинности антител однозначно указывают на то, что антиген представлен в той пространственной структуре, на которую были получены антитела. Иными словами, та структура пептидного антигена, которая вызовет формирование антисыворотки, обладающей наибольшей аффинностью к нативному антигену, имеет пространственную структуру, наиболее сходную с нативной. Для анализа взаимодействия специфических антипептидных антисывороток на полистероловые планшеты наносили 1 мкг нативного антигена вируса клещевого энцефалита в 100 мкл 0,05 М раствора двухзамещенного фосфатного буферного раствора, рН 8, и инкубировали при 37°С в течение 1 часа. Места неспецифического связывания насыщали 0,5% раствором казеина при 20°С в течение 60 минут. Затем инкубировали со специфическими антипептидными антисыворотками 1 час при 37°С. Образовавшиеся комплексы антиген-антитело выявляли путем нанесения в лунки микропланшета по 0,1 мл фосфатного буферного раствора, содержащего конъюгированные пероксидазой хрена антитела козы против иммуноглобулинов класса G мыши (Sigma), инкубирования при 37°С в течение 30 мин. Ферментную реакцию останавливали добавлением раствора серной кислоты, результат реакции учитывали по оптической плотности, измеренной с помощью планшетного ридера «Мультискан ЕХ» на длине волны 492 нм. Аффинность взаимодействие антиген-антитело характеризуется значением константы диссоциации комплекса антиген-антитело (Kd) – чем меньше Kd тем выше аффинность. Для определения Kd результаты измерения оптических плотностей при различных разведениях сыворотки представляли в координатах Скэтчарда (А, А/С), где А – оптическая плотность, а С – концентрация антител (сыворотки). Предполагалось, что имеет место взаимодействие антител с двумя типами связывающих центров: специфическое связывание антител с антигеном и неспецифическое взаимодействие антитела с пластиком. Тангенсы углов наклона касательных к кривой Скэтчарда соответствуют значениям Kd1 -1 специфического взаимодействия антиген-антитело и (Kd1 -1+Kd1 -2) специфического и неспецифического взаимодействия. Для двух типов связывающих центров, при условии, что один из них является высокоспецифичным, а второй неспецифическим, уравнение взаимодействия в координатах Скэтчарда имеет вид [Ворфоломеев С.Д., Зайцев С.В. «Кинетические методы в биохимических исследованиях». – Издательство Московского университета, 1982]: у=(((R1-x)/K1+(R2-x)/K2+(((R1-x)/K1+(R2-x)/K2)^2+4*x*(R1+R2-x)/(K1*K2)^0,5)/2), где у – количество комплексов антиген-антитело/концентрация антител; х – количество высокоспецифических комплексов и количество низкоспецифических комплексов; K1, K2 – константы диссоциации высокоспецифических и низкоспецифических комплексов; R1, R2 – количество высокоспецифических и низкоспецифических центров связывания. Рассчитанные с помощью уравнения Скэтчарда значения констант диссоциации высокоспецифического взаимодействия антипептидных антисывороток с нативным антигеном представлены в таблице 2.
Таким образом, антисыворотки, полученные на пептид №1-М, обладают меньшей Kd, или большей аффинностью к нативному антигену. Следовательно, N и С концевые модификации приводят к формированию пространственной структуры, сходной с нативной петлеобразной структурой района 89-119 белка Е вируса клещевого энцефалита. Иными словами, модифицирование N и С концов, приводящие к формированию гидрофобных радикалов по концам пептида, обеспечивает стабилизацию петлеобразной пространственной структуры пептида, аналогичной нативной петлеобразной структуре мимикрируемого района. Из приведенных примеров видно, что предлагаемый способ стабилизации изгибов и петлеобразных структур пептидов позволяет не затрагивать аминокислотную последовательность пептида, а также создавать пептиды, являющиеся готовым прекурсором для связывания их с носителями. При этом способ универсален, пригоден и для стабилизации изгибов и петлеобразных структур пептидов, имеющих произвольную аминокислотную последовательность.
Формула изобретения
1. Способ стабилизации петлеобразной структуры синтетических пептидов путем нековалентного взаимодействия гидрофобных углеводородов или их радикалов между собой, отличающийся тем, что гидрофобные углеводороды или их радикалы располагают на противоположных N и С концах пептида либо в непосредственной близости от них, причем названные гидрофобные углеводороды или их радикалы также нековалентно связывают с гидрофобным носителем. 2. Способ по п.1, отличающийся тем, что гидрофобные углеводороды или их радикалы ковалентно связывают с реакционноспособными активными группами пептида, содержащимися на его противоположных N и С концах или в непосредственной близости от них. 3. Способ по п.2, отличающийся тем, что реакционноспособными активными группами пептида, с которыми связывают углеводороды или их радикалы, являются карбоксильные группы. 4. Способ по п.2, отличающийся тем, что реакционноспособными активными группами пептида, с которыми связывают углеводороды или их радикалы, являются амидные группы. 5. Способ по п.2, отличающийся тем, что реакционноспособными активными группами пептида, с которыми связывают углеводороды или их радикалы, являются тиоловые группы. 6. Способ по п.1, отличающийся тем, что носитель выполнен из гидрофобного полимера. 7. Способ по п.1, отличающийся тем, что носителем является высокомолекулярная структура из амфипатических молекул. 8. Способ по п.1, отличающийся тем, что углеводороды или их радикалы являются алифатическими. 9. Способ по п.1, отличающийся тем, что углеводороды или их радикалы являются ароматическими.
РИСУНКИ
MM4A – Досрочное прекращение действия патента СССР или патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе
Дата прекращения действия патента: 18.10.2008
Извещение опубликовано: 20.03.2010 БИ: 08/2010
NF4A – Восстановление действия патента СССР или патента Российской Федерации на изобретение
Дата, с которой действие патента восстановлено: 27.04.2010
Извещение опубликовано: 27.04.2010 БИ: 12/2010
|
|||||||||||||||||||||||||||
 -hairpins”. PNAS, 98, (10), p.5578-5583; 2001]. Стабилизация бета-изгиба обуславливается гидрофобными взаимодействиями боковых групп пар молекул триптофана, расположенных симметрично относительно центра изгиба. Этот способ принят за прототип изобретения.
-hairpins”. PNAS, 98, (10), p.5578-5583; 2001]. Стабилизация бета-изгиба обуславливается гидрофобными взаимодействиями боковых групп пар молекул триптофана, расположенных симметрично относительно центра изгиба. Этот способ принят за прототип изобретения. -Trt для Asn и Gln; Nim-Trt для His; S-Trt для Cys; tBu для Thr и Ser; tBu-эфир для Asp; NG-Pbf для Arg; N
-Trt для Asn и Gln; Nim-Trt для His; S-Trt для Cys; tBu для Thr и Ser; tBu-эфир для Asp; NG-Pbf для Arg; N 3PO4; препаративную – на колонке 10×250 мм с фазой SynChropak RPP 100, 10 мкм (Eichrom Ind.), при скорости потока 4 мл/мин в линейном градиенте концентрации ацетонитрила (0-66%) в воде с 0,1% трифторуксусной кислоты. Детекцию осуществляли спектрофотометрически по поглощению при 226 нм с обработкой результатов с использованием системы MultiChrom (Ampersand). Фракции, содержащие основное вещество, упаривали в вакууме с изопропанолом и лиофилизовали из воды, содержащей 10% уксусной кислоты. Выход 42 мг (13%, для TFA-соли с м.в. 3250). Содержание SH-групп по реакции с 2,2′-дипиридилдисульфидом 0,58 мкмоль/мг (94,5%). При помощи препаративной ВЭЖХ был выделен мажорный продукт с чистотой около 85%, идентичность которого была доказана при помощи аминокислотного анализа. Элементный анализ проводили на CHN-анализаторе Perkin-Elmer, аминокислотный – на приборе Biotronik LC5001 после гидролиза пробы в 6 н. HCl, 110°, 24 ч. Количественное определение сульфгидрильных групп осуществляли при помощи теста с 2,2′-дипиридилдисульфидом (Carlsson et al., 1978), качественное – по реакции с нитропруссидом натрия.
3PO4; препаративную – на колонке 10×250 мм с фазой SynChropak RPP 100, 10 мкм (Eichrom Ind.), при скорости потока 4 мл/мин в линейном градиенте концентрации ацетонитрила (0-66%) в воде с 0,1% трифторуксусной кислоты. Детекцию осуществляли спектрофотометрически по поглощению при 226 нм с обработкой результатов с использованием системы MultiChrom (Ampersand). Фракции, содержащие основное вещество, упаривали в вакууме с изопропанолом и лиофилизовали из воды, содержащей 10% уксусной кислоты. Выход 42 мг (13%, для TFA-соли с м.в. 3250). Содержание SH-групп по реакции с 2,2′-дипиридилдисульфидом 0,58 мкмоль/мг (94,5%). При помощи препаративной ВЭЖХ был выделен мажорный продукт с чистотой около 85%, идентичность которого была доказана при помощи аминокислотного анализа. Элементный анализ проводили на CHN-анализаторе Perkin-Elmer, аминокислотный – на приборе Biotronik LC5001 после гидролиза пробы в 6 н. HCl, 110°, 24 ч. Количественное определение сульфгидрильных групп осуществляли при помощи теста с 2,2′-дипиридилдисульфидом (Carlsson et al., 1978), качественное – по реакции с нитропруссидом натрия.