Патент на изобретение №2294633
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) БИОЛОГИЧЕСКИЙ МАРКЕР ДЛЯ ОПРЕДЕЛЕНИЯ ПОРОДЫ РЫБ, НАБОР И СПОСОБ ОПРЕДЕЛЕНИЯ ПОРОДНОЙ ПРИНАДЛЕЖНОСТИ РЫБ
(57) Реферат:
Изобретение относится к биотехнологии и рыбоводству. Предложен способ определения породной принадлежности рыб с использованием полиморфных маркеров. Способ предусматривает сбор образца крови, выделение ДНК и исследование геномной ДНК с помощью ПЦР. При осуществлении способа используют полиморфные маркеры ДНК разного размера или длины. Визуализацию ДНК проводят при электрофорезе в акриламидном или агарозном геле. Предложены также биологические маркеры и набор для осуществления такого способа. 3 н. и 3 з.п. ф-лы, 2 ил., 2 табл.
Изобретение относится к биотехнологии, в частности генетическим исследованиям, к области, связанной с породной идентификацией рыб и их гибридов, а также для прижизненного генетического мониторинга природных и искусственных популяций рыб путем анализа полиморфизма дезоксирибонуклеиновой кислоты. Известен способ идентификации рыб и их гибридов, в котором в качестве экстракта белков используют слизь, взятую путем соскоба с наружной поверхности тела, и используют гели из полиакриламида, содержащего 4-метилумбеллиферилацетат, а электрофорез и сканирование проводят одновременно (а.с. СССР №1611303, БИ №45, 1990). Известен способ определения родства живых организмов, включающий выделение ДНК, обработку рестрикционными эндонуклеазами, электрофоретическое фракционирование фрагментов ДНК, перенос ДНК из геля, молекулярную гибридизацию ДНК из живого организма (а.с. СССР №1552642, 1994). Известен способ селективной амплификации ДНК, олигонуклеотид и набор для селективной амплификации (Патент РФ №2182176, 1992). Недостатком известных способов является непригодность их для породной идентификации рыб. Наиболее близким к предлагаемому способу определения пород карповых рыб и биологического маркера для их характеристики является способ подбора особей русского осетра ACIPENCER guldensatdti в аквакультуре, включающий свенс-анализ митохондриальной ДНК в области D-петли методом ПЦР-амплификации, в которой в процессе анализа используется прямой (OS1) и обратный (OS2) праймеры, а для воспроизводства подбирают особей, различающихся по длине ПЦР-продуктов мтДНК (патент РФ №2202178, 2001). Недостатками этого известного способа является невозможность определения породной принадлежности рыб, отсутствие праймеров и набора для идентификации и паспортизации карповых рыб, невозможность создания генетического паспорта породы рыб. Задачей предлагаемого изобретения является определение породной принадлежности и установление происхождения распространенных и практически значимых одомашненных форм различных рыб, в том числе из семейства карповых (Cyprinidae). В результате использования предлагаемого биологического маркера для определения породы рыб, набора и способа идентификации и паспортизации пород рыб появляется возможность создания генетического паспорта породы и повышение эффективности идентификации в генетико-селекционных мероприятиях, связанных с разведением рыб. Вышеуказанный результат достигается тем, что биологический маркер, содержащий полиморфную ДНК и характеризующий породную принадлежность, происхождение распространенных и практически значимых природных и одомашненных форм рыб семейства карповых и представляющий собой нуклеотидную последовательность из группы 5 пар праймеров следующего состава
Результат достигается также тем, что используют набор для определения породной принадлежности рыб, содержащий биологический маркер, реакционную смесь, состоящую из 60 мМ трис-HCl, 10 мМ сульфата аммония, 0,1% TWEEN 20, по 100 мкМ каждого dNTP, 0,5 мкМ MgCl2, 0,9 единиц Taq-полимеразы и эталонные ДНК для каждой породы. В способе определения породной принадлежности рыб семейства карповых, включающем сбор образца крови, выделение ДНК из него, исследование геномной ДНК с помощью ПЦР, ПЦР-анализ проводят используя предлагаемый набор, получают полиморфные маркеры ДНК разного размера, визуализируют эти маркерные ДНК с помощью электрофореза в акриламидном или агарозном геле, затем для типирования выявленных в изученной группе аллелей проводят сравнение генетического разнообразия определяемой и контрольной группы образцов амплифицируемой ДНК и по отсутствию или присутствию в исследуемой группе определенных единичных породоспецифических аллелей или аллельных сочетаний определяют породную принадлежность.
При этом для каждой породы и породной группы карповых рыб составляют генетический паспорт, представленный в таблице 1, в которой указано распределение 50-75 полиморфных микросателлитных маркеров, выявленных с помощью 5 пар праймеров (PR 4, 10, 19, 24, 26) по пяти соответствующим локусам (PRL 4, 10, 19, 24, 26), и по которой определяют породу рыб. Размер амплифицируемых продуктов определяют с помощью маркера молекулярных масс (10 bp Ladder). Набор контрольных амплификатов содержит образцы амплифицированной ДНК, характерные для каждой из изученных пород карповых рыб, в том числе карпа или сазана. В способе верификацию породной принадлежности проверяют с помощью многомерного анализа (например, дискриминантного анализа, метода главных компонент и т.п.), в котором в качестве обучающей выборки служит выборка аллелей и аллельных сочетаний одной из набора пород рыб. Способ генетической идентификации пород рыб, в том числе рыб семейства карповых, в том числе пород карпа и сазана с применением предлагаемых биологических маркеров и набора осуществляют следующим образом. Осуществляют геномный фингерпринтинг для выявления геномной вариабельности рыб и для идентификации и паспортизации различных пород рыб с использованием метода полимеразной цепной реакции (ПЦР или PCR). В предлагаемом способе получают молекулярно-генетические характеристики пород рыб, в том числе карповых рыб, в том числе наиболее распространенных на территории России пород карпа и сазана, предлагают породоспецифичные маркеры с целью паспортизации пород. При изучении пород и породных групп рыб выявляют несколько породоспецифичных маркеров, которые в дальнейшем используют в качестве эталона для определения породной принадлежности любой группы рыб. Для этого собирают образцы различных тканей (кровь, слизь, кусочки плавников, мышцы и т.п.), выделяют из них ДНК, проводят амплификацию определенных участков ДНК с определенными праймерами, визуализируют полученную изменчивость с помощью специального электрофоретического разделения амплификатов и специального окрашивания, проводят типирование полученной изменчивости путем сравнения с эталонными образцами. На заключительном этапе для каждой идентифицируемой группы составляют таблицу-паспорт с указанием распределения 50-70 аллельных вариантов по 5 полиморфным локусам. На основании этой таблицы с высокой вероятностью либо визуально, либо с помощью многомерного статистического анализа определяют принадлежность (или отсутствие принадлежности) исследуемой группы рыб к одной из наиболее распространенных пород. Для генотипирования породы собирают образцы тканей у 20-30 представителей породы (лучше образцы крови) по следующей методике отбора проб. Наилучшим материалом для последующего выделения суммарной ДНК являются: кровь (обычно, из хвостовой вены) и фрагменты плавников (грудных, брюшных, анальных или спинного), которые берут прижизненно, стараясь не наносить излишних повреждений донорам биопсийного материала. Используют также половые продукты рыб (овулировавшие яйцеклетки и сперму производителей), фрагменты ястыков и семенников на различных стадиях созревания. В тех случаях, когда существует возможность получения проб от умертвленных «ex tempora» особей, в качестве материала может дополнительно использоваться мышечная ткань, печень и другие внутренние органы, за исключением кишечника. Для кодирования наиболее часто используемых видов тканей предлагается использовать систему, принятую в Российской Национальной Коллекции эталонных генетических материалов (РНКЭГМ), которая демонстрируется табл.2.
Консервация (фиксация) проб. Для предотвращения деградации ДНК, полученные пробы помещают в фиксирующий раствор. Как правило, это – 70-96% этиловый спирт (этиловый ректификованный, высшей очистки). Мышцы, фрагменты плавников, печени, ястыков и семенников перед этанольной фиксацией ножницами, скальпелем или полулезвием безопасной бритвы измельчают до кусочков, размеры которых соответствуют приблизительно 0,5 см3. Первичную фиксацию осуществляют при соотношении спирт/образец 10/1. Вторичную – в среднем, через трое-четверо суток в соотношении 5/1. Кровь собирают из хвостовой вены при отрезании хвостового плавника (или шприцом). Капли крови собирают в стерильную пробирку с консервантом (0,5 М ЭДТА, рН 8,0) соотношение крови и консерванта 1:10. Не допускается попадания в пробирки слизи и других посторонних примесей, способствующих образованию в пробирке сгустков. Образцы, зафиксированные и перезалитые этанолом, могут храниться при комнатной температуре в течение 30 и более суток. Для более продолжительного хранения их следует поместить в свежий спиртовой раствор (не ниже 92%) в соотношении спирт/образец 5/1. Образцы крови, зафиксированные 0,5 М ЭДТА, хранят в холодильнике в течение 30 суток при низких плюсовых (2-4°С) температурах, но образец не должен подвергаться замораживанию. После замораживания все образцы хранят в холодильнике при отрицательных (не выше -50°С) температурах без размораживания. Но не дольше 90 суток. Для более продолжительного хранения пригодны кельвинаторы, поддерживающие температуру ниже -50°С, а также жидкоазотные сосуды Дюара. Выделение и хранение ДНК. 1. Выделение ДНК из крови. Образцы свежей крови, законсервированные в 0,05 М растворе ЭДТА (рН 8.0), хранят при температуре +4°С. Выделение ядерной ДНК проводили по методу Мэтью (Mathew, 1984). Выделение ДНК заключается в последовательном прохождении следующих фаз: лизис клеток и ядер, депротеинизация ДНК с помощью протеиназы К, фенол-хлороформная депротеинизация, осаждение и растворение ДНК. Для получения более очищенных образцов ДНК проводят дополнительную очистку с помощью набора реагентов DIAtom 2. Лизис клеток. К образцу крови добавляют лизирующий буфер в соотношении 1 объем крови: 30 объемов буфера. Состав лизирующего буфера: 320 мМ сахарозы, 1% тритон Х-100, 5 мМ MgCl2 и 10 мМ трис-HCl (рН 7,6). Смесь инкубируют 1 час 30 минут, затем центрифугируют 15 минут при 4°С (2000 g). Супернатант сливают, а осадок подсушивают в течение 30 минут при комнатной температуре. 3. Суспендирование и лизис ядер. К полученному осадку добавляют 4 мл буфера (25 мМ ЭДТА и 75 М NaCl). Затем проводят суспендирование ядер резким встряхиванием пробирки в течение 10 минут. К суспензии добавляют 0,4 мл 10% SDS (рН 8,0) и выдерживают в течение 10 минут, периодически плавно перемешивая. По завершению лизиса ядер к суспензии добавляют по каплям протеиназу К до конечной концентрации 50 мкг/мл, осторожно перемешивают и инкубируют смесь 4 часа при 45°С или оставляют на ночь при 37°С. 4. Депротеинизация ДНК. В пробирку со смесью, содержащей лизированные ядра, добавляют 0,3 мл 5 М ацетата натрия или 0,3 мл 5 М ацетата аммония, или 0,5 мл 3 М перхлората натрия и аккуратно перемешивают. К смеси добавляют равный объем фенола, насыщенного трис-HCl (рН>7,6), плавно перемешивают 5 минут и приливают 5 мл водонасыщенного хлороформа, снова плавно перемешивают 5 минут и центрифугируют при 4°С в течение 20 минут (2000 g). Верхнюю фазу с растворенной ДНК отбирают пипеткой, стараясь не захватывать интерфазу, содержащую белки. Фенол-хлороформная депротеинизация повторяется 2-3 раза до полной очистки раствора ДНК от белков. На следующей стадии к отобранному супернатанту добавляют 10 мл водонасыщенного хлороформа, плавно перемешивают 5 минут и центрифугируют в течение 10 минут (2000 g) и отбирают верхнюю фазу с растворенной в ней ДНК. 5. Осаждение ДНК. К 1 мл растворенной ДНК приливают 20 мкл 5 М ацетата натрия и осторожно перемешивают. Затем добавляют тройной объем 96° этанола и плавно перемешивают до выпадения ДНК в виде тонких белых нитей или белого осадка. ДНК однократно промывают в 70° этаноле для удаления избытка соли, затем – в 90° этаноле. На следующем этапе спирт сливают, ДНК высушивают на воздухе до полного испарения спирта. Высушенную ДНК растворяют в 100 мкл дистиллированной воды. 6. Дополнительная очистка ДНК В случае некачественного выделения ДНК из крови препараты ДНК необходимо дополнительно очистить. Один из методов дополнительной очистки состоит в следующем: к 40 мкл растворенной ДНК добавляют 160 мкл лизирующего агента и перемешивают содержимое. После термостатирования в течение 5-7 минут при температуре 65°С смесь центрифугируют и супернатант переносят в чистую пробирку. Далее в супернатант добавляют 8 мкл суспензии сорбента NucleoS В отдельных случаях для быстроты можно использовать различные наборы для выделения ДНК. Ниже приводим метод выделения ДНК из различных тканей рыб с помощью одного из таких наборов, а именно набора реагентов Diatom Набор реагентов Diatom Продолжительность выделения ДНК из жидких проб составляет 25 минут. Выделение ДНК из сухих пятен или мелкоизмельченного твердого материала может длиться до нескольких часов в зависимости от обрабатываемого материала. 7. Аналитический электрофорез Для определения концентрации и качества выделенных образцов используют метод электрофореза в агарозном геле. Гель-электрофорез проводится в горизонтальной камере в 1х трис-боратном буфере (ТВЕ), рН 7,5-7,8 (0,089 М трис-бората, 0,089 М борной кислоты и 0,002 М ЭДТА). При внесении ДНК в карманы геля используется буфер для нанесения (ксиленцианол, бромфеноловый синий и глицерин). После электрофореза гель окрашивают в растворе бромистого этидия в течение 5-10 минут или добавляют бромистый этидий непосредственно в гель. Вхождение ДНК в гель осуществляют при 20 V в течение 15-20 минут, далее электрофорез ведут при 60 V в течение 2-3 часов. В качестве маркера концентрации используется ДНК фага Полимеразная цепная реакция (PCR). 1. Проведение PCR. Выделенную ДНК используют в качестве матрицы в реакции амплификации. Амплификацию с единичными праймерами проводят по стандартной методике на термоциклере «MJ Research» (USA), имеющем металлический нагревательный блок на 60 образцов. Реакционная смесь объемом 25 мкл содержит 60 мМ трис-HCl, 10 мМ сульфата аммония, 0,1% TWEEN 20, по 100 мкМ каждого dNTP, 0,5 мкМ MgCl2, 0,1 мкМ каждого праймера, 0,9 единиц Taq-полимеразы и 25-100 нг тотальной ДНК. Для проведения PCR используют пять пар праймеров следующего состава:
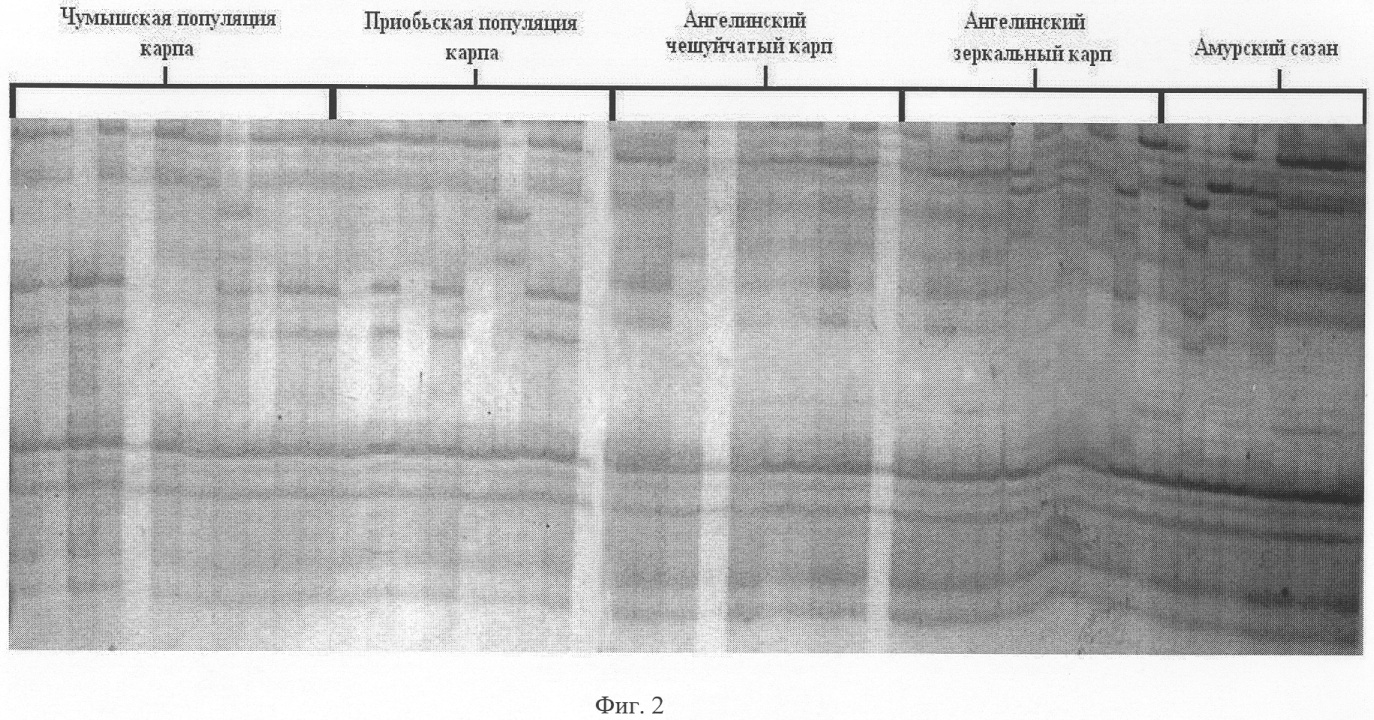
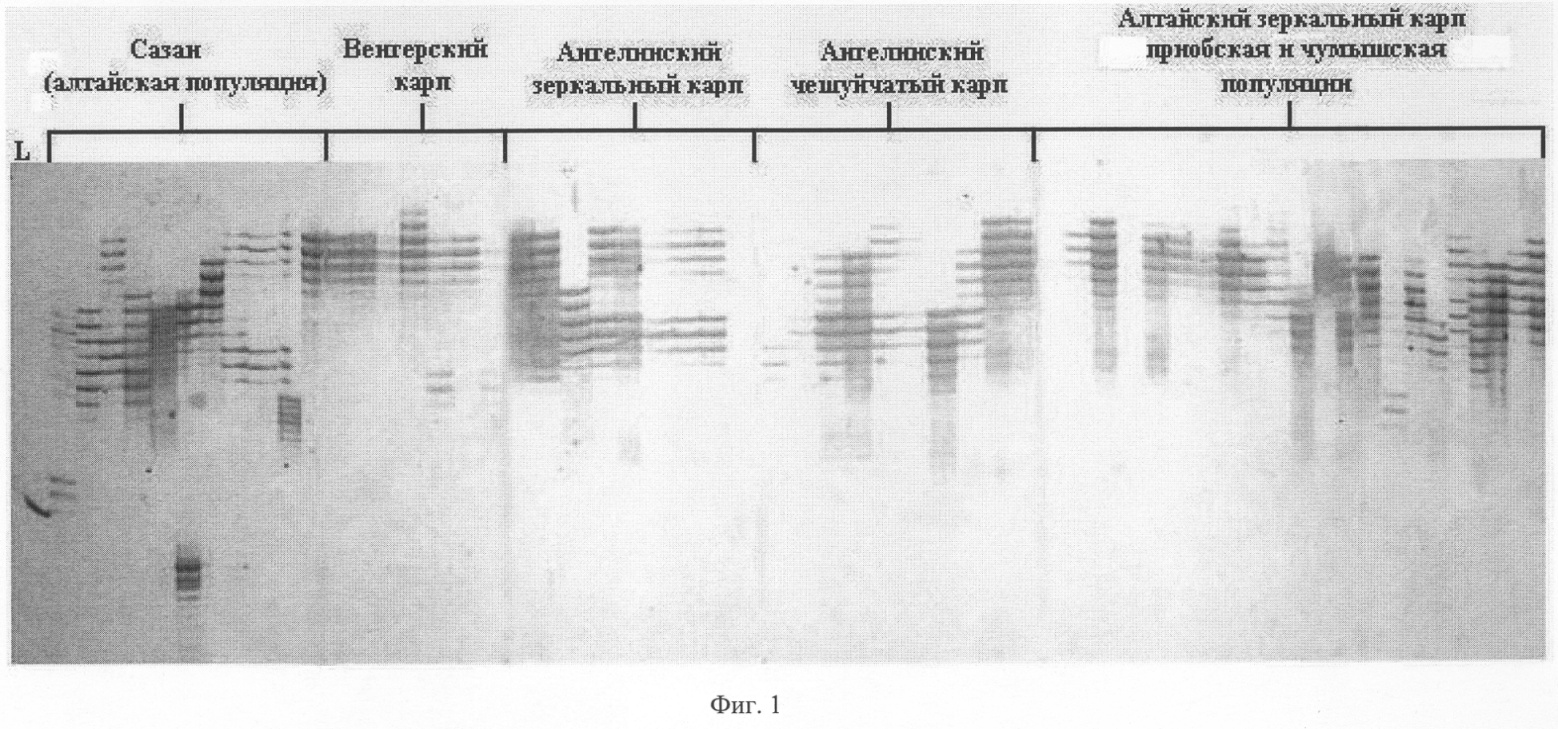
Для предотвращения испарения с поверхности раствора во время амплификации в каждую пробирку добавляют каплю (около 15 мкл) минерального масла. Каждый цикл полимеразной реакции состоит из следующих этапов денатурации (94°С, 2 мин), отжига (36°С, 1 мин) и элонгации (72°С, 2 мин). Циклы повторяются 35 раз. Первый цикл предваряли денатурацией в течение 5 минут при температуре 94°С. После окончательной достройки амплифицированной ДНК (72°С, 2 мин) температуру снижают до 4°С. 2. Электрофоретическое разделение продуктов и визуализация ПЦР-продуктов. Продукты амплификации подвергают аналитическому электрофорезу в 1.5% агарозном геле толщиной 7-10 мм, содержащем бромистый этидий. В качестве маркеров молекулярного веса используется 50 bp Ladder (Fermentas). Вхождение в гель осуществляют при 20 V в течение 20 минут, а разделение фрагментов – в течение 10-20 часов при 40-50 V. В случае эффективной амплификации продукты амплификации подвергают электрофоретическому разделению в вертикальной камере в 6% ПААГе, содержащем 7% мочевину. Визуализацию осуществляют путем окрашивания гелей азотнокислым серебром из набора Silver Sequence Staining Kit [Promega, USA] согласно инструкции производителя, с последующим фотографированием цифровым фотоаппаратом Nicon. Размер амплифицируемых продуктов определяют с помощью маркера молекулярных масс (50 bp Ladder). Затем проводят статистическую обработку результатов и составляют породный генетический паспорт. Для типирования (определения размера в п.н.) выявленных в изученной группе рыб аллелей необходимо провести обязательное сравнение генетического разнообразия данной и контрольной (эталонной) группы образцов амплифицируемой ДНК. Набор контрольных амплификатов может содержать от 5 до 20 образцов амплификатов, характерных для 5-20 микросателлитных аллелей. Отсутствие или присутствие в исследуемой группе определенных аллелей или аллельных сочетаний позволяет надежно определять породную принадлежность. Верификацию породной принадлежности проводят с помощью различных вариантов многомерного (дискриминантного анализа или метода главных компонент), в котором в качестве обучающей выборки может служить выборка аллелей и аллельных сочетаний одной из 9 ранее изученных пород карпа и сазана. Список пород и породных групп карпа и сазана, которые могут служить обучающей выборкой и для которых известно генетическое разнообразие и породоспецифичные аллели: карп зеркальный и чешуйчатый Ангелинский, карп Венгерский, карп Ропшинский, Черепецкий рамчатый и чешуйчатый карп, ставропольский карп, Алтайский и Амурский сазан. На фиг.1 представлен набор полиморфных ДНК, выявляемых с помощью праймера PR-10 у различных пород карпа. На фиг.2 представлен набор полиморфных ДНК, выявляемых с помощью праймера PR-24 у различных пород карпа. Примеры выполнения предлагаемого способа. Пример 1. Способ определения породы карпа с помощью праймера PR-10 Из образцов крови, полученных из особей Венгерского карпа, Алтайского карпа, Ангелинского карпа (зеркального и чешуйчатого) и Алтайского сазана, выделяют ДНК способом, указанным выше. На каждой из выделенных ДНК осуществляют ПЦР с одной из пяти пар предлагаемых праймеров, а именно PR-10. Полученные амплификационные продукты визуализируют путем электрофореза в гелях, и после окрашивания гелей серебром идентифицируют по размеру (длине) выявляемые полиморфные маркеры. На фиг.1 представлены полиморфные маркеры для каждой породы Венгерского карпа (дорожки 11-17), Алтайского карпа (дорожки 36-56), Ангелинского зеркального (дорожки 18-25) и чешуйчатого (дорожки 26-35) карпа и Алтайского сазана (дорожки 1-10). Каждая из этих пород характеризуется определенным набором или сочетанием аллелей, часть из которых представлена в таблице 1. Это распределение служит эталоном при сравнении и идентификации неизвестных пород карповых рыб. Путем сравнения распределения полиморфных ДНК разной длины среди исследуемых и эталонных образцов определяют породную принадлежность. Пример 2. Способ определения породы карпа с помощью праймера PR-24. Из образцов крови, полученных из Чумышской и Приобской популяции Алтайского карпа, Ангелинского чешуйчатого карпа и Ангелинского зеркального карпа, Амурского сазана, выделяют ДНК способом, указанным выше. На каждый из выделенных ДНК осуществляют ПЦР с одной из пяти пар предлагаемых праймеров, а именно PR -10. Полученные амплификационные продукты визуализируют путем электрофореза в гелях, и после окрашивания гелей серебром идентифицируют по размеру (длине) выявляемые полиморфные маркеры. На фиг.2 представлены полиморфные маркеры для каждой породы Чумышской (дорожки 1-10) и Приобской (дорожки 11-20), популяции Алтайского карпа, Ангелинского чешуйчатого карпа (дорожки 21-30), и Ангелинского зеркального карпа (дорожки 31-40), Амурского сазана (дорожки 41-49). Каждая из этих пород характеризуется определенным набором или сочетанием аллелей, часть из которых представлена в таблице 1. Это распределение служит эталоном при сравнении и идентификации неизвестных пород карповых рыб. Путем сравнения распределения полиморфных ДНК разной длины среди исследуемых и эталонных образцов определяют породную принадлежность. Генетический паспорт составляется для каждой известной породы и породной группы и представлен в таблице 1, в которой указано распределение 50-70 полиморфных маркеров по 5 парам праймеров. Аналогичный паспорт можно составить для любой выборки карпа неизвестного происхождения.
Формула изобретения
1. Биологический маркер, содержащий полиморфную ДНК и характеризующий породную принадлежность, происхождение распространенных и практически значимых природных и одомашненных форм рыб семейства карповых и представляющий собой нуклеотидную последовательность из группы 5 пар праймеров следующего состава:
2. Набор для определения породной принадлежности рыб семейства карповых, содержащий биологический маркер по п.1, реакционную смесь, состоящую из 60 мМ трис-HCl, 10 мМ сульфата аммония, 0,1% TWEEN 20, по 100 мкМ каждого dNTP, 0,5 мкМ MgCl2, 0,9 единиц Taq-полимеразы и эталонные ДНК для каждой породы. 3. Способ паспортизации пород рыб семейства карповых с использованием полиморфных маркеров, включающий сбор образца крови, выделение ДНК из него, исследование геномной ДНК с помощью ПЦР, отличающийся тем, что ПЦР-анализ проводят, используя набор по п.2, получают полиморфные маркеры ДНК разного размера или длины, визуализируют эти ДНК при электрофорезе в акриламидном или агарозном геле, затем для типирования выявленных в изученной группе аллелей проводят сравнение генетического разнообразия определяемой и контрольной группы образцов амплифицируемой ДНК, и по отсутствию или присутствию в исследуемой группе определенных единичных породоспецифических аллелей или аллельных сочетаний определяют породную принадлежность, при этом для каждой породы и породной группы рыб составляют генетический паспорт, представленный в таблице 1, в которой указано распределение 50-75 полиморфных микросателлитных маркеров, выявленных с помощью 5 пар праймеров (PR 4, 10, 19, 24, 26) по пяти соответствующим локусам (PRL 4, 10, 19, 24, 26), и по которой определяют породу рыб. 4. Способ по п.3, отличающийся тем, что размер амплифицируемых продуктов определяют с помощью маркера молекулярных масс (10 bp Ladder). 5. Способ по п.3, отличающийся тем, что набор контрольных амплификатов содержит образцы амплифицированной ДНК, характерные для каждой из изученных пород карповых рыб, в том числе карпа или сазана. 6. Способ по пп.3, 4 и 5, отличающийся тем, что верификацию породной принадлежности проверяют с помощью многомерного анализа, например путем дискриминантного анализа, метода главных компонент, в котором в качестве обучающей выборки служит выборка аллелей и аллельных сочетаний одной из набора пород рыб.
РИСУНКИ
MM4A – Досрочное прекращение действия патента СССР или патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе
Дата прекращения действия патента: 26.03.2007
Извещение опубликовано: 10.07.2008 БИ: 19/2008
NF4A – Восстановление действия патента СССР или патента Российской Федерации на изобретение
Дата, с которой действие патента восстановлено: 27.04.2009
Извещение опубликовано: 27.04.2009 БИ: 12/2009
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
 Prep 200 (Москва), основанного на использовании лизирующего реагента с гуанидинтиоционатом, в присутствии которого ДНК сорбируется на сорбенте и отмывается затем раствором Экстра-Гена
Prep 200 (Москва), основанного на использовании лизирующего реагента с гуанидинтиоционатом, в присутствии которого ДНК сорбируется на сорбенте и отмывается затем раствором Экстра-Гена , внесенная в разных количествах, например 0,5; 1; 2 мкг.
, внесенная в разных количествах, например 0,5; 1; 2 мкг.