Патент на изобретение №2293117
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) ГЕН STREPTOMYCES AVERMITILIS И ЕГО ПРИМЕНЕНИЕ ДЛЯ ИЗМЕНЕНИЯ СООТНОШЕНИЯ АВЕРМЕКТИНОВ B2:B1
(57) Реферат:
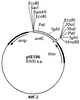
Изобретение относится к области биотехнологии и может быть использовано при производстве антибиотиков авермектинов. Изобретение относится к полинуклеотиду, содержащему нуклеотидные последовательности, кодирующие продукт гена aveC, молекулы которого можно использовать для изменения соотношения количества авермектинов класса 2:1, продуцируемых в ферментационных культурах S. avermitilis. Нуклеотидная последовательность полинуклеотида по существу аналогична последовательности аллеля aveC S. avermitilis, последовательности плазмиды pSE186 (ATCC 209604), кодирующей продукт гена AveC S. avermitilis, нуклеотидной последовательности (НП) открытой рамки считывания (ORF) aveC, но при этом НП содержит мутации, кодирующие комбинацию замен аминокислот, благодаря чему штамм S. avermitilis ATCC 53692, в клетках которого инактивирован аллель aveC дикого типа, способен продуцировать авермектины класса 2:1 в соотношении 0,35:1 или меньше (когда авермектины являются циклогексилом В2 и циклогексилом В1 соответственно). Изобретение относится также к векторам, клеткам-хозяевам и мутантным штаммам S. avermitilis, в которых ген aveC инактивирован или мутирован с возможностью изменения соотношения количества авермектинов класса 2:1. Изобретение касается способа получения мутированного штамма (комбинации замен аминокислот приводятся в формуле) и способа получения авермектинов класса 2:1 в уменьшенном соотношении, а также композиции, полученной указанным способом. Использование изобретения позволяет получить композицию авермектинов класса 2:1 со сниженным количеством в смеси авермектина В2. 7 н. и 10 з.п. ф-лы, 6 ил., 4 табл.
Описание 1. Область техники Настоящее изобретение относится к композициям, содержащим авермектины, и способам эффективного получения авермектинов, таких как “дорамектин”, которые находят широкое применение в ветеринарии. В частности, настоящее изобретение относится к молекулам полинуклеотидов, содержащим нуклеотидные последовательности, кодирующие продукт гена AveC, который можно использовать для модуляции отношения авермектинов класса 2:1, полученных ферментацией культур Streptomyces avermitilis. Настоящее изобретение далее относится к векторам, трансформированным клеткам-хозяевам и новым мутантным штаммам S. avermitilis, в которых ген aveC мутирован с возможностью модуляции соотношения продуцируемых авермектинов класса 2:1. 2. Предпосылки изобретения 2.1. Авермектины Виды из рода Streptomyces продуцируют целый ряд вторичных метаболитов, в том числе авермектины, которые образуют серию из восьми родственных шестнадцатичленных макроциклических лактонов, обладающих сильной антигельминтной и инсектицидной активностью. Восемь разных, но близко родственных соединений обозначены как А1а, А1b, A2a, A2b, B1a, B1b, B2a и B2b. Серия “а” указанных соединений включает природный авермектин, в котором заместитель в положении С25 является (S)-втор-бутилом, и серия “b” включает такие соединения, в которых заместитель в положении С25 является изопропилом. Символы “А” и “В” служат для обозначения авермектинов, в которых заместитель в положении С5 является соответственно метоксильной и гидроксильной группой. Цифрой “1” обозначены авермектины, у которых в положении С22,23 находится двойная связь, и цифрой “2” обозначены авермектины, содержащие атом водорода в положении С22 и гидроксильную группу в положении С23. Считается, что среди родственных авермектинов особенно сильной противопаразитарной и пестицидной активностью обладает авермектин типа В1, такой как дорамектин, который является наиболее привлекательным в коммерческом отношении. Авермектины и способы их получения методом аэробной ферментации штаммов S. avermitilis описаны в патентах США №№ 4310519 и 4429042. Считается, что биосинтез природных авермектинов инициируется эндогенно СоА-аналогами сложного тиоэфира изомасляной кислоты и S-(+)-2-метилмасляной кислоты. Улучшение штамма в результате неспецифического мутагенеза и использование экзогенно вводимых жирных кислот сделало возможным эффективное продуцирование аналогов авермектина. Мутанты S. avermitilis, в которых отсутствует 3-кетокислота-дегидрогеназа с разветвленной цепью (мутанты с отсутствием bkd), могут продуцировать авермектины только в процессе ферментации с введением жирных кислот. Скрининг и выделение мутантов, не обладающих активностью дегидрогеназы с разветвленной цепью (например, S. avermitilis, АТСС 53567), описаны в европейском патенте (ЕР) № 276103. Ферментация таких мутантов в присутствии экзогенно вводимых жирных кислот позволяет получить только четыре авермектина, соответствующих используемым жирным кислотам. Так, в результате ферментации S. avermitilis (АТСС 53567) с введением S-(+)-2-метилмасляной кислоты образуются природные авермектины А1а, А2а, В1а и В2а; в результате ферментации с введением изомасляной кислоты образуются природные авермектины A1b, A2b, B1b и B2b; и в результате ферментации с введением циклопентанкарбоновой кислоты образуются четыре новых циклопентилавермектина А1, А2, В1 и В2. При введении других жирных кислот образуются новые авермектины. В результате скрининга более 800 предполагаемых предшественников было идентифицировано более 60 новых авермектинов. (См., например, Dutton et al., 1991, J. Antibiot. 44:357-365; and Banks et al., 1994, Roy. Soc. Chem. 147:16-26). Кроме того, мутанты S. avermitilis, не обладающие активностью 5-О-метилтрансферазы, продуцируют в основном только аналоги авермектинов В. Следовательно, мутанты S. avermitilis, у которых отсутствует как 2-кетокислота-дегидрогеназа с разветвленной цепью, так и 5-О-метилтрансфераза, продуцируют только авермектины В, соответствующие жирной кислоте, используемой в процессе ферментации. Таким образом, при добавлении к таким двойным мутантам S-(+)-2-метилмасляной кислоты образуются только природные авермектины В1а и В2а, в то время как при добавлении изомасляной кислоты или циклопентанкарбоновой кислоты образуются соответственно природные авермектины B1b и B2b или новые циклопентилавермектины В1 и В2. Добавление к штамму с двумя мутациями циклогексанкарбоновой кислоты является предпочтительным способом получения коммерчески важного нового авермектина, такого как циклогексилавермектин В1 (дорамектин). Способы выделения и характеристики таких двойных мутантов, например S. avermitilis (АТСС 53692), описаны в европейском патенте № 276103. 2.2. Гены, участвующие в биосинтезе авермектина Обнаружено, что во многих случаях гены, участвующие в продуцировании вторичных метаболитов, и гены, кодирующие определенный антибиотик, образуют в хромосоме совместные кластеры. Именно таким кластером является кластер генов поликетидсинтезы Streptomyces (PKS) (см. Hopwood and Sherman, 1990, Ann. Rev. Genet. 24:37-66). Таким образом, одним методом клонирования генов в процессе биосинтеза является выделение гена лекарственной устойчивости и исследование смежных областей хромосомы с целью обнаружения других генов, имеющих отношение к биосинтезу данного антибиотика. Другим методом клонирования генов, участвующих в биосинтезе важных метаболитов, является комплементация мутантов. Например, части библиотеки ДНК из организма, способного продуцировать определенный метаболит, вводят в мутант, не продуцирующий такой метаболит, и ведут поиск трансформантов, продуцирующих данный метаболит. Кроме того, для идентификации и клонирования генов в процессе биосинтеза используют гибридизацию библиотеки при помощи зондов, выделенных из других видов из рода Strepromyces. Установлено, что гены, участвующие в биосинтезе авермектина (гены ave), подобно генам, необходимым для биосинтеза других вторичных метаболитов Strepromyces (например, PKS), образуют кластеры в хромосоме. Был успешно клонирован ряд генов ave с использованием векторов для комплементации мутантов S. avermitilis, блокируемых в процессе биосинтеза авермектина. Клонирование таких генов описано в патенте США № 5252474. Кроме того, Ikeda et al., 1995, J. Antibiot. 48: 532-534, описали локализацию области хромосомы, имеющей непосредственное отношение к дегидратации в положениях С22,23 (aveC) фрагмента BamHI длиной 4,82 т.п.о. S. avermitilis, а также к мутациям гена aveC, в результате чего образуется однокомпонентный продуцент В2а. Так как ивермектин, сильнодействующее антигельминтное соединение, можно получить химическим путем из авермектина В2а, то однокомпонентный продуцент авермектина В2а считается особенно пригодным для коммерческого получения ивермектина. В патенте США № 6248579, выданном Stutzman-Engwall 19 июня 2001 г., описаны некоторые мутации гена aveC Streptomyces avermitilis, позволяющие уменьшить соотношение авермектинов циклогексил В2 : циклогексил В1 примерно до 0,75:1. В заявке РСТ WO 01/12821 компании Pfizer Products Inc., опубликованной 22 февраля 2001 г., описаны некоторые дополнительные мутации гена AveCa Streptomyces avermitilis, вызывающие дальнейшее уменьшение соотношения авермектинов циклогексил В2 : циклогексил В1 до 0,40:1. Идентификация дополнительных мутаций или комбинаций мутаций в гене AveC, позволяющих значительно упростить продуцирование авермектина, таких как, например, мутации, уменьшающие соотношение авермектинов В2 : В1, дожна облегчить получение и очистку коммерчески важных авермектинов. 3. Краткое изложение существа изобретения Настоящее изобретение относится к молекуле полинуклеотида, содержащей нуклеотидную последовательность, которая по существу аналогична аллелю aveC Streptomyces avermitilis, последовательности плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC S. avermitilis, нуклеотидной последовательности открытой рамки считывания (ORF) aveC S. avermitilis, показанной на фигуре 1 (SEQ ID No.1), или их вырожденному варианту, но при этом имеет мутации, кодирующие комбинацию замен аминокислот в положении аминокислотных остатков, соответствующих положениям аминокислот SEQ ID No.2, благодаря чему клетки штамма S. avermitilis АТСС 53692, в которых инактивирован аллель aveC дикого типа и которые экспрессируют молекулу полинуклеотида, содержащую мутированную нуклеотидную последовательность, способны продуцировать авермектины класса 2:1 с меньшим соотношением по сравнению с соотношением, продуцируемым клетками штамма S. avermitilis АТСС 53692, экспрессирующими только аллель aveC дикого типа, причем, когда авермектины класса 2:1 являются авермектинами циклогексил В2:циклогексил В1, соотношение авермектинов класса 2:1 составляет 0,35:1 или меньше. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,30:1 или меньше. В еще более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,25:1 или меньше. В наиболее предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,20:1 или меньше. В конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию, выбираемую из нижеследующего списка:
Настоящее изобретение далее относится к молекуле полинуклеотида, содержащей нуклеотидную последовательность, которая по существу аналогична аллелю aveC Strepromyces avermitilis, последовательности плазмиды pSЕ186 (АТСС 209604), кодирующей продукт гена AveC S. avermitilis, нуклеотидной последовательности открытой рамки считывания aveC S. avermitilis, показанной на фигуре 1 (SEQ ID No.1), или их вырожденному варианту, но при этом имеет мутации, кодирующие комбинацию замен аминокислот в положении аминокислотных остатков, соответствующих положениям аминокислот SEQ ID No.2, благодаря чему клетки штамма S. avermitilis АТСС 53692, в которых инактивирован аллель aveC дикого типа и которые экспрессируют молекулу полинуклеотида, содержащую мутированную нуклеотидную последовательность, способны продуцировать авермектины класса 2:1 с меньшим соотношением по сравнению с соотношением, продуцируемым клетками штамма S. avermitilis АТСС 53692, экспрессирующими только аллель aveC дикого типа, причем, когда авермектины класса 2:1 являются авермектинами циклогексил В2: циклогексил В1, соотношение авермектинов класса 2:1 уменьшается примерно до 0,40:1 или меньше, и комбинация замен аминокислот включает комбинацию, выбираемую из нижеследующей группы: (bf) D48E, S138T, A139T, G179S, E238D; и (bg) Y28C, Q38R, D48E, L136P, G179S, E238D. Настоящее изобретение далее относится к рекомбинантному вектору, содержащему молекулу полинуклеотида по настоящему изобретению. Настоящее изобретение далее относится к клетке-хозяину, содержащей молекулу полинуклеотида или рекомбинантный вектор по настоящему изобретению. В предпочтительном варианте осуществления изобретения клетка-хозяин является клеткой Streptomyces. В более предпочтительном варианте осуществления изобретения клетка-хозяин является клеткой Streptomyces avermitilis. Настоящее изобретение далее относится к способу получения нового штамма Streptomyces avermitilis, который включает (i) мутирование аллеля aveC в клетке штамма S. avermitilis, вызывающее комбинацию замен аминокислот в продукте гена AveC, или (ii) введение в клетку штамма S. avermitilis мутированного аллеля aveC или его вырожденного варианта, кодирующего продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из пунктов (а)-(be) приведенного выше списка. Настоящее изобретение далее относится к способу получения нового штамма Strepromyces avermitilis, который включает (i) мутирование аллеля aveC в клетке штамма S. avermitilis, вызывающее комбинацию замен аминокислот в продукте гена AveC, или (ii) введение в клетку штамма S. avermitilis мутированного аллеля aveC или его вырожденного варианта, кодирующего продукт гена AveC, включающий комбинацию замен аминокислот, благодаря чему клетки, содержащие мутированный аллель aveC или его вырожденный вариант, способны продуцировать авермектины циклогексил В2 : циклогексил В1 в соотношении 0,35:1 или меньше. В неограничивающем варианте осуществления изобретения мутированный аллель aveC или его вырожденный вариант кодирует продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. В предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,30:1 или меньше. В неограничивающем варианте осуществления изобретения мутированный аллель aveC или его вырожденный вариант кодирует продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (f)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,25:1 или меньше. В неограничивающем варианте осуществления изобретения мутированный аллель aveC или его вырожденный вариант кодирует продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (w)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,20:1 или меньше. В неограничивающем варианте осуществления изобретения мутированный аллель aveC или его вырожденный вариант кодирует продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (ао)-(be) приведенного выше списка. Настоящее изобретение далее относится к способу получения нового штамма Streptomyces avermitilis, который включает (i) мутирование аллеля aveC в клетке штамма S. avermitilis, вызывающее комбинацию замен аминокислот в продукте гена AveC, или (ii) введение в клетку штамма S. avermitilis мутированного аллеля aveC или его вырожденного варианта, кодирующего продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (bf) и (bg) приведенного выше списка. В предпочтительном варианте осуществления изобретения клетки S. avermilitis, содержащие такой мутированный аллель aveC или его вырожденный вариант, способны продуцировать авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,40:1 или меньше. Настоящее изобретение далее относится к клетке вида из рода Streptomyces, которая содержит мутированный аллель aveC S. avermitilis или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. В предпочтительном варианте осуществления изобретения вид из рода Streptomyces является видом из рода S. avermitilis. Настоящее изобретение далее относится к клетке Streptomyces avermitilis, способной продуцировать авермектины циклогексил В2: циклогексил В1 в соотношении 0,35:1 или меньше. В неограничивающем варианте осуществления изобретения указанная клетка содержит мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. В предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2:циклогексил В1 составляет примерно 0,30:1 или меньше. В неограничивающем варианте осуществления изобретения указанные клетки содержат мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (f)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,25:1 или меньше. В неограничивающем варианте осуществления изобретения указанные клетки содержат мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (w)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2:циклогексил В1 составляет примерно 0,20:1 или меньше. В неограничивающем варианте осуществления изобретения указанные клетки содержат мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (ао)-(be) приведенного выше списка. Настоящее изобретение далее относится к клетке вида из рода Streptomyces, которая содержит мутированный аллель aveC S. avermitilis или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (bf) и (bg) приведенного выше списка. В предпочтительном варианте осуществления изобретения вид из рода Streptomyces является видом из рода S. avermitilis. В более предпочтительном варианте осуществления изобретения указанная клетка является клеткой S. avermitilis, способной продуцировать авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,40:1 или меньше. Настоящее изобретение далее относится к способу получения авермектинов, который включает культивирование штамма клеток Streptomyces avermitilis по настоящему изобретению в культуральной среде в условиях, стимулирующих или индуцирующих продуцирование авермектинов, и выделение указанных авермектинов из культуры. Настоящее изобретение далее относится к композиции авермектинов циклогексил В2 : циклогексил В1, продуцируемой клетками Streptomyces avermitilis в культуральной среде, используемой для культивирования клеток, в которой соотношение авермектинов циклогексил В2 : циклогексил В1 составляет 0,35:1 или меньше, предпочтительно около 0,30:1 или меньше, более предпочтительно около 0,25:1 или меньше и наиболее предпочтительно около 0,20:1 или меньше. В конкретном варианте осуществления изобретения композицию авермектинов циклогексил В2 : циклогексил В1 продуцируют клетки штамма S. avermitilis, экспрессирующие мутированный аллель aveC или его вырожденный вариант, кодирующий генный продукт, благодаря чему уменьшается соотношение авермектинов циклогексил В2 : циклогексил В1 класса 2:1, продуцированных указанными клетками, по сравнению с клетками такого же штамма S. avermitilis, которые экспрессируют только аллель aveC дикого типа и не экспрессируют мутированный аллель aveC. В предпочтительном варианте осуществления изобретения, в котором указанная композиция включает авермектины циклогексил В2 : циклогексил В1 в соотношении 0,35:1 или меньше, данную композицию продуцируют клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения, в котором указанная композиция включает авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,30:1 или меньше, данную композицию продуцируют клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (f) и (be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения, в котором указанная композиция включает авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,25:1 или меньше, данную композицию продуцируют клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (w)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения, в котором указанная композиция включает авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,20:1 или меньше, данную композицию продуцируют клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (ао)-(be) приведенного выше списка. Настоящее изобретение далее относится к композиции авермектинов циклогексил В2 :циклогексил В1, продуцированной клетками Streptomyces avermitilis в культуральной среде, используемой для культивирования указанных клеток, в которой соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,40:1 или меньше, а также клетками, содержащими мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (bf) и (bg) приведенного выше списка. 4. Краткое описание чертежей Фигура 1. Последовательность ДНК (SEQ ID No.1), содержащая открытую рамку считывания aveC S. avermitilis и выделенную аминокислотную последовательность (SEQ ID No.2). Фигура 2. Плазмидный вектор pSE186 (АТСС 209604), содержащий полную открытую рамку считывания гена aveC S. avermitilis. Фигура 3. Вектор замещения гена pSE180 (АТСС 209605), содержащий ген ermE Sacc. erythraea, встроенный в открытую рамку считывания aveC S. avermitilis. Фигура 4. Рестрикционная карта BamHI кластера генов поликетидсинтазы авермектина в S. avermitilis с пятью идентифицированными перекрывающимися космидными клонами (то есть pSE65, pSE66, pSE67, pSE68, pSE69). Указана также взаимосвязь между pSE118 и pSE119. Фигура 5. Анализ методом ВЭЖХ продуктов ферментации, продуцированных штаммами S. avermitilis. Количественное определение пиков выполнено путем сравнения со стандартными количествами циклогексилавермектина В1. Время удерживания циклогексилавермектина В2 равно 7,4-7,7 мин; время удерживания циклогексилавермектина В1 равно 11,9-12,3 мин. Фиг. 5А. Штамм SE180-11 S. avermitilis с открытой рамкой считывания инактивированного аллеля aveC. Фиг. 5В. Штамм SE180-11 S. avermitilis, трансформированный плазмидой pSE186 (АТСС 209604). Фиг. 5С. Штамм SE180-11 S. avermitilis, трансформированный плазмидой pSE187. Фиг. 5D. Штамм SE180-11 S. avermitilis, трансформированный плазмидой pSE188. Фигура 6А-М. Компилированный список комбинаций замен аминокислот, кодированных мутациями в аллеле aveC, определенный во втором цикле “перестановки генов”, и их влияние на соотношение продуцируемых авермектинов циклогексил В2 : циклогексил В1. Для каждой плазмиды в верхней блоке колонки, озаглавленной “Мутации”, указаны замены аминокислот и в нижнем блоке перечислены замены нуклеотидных оснований, приводящие к замене указанных аминокислот. Замены нуклеотидных оснований, указанные в скобках, являются молчащими заменами, то есть они не вызывают изменений аминокислотной последовательности. 5. Подробное описание изобретения Настоящее изобретение относится к идентификации и исследованию молекул полинуклеотидов, содержащих нуклеотидные последовательности, кодирующие продукт гена AveC, продуцированного Streptomyces avermitilis, к созданию новых штаммов S. avermitilis, которые можно использовать для скрининга мутированных продуктов гена AveC с целью определения их влияния на продуцирование авермектина, и к открытию того, что некоторые мутированные продукты гена AveC могут уменьшать соотношение авермектинов В2:В1, продуцированных S. avermitilis. В качестве примера в нижеследующих разделах данного изобретения описана молекула полинуклеотида, содержащая нуклеотидную последовательность, которая аналогична последовательности плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC S. avermitilis, или нуклеотидной последовательности открытой рамки считывания, показанной на фигуре 1 (SEQ ID No.1), и молекулы полинуклеотидов, содержащие мутированные нуклеотидные последовательности и их вырожденные варианты. Однако принципы, изложенные в настоящем изобретении, в равной степени применимы к другим молекулам полинуклеотидов, включающим наряду с прочими гомологичные гены aveC, продуцируемые другими видами из рода Streptomyces, например, S. hygroscopicus и D. griseochromogenes. 5.1. Молекулы полинуклеотидов, кодирующие продукт гена AveC S. avermitilis Настоящее изобретение относится к выделенной молекуле полинуклеотида, содержащей полную открытую рамку считывания aveC S. avermitilis или ее значительную часть, причем в указанной молекуле полинуклеотида отсутствует следующая полная открытая рамка считывания, расположенная внизу от открытой рамки считывания aveC in situ в хромосоме S. avermitilis. Выделенная молекула полинуклеотида по настоящему изобретению предпочтительно содержит нуклеотидную последовательность, которая аналогична последовательности плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC S. avermitilis, нуклеотидной последовательности открытой рамки считывания, показанной на фигуре 1 (SEQ ID No.1), или ее значительной части. В используемом здесь значении термин “значительная часть” выделенной молекулы полинуклеотида, содержащей нулеотидную последовательность, кодирующую продукт гена AveC S. avermitilis, означает выделенную молекулу полинуклеотида, которая содержит по крайней мере около 70% последовательности полной открытой рамки считывания aveC, показанной на фигуре 1 (SEQ ID No.1), и кодирует функционально эквивалентный продукт гена AveC. Таким образом, “фукнционально эквивалентный” продукт гена AveC определяется как генный продукт, который при экспрессии в штамме S. avermitilis АТСС 53692, в котором инактивирован нативный аллель aveC, вызывает продуцирование авермектинов по существу в таком же соотношении и количестве, что и при продуцировании штаммом S. avermitilis АТСС 53692, экспрессирующим только функциональный аллель aveC дикого типа, который является нативным для штамма S. avermitilis АТСС 53692. Помимо нуклеотидной последовательности открытой рамки считывания aveC выделенная молекула полинуклеотида по настоящему изобретению может далее содержать нуклеотидные последовательности, которые фланкируют ген aveC in situ в S. avermitilis, в частности последовательности, фланкирующие нуклеотидные последовательности, показанные на фигуре 1 (SEQ ID No.1). Настоящее изобретение далее относится к выделенной молекуле полинуклеотида, содержащей нуклеотидную последовательность SEQ ID No.1 или ее вырожденный вариант на основе известной вырожденности генетического кода. В используемом здесь значении термины “молекула полинуклеотида”, “полинуклеотидная последовательность”, “кодирующая последовательность”, “открытая рамка считывания” и “ORF” означают молекулы ДНК и РНК, которые могут быть одноцепочечными или двухцепочечными и могут быть транскрибированы и транслированы (ДНК) или транслированы (РНК) в продукт гена AveC или полипептид, гомологичный продукту гена AveC, в соответствующей экспрессирующей системе клетки-хозяина под контролем соответствующих регуляторных элементов. Кодирующая последовательность может включать, не ограничиваясь ими, прокариотические последовательности, последовательности кДНК, последовательности геномной ДНК и последовательности химически синтезированных ДНК и РНК. Нуклеотидная последовательность, показанная на фигуре 1 (SEQ ID No.1), содержит четыре разных кодона GTG в положениях пар оснований 42, 174, 177 и 180. Как ранее было описано в патенте США № 6248579, несколько делеций в 5′-концевой области открытой рамки считывания aveC (фигура 1; SEQ ID No.1) были созданы для определения того, какие из указанных кодонов могут функционировать в открытой рамке считывания aveC в качестве сайтов инициации экспрессии белка. Делеция первого сайта GTG в положении пары оснований 42 не устраняла активность гена AveC. Дополнительная делеция всех кодонов GTG в положениях пар оснований 174, 177 и 180 подавляла активность гена AveC, что указывало на необходимость данной области для экспрессии белка. Таким образом, в объем настоящего изобретения входят открытые рамки считывания aveC переменной длины. Настоящее изобретение далее относится к молекуле полинуклеотида, содержащей нуклеотидную последовательность, гомологичную последовательности плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC S. avermitilis, или нуклеотидной последовательности открытой рамки считывания aveC, показанной на фигуре 1 (SEQ ID No.1), или ее значительной части. Термин “гомологичный” применительно к молекуле полинуклеотида, которая гомологична последовательности, кодирующей продукт гена AveC S. avermitilis, означает молекулу полинуклеотида, содержащую нуклеотидную последовательность, которая (а) кодирует такой же продукт гена AveC, что и последовательность плазмиды pSE186 (АТСС 209604), кодирующая продукт гена AveC S. avermitilis, или кодирует такой же продукт гена AveC, что и нуклеотидная последовательность открытой рамки считывания aveC, показанная на фигуре 1 (SEQ ID No.1), но включает одну или несколько молчащих замен в нуклеотидной последовательности в соответствии с вырожденностью генетического кода (то есть вырожденный вариант); или (b) гибридизирует с комплементом молекулы полинуклеотида, содержащей нуклеотидную последовательность, которая кодирует аминокислотную последовательность, кодированную последовательностью плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC, и кодирует аминокислотную последовательность, показанную на фигуре 1 (SEQ ID No.2), в умеренно строгих условиях, то есть при осуществлении гибридизации с фильтрсвязанной ДНК в 0,5 М NaHPO4, 7% додецилсульфата натрия (SDS), 1 мМ EDTA при 65°С и выполнении промывки в 0,2×SSC/0,1% SDS при 42°С (см. Ausubel et al. (eds.), 1989, Current Protocols in Molecular Biology, Vol. I, Green Publishing Associates, Inc., and John Wiley & Sons, Inc., New York, at p. 2.10.3), и кодирует вышеуказанный функционально эквивалентный продукт гена AveC. В предпочтительном варианте осуществления изобретения гомологичная молекула полинуклеотида гибридизирует с комплементом нуклеотидной последовательности плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC, или с комплементом нуклеотидной последовательности открытой рамки считывания aveC, показанной на фигуре 1 (SEQ ID No.1), или ее значительной части в очень строгих условиях, то есть при осуществлении гибридизации с фильтрсвязанной ДНК в 0,5 М NaHPO4, 7% SDS, 1 мМ EDTA при 65°С и выполнении промывки в 0,1×SSC/0,1% SDS при 68°С (Ausubel et al., 1989, см. выше), и кодирует вышеуказанный фукционально эквивалентный продукт гена AveC. Активность продукта гена AveC и его потенциальных функциональных эквивалентов можно определить при помощи анализа методом ВЭЖХ продуктов ферментации, как это описано в нижеследующих примерах. Молекулы полинуклеотидов, содержащие нуклеотидные последовательности, которые кодируют функциональные эквиваленты продукта гена AveC S. avermitilis, могут включать естественные гены AveC, присутствующие в других штаммах S. avermitilis, гомологичные гены AveC, присутствующие в других видах из рода Streptomyces, и мутированные аллели aveC, которые являются естественными или созданными методами рекомбинантных ДНК. Настоящее изобретение далее относится к молекуле полинуклеотида, содержащей нуклеотидную последовательность, которая кодирует полипептид, имеющий аминокислотную последовательность, гомологичную аминокислотной последовательности, кодированной последовательностью плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC, или аминокислотной последовательности, показанной на фигуре 1 (SEQ ID No.2), или ее значительной части. В используемом здесь значении термин “значительная часть” аминокислотной последовательности, показанной на фигуре 1 (SEQ ID No.2), означает полипептид, содержащий по крайней мере около 70% аминокислотной последовательности, показанной на фигуре 1 (SEQ ID No.2), и представляющий собой вышеуказанный функционально эквивалентный продукт гена AveC. В используемом здесь значении применительно к аминокислотным последовательностям, гомологичным аминокислотной последовательности продукта гена AveC, продуцированного штаммом S. avermitilis, термин “гомологичный” относится к полипептиду, который имеет аминокислотную последовательность, показанную на фигуре 1 (SEQ ID No.2), в которой один или несколько аминокислотных остатков консервативно заменены другим аминокислотным остатком, при этом указанная аминокислотная последовательность по крайней мере примерно на 70%, более предпочтительно по крайней мере примерно на 80% и наиболее предпочтительно по крайней мере примерно на 90% идентична полипептиду, кодированному последовательностью плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC, или аминокислотной последовательности, показанной на фигуре 1 (SEQ ID No.2), при определении с помощью стандартного алгорифма идентичности аминокислотных последовательностей, такого как алгорифм BLASTP (GENBANK, NCBI), и, кроме того, такая консервативная замена позволяет получить вышеуказанный функционально эквивалентный генный продукт. Консервативные замены аминокислот хорошо известны в данной области. Правила выполнения таких замен описаны наряду с другими научными работами в публикации Dayhof, M.D., 1978, Nat. Biomed. Res. Found., Washington, D.C., Vol. 5, Sup. 3. В частности, консервативные замены аминокислот представляют собой такие замены, которые обычно происходят в семействе аминокислот, родственных в отношении кислотности или полярности. Генетически кодированные аминокислоты обычно делят на четыре группы: (1) кислые = аспартат, глутамат; (2) основные = лизин, аргинин, гистидин; (3) неполярные = аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженные полярные = глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин. Фенилаланин, триптофан и тирозин классифицированы также как ароматические аминокислоты. Одна или несколько замен в любой из указанных групп, например замена лейцина изолейцином или валином, аспартата глутаматом, треонина серином или любого другого аминокислотного остатка структурно родственным аминокислотным остатком, например аминокислотным остатком с одинаковой кислотностью или полярностью либо сходной комбинацией свойств, обычно оказывает незначительное воздействие на функцию полипептида. Методы получения и обработки молекул полинуклеотидов по данному изобретению известны в данной области и могут быть выполнены методами рекомбинантных ДНК, описанными, например, в публикациях Maniatis, et al., 1989, Molecular Cloning, A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; Ausubel, et al., 1989, Current Protocols In Molecular Biology, Greene Publishing Associates & Wiley Interscience, NY; Sambrook, et al., 1989, Molecular Cloning: A Laboratory Manual, 2d ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY; Innis et al. (eds), 1995, PCR Strategies, Academic Press, Inc., San Diego; and Erlich (ed), 1992, PCR Technology, Oxford University Press, New York, которые включены в данное описание изобретения в качестве ссылки. Клоны полинуклеотидов, кодирующие продукты гена AveC или продукты гомологичного гена AveC, можно идентифицировать любыми методами, известными в данной области, которые включают, не ограничиваясь ими, методы, рассмотренные в нижеследующем разделе 7. Библиотеки геномных ДНК можно исследовать с целью обнаружения последовательностей, кодирующих aveC и гомолог aveC, при помощи методов, описанных в публикации Benton and Davis, 1977, Science 196:180 для скрининга библиотек бактериофагов, и в публикации Grunstein and Hogness, 1975, Proc. Natl. Acad. Sci. USA, 72:3961-3965 для скрининга плазмидных библиотек. Молекулы полинуклеотидов, содержащие нуклеотидные последовательности, которые, как известно, включают открытую рамку считывания aveC, имеющуюся, например, в плазмиде pSE186 (АТСС 209604) или в плазмиде pSE119 (описанные в нижеследующем разделе 7), можно использовать в качестве зондов в указанных экспериментах по скринингу. Альтернативно, можно синтезировать олигонуклеотидные зонды, которые соответствуют нуклеотидным последовательностям, выделенным из неполных или полных аминокислотных последовательностей очищенного продукта гомологичного гена AveC. 5.2. Рекомбинантные системы 5.2.1. Клонирующие и экспрессирующие векторы Настоящее изобретение далее относится к рекомбинантным клонирующим векторам и экспрессирующим векторам, предназначенным для клонирования или экспрессии молекул полинуклеотидов по настоящему изобретению, которые включают, например, открытую рамку считывания aveC S. avermitilis или открытые рамки считывания любых гомологов aveC. В соответствии с неограничивающим вариантом осуществления настоящее изобретение относится к плазмиде pSE186 (АТСС 209604), которая содержит полную открытую рамку считывания гена aveC S. avermitilis. Все нижеследующее описание открытой рамки считывания aveC из S. avermitilis, молекулы полинуклеотида, содержащей открытую рамку считывания aveC из S. avermitilis или ее часть, или продукта гена AveC S. avermitilis относится также к мутированным аллелям aveC, рассматриваемым ниже, за исключением четко оговоренных случаев или случаев, следующих из контекста. Для конкретного использования в штамме Streptomyces создан целый ряд разных векторов, включающих фаг, мультикопийные плазмиды, малокопийные плазмиды и шаттл-векторы E. coli Streptomyces, и любые из указанных векторов могут быть использованы при осуществлении настоящего изобретения. Кроме того, в штамме Streptomyces был клонирован ряд генов лекарственной устойчивости, и некоторые из указанных генов были встроены в векторы в качестве селектируемых маркеров. Примеры существующих векторов, пригодных для использования в штамме Streptomyces, приведены в публикации Hutchinson, 1980, Applied Biochem. Biotech. 16:169-190. Рекомбинантные векторы по настоящему изобретению, в частности экспрессирующие векторы, предпочтительно сконструированы так, чтобы кодирующая последовательность молекулы полинуклеотида по данному изобретению была функционально связана с одним или несколькими регуляторными элементами, необходимыми для транскрипции и трансляции кодирующей последовательности для продуцирования полипептида. В используемом здесь значении термин “регуляторный элемент” означает, не ограничиваясь ими, нуклеотидные последовательности, кодирующие индуцируемые и неиндуцируемые промоторы, энхансеры, операторы и другие элементы, которые, как известно в данной области, служат для инициации и/или регулирования экспрессии кодирующих полинуклеотид последовательностей. Кроме того, в используемом здесь значении кодирующая последовательность “функционально связана” с одним или несколькими регуляторными элементами, которые эффективно регулируют и делают возможной транскрипцию кодирующей последовательности или трансляцию ее мРНК либо обеспечивают выполнение обеих функций. Типичные плазмидные векторы, которые могут содержать молекулу полинуклеотида по настоящему изобретению, включают наряду с прочими pCR-Blunt, pCR2.1 (Invitrogen), pGEM3Zf (Promega) и шаттл-вектор pWHM3 (Vara et al., 1989, J. Bact. 171:5872-5881). В данной области хорошо известны методы создания рекомбинантных векторов, содержащих определенные кодирующие последовательности, функционально связанные с соответствующими регуляторными элементами, и такие методы могут быть использованы при осуществлении настоящего изобретения. Указанные методы включают методы рекомбинантных ДНК in vitro, методы синтеза и методы генетической рекомбинации in vivo. См., например, методы, описанные в публикациях Maniatis et al., 1989, приведена выше; Ausubel et al., 1989, приведена выше; Sambrook et al., 1989, приведена выше; Innis et al., 1995, приведена выше, и Erlich, 1992, приведена выше. Регуляторные элементы указанных векторов могут характеризоваться разной силой и специфичностью. В зависимости от применяемой системы хозяин/вектор можно использовать любые элементы, пригодные для транскрипции и трансляции. Неограничивающие примеры областей регуляции транскрипции или промоторов для бактерий включают промотор Векторы экспрессии слитого белка можно использовать для экспрессии слитого белка продукта гена AveC. Очищенный слитый белок можно использовать для получения антисыворотки против продукта гена AveC, исследования биохимических свойств продукта гена AveC, создания слитых белков AveC, обладающих разной биохимической активностью, или облегчения идентификации или очистки экспрессированного продукта гена AveC. Возможные векторы экспрессии слитого белка включают, не ограничиваясь ими, векторы, содержащие последовательности, кодирующие продукты слияния Слитые белки AveC могут содержать область, пригодную для очистки. Например, продукты слияния AveC, мальтозы и связывающего белка могут быть очищены при помощи амилозной смолы; белки, полученные в результате слияния AveC, глутатиона и S-трансферазы, могут быть очищены при помощи глутатион-агарозных шариков, и продукты слияния AveC и полигистидина могут быть очищены при помощи смолы, содержащей двухвалентный никель. Альтернативно для очистки слитого белка при помощи аффинной хроматографии можно использовать антитела против белка-носителя или пептида. Например, нуклеотидная последовательность, кодирующая эпитоп-мишень моноклонального антитела, может быть функционально связана в экспрессирующем векторе с регуляторными элементами и расположена с возможностью слияния экспрессированного эпитопа с полипептидом гена AveC. Например, нуклеотидная последовательность, кодирующая меченый эпитоп FLAG Экспрессирующий вектор, кодирующий слитый белок AveC, может также содержать полилинкерные последовательности, кодирующие сайты расщепления специфической протеазой, благодаря чему экспрессированный полипептид AveC может быть выделен из несущей области или продукта слияния в результате обработки специфической протеазой. Например, вектор слитого белка может содержать последовательности ДНК, кодирующие сайты расщепления тромбином или Ха-фактором. При помощи известных методов в экспрессирующий вектор может быть встроена сигнальная последовательность, расположенная в рамке считывания вверху от открытой рамки считывания aveC, которая направляет миграцию и секрецию экспрессированного генного продукта. Неограничивающие примеры сигнальных последовательностей включают последовательности, выделенные из Для облегчения выбора клеток-хозяев, трансформированных или трансфецированных клонирующими или экспрессирующими векторами по настоящему изобретению, указанный вектор может далее содержать кодирующую последовательность для продукта гена-репортера или другого селектируемого маркера. Такая кодирующая последовательность предпочтительно функционально связана с вышеописанными последовательностями, кодирующими регуляторные элементы. Гены-репортеры, пригодные для использования в данном изобретении, хорошо известны в данной области и включают гены-репортеры, кодирующие зеленый флуоресцирующий белок, люциферазу, xyIE и тирозиназу. Нуклеотидные последовательности, кодирующие селектируемые маркеры, хорошо известны в данной области и включают последовательности, которые кодируют генные продукты, сообщающие устойчивость к антибиотикам или антиметаболитам, или удовлетворяют требованию ауксотрофности. Примеры таких последовательностей включают последовательности, кодирующие устойчивость к эритромицину, тиострептону или канамицину наряду с прочими антибиотиками. 5.2.2. Трансформация клеток-хозяев Настоящее изобретение далее относится к трансформированным клеткам-хозяевам, содержащим молекулу полинуклеотида или рекомбинантный вектор по данному изобретению, и выделенным из них новым штаммам или линиям клеток. Клетки-хозяева, пригодные для осуществления данного изобретения, предпочтительно являются клетками Streptomyces, хотя можно также использовать другие прокариотические или эукариотические клетки. Такие трансформированные клетки-хозяева обычно включают, не ограничиваясь ими, микроорганизмы, такие как бактерии, трансформированные рекомбинантными векторами ДНК бактериофага, плазмидной ДНК или космидной ДНК, или дрожжи, трансформированные рекомбинантными векторами наряду с прочими векторами. Молекулы полинуклеотидов по настоящему изобретению предназначены для функционирования в клетках Strepromyces, но могут быть также введены в другие бактериальные или эукариотические клетки, например, с целью клонирования или экспрессии. Обычно используют штамм E. coli, такой как, например, штамм DH5 Рекомбинантный эксперссирующий вектор по данному изобретению предпочтительно вводят, например, путем трансформации или трансфекции в одну или несколько клеток-хозяев по существу гомогенной культуры клеток. Экспрессирующий вектор обычно вводят в клетки-хозяева известными методами, такими как, например, трансформация протопластов, осаждение фосфатом кальция, обработка хлоридом кальция, микроинъекция, электропорация, трансфекция при контактировании с рекомбинированным вирусом, трансфекция, опосредуемая липосомами, трансфекция DEAE-декстраном, трансдукция, конъюгирование или бомбардировка микрочастицами. Трансформанты можно выбрать стандартными методами, например, выбирая клетки, экспрессирующие вышеописанный селектируемый маркер, такой как устойчивость к антибиотикам, связанный с рекомбинантным вектором. После введения экспрессирующего вектора в клетку-хозяина интеграцию и сохранение последовательности, кодирующей aveC, в хромосоме клетки-хозяина или эписоме можно проверить стандартными методами, например при помощи анализа гибридизации методом саузерн-блоттинга, анализа рестрикционного фермента, анализа PCR, включая PCR с использованием обратной транскриптазы (rt-PCR) или иммунологического анализа, предназначенного для обнаружения предполагаемого генного продукта. Клетки-хозяева, содержащие и/или экспрессирующие последовательность, кодирующую рекомбинантный aveC, можно идентифицировать любым по крайней мере из четырех общих методов, хорошо известных в данной области, которые включают: (i) гибридизацию ДНК-ДНК, ДНК-РНК или РНК-антисмысловая РНК; (ii) обнаружение наличия функций гена-“маркера”; (iii) оценку уровня транскрипции путем измерения экспрессии aveC-специфических транскриптов мРНК в клетке-хозяине; и (iv) обнаружение присутствия зрелого полипептида, определяемого, например, при помощи иммуноанализа или по наличию биологической активности AveC (например, продуцирование в клетках-хозяевах S. avermitilis авермектинов, являющихся показателем активности AveC, в соответствующих соотношениях и количествах). 5.2.3. Экспрессия и исследование продукта рекомбинантного гена AveC После того как последовательность, кодирующая нативный или мутированный aveC, устойчиво введена в соответствующую клетку-хозяин, трансформированную клетку-хозяина размножают клонированием и образовавшиеся клетки выращивают в условиях, обеспечивающих максимальное продуцирование продукта нативного или мутированного гена AveC. Такие условия обычно включают выращивание клеток до высокой плотности. Если экспрессирующий вектор содержит индуцируемый промотор, создают условия, необходимые для индукции экспрессии, которые включают, например, изменение температуры, удаление питательных веществ, добавление приемлемых индукторов (например, аналогов углеводородов, таких как изопропил- Если продукт экспрессированного гена AveC остается внутри клеток-хозяев, клетки собирают и лизируют, продукт выделяют и очищают от лизата в условиях экстракции, известных в данной области, которые позволяют свести до минимума разрушение белка, например, при 4°С или в присутствии ингибиторов протеазы либо при создании обоих условиях. Если продукт экспрессированного гена AveC секретируется из клеток-хозяев, можно просто собрать истощенную питательную среду и выделить из нее продукт. Продукт экспрессированного гена AveC можно выделить или очистить от клеточных лизатов или культуральной среды при помощи стандартных методов, которые включают, не ограничиваясь ими, любую комбинацию нижеследующих методов: осаждение сульфатом аммония, вытеснительное фракционирование, ионообменную хроматографию, ВЭЖХ, центрифугирование в градиенте плотности и аффинную хроматографию. Если продукт экспрессированного гена AveC обладает биологической активностью, более высокую степень чистоты препарата можно контролировать на каждой стадии очистки при помощи соответствующего анализа. Независимо от того, обладает продукт экспрессированного гена AveC биологической активностью или нет, его можно обнаружить на основании величины выхода, способности взаимодействовать с антителом, специфическим для AveC, или наличия метки слияния. В используемом здесь значении продукт гена AveC считается “по существу очищенным”, если данный продукт составляет примерно более 20 мас.% от содержания белка в конкретном препарате. Кроме того, в используемом здесь значении продукт гена AveC считается “выделенным”, если данный продукт составляет по крайней мере около 80 мас.% от содержания белка в конкретном препарате. Таким образом, настоящее изобретение относится к выделенному или по существу очищенному продукту рекомбинантно экспрессированного гена AveC S. avermitilis, содержащему аминокислотную последовательность, кодированную последовательностью плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC, или аминокислотную последовательность, показанную на фигуре 1 (SEQ ID No.2), или ее значительную часть, а также их мутированные и вырожденные варианты. Настоящее изобретение далее относится к способу получения про-дукта гена AveC, который включает культивирование клетки-хозяина, трансформированной рекомбинантным экспрессирующим вектором, содержащим молекулу полинуклеотида, имеющую нуклеотидную последовательность, кодирующую продукт гена AveC, причем молекула полинуклеотида функционально связана с одним или несколькими регуляторными элементами, контролирующими экспрессию молекулы полинуклеотида в клетке-хозяине в условиях, необходимых для продуцирования продукта рекомбинантного гена AveC, и выделение продукта гена AveC из культуры клеток. Продукт рекомбинантно экспрессированного гена AveC S. avermitilis пригоден для разных целей, включая скрининг соединений, изменяющих функцию продукта гена AveC и модулирующих таким образом биосинтез авермектина, и получение антител против продукта гена AveC. После получения продукта гена AveC достаточной чистоты его можно исследовать стандартными методами, включающими SDS-PAGE, вытеснительную хроматографию, анализ аминокислотной последовательности, анализ биологической активностью при продуцировании соответствующих продуктов в процессе биосинтеза авермектина и т.д. Например, аминокислотную последовательность продукта гена AveC можно определить стандартными методами секвенирования пептида. Продукт гена AveC можно далее исследовать при помощи анализа гидрофильности (см., например, Hopp and Woods, 1981, Proc. Natl. Acad. Sci. USA 78:3824) или аналогичных программных алгорифмов для идентификации гидрофобных и гидрофильных областей в продукте гена AveC. Можно выполнить структурный анализ для идентификации областей продукта гена AveC, которые предполагают наличие специфических вторичных структур. Биофизические методы, такие как рентгеновская кристаллография (Engstrom, 1974, Biochem. Exp. Biol. 11:7-13), компьютерное моделирование (Fletterick and Zoller (eds.), 1986, in: Current Communications in Molecular Biology. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY) и ядерный магнитный резонанс (ЯМР), можно использовать для картирования и исследования сайтов взаимодействия между продуктом гена AveC и его субстратом. Информацию, полученную в результате выполнения вышеуказанных исследований, можно использовать для выбора новых сайтов для мутации в открытой рамке считывания aveC, получения новых штаммов S. avermitilis, обладающих более привлекательными характеристиками продуцирования авермектина. 5.3. Создание и использование мутантов AveC Главным объектом настоящего изобретения является идентификация новых мутаций в аллеле aveC S. avermitilis, которые вызывают изменение и наиболее предпочтительно уменьшение соотношения авермектинов В2:В1. Таким образом, настоящее изобретение относится к молекулам полинуклеотидов, пригодным для получения новых штаммов клеток S. avermitilis, которые характеризуются поддающимся обнаружению изменением продуцирования авермектина по сравнению с клетками такого же штамма, экспрессирующими только аллель aveC дикого типа. В предпочтительном варианте осуществления изобретения такие молекулы полинуклеотидов пригодны для получения новых штаммов клеток S. avermitilis, которые продуцируют авермектины класса 2:1 в меньшем соотношении по сравнению с клетками такого же штамма, экспрессирующими только аллель aveC дикого типа. Клетки таких штаммов могут также содержать дополнительные мутации, способствующие продуцированию большего количества авермектинов по сравнению с клетками такого же штамма, экспрессирующими только аллель aveC дикого типа. Мутации аллеля aveC или кодирующей последовательности включают любые мутации, которые вызывают замену, делецию и/или добавление одной или нескольких аминокислот в продукт гена AveC или укорачивание продукта гена AveC, либо любые комбинации указанных мутаций, а также мутации, позволяющие достичь требуемого результата. Предполагается, что такие последовательности мутированного аллеля aveC включают любые вырожденные варианты. Например, настоящее изобретение относится к молекуле полинуклеотида, содержащей нуклеотидную последовательность аллеля aveC или ее вырожденный вариант, последовательность плазмиды pSE186 (АТСС 209604), кодирующую продукт гена AveC, или ее вырожденный вариант или нуклеотидную последовательность открытой рамки считывания aveC S. avermitilis, показанную на фигуре 1 (SEQ ID No.1), или ее вырожденный вариант, которая далее включает мутации, кодирующие комбинацию замен аминокислот в выбранных положениях структуры продукта гена AveC. В неограничивающем варианте осуществления изобретения такие замены происходят в положении одной или нескольких аминокислот продукта гена AveC, соответствующих положениям аминокислот 25, 28, 35, 36, 38, 40, 41, 48, 55, 61, 78, 84, 89, 90, 99, 107, 108, 111, 120, 123, 136, 138, 139, 141, 154, 159, 163, 179, 192, 196, 198, 200, 202, 220, 228, 229, 230, 231, 234, 238, 239, 250, 252, 266, 275, 278, 289 или 298 SEQ ID No.2. Предпочтительные комбинации положений аминокислот, в которых происходит замена, включают один или несколько аминокислотных остатков D48, A61, A89, L136, S138, A139, R163, G179, V196, A198, E238 и Р289. Особенно предпочтительные комбинации замен аминокислот включают замены в положениях D48 и G179, более конкретно D48E и G179S. Конкретные примеры комбинаций замен аминокислот, вызывающих уменьшение соотношения авермектинов циклогексил В2 : циклогексил В1, показаны на фигуре 6А-J. Таким образом, настоящее изобретение относится к молекуле полинуклеотида, содержащей нуклеотидную последовательность, которая по существу аналогична аллелю aveC Streptomyces avermitilis, последовательности плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC S. avermitilis, нуклеотидной последовательности открытой рамки считывания aveC S. avermitilis, показанной на фигуре 1 (SEQ ID No.1), или их вырожденному варианту, но при этом имеет мутации, кодирующие комбинацию замен аминокислот в положениях аминокислотных остатков, соответствующих положениям аминокислот SEQ ID No.2, благодаря чему клетки штамма S. avermitilis АТСС 53692, в которых инактивирован аллель aveC дикого типа и которые экспрессируют молекулу полинуклеотида, содержащую мутированную нуклеотидную последовательность, способны продуцировать авермектины класса 2:1 в меньшем соотношении по сравнению с соотношением, продуцируемым клетками штамма S. avermitilis АТСС 53692, экспрессирующими только аллель aveC дикого типа, причем когда авермектины класса 2:1 являются авермектинами циклогексил В2 : циклогексил В1, соотношение авермектинов класса 2:1 составляет 0,35:1 или меньше. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,30:1 или меньше. В еще более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,25:1 или меньше. В наиболее предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,20:1 или меньше. В конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (а): D48E, A61T, A89T, S138T, A139T, G179S, A198G, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE538 (см. фигуру 6). В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (b): G40S, D48E, L136P, G179S, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE559. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (с): D48E, L136P, R163Q, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE567. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (d): D48E, L136P, R163Q, G179S, E238D. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE570 и pSE572. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (е): D48E, L136P, R163Q, G179S, A200G, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE571. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (f): D48E, L136P, G179S, E238D. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE501 и pSE546. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (g): D48E, A61T, L136P, G179S, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE510. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (h): D48E, A61T, L136P, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE512. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (i): D48E, A89T, S138T, A139T, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE519. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (j): D48E, A61T, L136P, G179S, A198G, P202S, E238D, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE526. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (k): D48E, A61T, L136P, S138T, A139F, G179S, E238D, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE528. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (l): D48E, L136P, G179S, A198G, E238D, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE530. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (m): D48E, A61T, S138T, A139F, G179S, A198G, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE531. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (n): D48E, L84P, G111V, S138T, A139T, G179S, A198G, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE534. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (o): Y28C, D48E, A61T, A89T, A138T, A139T, G179S, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE535. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (p): D48E, A61T, A107T, S108G, L136P, G179S, S192A, E238D, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE542. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (q): D48E, L136P, G179S, R250W. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE545. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (r): D48E, A89T, S138T, A139T, R163Q, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE548. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (s): D48E, L136P, G179S, A198G, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE552. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (t): D48E, F78L, A89T, L136P, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE557. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (u): D48E, A89T, S138T, A139T, G179S, E238D, F278L. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE564 и pSE565. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (v): D48E, A89T, L136P, R163Q, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE568. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (w): D48E, A61T, A89T, G111V, S138T, A139F, G179S, E238D, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE543. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (x): D25G, D48E, A89T, L136P, S138T, A139T, V141A, I159T, R163Q, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE504. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (y): D48E, A89T, S90G, L136P, R163Q, G179S, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE508. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (z): D48E, A61T, A89T, G111V, S138T, A139T, G179S, E238D, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE511. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (aa): D48E, A89T, S138T, A139T, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE520. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ab): D48E, L136P, R163Q, G179S, S231L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE523. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ac): D48E, L136P, S138T, A139F, G179S, V196A, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE527. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ad): D48E, A61T, A89T, F99S, S138T, A139T, G179S, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE539. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ae): G35S, D48E, A89T, S138T, A139T, G179S, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE540. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (af): D48E, A61T, A89T, S138T, A139T, G179S, V196A, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE547. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ag): D48E, A89T, G111V, S138T, A139T, G179S, A198G, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE550. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ah): S41G, D48E, A89T, L136P, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE558. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ai): D48E, A89T, L136P, R163Q, G179S, P252S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE563. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (aj): D48E, A89T, L136P, G179S, F234S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE566. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ak): D48E, A89T, L136P, R163Q, G179S, E238D. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE573 и pSE578. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (al): Q36R, D48E, A89T, L136P, G179S, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE574. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (am): D48E, A89T, L136P, R163Q, G179S. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE575 и pSE576. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (an): D48E, A89T, S138T, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE577. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ao): D48E, A89T, L136P, G179S, E238D. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE502 и pSE524. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ap): D48E, A89T, L136P, K154E, G179S, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE503. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (aq): D48E, A89T, S138T, A139T, K154R, G179S, V196A, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE505. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ar): D48E, A89T, S138T, A139F, G179S, V196A, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE506. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (as): D48E, A61T, A89T, L136P, G179S, V196A, A198G, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE507. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (at): D48E, A61T, S138T, A139F, G179S, G196A, E238D, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE509. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (au): D48E, A89T, L136P, G179S. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE514 и pSE525. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (av): D48E, A89T, V120A, L136P, G179S. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE515. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (aw): D48E, A61T, A89T, S138T, A139F, G179S, V196A, A198G, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE517. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ax): D48E, A61T, A89T, G111V, S138T, A139F, G179S, V196A, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE518. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ay): D48E, A61T, A89T, S138T, A139T, G179S, V196A, E238D, P289L. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE529 и pSE554. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (az): D48E, A61T, A89T, L136P, S138T, A139F, G179S, A198G, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE532. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (ba): D48E, A89T, S138T, A139F, G179S, A198G, V220A. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE536. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (bb): D48E, A61T, A89T, S138T, A139T, G179S, V196A, E238D, R239H, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE537. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (bc): D48E, A61T, A89T, L136P, G179S, P289L. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE541. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (bd): D48E, A89T, S138T, A139T, G179S, V196A, E238D, P289L. Неограничивающими примерами плазмид, кодирующих указанные замены аминокислот, являются плазмиды pSE549 и pSE553. В другом конкретном варианте осуществления изобретения комбинация замен аминокислот включает комбинацию группы (be): D48E, A61T, A89T, S138T, A139F, G179S, V196A, E238D. Неограничивающим примером плазмиды, кодирующей указанные замены аминокислот, является плазмида pSE551. Настоящее изобретение далее относится к молекуле полинуклеотида, содержащей нуклеотидную последовательность, которая по существу аналогична аллелю aveC Strepromyces avermitilis, последовательности плазмиды pSЕ186 (АТСС 209604), кодирующей продукт гена AveC S. avermitilis, нуклеотидной последовательности открытой рамки считывания aveC S. avermitilis, показанной на фигуре 1 (SEQ ID No.1), или их вырожденному варианту, но при этом имеет мутации, кодирующие комбинацию замен аминокислот в положении аминокислотных остатков, соответствующих положениям аминокислот SEQ ID No.2, благодаря чему клетки штамма S. avermitilis АТСС 53692, в которых инактивирован аллель aveC дикого типа и которые экспрессируют молекулу полинуклеотида, содержащую мутированную нуклеотидную последовательность, способны продуцировать авермектины класса 2:1 с меньшим соотношением по сравнению с соотношением, продуцируемым клетками штамма S. avermitilis АТСС 53692, экспрессирующими только аллель aveC дикого типа, причем когда авермектины класса 2:1 являются авермектинами циклогексил В2 : циклогексил В1, соотношение авермектинов класса 2:1 уменьшается примерно до 0,40:1 или меньше, и комбинация замен аминокислот включает комбинацию, выбираемую из нижеследующей группы: (bf) D48E, S138T, A139T, G179S, E238D; и (bg) Y28C, Q38R, D48E, L136P, G179S, E238D. Неограничивающими примерами плазмиды, кодирующей замены аминокислот группы (bf), являются pSE556 и pSE569. Неограничивающим примером плазмиды, кодирующей замены аминокислот группы (bg), является pSE561. В соответствии с настоящим изобретением любые вышеуказанные замены аминокислот могут быть выполнены в результате любой модификации нуклеотидной последовательности аллеля aveC или ее вырожденного варианта, которая вызывает такие замены. Например, большинство вышеописанных замен аминокислот можно произвести, заменяя последовательность нативного кодона или ее вырожденный вариант любым альтернативным кодоном, кодирующим такую же замену аминокислот. Все возможные последовательности, которые могут кодировать вышеуказанные замены аминокислот, должны быть очевидны специалисту в данной области на основании данного описания изобретения и известной вырожденности генетического кода. В неограничивающем варианте осуществления изобретения любая вышеуказанная комбинация замен аминокислот включает немолчащие замены нуклеотидов, показанные на фигуре 6. В используемом здесь значении фраза “комбинация замен аминокислот включает комбинацию группы…” и подобные фразы означают, что замены аминокислот в продукте гена AveC по настоящему изобретению включают по крайней мере специально указанные замены и могут включать другие замены, делеции, добавления аминокислот или их комбинации, благодаря чему экспрессия продукта гена AveC в клетке S. avermitilis характеризуется требуемым уменьшением соотношения авермектинов В2:В1. Мутации в аллеле aveC или его вырожденном варианте могут быть выполнены целым рядом известных методов, таких как PCR с возможностью ошибок или кластерный мутагенез. В частности, олигонуклеотид-направленный мутагенез можно использовать для целевого изменения последовательности аллеля aveC или открытой рамки считывания, например, для введения одного или нескольких сайтов рестрикции или терминирующего кодона в специфические области аллеля aveC или открытой рамки считывания. Методы, аналогичные описанным в патентах США №№ 5605793, 5830721 и 5837458, которые включают неспецифическую фрагментацию, повторные циклы мутагенеза и перестановку нуклеотидов, можно также использовать для создания крупных библиотек полинуклеотидов, содержащих нуклеотидные последовательности, кодирующие мутации aveC. Могут быть выполнены направленные мутации, особенно в тех случаях, когда они позволяют изменить один или несколько консервативных аминокислотных остатков в продукте гена AveC. Например, сравнение выделенной аминокислотной последовательности продукта гена AveC S. avermitilis (SEQ ID No.2) с продуктами гомологичного гена AveC, продуцированными S. griseochromogenes (SEQ ID No.5) и S. hygroscopicus (SEQ ID No.4), описанными в патенте США № 6248579, позволяет выявить сайты наиболее консервативных аминокислотных остатков в двух видах. Направленный мутагенез, вызывающий замену одного или нескольких указанных консервативных аминокислотных остатков, может быть весьма эффективным для получения новых мутантных штаммов, продуцирующих авермектин с требуемыми изменениями. Кроме того, хорошие результаты могут быть получены при выполнении неспецифического мутагенеза в клетках S. avermitilis под воздействием ультрафиолетового или рентгеновского излучения либо химических мутагенов, таких как N-метил-N’-нитрозогуанидин, этилметансульфонат, азотистая кислота или горчичный газ. См., например, приведенную выше публикацию Ausubel, 1989, для ознакомления с методами мутагенеза. Полученные мутированные молекулы полинуклеотидов исследуют для определения их способности модулировать биосинтез авермектинов в S. avermitilis. В предпочтительном варианте осуществления изобретения молекулу полинуклеотида, содержащую мутированную нуклеотидную последовательность, испытывают методом комплементации штамма S. avermitilis, в котором был инактивирован ген aveC, с целью создания отрицательного фона для aveC (aveC’). При осуществлении способа, не ограничивающего объем изобретения, мутированную молекулу полинуклеотида сплайсируют с получением экспрессирующей плазмиды, функционально связанной с одним или несколькими регуляторными элементами, которая также предпочтительно содержит один или несколько генов лекарственной устойчивости, позволяющих выбрать трансформированные клетки. Указанный вектор затем вводят в клетки-хозяева с aveC’ при помощи известных методов, после чего трансформированные клетки отбирают и культивируют в соответствующих ферментационных средах в условиях, стимулирующих или индуцирующих продуцирование авермектина, например, в результате введения в среду соответствующих инициирующих субъединиц и культивирования в условиях, оптимальными для продуцирования авермектина. Продукты ферментации анализируют ВЭЖХ для определения способности мутированной молекулы полинуклеотида дополнять клетку-хозяина. Несколько плазмидных векторов, несущих мутированные молекулы полинуклеотидов, способные уменьшать соотношение авермектинов В2:В1, в том числе pSE188, pSE199, рSE231, pSE239 и pSE290-pSE297, рассмотрены в нижеследующем разделе 8.3. Другие примеры таких плазмидных векторов приведены на фигуре 6. Любые вышеуказанные способы по настоящему изобретению можно осуществлять, используя ферментационные культуральные среды, предпочтительно содержащие циклогексанкарбоновую кислоту, хотя можно также использовать другие приемлемые предшественники жирных кислот, в частности, предшественники жирных кислот, приведенные в таблице 1, или метилтиомолочную кислоту. После идентификации мутированной молекулы полинуклеотида, модулирующей продуцирование авермектина в требуемом направлении, можно определить расположение мутации в нуклеотидной последовательности. Например, молекулу полинуклеотида, содержащую нуклеотидную последовательность, кодирующую продукт мутированного гена AveC, можно выделить методом PCR или произвести анализ последовательности ДНК известными методами. Сравнивая последовательность ДНК мутированного аллеля aveC с последовательностью аллеля aveC дикого типа, можно определить мутации, ответственные за изменение продуцирования авермектина. Например, у продуктов гена AveC S. avermitilis, содержащих одинарные замены аминокислот в положении любых остатков 55 (S55F), 138 (S138T), 139 (A139T) или 230 (G230D) или двойные замены в положениях 138 (S138T) и 139 (А139Т или A139F), произошло изменение функции продукта гена AveC, в результате чего изменилось соотношение авермектинов класса 2:1 (см. нижеследующий раздел 8); указанные положения аминокислот соответствуют показанным на фигуре 1 (SEQ ID No.2). Кроме того, было установлено, что семь нижеследующих комбинаций мутаций эффективно уменьшают соотношение авермектинов класса 2:1: (1) D48E/A89T; (2) S138T/A139T/G179S; (3) Q38P/L136P/E238D; (4) F99S/S138T/A139T/G179S; (5) A139T/M228T; (6) G111V/P289L; (7) A139T/K154E/Q298H. В объем настоящего изобретения входят пятьдесят девять (59) дополнительных комбинаций мутаций, которые, как установлено, уменьшают соотношение авермектинов циклогексил В2 : циклогексил В1; указанные мутации показаны на фигуре 6 и приведены в прилагаемой формуле изобретения. Используемые в данном описании изобретения вышеуказанные обозначения, такие как А139Т, содержат однобуквенное название аминокислотного остатка, которым в данном примере является аланин (А), в указанном положении, которым в данном примере является положение 139 (см. SEQ ID No.2) полипептида, с последующим указанием аминокислотного остатка, заменяющего исходный аминокислотный остаток, которым в данном примере является треонин (Т). Когда аминокислотный остаток, кодированный аллелем aveC в хромосоме S. avermitilis, векторе или выделенной молекуле полинуклеотида по настоящему изобретению, “соответствует” определенному аминокислотному остатку SEQ ID No.2 или когда замена аминокислоты происходит в определенном положении, “соответствующем” положению конкретного аминокислотного остатка SEQ ID No.2, предполагается, что такой аминокислотный остаток находится в таком же относительном положении в продукте гена AveC, которое специалист в данной области может легко определить, обратившись к аминокислотной последовательности, обозначенной в данном описании изобретении как SEQ ID No.2. Когда конкретные мутации в аллеле aveC, кодирующем определенные мутации, представляют собой замены оснований в положениях конкретных нуклеотидов в аллеле aveC, “соответствующих” положениям определенных нуклеотидов, показанным в SEQ ID No.1, или когда положение нуклеотида в аллеле aveC “соответствует” положению определенного нуклеотида в SEQ ID No.1, предполагается, что указанный нуклеотид находится в таком же относительном положении в нуклеотидной последовательности aveC или ее вырожденном варианте, которое специалист в данной области может легко определить, обратившись к нуклеотидной последовательности, обозначенной в данном описании изобретения как SEQ ID No.1. В определении соотношений авермектинов циклогексил В2 : циклогексил В1 термин “примерно” определяет данное числовое значение с возможным отклонением плюс или минус 10% от указанного значения. Молекула полинуклеотида по настоящему изобретению может быть определена термином “выделенная”, который означает, что данная молекула (i) очищена в такой степени, что она по существу не загрязнена другими молекулами полинуклеотидов, содержащими другие нуклеотидные последовательности, (ii) находится в окружении, в котором она не встречается в естественных условиях, например, когда аллель aveC, выделенный из S. avermitilis, или его мутированный вариант присутствует в клетке, не являющейся клеткой S. avermitilis, или (iii) имеет форму, не присущую ей в естественных условиях, например, представляет собой более короткий фрагмент ДНК, такой как рестрикционный фрагмент, выделенный из бактериальной хромосомы и содержащий в основном aveC-кодирующую область или ее мутированный вариант с какими-либо связанными регуляторными последовательностями или без них; встроена в гетерологичный фрагмент ДНК, такой как хромосома бактериальной клетки (не являющейся клеткой S. avermitilis), или в ДНК вектора, такого как плазмида или фаг, или встроена в хромосому S. avermitilis в локусе, не являющемся локусом нативного аллеля aveC. Настоящее изобретение далее относится к рекомбинантному вектору, содержащему молекулу полинуклеотида по настоящему изобретению. Такой рекомбинантный вектор можно использовать для направленной доставки любых молекул полинуклеотидов, содержащих мутированные нуклеотидные последовательности по настоящему изобретению, к сайту аллеля aveC хромосомы S. avermitilis для вставки в открытую рамку считывания aveC или замены указанной рамки считывания или ее части, например, путем гомологичной рекомбинации. Однако молекула полинуклеотида, содержащая мутированную нуклеотидную последовательность по настоящему изобретению, может также модулировать биосинтез авермектина при введении в хромосому S avermitilis на сайте, не относящемся к аллелю aveC, или при нахождении в эписоме клеток S. avermitilis. Таким образом, настоящее изобретение далее относится к векторам, содержащим молекулу полинуклеотида, имеющую мутированную нуклеотидную последовательность по настоящему изобретению, которые можно использовать для вставки молекулы полинуклеотида на сайт в хромосоме S. avermitilis, не относящийся к гену aveC, или для сохранения в эписоме. В неограничивающем варианте осуществления изобретения вектор является вектором замещения гена, который можно использовать для введения мутированного аллеля aveC или его вырожденного варианта по настоящему изобретению в клетки штамма S. avermitilis с образованием новых штаммов S. avermitilis, клетки которых могут продуцировать авермектины класса 2:1 в меньшем соотношении по сравнению с клетками такого же штамма, которые экспрессируют только аллель aveC дикого типа. Такие векторы замещения гена могут быть созданы с использованием мутированных молекул полинуклеотидов, присутствующих в экспрессирующих векторах, рассмотренных в данном описании изобретения, таких как экспрессирующие векторы, рассмотренные в нижеследующем разделе 8. Настоящее изобретение далее относится к векторам, которые можно использовать для введения мутированного аллеля aveC или его вырожденного варианта в клетки штамма S. avernmitilis с целью создания новых штаммов клеток, продуцирующих авермектины в измененных количествах по сравнению с клетками такого же штамма, которые экспрессируют только аллель aveC дикого типа. В предпочтительном варианте осуществления изобретения увеличивается количество авермектинов, продуцируемых клетками. В конкретном варианте осуществления изобретения, не ограничивающем объем изобретения, такой вектор содержит сильный промотор, известный в данной области, например, сильный структурный промотор ermE, выделенный из Saccharopolyspora erythraea, который расположен вверху от открытой рамки считывания aveC и функционально связан с указанной рамкой. Такие векторы можно создать способами, описанными в патенте США № 6248579, используя мутированный аллель aveC плазмиды pSE189. Настоящее изобретение относится к векторам замещения гена, которые пригодны для инактивации гена aveC в штамме S. avermitilis дикого типа. В неограничивающем варианте осуществления изобретения такие векторы замещения гена могут быть созданы с использованием мутированной молекулы полинуклеотида, находящейся в плазмиде pSE180 (АТСС 209605), которая рассмотрена в нижеследующем разделе 8.1 (фигура 3). Настоящее изобретение далее относится к векторам замещения гена, которые содержат молекулу полинуклеотида, имеющую нуклеотидные последовательности, которые в естественном состоянии фланкируют ген aveC in situ в хромосоме S. avermitilis, включая последовательности, фланкирующие нуклеотидные последовательности, показанные на фигуре 1 (SEQ ID No.1), причем указанные векторы можно использовать для удаления открытой рамки считывания aveC S. avermitilis. Настоящее изобретение далее относится к клетке-хозяину, содержащей молекулу полинуклеотида или рекомбинантный вектор по настоящему изобретению. Клетка-хозяин может быть любой прокариотической или эукариотической клеткой, которую можно использовать в качестве хозяина для молекулы полинуклеотида или рекомбинантного вектора. В предпочтительном варианте осуществления изобретения клетка-хозяин является бактериальной клеткой. В более предпочтительном варианте осуществления изобретения клетка-хозяин является клеткой Streptomyces. В наиболее предпочтительном варианте осуществления изобретения клетка-хозяин является клеткой Streptomyces avermitilis. Настоящее изобретение далее относится к способу получения нового штамма Streptomyces avermitilis, который включает (i) мутирование аллеля aveC в клетке штамма S. avermitilis, вызывающее комбинацию замен аминокислот в продукте гена AveC, или (ii) введение в клетку штамма S. avermitilis мутированного аллеля aveC или его вырожденного варианта, кодирующего продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. Настоящее изобретение далее относится к способу получения нового штамма S. avermitilis, который включает (i) мутирование аллеля aveC в клетке штамма S. avermitilis, вызывающее комбинацию замен аминокислот в продукте гена AveC, или (ii) введение в клетку штамма S. avermitilis мутированного аллеля aveC или его вырожденного варианта, кодирующего продукт гена AveC, включающий комбинацию замен аминокислот, благодаря чему клетки, содержащие мутированный аллель aveC или его вырожденный вариант, могут продуцировать авермектины циклогексил В2 : циклогексил В1 в соотношении 0,35:1 или меньше. В неограничивающем варианте осуществления изобретения мутированный аллель aveC или его вырожденный вариант кодирует продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. В предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,30:1 или меньше. В неограничивающем варианте осуществления изобретения мутированный аллель aveC или его вырожденный вариант кодирует продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (f)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,25:1 или меньше. В неограничивающем варианте осуществления изобретения мутированный аллель aveC или его вырожденный вариант кодирует продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (w)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,20:1 или меньше. В неограничивающем варианте осуществления изобретения мутированный аллель aveC или его вырожденный вариант кодирует продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (ао)-(be) приведенного выше списка. Настоящее изобретение далее относится к способу получения нового штамма Streptomyces avermitilis, который включает (i) мутирование аллеля aveC в клетке штамма S. avermitilis, вызывающее комбинацию замен аминокислот в продукте гена AveC, или (ii) введение в клетку штамма S. avermitilis мутированного аллеля aveC или его вырожденного варианта, кодирующего продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (bf) и (bg). В предпочтительном варианте осуществления изобретения клетки S. avermitilis, содержащие такой мутированный аллель aveC или его вырожденный вариант, могут продуцировать авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,40:1 или меньше. В результате мутирования аллеля aveC в клетке или введения в клетку мутированного аллеля aveC или его вырожденного варианта в соответствии с вышеуказанными пунктами получают новый штамм S. avermitilis. Настоящее изобретение далее относится к клетке вида из рода Streptomyces, которая содержит мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из пунктов (а)-(be) приведенного выше списка. В предпочтительном варианте осуществления изобретения вид из рода Streptomyces является видом из рода S. avermitilis. Настоящее изобретение далее относится к клетке S. avermitilis, способной продуцировать авермектины циклогексил В2 : циклогексил В1 в соотношении 0,35:1 или меньше. В неограничивающем варианте осуществления изобретения указанная клетка содержит мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. В предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,30:1 или меньше. В неограничивающем варианте осуществления изобретения указанная клетка содержит мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (f)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,25:1 или меньше. В неограничивающем варианте осуществления изобретения указанная клетка содержит мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (w)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,20:1 или меньше. В неограничивающем варианте осуществления изобретения указанная клетка содержит мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (ао)-(be) приведенного выше списка. Настоящее изобретение далее относится к клетке вида из рода Streptomyces, содержащей мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (bf) и (bg) приведенного выше списка. В предпочтительном варианте осуществления изобретения вид из рода Streptomyces является видом из рода S. avermitilis. В более предпочтительном варианте осуществления изобретения указанная клетка является клеткой S. avermitilis, способной продуцировать авермектины циклогексил В2:циклогексил В1 в соотношении около 0,40:1 или меньше. Хотя все вышеуказанные мутации могут присутствовать в клетках по настоящему изобретению во внехромосомном элементе, таком как плазмида, желательно, чтобы такие мутации находились в aveC-кодирующей последовательности, встроенной в хромосому S. avermitilis, и предпочтительно, хотя и необязательно, на сайте нативного аллеля aveC. Такие новые штаммы клеток пригодны для крупномасштабного производства коммерчески необходимых авермектинов, таких как дорамектин. Настоящее изобретение далее относится к способу получения авермектинов, который включает культивирование клеток S. avermitilis по настоящему изобретению в культуральных средах в условиях, стимулирующих или индуцирующих продуцирование авермектинов, и выделение указанных авермектинов из культуры. В предпочтительном варианте осуществления изобретения клетки, используемые в данном способе, продуцируют авермектины циклогексил В2 : циклогексил В1 в соотношении 0,35:1 или меньше, более предпочтительно в соотношении около 0,30:1 или меньшее, еще предпочтительнее в соотношении около 0,25:1 или меньше и наиболее предпочтительно в соотношении около 0,20:1 или меньше. В предпочтительном варианте осуществления изобретения клетки, продуцирующие авермектины циклогексил В2 : циклогексил В1 в соотношении 0,35:1 или меньше, содержат мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения клетки, продуцирующие авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,30:1 или меньше, содержат мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (f)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения, клетки, продуцирующие авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,25,1 или меньше, содержат мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (w)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения клетки, продуцирующие авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,20:1 или меньше, содержат мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (ао)-(be) приведенного выше списка. В другом варианте осуществления изобретения клетки продуцируют авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,40:1 или меньше и содержат мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (bf) и (bg) приведенного выше списка. При осуществлении способа по данному изобретению достигается более высокая эффективность получения коммерчески ценных авермектинов, таких как дорамектин. Настоящее изобретение далее относится к композиции авермектинов циклогексил В2 : циклогексил В1, продуцированной клетками Streptomyces avermitilis в культуральной среде, используемой для культивирования указанных клеток, в которой соотношение авермектинов циклогексил В2 : циклогексил В1 составляет примерно 0,35:1 или меньше, предпочтительно около 0,30:1 или меньше, более предпочтительно около 0,25:1 или меньше и наиболее предпочтительно около 0,20:1 или меньше. В конкретном варианте осуществления изобретения композицию авермектинов циклогексил В2 : циклогексил В1 продуцируют клетки штамма S. avermitilis, которые экспрессируют мутированный аллель aveC или его вырожденный вариант, кодирующий генный продукт, в результате чего уменьшается соотношение авермектинов циклогексил В2 : циклогексил В1, продуцируемых указанными клетками, по сравнению к клетками такого же штамма S. avermitilis, не содержащими мутированный аллель aveC и экспрессирующими только аллель aveC дикого типа. В предпочтительном варианте осуществления изобретения, в котором указанная композиция включает авермектины циклогексил В2 : циклогексил В1 в соотношении 0,35:1 или меньше, данную композицию продуцируют клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (а)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения, в котором указанная композиция включает авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,30:1 или меньше, данную композицию продуцируют клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (f)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения, в котором указанная композиция включает авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,25:1 или меньше, данную композицию продуцируют клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (w)-(be) приведенного выше списка. В более предпочтительном варианте осуществления изобретения, в котором указанная композиция включает авермектины циклогексил В2 : циклогексил В1 в соотношении около 0,20:1 или меньше, данную композицию продуцируют клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (ао)-(be) приведенного выше списка. Настоящее изобретение далее относится к композиции авермектинов циклогексил В2 : циклогексил В1, продуцированной клетками Streptomyces avermitilis в культуральной среде, используемой для культивирования указанных клеток, в которой соотношение авермектинов циклогексил В2 :циклогексил В1 составляет примерно 0,40:1, а также клетками, содержащими мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, содержащий комбинацию замен аминокислот, выбираемую из группы, включающей пункты (bf) и (bg) приведенного выше списка. Хотя желательно, чтобы новая композиция авермектинов находилась в культуральной среде, используемой для культивирования клеток, например, в частично или полностью истощенной ферментационной культуральной среде, альтернативно композиция авермектинов может быть частично или по существу полностью очищена от культуральной среды известными биохимическими методами очистки, такими как осаждение сульфатом аммония, диализ, вытеснительное фракционирование, ионообменная хроматография, ВЭЖХ и т.д. Помимо получения новых штаммов S. avermitilis, содержащих клетки, способные продуцировать авермектины циклогексил В2 : циклогексил В1 в меньших соотношениях, объектом настоящего изобретения являются дополнительные мутации, вводимые в клетки S. avermitilis для улучшения характеристик продуцирования авермектинов. В неограничивающем варианте осуществления изобретения клетки по настоящкму изобретению могут включать модификации, повышающие уровни продуцирования авермектинов. В одном варианте осуществления изобретения такие клетки получают в результате (i) мутирования аллеля aveC в клетке S. avermitilis или (ii) введения мутированного аллеля aveC или его вырожденного варианта в клетки штамма S. avermitilis, при этом экспрессия мутированного аллеля вызывает увеличение количества авермектинов, продуцируемых клетками штамма S. avermitilis, экспрессирующими мутированный аллель aveC, по сравнению с клетками такого же штамма, экспрессирующими только аллель aveC дикого типа, и последующего отбора трансформированных клеток, продуцирующих авермектины в большем количестве по сравнению с клетками такого же штамма, экспрессирующими только аллель aveC дикого типа. Например, аллель aveC можно модифицировать так, чтобы он включал сильный промотор, например сильный структурный промотор ermE, выделенный из Saccharopolyspora erythraea, расположенный вверху от открытой рамки считывания aveC и функционально связанный с указанной рамкой считывания. В другом варианте осуществления изобретения одну или несколько мутаций можно ввести в гены aveR1 и/или aveR2 S. avermitilis, в результате чего увеличивается продуцирование авермектинов, как это описано в патенте США № 6197591, выданном Stutzman-Engwall et al. 6 марта 2001 г. 5.4. Применение авермектинов Авермектины являются весьма активными противопаразитарными средствами, применяемыми в качестве антигельминтных средств, эктопаразитицидов, инсектицидов и акарицидов. Соединения авермектинов, полученные способами по настоящему изобретению, пригодны для любых вышеуказанных целей. Например, соединения авермектинов, полученные по настоящему изобретению, можно использовать для лечения разных заболеваний или нарушений у человека, в частности, заболеваний или нарушений, вызванных паразитарными инфекциями. См., например, Ikeda and Omura, 1997, Chem. Rev. 97(7):2591-2609. В частности, соединения авермектинов, полученные по настоящему изобретению, можно эффективно использовать для лечения разных заболеваний или нарушений, вызываемых эндопаразитами, такими как паразитарные нематоды, которые могут инфицировать человека, домашних животных, свиней, овец, домашнюю птицу, лошадей или крупный рогатый скот. В частности, соединения авермектинов, полученные по настоящему изобретению, эффективно воздействуют на нематоды, инфицирующие человека, а также на нематоды, инфицирующие животных разных видов. Указанные нематоды включают желудочно-кишечных паразитов, таких как Ancylostoma, Necator, Ascaris, Strongyloides, Trichinella, Capillaria, Trichuris, Enterobius, Dirofilaria, паразитов, обнаруживаемых в крови и других тканях или органах, таких как филярия, и паразитов, вызывающих острые кишечные расстройства, таких как Strongyloides и Trichinella. Соединения авермектинов, полученные по настоящему изобретению, пригодны также для лечения эктопаразитарных инфекций, включающих, например, инвазию млекопитающих и птиц членистоногими, в частности клещами, вшами, блохами, падальными мухами, жалящими насекомыми или мигрирующими личинками двукрылых насекомых, которые могут поражать крупный рогатый скот и лошадей наряду с прочими животными. Соединения авермектинов, полученные по настоящему изобретению, можно также использовать в качестве инсектицидов против домашних паразитов, таких как, например, тараканы, платяная моль, кожеед и комнатная муха, а также против вредителей, портящих запасы зерна и сельскохозяйственные растения, которые включают наряду с прочими вредителями клещика паутинного, тлю, гусениц и прямокрылых насекомых, таких как саранча. Соединения авермектина, полученные по настоящему изобретению, можно использовать для лечения таких животных, как овцы, крупный рогатый скот, лошади, олени, козы, свиньи, птицы, включая домашнюю птицу, собаки и кошки. Соединение авермектина, полученное по настоящему изобретению, вводят в виде препарата, соответствующего конкретному способу введения, определенному виду животного, нуждающегося в лечении, и паразитам или насекомым, подлежащим уничтожению. При использовании в качестве паразитицида соединение авермектина, полученное по настоящему изобретению, можно вводить перорально в виде капсулы, болюса, таблетки или жидкости или альтернативно можно вводить в виде обливания, инъекции или имплантата. Такие препараты получают обычным способом в соответствии со стандартной практикой, принятой в ветеринарии. Так, капсулы, болюсы или таблетки можно получить, смешивая активный ингредиент с приемлемым тонко измельченным разбавителем или носителем, дополнительно содержащим вещество, улучшающее распадаемость, и/или связывающее вещество, такое как крахмал, лактоза, тальк, стеарат магния и т.д. Смачивающий препарат можно получить, диспергируя активный ингредиент в водном растворе с диспергирующим или смачивающим агентом и т.д. Инъекционные препараты можно получить в виде стерильного раствора, который может содержать другие вещества, такие как, например, соли и/или глюкоза, в количестве, достаточном для получения изотонического раствора по отношению к крови. Такие препараты могут содержать разное количество активного соединения в зависимости от состояния нуждающегося субъекта или вида животного, подлежащего лечению, тяжести заболевания, типа инфекции и массы тела хозяина. Доза активного соединения, предназначенная для перорального введения, обычно составляет от около 0,001 до 10 мг/кг массы тела нуждающегося субъекта или животного при введении в виде однократной дозы или раздельных доз в течение 1-5 дней. Однако в некоторых случаях могут быть необходимы более высокие или более низкие дозы, определяемые лечащим врачом или ветеринаром на основании клиническим симптомов. В качестве альтернативы соединение авермектина, полученное по настоящему изобретению, можно вводить вместе с кормом для животных, и для указанной цели может быть получена концентрированная кормовая добавка или премикс, предназначенные для смешивания с обычным кормом для животных. При использовании в качестве инсектицида и для уничножения сельскохозяйственных вредителей соединение авермектина, полученное по настоящему изобретению, можно применять в виде раствора для опрыскивания, пылевидного препарата, эмульсии и подобных препаратов в соответствии с обычной сельскохозяйственной практикой. 6. Пример. Ферментация Streptomyces avermitilis и анализ авермектинов В2:В1 Штаммы, в которых отсутствует 2-кетокислота-дегидрогеназа с разветвленной цепью и 5-О-метилтрансфераза, не продуцируют авермектины без добавления в ферментационную среду жирных кислот. Данный пример показывает, что в таких мутантах можно получить авермектины В2:В1 в разных соотношениях, если инициировать биосинтез в присутствии разных жирных кислот. 6.1. Вещества и способы Streptomyces avermitilis АТСС 53692 хранили при -70°С в виде цельного бульона, полученного в среде для посева, включающей: крахмал (Nadex, Laing National) – 20 г; фармацевтическую среду (Trader’s Protein, Memphis, TN) – 15 г; ардамин рН (Yeast Products Inc.) – 5 г; карбонат кальция – 1 г. Конечный объем доводили до 1 литра, добавляя водопроводную воду, рН доводили до 7,2, после чего среду обрабатывали в автоклаве при 121°С в течение 25 минут. Два мл оттаявшей суспензии вышеуказанного препарата использовали для инокуляции 50 мл аналогичной среды, находящейся в колбе. Смесь инкубировали при 28°С в течение 48 часов в роторном шейкере со скоростью вращения 180 оборотов/мин, 2 мл бульона использовали для инокуляции 50 мл продуцирующей среды, находящейся в колбе, которая содержит: крахмал – 80 г; карбонат кальция – 7 г; фармацевтическую среду – 5 г; вторичный кислый фосфат калия – 1 г; сульфат магния – 1 г; глутаминовую кислоту – 0,6 г; гептагидрат сульфата железа – 0,01 г; сульфат цинка – 0,001 г; сульфат магния (II) – 0,001 г. Конечный объем доводили до 1 литра, добавляя водопроводную воду, рН доводили до 7,2, после чего среду обрабатывали в автоклаве при 121°С в течение 25 минут. Разные субстраты на основе карбоновой кислоты (см. таблицу 1) растворяли в метаноле и добавляли к ферментационному бульону через 24 часа после инокуляции до конечной концентрации 0,2 г/литр. Ферментационный бульон инкубировали в течение 14 дней при 28°С, затем бульон центрифугировали (2500 оборотов/мин в течение 2 минут) и супернатант декантировали. Дебрис мицелия экстрагировали ацетоном (15 мл) и дихлорметаном (30 мл), органическую фазу отделяли, фильтровали и упаривали досуха. Остаток поглощали метанолом (1 мл) и анализировали методом ВЭЖХ в жидкостном хроматографе Hewlett-Packard 1090A, оснащенном сканирующим диодным детектором, настроенным на длину волны 240 нм. Использовали колонку Backman Ultrasphere C-18, 5 мкм, 4,6 мм × 25 см, находящуюся при температуре 40°С. В колонку инъецировали двадцать пять мкл вышеуказанного раствора в метаноле. Элюирование производили линейным градиентом смеси метанол-вода от 80:20 до 95:5 в течение 40 минут со скоростью 0,85 мл/мин. Для калибровки чувствительности детектора использовали две стандартные концентрации авермектина циклогексил В1 и измеряли площадь под кривыми для авермектинов В2 и В1. 6.2. Результаты Значения времени удерживания при выполнении ВЭЖХ, полученные для авермектинов В2 и В1, и соотношения 2:1 приведены в таблице 1.
Данные, приведенные в таблице 1, свидетельствуют о чрезвычайно широком диапазоне соотношений авермектинов В2:В1 в продукте, указывая на значительную разницу результатов дегидративного превращения соединений класса 2 в соединения класса 1 в зависимости от инициирующего звена в боковой цепи жирной кислоты. Полученные данные показывают, что изменение соотношений В2:В1 в результате перестройки белка AveC может быть специфическим для конкретных субстратов. Поэтому скрининг мутантов, характеризующихся изменением соотношения В2:В1, полученных при использовании определенного субстрата, необходимо производить в присутствии данного субстрата. В приведенных ниже примерах в качестве субстрата для скрининга использована циклогексанкарбоновая кислота. Однако данный субстрат был использован только в качестве примера его возможного применения, и объем настоящего изобретения не ограничен только указанным субстратом. 7. Пример. Выделение гена aveC В данном примере описано выделение и исследование области хромосомы Streptomyces avermitilis, кодирующей продукт гена AveC. Как показано ниже, было установлено, что ген aveC может изменять соотношение продуцируемых авермектинов циклогексил В2 : циклогексил В1 (В2:В1). 7.1. Вещества и способы 7.1.1. Выращивание Streptomyces для выделения ДНК Streptomyces выращивали нижеследующим способом. Единичные колонии S. avermitilis АТСС 31272 (изолят единичной колонии №2) выделяли в 1/2-кратной среде YPD-6, содержащей дрожжевой экстракт Difco – 5 г; бактопептон Difco – 5 г; декстрозу – 2,5 г; MOPS – 5 г; бакто-агар Difco – 15 г. Конечный объем доводили до 1 литра, добавляя дистиллированную воду, рН доводили до 7,0, после чего среду обрабатывали в автоклаве при 121°С в течение 25 минут. Мицелий, выращенный в вышеуказанной среде, использовали для инокуляции 10 мл среды TSB (триптиказо-соевый бульон Difco – 30 г в 1 литре дистиллированной воды, обработанный в автоклаве при 121°С в течение 25 минут) в пробирке 25 мм × 150 мм, которую встряхивали (300 оборотов/мин) при 28°С в течение 48-72 часов. 7.1.2. Выделение хромосомной ДНК из Streptomyces Аликвоты (0,25 мл или 0,5 мл) мицелия, выращенного вышеописанным способом, вводили в 1,5 мл микроцентрифужные пробирки и клетки концентрировали центрифугированием с ускорением 12000×g в течение 60 секунд. Супернатант декантировали и клетки вновь суспендировали в 0,25 мл буфера TSE (20 мл 1,5 М сахарозы, 2,5 мл 1 М трис-буфера с HCl, рН 8,0, 2,5 мл 1 М EDTA, рН 8,0 и 75 мл дистиллированной воды), содержащем 2 мг/мл лизоцима. Образцы инкубировали, встряхивая, при 37°С в течение 20 минут, вводили в аппарат для автоматического выделения нуклеиновой кислоты AutoGen 540 Альтернативно, 5 мл мицелия вводили в пробирку 17 мм × 100 мм, клетки концентрировали центрифугированием со скоростью 3000 оборотов/мин в течение 5 минут и удаляли супернатант. Клетки вновь суспендировали в 1 мл буфера TSE, концентрировали центрифугированием со скоростью 3000 оборотов/мин в течение 5 минут и удаляли супернатант. Клетки вновь суспендировали в 1 мл буфера TSE, содержащего 2 мг/мл лизоцима, и инкубировали, встряхивая, при 37°С в течение 30-60 минут. После окончания инкубации добавляли 0,5 мл 10% раствора додецилсульфата натрия (SDS) и инкубировали клетки при 37°С до окончания лизиса. Лизат инкубировали при 65°С в течение 10 минут, охлаждали до комнатной температуры, вводили в две 1,5 мл пробирки Эппендорфа и экстрагировали 1 раз 0,5 мл смеси фенол/хлороформ (50% фенола, предварительно уравновешенного 0,5М трис-буфера, рН 8,0; 50% хлороформа). Водную фазу удаляли и экстрагировали 2-5 раз смесью хлороформ : изоамиловый спирт (24:1). ДНК осаждали, для чего добавляли 1/10 объема 3М раствора ацетата натрия, рН 4,8, инкубировали смесь на льду в течение 10 минут, центрифугировали смесь со скоростью 15000 оборотов/мин при 5°С в течение 10 минут и переносили супернатант в чистую пробирку, в которую добавляли 1 объем изопропанола. Смесь супернатанта с изопропанолом инкубировали на льду в течение 20 минут, центрифугировали со скоростью 15000 оборотов/мин в течение 20 минут при 5°С, супернатант удаляли и осадок ДНК промывали 1 раз 70% этанолом. Затем осадок сушили и ДНК вновь суспендировали в буфере ТЕ (10 мМ трис-буфера, 1 мМ EDTA, рН 8,0). 7.1.3. Выделение плазмидной ДНК из Streptomyces Аликвоту (1,0 мл) мицелия вводили в 1,5 мл микроцентрифужные пробирки и клетки концентрировали центрифугированием с ускорением 12000×g в течение 60 секунд. Супернатант декантировали, клетки вновь суспендировали в 1,0 мл 10,3% раствора сахарозы, концентрировали центрифугированием с ускорением 12000×g в течение 60 секунд и супернатант декантировали. Клетки вновь суспендировали в 0,25 мл буфера TSE, содержащего 2 мг/мл лизоцима, инкубировали, встряхивая, при 37°С в течение 20 минут и вводили в аппарат для автоматического выделения нуклеиновой кислоты AutoGen 540 Альтернативно 1,5 мл мицелия вводили в 1,5 мл микроцентрифужные пробирки и клетки концентрировали центрифугированием с ускорением 12000×g в течение 60 секунд. Супернатант декантировали, клетки вновь суспендировали в 1,0 мл 10,3% раствора сахарозы, концентрировали центрифугированием с ускорением 12000×g в течение 60 секунд и супернатант декантировали. Клетки вновь суспендировали в 0,5 мл буфера TSE, содержащего 2 мг/мл лизоцима, и инкубировали при 37°С в течение 15-30 минут. После окончания инкубации добавляли 0,25 мл щелочного раствора SDS (0,3 н. раствор в NaOH, 2% SDS) и клетки инкубировали при 55°С в течение 15-30 минут или до тех пор, пока раствор не становился прозрачным. К раствору ДНК добавляли ацетат натрия (0,1 мл, 3М, рН 4,8) и инкубировали на льду в течение 10 минут. Образцы ДНК центрифугировали со скоростью 14000 оборотов/мин в течение 10 минут при 5°С. Супернатант переносили в чистую пробирку, добавляли 0,2 мл смеси фенол : хлороформ (50% фенола : 50% хлороформа) и осторожно смешивали. Раствор ДНК центрифцугировали со скоростью 14000 оборотов/мин в течение 10 минут при 5°С и верхний слой переносили в чистую пробирку Эппендорфа. Добавляли изопропанол (0,75 мл), раствор осторожно смешивали и инкубировали при комнатной температуре в течение 20 минут. Раствор ДНК центрифугировали со скоростью 14000 оборотов/мин в течение 15 минут при 5°С, супернатант удаляли и осадок ДНК промывали 70% этанолом, сушили и вновь суспендировали в буфере ТЕ. 7.1.4. Выделение плазмидной ДНК из E. coli Единичную трансформированную колонию E. coli инокулировали в 5 мл среды Луриа-Бертани (LB) (бактотриптон – 10 г, бактодрожжевой экстракт – 5 г и NaCl – 10 г в 1 литре дистиллированной Н2О, рН 7,0, обрабатывали в автоклаве при 121°С в течение 25 минут и добавляли 100 мкг/мл ампициллина). Культуру инкубировали в течение ночи и 1 мл аликвоту вводили в 1,5 мл микроцентрифужную пробирку. Образцы культуры вводили в аппарат для автоматического выделения нуклеиновой кислоты AutoGen 540 7.1.5. Получение и трансформация протопластов S. avermitilis Единичные колонии S. avermitilis выделяли в 1/2-кратной среде YPD-6. Мицелий использовали для инокуляции 10 мл среды TSB в пробирке 25 мм × 150 мм и инкубировали, встряхивая (300 оборотов/мин), при 28°С в течение 48 часов. Один мл мицелия использовали для инокуляции 50 мл среды YEME. Среда YEME содержит в одном литре: дрожжевой экстракт Difco – 3 г; бактопептон Difco – 5 г; солодовый экстракт Difco – 3 г; сахарозу – 300 г. Культуру обрабатывали в автоклаве при 121°С в течение 25 минут и добавляли 2,5 М раствор MgCl2·6H2O (отдельно обработанный в автоклаве при 121°С в течение 25 мин) – 2 мл и глицин (20%) (стерилизованный фильтрацией) – 25 мл. Мицелий выращивали при 30°С в течение 48-72 часов и собирали в 50 мл центрифужной пробирке (Falcon) центрифугированием со скоростью 3000 оборотов/мин в течение 20 минут. Супернатант декантировали и мицелий вновь суспендировали в буфере Р, содержащем: сахарозу – 205 г; K2SO4 – 0,25 г; MgCl2·6H2O – 2,02 г; Н2О – 600 мл; К2РО4 (0,5%) – 10 мл; раствор микроэлементов* – 20 мл; CaCl2·2H2O (3,68%) – 100 мл и буфер MES (1,0 М, рН 6,5) – 10 мл. (*Раствор микроэлементов содержит в одном литре: ZnCl2 – 40 мг; FeCl3·6H2O – 200 мг; CuCl2·2H2O – 10 мг; MnCl2·4H2O – 10 мг; Na2B4O7·10H2O – 10 мг; (NH4)6Mo7O24·4H2O – 10 мг). Показатель рН доводили до 6,5, конечный объем доводили до 1 литра и среду фильтровали в горячем состоянии через фильтр с размером отверстий 0,45 мкм. Мицелий осаждали со скоростью 3000 оборотов/мин в течение 20 минут, супернатант декантировали и мицелий вновь суспендировали в 20 мл буфера Р, содержащего 2 мг/мл лизоцима. Мицелий инкубировали, встряхивая, при 35°С в течение 15 минут и исследовали под микроскопом для определения степени образования протопластов. После окончания образования протопластов указанные протопласты центрифугировали со скоростью 8000 оборотов/мин в течение 10 минут. Супернатант удаляли и протопласты вновь суспендировали в 10 мл буфера Р. Протопласты центрифугировали со скоростью 8000 оборотов/мин в течение 10 минут, супернатант удаляли, протопласты вновь суспендировали в 2 мл буфера Р и примерно 1×109 протопластов помещали в 2,0 мл криогенные сосуды (Nalgene). Сосуд, содержащий 1×109 протопластов, центрифугировали со скоростью 8000 оборотов/мин в течение 10 минут, супернатант удаляли и протопласты вновь суспендировали в 0,1 мл буфера Р. К протопластам добавляли 2-5 мкг трансформирующей ДНК, после чего сразу же добавляли 0,5 мл рабочего буфера Т. Основной буфер Т содержит: PEG-1000 (Sigma) – 25 г; сахарозу – 2,5 г; Н2О – 83 мл. Показатель рН доводили до 8,8, добавляя 1 н. раствор NaOH (стерилизованный фильтрацией), после чего основной буфер Т стерилизовали фильтрацией и хранили при 4°С. Рабочий буфер Т, полученный в день использования, содержал основной буфер Т – 8,3 мл; К2РО4 (4 мМ) – 1,0 мл; CaCl2.2H2O (5 M) – 0,2 мл и TES (1 M, pH 8) – 0,5 мл. Все компоненты рабочего буфера Т отдельно стерилизовали фильтрацией. Через 20 секунд после добавления к протопластам буфера Т добавляли 1,0 мл буфера Р и протопласты центрифугировали со скоростью 8000 оборотов/мин в течение 10 минут. Супернатант декантировали и протопласты вновь суспендировали в 0,1 мл буфера Р. Затем протопласты культивировали на среде RM14, содержащей: сахарозу – 205 г; K2SO4 – 0,25 г; MgCl2·6H2O – 10,12 г; глюкозу – 10 г; касаминокислоты Difco – 0,1 г; дрожжевой экстракт Difco – 5 г; агар на отваре из овсяной муки Difco – 3 г; бактоагар Difco – 22 г; дистиллированную Н2О – 800 мл. Раствор обрабатывали в автоклаве при 121°С в течение 25 минут. После обработки в автоклаве добавляли нижеследующие стерильные компоненты: К2РО4 (0,5%) – 10 мл; CaCl2·2H2O (5 M) – 5 мл; L-пролин (20%) – 15 мл; буфер MES (1,0 М, рН 6,5) – 10 мл; раствор микроэлементов (аналогичный вышеуказанному) – 2 мл; циклогексимид (25 мг/мл) – 40 мл и 1 н. раствор NaOH – 2 мл. На планшет наносили двадцать пять мл среды RM14 и сушили в течение 24 часов перед использованием. Протопласты инкубировали в условиях 95% влажности при 30°С в течение 20-24 часов. Для отбора трансформантов, устойчивых к тиострептону, 1 мл покрывающего буфера, содержащего 125 мкг/мл тиострептона, равномерно распределяли поверх планшетов для регенерации RM14. Покрывающий буфер содержит в 100 мл: сахарозу – 10,3 г; раствор микроэлементов (аналогичный вышеуказанному) – 0,2 мл и буфер MES (1М, рН 6,5) – 1 мл. Протопласты инкубировали в условиях 95% влажности при 30°С в течение 7-14 дней до выявления колоний, устойчивых к тиострептону (Thior). 7.1.6. Трансформация протопластов Streptomyces lividans В некоторых случаях для трансформации использовали протопласты S. lividans TK64 (предоставлены институтом John Innes Institute, Norwich, U.K.). В процессе исследования были использованы способы и композиции для выращивания, образования и трансформации протопластов S. lividans, описанные в публикации Hopwood et al., 1985, Genetic Manipulation of Streptomyces, A Laboratory Manual, John Innes Foundation, Norwich, U.K. Плазмидную ДНК выделяли из трансформантов S. lividans в соответствии с описанием, приведенным в разделе 7.1.3 вышеуказанной публикации. 7.1.7. Анализ ферментации штаммов S. avermitilis Мицелий S. avermitilis, выращиваемый в 1/2-кратной среде YPD-6 в течение 4-7 дней, инокулировали в пробирки 2,5 см × 15 см (1×6 дюймов), содержащие 8 мл предварительно полученной среды и два 5 мм стеклянных шарика. Предварительно полученная среда содержит: растворимый крахмал (крахмал, проваренный в жидком состоянии, или KOSO, Japan Corn Starch Co., Nagoya) – 20 г/л; фармацевтическую среду – 15 г/л; ардамин рН – 5 г/л (Champlain Ind., Clifton, NJ); СаСО3 – 2 г/л; двукратный объем жирных кислот с разветвленной цепью (bcfa), находящихся в среде в конечной концентрации, равной 50 ч/млн 2-(+)-метилмасляной кислоты, 60 ч/млн изомасляной кислоты и 20 ч/млн изовалериановой кислоты. Показатель рН доводили до 7,2 и среду обрабатывали в автоклаве при 121°С в течение 25 минут. Пробирку встряхивали под углом 17° со скоростью 215 оборотов/мин при 29°С в течение 3 дней. 2 мл аликвоту посевной культуры использовалия для инокуляции 25 мл продуцирующей среды в 300 мл колбе Эрленмейера, содержащей крахмал (крахмал, проваренный в жидком состоянии, или KOSO) – 160 г/л; нутрисой (Archer Daniels Midland, Decatur, IL) – 10 г/л; ардамин рН – 10 г/л; К2НРО4 – 2 г/л; MgSO4·4H2O – 2 г/л; FeSO4·7H2O – 0,02 г/л; MnCl2 – 0,002 г/л; ZnSO4·7H2O – 0,002 г/л; СаСО3 – 14 г/л; двукратный объем жирных кислот с разветвленной цепью (bcfa) (аналогичных вышеуказанным) и циклогексанкарбоновую кислоту (СНС) (полученную в виде 20% раствора с рН 7,0) – 800 ч/млн. Показатель рН доводили до 6,9 и среду обрабатывали в автоклаве при 121°С в течение 25 минут. (Как указывалось выше, вместо СНС можно использовать другие инициирующие звенья (см., например, таблицу 1). После инокуляции колбу инкубировали, встряхивая со скоростью 200 оборотов/мин при 29°С в течение 12 дней. После окончания инкубации из колбы удаляли 2 мл образец, разводили его в 8 мл метанола, смешивали и смесь центрифугировали с ускорением 1250×g в течение 10 минут до образования дебриса. Супернатант анализировали методом ВЭЖХ, используя колонку Beckman Ultrasphere ODS (25 см × 4,6 мм внутренний диаметр) со скоростью потока 0,75 мл/мин, и производили исследование при оптической плотности 240 нм. В качестве подвижной фазы использовали смесь метанол/вода/ацетонитрил (86:8,9:5,1). 7.1.8. Выделение генов PKS S. avermitilis Была получена космидная библиотека хромосомной ДНК S. avermitilis (АТСС 31272, SC-2), которую гибридизировали с зондом на основе кетосинтазы (KS), полученным из фрагмента гена поликетидсинтазы (PKS) Saccharopolyspora erythraea. Получение космидных библиотек подробно описано в приведенной выше публикации Sambrook et al., 1989. Получение библиотек хромосомной ДНК Streptomyces подробно описано в приведенной выше публикации Hopwood et al., 1985. Космидные клоны, содержащие области, гибридизирующие с кетосинтазой, были идентифицированы в результате гибридизации с фрагментом NdeI/Eco47III длиной 2,7 т.п.о., выделенным из плазмиды рЕХ26 (любезно предоставленной д-ром P. Leadlay, Cambridge, UK). Примерно 5 нг рЕХ25 гидролизовали, используя NdeI и Eco47III. Реакционную смесь помещали на 0,8% агарозный гель SeaPlaque GTG (FMC BioProducts, Rockland, ME). После выполнения электрофореза из геля вырезали фрагмент NdeI/Eco47III длиной 2,7 т.п.о. и методом Fast Protocol выделяли ДНК, используя GELазу Методом гибридизации колоний было исследовано примерно 1800 космидных клонов. Было идентифицировано десять клонов, которые интенсивно гибридизировали с KS-зондом Sacc. erythraea. Колонии E. coli, содержащие космидную ДНК, выращивали в жидкой среде LB, после чего космидную ДНК выделяли из всех культур в аппарате для автоматического выделения нуклеиновой кислоты AutoGen 540 7.1.9. Идентификация ДНК, модулирующей соотношения авермектинов В2:В1, и идентификация открытой рамки считывания aveC Нижеследующие способы были использованы для исследования способности субклонированных фрагментов, выделенных из космидного клона pSЕ66, модулировать соотношения авермектинов В2:В1 в мутантах AveC. Плазмиду pSE66 (5 мкг) гидролизовали Sacl и BamHI. Реакционную смесь помещали на 0,8% агарозный гель GTG SeaPlaque Полученные протопласты штамма 1100-SC38 S. avermitilis (штамм компании Pfizer) трансформировали плазмидой pSE119 в соответствии с описанием, приведенным в разделе 7.1.5. Штамм 1100-SC38 является мутантом, продуцирующим значительно больше авермектина циклогексил В2 по сравнению с авермектином циклогексил В1 при добавлении циклогексанкарбоновой кислоты (соотношение В2:В1 равно примерно 30:1). Плазмиду pSE119, использованную для трансформации протопластов S. avermitilis, выделяли из штамма GM2163 E. coli (предоставленного д-ром B.J. Bachmann, Curator, E. coli Genetic Stock Center, Yale University), штамма DM1 (BRL) E. coli или штамма ТК64 S. lividans. Устойчивые к тиострептону трансформанты штамма 1100-SC38 выделяли и исследовали, анализируя методом ВЭЖХ продукты ферментации. Трансформанты штамма 1100-SC38 S. avermitilis, содержащие плазмиду pSE119, продуцировали авермектины циклогексил В2 : циклогексил В1 в измененном соотношении, равном примерно 3,7:1 (таблица 2). Установив, что плазмида pSE119 способна модулировать соотношение авермектинов В2:В1 в мутанте AveC, ДНК-вставку секвенировали. При помощи набора для выделения плазмидной ДНК (Qiagen, Valencia, CA), используемого в соответствии с инструкциями изготовителя, было выделено примерно 10 мкг плазмиды pSE119, которую секвенировали в автоматическом секвенаторе ДНК ABI 373A (Perkin Elmer, Foster City, CA). Данные, полученные для последовательности, были скомпонованы и отредактированы при помощи программы Genetic Computer Group (GCG, Madison, WI). Последовательность ДНК и открытая рамка считывания aveC показаны на фигуре 1 (SEQ ID No.1). Новая плазмида, получившая название pSE118, была создана следующим образом. Примерно 5 мкл pSE66 гидролизовали SphI и BamHI. Реакционную смесь помещали на 0,8% агарозный гель GTG SeaPlaque (FMC BioProducts), после выполнения электрофореза из геля вырезали фрагмент SphI/BamHI длиной 2,8 т.п.о. и методом Fast Protocol из геля выделяли ДНК, используя GELазу Протопласты штамма 1100-SC38 S. avermitilis трансформировали плазмидой pSE118 вышеуказанным способом. Трансформанты, устойчивые к тиострептону, штамма 1100-SC38 выделяли и исследовали, анализируя методом ВЭЖХ продукты ферментации. У трансформантов штамма 1100-SC38 S. avermitilis, содержащих плазмиду pSE118, не были изменены соотношения авермектинов циклогексил В2:циклогексил В1 по сравнению со штаммом 1100-SC38 (таблица 2). 7.1.10. Амплификация гена aveC из хромосомной ДНК S. avermitilis методом PCR Фрагмент длиной 1,2 т.п.о., содержащий открытую рамку считывания aveC, выделяли из хромосомной ДНК S. avermitilis при помощи амплификацим методом PCR, используя затравки, созданные на основе полученной выше нуклеотидной последовательности aveC. Затравки для PCR были предоставлены компанией Genosys Biotechnologies, Inc. (Texac). Нижняя затравка представляла собой: 5′-TCACGAAACCGGACACAC-3′ (SEQ ID No.6); и верхняя затравка представляла собой: 5′-CATGATCGCTGAACCGAG-3′ (SEQ ID No.7). Реакцию PCR выполняли с использованием полимеразы Deep Vent 72°С в течение 2 минут (стадия удлинения). 24 последующих цикла имели аналогичный тепловой профиль за исключением того, что стадия денатурации была сокращена до 45 секунд и стадия отжига была сокращена до 1 минуты. Продукт PCR подвергали электрофорезу в 1% агарозном геле, при этом была обнаружена одна полоса ДНК длиной 1,2 т.п.о. Данную ДНК очищали от геля и лигировали с 25 нг линейного, дефосфорилированного вектора pCR-Blunt (Invetrogen) в молярном соотношении вектор:вставка, равном 1:10, в соответствии с инструкциями изготовителя. Лигированную смесь использовали для трансформации компетентных клеток E. coli One Shot ДНК-вставку из плазмиды pSE179 выделяли гидролизом BamHI/XbaI, отделяли электрофорезом, очищали от геля и лигировали с шаттл-вектором pWHM3, который также предварительно гидролизовали BamHI/XbaI, с достижением концентрации полной ДНК, равной 1 мкг, при молярном соотношении вектор : вставка, равном 1:5. Лигированную смесь использовали для троансформации компетентных клеток DH5 7.2. Результаты Установлено, что фрагмент SacI/BamHI длиной 2,9 т.п.о., выделенный из плазмиды pSE119, при введении в штамм 1100-SC38 S. avermitilis значительно изменяет соотношение продуцируемых авермектинов В2:В1. Штамм 1100-SC38 S. avermitilis обычно характеризовался соотношением В2:В1, равным примерно 30:1, но в результате трансформации вектором, содержащим фрагмент SacI/BamHI длиной 2,9 т.п.о., соотношение авермектинов В2:В1 уменьшилось примерно до 3,7:1. Результаты анализа культур трансформантов после ферментации подтвердили наличие трансформирующей ДНК. Фрагмент pSE119 длиной 2,9 т.п.о. секвенировали и идентифицировали открытую рамку считывания длиной 0,9 т.п.о. (фигура 1) (SEQ ID No.1), включающую фрагмент PstI/SphI, который предварительно был мутирован с возможностью продуцирования только продуктов В2 (Ikeda et al., 1995, см. выше). Сравнение данной открытой рамки считывания или ее соответствующего выделенного полипептида с известными базами данных (GenEMBL, SWISS-PROT) не выявило какой-либо значительной гомологии с известными последовательностями ДНК или белка. В таблице 2 приведены результаты анализа ферментации штамма 1100-SC38 S. avermitilis, трансформированного разными плазмидами.
8. Пример. Создание мутантов AveC S. Avermitilis В данном примере описано создание нескольких разных мутантов AveC S. avermitilis с использованием вышеописанных композиций и способов. Общее описание методов введения мутаций в ген Streptomyces дано в статье Kieser and Hopwood, 1991, Meth. Enzym. 204:430-458. Более подробно указанные методы описаны в статьях Anzai et al., 1988, J. Antibiot. XLI(2):226-233 и Stutzman-Engwall et al., 1992, J. Bacteriol. 174(1):144-154. Указанные публикации полностью включены в данное описание в качестве ссылки. 8.1. Инактивация гена aveC S. avermitilis Мутанты AveC, содержащие инактивированные гены aveC, были созданы несколькими способами, подробно описанными ниже. В соответствии с первым способом фрагмент SphI/PstI длиной 640 п.о., находящийся внутри гена aveC плазмиды pSE119 (плазмида, описанная в разделе 7.1.9.), заменяли геном ermE (ген устойчивости к эритромицину) из Sacc. erytraea. Ген ermE выделяли из плазмиды pIJ4026 (предоставлена институтом John Innes Institute, Norwich, U.K.; см. также Bibb et al., 1985, Gene 41:357-368) путем гидролиза рестрикционными ферментами BglII и EcoRI с последующим выполнением электрофореза и очищали от геля. Полученный фрагмент длиной 1,7 т.п.о. лигировали с плазмидой pGEM7Zf (Promega), которую гидролизовали BamHI и EcoRI, и лигированную смесь вводили в компетентные клетки DH5 Плазмиду pSE118 (описанную в разделе 7.1.9.) гидролизовали SphI и BamHI, гидролизат подвергали электрофорезу и вставку SphI/BamHI длиной 2,8 т.п.о. очищали от геля. Плазмиду pSE119 гидролизовали PstI и EcoRI, гидролизат подвергали электрофорезу и вставку PstI/EcoRI длиной 1,5 т.п.о. очищали от геля. Шаттл-вектор pWHM3 гидролизовали BamHI и EcoRI. Плазмиду pSE27 гидролизовали PstI и SphI, гидролизат подвергали электрофорезу и вставку PstI/SphI длиной 1,7 т.п.о. очищали от геля. Все четыре фрагмента (то есть фрагменты длиной 2,8 т.п.о., 1,5 т.п.о., 7,2 т.п.о., 1,7 т.п.о.) лигировали друг с другом методом четырехстороннего лигирования. Лигированную смесь вводили в компетентные клетки DH5 Плазмиду pSE180 вводили в клетки ТК64 S. lividans и трансформированные колонии идентифицировали по устойчивости к тиострептону и эритромицину. Плазмиду pSE180 выделяли из S. lividans и использовали для трансформации протопластов S. avermitilis. Были идентифицированы четыре устойчивых к тиострептону трансформанта S. avermitilis, после чего полученные протопласты культивировали в неизбирательных условиях на среде RM14. После регенерации протопластов единичные колонии исследовали на наличие устойчивости к эритромицину и отсутствие устойчивости к тиострептону, что свидетельствовало о встраивании в хромосому инактивированного гена aveC и утрату свободного репликона. Был идентифицирован один трансформант Ermr Thios, который получил название штамма SE180-11. Из штамма SE180-11 выделяли полную хромосомную ДНК, гидролизовали рестрикционными ферментами BamHI, HindIII, PstI или SpjI, выделяли электрофорезом в 0,8% агарозном геле, переносили на найлоновые мембраны и гибридизировали с зондом ermE. Указанные анализы показали, что встраивание гена устойчивости ermE в хромосому и одновременная делеция фрагмента PstI/SphI длиной 640 п.о. происходили в результате двойного кроссинговера. Анализ продуктов ферментации штамма SЕ180-11 методом ВЭЖХ показал отсутствие продуцирования нормальных авермектинов (фигура 5А). В соответствии со вторым способом инактивации гена aveC из хромосомы штамма SE180-11 S. avermitilis удаляли ген ermE длиной 1,7 т.п.о., оставляя делецию PstI/SphI длиной 640 п.о. в гене aveC. Плазмида замещения гена была создана следующим образом: плазмиду pSE180 частично гидролизовали XbaI и фрагмент длиной 11,4 т.п.о. очищали от геля. В полосе, соответствующей фрагменту длиной 11,4 т.п.о., отсутствует ген устойчивости ermE длиной 1,7 т.п.о. Затем ДНК лигировали и вводили в клетки DH5 В соответствии с третьим способом инактивации гена aveC в хромосомном гене aveC вызывали сдвиг рамки считывания, добавляя два остатка G после С в положении нуклеотида 471 при помощи PCR, в результате чего был создан сайт BspE1. Наличие сконструированного сайта BspE1 позволило обнаружить замену гена. Затравки для PCR, созданные с возможностью введения мутации со сдвигом рамки считывания в ген aveC, были предоставлены компанией Genosys Biotechnologies, Inc. Нжняя затравка представляла собой: 5′-GGTTCCGGATGCCGTTCTCG-3′ (SEQ ID No.8), и верхняя затравка представляла собой: 5′-AACTCCGGT CGACTCCCCTTC-3′ (SEQ ID No.9). Условия выполнения PCR были аналогичны описанным в разделе 7.1.10. Продукт PCR длиной 666 п.о. гидролизовали SphI с образованием двух фрагментов длиной соответственно 278 п.о. и 388 п.о. Фрагмент длиной 388 п.о. очищали от геля. Плазмида замещения гена была создана следующим образом: шаттл-вектор pWHM3 гидролизовали EcoRI и BamHI. Плазмиду pSE119 гидролизовали BamHI и SphI, гидролизат подвергали электрофорезу и фрагмент длиной 840 п.о. очищали от геля. Плазмиду pSE119 гидролизовали EcoRI и XmnI, гидролизат выделяли электрофорезом и фрагмент длиной 1,7 т.п.о. очищали от геля. Все четыре фрагмента (то есть фрагменты длиной 7,2 т.п.о., 840 п.о., 1,7 т.п.о. и 388 п.о.) лигировали друг с другом методом четырехстороннего лигирования. Лигированную смесь вводили в компетентные клетки DH5 Плазмида pSE185 существенно не изменяла соотношение авермектинов В2:В1 при введении в штамм 1100-SC38 S. avermitilis (таблица 2). Плазмиду pSE185 использовали для трансформации протопластов S. avermitilis с получением мутации со сдвигом рамки считывания в хромосомном гене aveC. Протопласты получали из трансформантов, устойчивых к тиострептону, и культивировали в виде единичных колоний на среде RM14. После регенерации протопластов единичные колонии исследовали на отсутствие устойчивости к тиострептону. Из колоний, чувствительных к тиострептону, выделяли хромосомную ДНК и исследовали при помощи PCR на наличие мутации со сдвигом рамки считывания, встроенной в хромосому. Затравки для PCR, полученные на основе нуклеотидной последовательности aveC, были предоставлены компанией Genosys Biotechnologies, Inc. (Техас). Нижняя затравка для PCR представляла собой: 5′-GCAAGGATACGGGGACTAC-3′ (SEQ ID No.10), верхняя затравка для PCR представляли собой: 5′-GAACCGACCGCCTGATAC-3′ (SEQ ID No.11), и условия выполнения PCR были аналогичны описанным в разделе 7.1.10. Продукт PCR, содержащий 543 п.о., гидролизовали BspE1, в результате чего были получены три фрагмента длиной 368 п.о., 96 п.о. и 79 п.о., что свидетельствовало о встраивании в хромосому инактивированного гена aveC и утрате свободного репликона. Анализ ферментации мутантов S. avermiotilis, содержащих мутацию со сдвигом рамки считывания в гене aveC, показал отсутствие продуцирования нормальных авермектинов, при этом указанные мутанты имели такой же профиль ферментации при анализе ВЭЖХ, что и штаммы SE180-11 и SE184-1-13. Был идентифицирован один трансформант Thios, получивший название штамма SE185-5a. Кроме того, в гене aveC была создана мутация с заменой в положении нуклеотида 520 остатка G остатком А, в результате чего происходит замена кодона, кодирующего триптофан (W), в положении 116 терминирующим кодоном. Штамм S. avermitilis c указанной мутацией не продуцировал нормальные авермектины и имел такой же профиль ферментации, что и штаммы SE180-11, SE184-1-13 и SE185-5a. Кроме того, в гене aveC были созданы мутации, в результате которых просходит замена: (i) остатка G остатком А в положении нуклеотида 970, что вызывает замену глицина (G) аспартатом (D) в положении аминокислоты 266, и (ii) остатка Т остатком С в положении нуклеотида 996, что вызывает замену тирозина (Y) гистидином (Н) в положении аминокислоты 275. Штамм S. avermitilis с указанными мутациями (G266D/Y275H) не продуцировал нормальные авермектины и имел такой же профиль ферментации, что и штаммы SE180-11, SE184-1-13 и SE185-5а. Инактивация гена aveC S. avermitilis в мутантных штаммах SE180-11, SE184-1-13, SE185-5a и других полученных штаммах позволяет оценить влияние других мутаций в гене aveC. Плазмиду pSE186, содержащую копию гена aveC дикого типа, вводили в DM1 E. coli и плазмидную ДНК выделяли из трансформантов, устойчивых к ампициллину. Полученную ДНК pSE186 использовали для трансформации протопластов штамма SE180-11 S. avermitilis. Устойчивые к тиострептону трансформанты штамма SE180-11 выделяли, определяли у них наличие устойчивости к эритромицину и исследовали трансформанты Thior Ermr, анализируя методом ВЭЖХ продукты ферментации. Наличие фуцнкционального гена aveC в трансформантах позволяло восстановить продуцирование нормальных авермектинов в штамме SE180-11 (фигура 5В). 8.2. Анализ мутаций в гене aveC, изменяющих соотношение авермектинов класса В2:В1 Как отмечалось выше, штамм SE180-11 S. avermitilis, содержащий неактивный ген aveC, дополнительно был трансформирован плазмидой, содержащей функциональный ген aveC (pSE186). Штамм SE180-11 также использовали в качестве штамма-хозяина для исследования других мутаций в гене aveC в соответствии с приведенным ниже описанием. Хромосомную ДНК выделяли из штамма 1100-SC38 и использовали в качестве матрицы для амплификации гена aveC при помощи PCR. Открытую рамку считывания длиной 1,2 т.п.о. выделяли путем амплификации при помощи PCR, используя затравки, созданные на основе нуклеотидной последовательности aveC. Нижняя затравка имела SEQ ID No.6, и верхняя затравка имела SEQ ID No.7 (см. приведенный выше раздел 7.1.10). Условия выполнения PCR и субклонирования были аналогичны описанным в разделе 7.1.10. Анализ последовательности ДНК открытой рамки считывания длиной 1,2 т.п.о. показывает наличие мутации в гене aveC, вызывающей замену остатка С остатком Т в положении нуклеотида 337, в результате чего происходит замена серина (S) фенилаланином (F) в положении аминокислоты 55. Ген aveC, содержащий мутацию S55F, субклонировали в векторе pWHM3 с образованием плазмиды, которая получила название pSE187 и была использована для трансформации протопластов штамма SE180-11 S. avermitilis. Устойчивые к тиострептону трансформанты штамма SE180-11 выделяли, определяли наличие устойчивости к эритромицину и исследовали трансформанты Thior Ermr, анализируя методом ВЭЖХ продукты ферментации. Наличие гена aveC, кодирующего замену аминокислотного остатка 55 (S55F), позволяло восстановить продуцирование нормальных авермектинов в штамме SE180-11 (фигура 5С); однако соотношение авермектинов циклогексил В2 : циклогексил В1 составляло примерно 26:1 по сравнению со штаммом SE180-11, трансформированным плазмидой pSE186, в котором соотношение В2:В1 было равно примерно 1,6:1 (таблица 3), свидетельствуя о том, что одна мутация (S55F) модулирует количество продуцируемого авермектина циклогексил В2 по отношению к авермектину циклогексил В1. В гене aveC была идентифицирована другая мутация, вызывающая замену остатка G остатком А в положении нуклеотида 862, в результате чего происходит замена глицина (G) аспартатом (D) в положении аминокислоты 230. Штамм S. avermitilis, имеющий указанную мутацию (G230D), продуцирует авермектины при соотношении В2:В1 около 30:1. 8.3. Мутации, уменьшающие соотношение авермектинов В2:В1 Несколько мутаций, уменьшающий количество продуцируемого авермектина циклогексил В2 по отношению к авермектину циклогексил В1, было получено описанным ниже способом. Была идентифицирована мутация в гене aveC, которая вызывает замену остатка G остатком А в положении нуклеотида 588, в результате чего происходит замена аланина (А) треонином (Т) в положении аминокислоты 139. Ген aveC, содержащий мутацию А139Т, субклонировали в векторе pWHM3 с образованием плазмиды, которая получила название pSE188 и была использована для трансформации протопластов штамма SE180-11 S. avermitilis. Устойчивые к тиострептону трансформанты штамма SE180-11 выделяли, определяли наличие устойчивости к эритромицину и исследовали трансформанты Thior Ermr, анализируя методом ВЭЖХ продукты ферментации. Наличие мутированного гена aveC, кодирующего замену аминокислотного остатка в положении 139 (А139Т), позволяло восстановить продуцирование нормальных авермектинов в штамме SE180-11 (фигура 5D); однако соотношение В2:В1 составляло примерно 0,94:1, показывая, что данная мутация уменьшает количество продуцируемого авермектина циклогексил В2 по отношению к авермектину циклогексил В1. Такой результат был весьма неожиданным, так как опубликованные данные, а также описанные выше результаты мутаций свидетельствовали только об инактивации гена aveC или более высоком продуцировании авермектина В2 по отношению к авермектину В1 (таблица 3). Так как мутация А139Т изменяла соотношение В2:В1 в более благоприятном направлении для авермектина В1, была создана мутация, кодирующая треонин вместо серина в положении аминокислоты 138. Так, плазмиду pSE186 гидролизовали EcoRI и клонировали в pGEM3Zf (Promega), которую гидролизовали EcoRI. Данную плазмиду, которая получила название pSE186a, гидролизовали ApaI и KpnI, фрагменты ДНК разделяли на агарозном геле и два фрагмента длиной 3,8 т.п.о. и 0,4 т.п.о. очищали от геля. ДНК-вставку длиной 1,2 т.п.о., выделенную из плазмиды pSE186, использовали в качестве матрицы для PCR, служащей для замены одного основания в положении нуклеотида 585. Затравки для PCR, предназначенные для введения мутации в положение нуклеотида 585, были предоставлены компанией Genosys Bioterchnologies, Inc. (Техас). Нижняя затравка для PCR представляла собой: 5′-GGGGGCGGGCCCGGGTGCGGAGGCGGAAATGCCCCTGGCGACG-3′ (SEQ ID No.12) и верхняя затравка для PCR представляла собой: 5′-GGAACCGACCGCCTGATACA-3′ (SEQ ID No.13). Реакцию PCR выполняли, используя набор для геномной PCR Advantage GC (Clonetech Laboratories, Palo Alto, CA), в буфере, предоставленном изготовителем, в присутствии смеси 200 мкМ dNTP, 200 пмоль каждой затравки, 50 нг матричной ДНК, 1,0М GC-расплава и 1 единицы полимеразной смеси KlenTaq в конечном объеме 50 мкл. Тепловой профиль первого цикла характеризовался температурой 94°С в течение 1 минуты, затем 25 циклов при температуре 94°С в течение 30 секунд и 68°С в течение 2 минут; и 1 цикл при температуре 68°С в течение 3 минут. Продукт PCR длиной 295 п.о. гидролизовали ApaI и KpnI и выделяли фрагмент длиной 254 п.о., который отделяли электрофорезом и очищали от геля. Все три фрагмента (фрагменты длиной 3,8 т.п.о., 0,4 т.п.о. и 254 п.о.) лигировали друг с другом путем трехстороннего лигирования. Лигированную смесь вводили в компетентные клетки DH5 Плазмиду pSE198 гидролизовали EcoRI, клонирования в векторе pWHM3, который предварительно гидролизовали EcoRI, и вводили в клетки DH5 Другая мутация была создана для введения треонина в положения аминокислот 138 и 139. ДНК-вставку длиной 1,2 т.п.о., выделенную из плазмиды pSE186, использовали в качестве матрицы для PCR. Затравки для PCR, предназначенные для введения мутаций в положения нуклеотидов 585 и 588, были предоставлены компанией Genosys Biotechnologies, Inc. (Техас). Нижняя затравка для pCR представляла собой: 5′-GGGGGCGGGCCCGGGTGCGGAGGCGGAAATGCCGCTGGCGACGACC-3′ (SEQ ID No.14), и верхняя затравка для PCR представляла собой: 5′-GGAACATCACGGCATTCACC-3′ (SEQ ID No.15). Реакцию PCR выполняли в условиях, описанных выше в данном разделе. Продукт PCR длиной 449 п.о. гидролизовали ApaI и KpnI и выделяли фрагмент длиной 254 п.о., который отделяли электрофорезом и очищали от геля. Плазмиду pSE186a гидролизовали ApaI и KpnI, фрагменты ДНК разделяли на агарозном геле и два фрагмента длиной 3,8 т.п.о. и 0,4 т.п.о. очищали от геля. Все три фрагмента (фрагменты длиной 3,8 т.п.о., 0,4 т.п.о. и 254 п.о.) лигировали друг с другом путем трехстороннего лигирования и лигированную смесь вводили в компетентные клетки DH5 Плазмиду pSE230 гидролизовали EcoRI, клонировали в векторе pWHM3, который предварительно гидролизовали EcoRI, и вводили в клетки DH5 Другая мутация была создана для дальнейшего уменьшения количества продуцируемого авермектина циклогексил В2 по отношени к авермектину циклогексил В1. Так как мутации S138T/A139T изменяли соотношение В2:В1 в более благоприятном направлении для авермектина В1, была создана мутация для введения треонина в положение аминокислоты 138 и фенилаланина в положение аминокислоты 139. ДНК-вставку длиной 1,2 т.п.о., выделенную из плазмиды pSE186, использовали в качестве матрицы для PCR. Затравки для PCR, предназначенные для введения мутаций в положения нуклеотидов 585 (замена остатка Т остатком А), 588 (замена остатка G остатком Т) и 589 (замена остатка С остатком Т), были предоставлены компанием Genosys Biotechnologies, Inc. (Техас). Нижняя затравка для PCR представляла собой: 5′-GGGGGCGGGCCCGGGTGCGGAGGCGGAAATGCCGCTGGCGACGTTC-3′ (SEQ ID No.16), и верхняя затравка для PCR представляла собой: 5′-GGAACATCACGGCATTCACC-3′ (SEQ ID No.5). Реакцию PCR выполняли, используя набор для геномной PCR Advantage GC (Clonetech Laboratories, Palo Alto, CA), в буфере, предоставленном изготовителем, в присутствии 200 мкМ dNTP, 200 пмоль каждой затравки, 50 нг матричной ДНК, 1,1 мМ ацетата магния, 1,0 М GC-расплава и 1 единицы ДНК-полимеразы в конечном объеме, равном 50 мкл. Тепловой профиль первого цикла характеризовался температурой 94°С в течение 1 минуты; затем 25 циклов при температуре 94°С в течение 30 секунд и 68°С в течение 2 минут; и 1 цикл при температуре 68°С в течение 3 минут. Продукт PCR длиной 449 п.о. гидролизовали ApaI и KpnI и выделяли фрагмент длиной 254 п.о., который отделяли электрофорезом и очищали от геля. Все три фрагмента (фрагменты длиной 3,8 т.п.о., 0,4 т.п.о. и 254 п.о.) лигировали друг с другом путем трехстороннего лигирования. Лигированную смесь вводили в компетентные клетки DH5 Плазмиду pSE238 гидролизовали EcoRI, клонировани в векторе pWHM3, который предварительно гидролизовали EcoRI, и вводили в клетки DH5
Дополнительные мутации были созданы для дальнейшего уменьшения количества продуцируемого авермектина циклогексил В2 по отношению к авермектину циклогексил В1 методом перестановки ДНК, описанным в публикациях Stemmer, 1994, Nature 370:389-391; и Stemmer, 1994, Proc. Natl. Acad. Sci. USA 91:10747-10751, а также в патентах США №№ 5605793, 5811238, 5830721 и 5837458. Плазмиды с перестановкой ДНК, содержащие мутированные гены aveC, вводили в компетентные клетки dam dcm E. coli. Плазмидную ДНК выделяли из трансформантов, устойчивых к ампициллину, и использовали для трансформации протопластов штамма SE180-11 S. avermitilis. Устойчивые к тиострептону трансформанты штамма SE180-11 выделяли и исследовали на продуцирование авермектинов с соотношением циклогексил В2 : циклогексил В1, равным 1:1 или меньше. Определяли последовательность плазмидной ДНК, выделенной из трансформантов SE180-11, продуцирующих авермектины с соотношением В2:В1, равным 1:1 или меньше. Было идентифицировано восемь трансформантов, продуцирующих меньшие количества авермектина циклогексил В2 по отношению к авермектину циклогексил В1. Наименьшее соотношение В2:В1, достигаемое в указанных трансформантах, составляло 0,40:1 (таблица 4). Плазмидную ДНК выделяли из всех восьми трансформантов и определяли последовательность ДНК для идентификации мутаций в гене aveC. Были обнаружены следующие мутации. Плазмида pSE290 содержит 4 нуклеотидные мутации, включающие замену остатка Т остатком А в положении нуклеотида 317, замену остатка С остатком А в положении нуклеотида 353, замену остатка G остатком А в положении нуклеотида 438 и замену остатка Т остатком А в положении нуклеотида 1155. Замена нуклеотида в положении 317 вызывает замену остатка D остатком Е в положении аминокислоты 48, и замена нуклеотида в положении 438 вызывает замену остатка А остатком Т в положении аминокислоты 89. Соотношение В2:В1, продуцируемое клетками, несущими данную плазмиду, составило 0,42:1 (таблица 4). Плазмида pSE291 содержит 4 нуклеотидные мутации, включающие замену остатка G остатком А в положении нуклеотида 272, замену остатка Т остатков А в положении нуклеотида 585, замену остатка G остатком А в положении нуклеотида 588 и замену остатка G остатком А в положении нуклеотида 708. Замена нуклеотида в положении 585 вызывает замену остатка S остатком Т в положении аминокислоты 138, замена нуклеотида в положении 588 вызывает замену остатка А остатком Т в положении аминокислоты 139, и замена нуклеотида в положении 708 вызывает замену остатка G остатком S в положении аминокислоты 179. Соотношение В2:В1 продуцируемое клетками, несущими указанную плазмиду, составило 0,57:1 (таблица 4). Плазмида pSE292 содержит такие же четыре нуклеотидные мутации, что и плазмида pSE290. Соотношение В2:В1, продуцируемое клетками, несущими указанную плазмиду, составило 0,40:1 (таблица 4). Плазмида pSE293 содержит 6 нуклеотидных мутаций, включающих замену остатка А остатком G в положении нуклеотида 24, замену остатка А остатком С в положении нуклеотида 286, замену остатка Т остатком С в положении нуклеотида 497, замену остатка С остатком Т в положении нуклеотида 554, замену остатка Т остатком С в положении нуклеотида 580 и замену остатка А остатком Т в положении нуклеотида 886. Замена нуклеотида в положении 286 вызывает замену остатка Q остатком Р в положении аминокислоты 38, замена нуклеотида в положении 580 вызывает замену остатка L остатком Р в положении аминокислоты 136, и замена нуклеотида в положении 886 вызывает замену остатка Е остатком D в положении аминокислоты 238. Соотношение В2:В1, продуцируемое клетками, несущими указанную плазмиду, составило 0,68:1 (таблица 4). Плазмида pSE294 содержит 6 нуклеотидных мутаций, включающих замену остатка Т остатком С в положении нуклеотида 469, замену остатка Т остатком А в положении нуклеотида 585, замену остатка G остатком А в положении нуклеотида 588, замену остатка G остатком А в положении нуклеотида 708, замену остатка С остатком Т в положении нуклеотида 833 и замену остатка G остатком А в положении нуклеотида 1184. Кроме того, были удалены нуклеотиды в положениях 173, 174 и 175. Замена нуклеотида в положении 469 вызывает замену остатка F остатком S в положении аминокислоты 99, замена нуклеотида в положении 585 вызывает замену остатка S остатком Т в положении аминокислоты 138, замена нуклеотида в положении 588 вызывает замену остатка А остатком Т в положении аминокислоты 139, и замена нуклеотида в положении 708 вызывает замену остатка G остатком S в положении аминокислоты 179. Соотношение В2:В1, продуцируемое клетками, несущими указанную плазмиду, составило 0,53:1 (таблица 4). Плазмида pSE295 содержит 2 нуклеотидные мутации, включающие замену остатка G остатком А в положении нуклеотида 588 и замену остатка Т остатком С в положении нуклеотида 856. Замена нуклеотида в положении 588 вызывает замену остатка А остатком Т в положении аминокислоты 139, и замена нуклеотида в положении 856 вызывает замену остатка М остатком Т в положении аминокислоты 228. Соотношение В2:В1, продуцируемое клетками, несущими указанную плазмиду, составило 0,80:1 (таблица 4). Плазмида pSE296 содержит 5 нуклеотидных мутаций, включающих замену остатка Т остатком С в положении нуклеотида 155, затему остатка G остатком Т в положении нуклеотида 505, замену остатка С остатком Т в положении нуклеотида 1039, замену остатка С остатком Т в положении нуклеотида 1202 и замену остатка Т остатком С в положении нуклеотида 1210. Замена нуклеотида в положении 505 вызывает замену остатка G остатком V в положении аминокислоты 111, и замена нуклеотида в положении 1039 вызывает замену остатка Р остатком L в положении аминокислоты 289. Соотношение В2:В1, продуцируемое клетками, несущими указанную плазмиду, составило 0,73:1 (таблица 4). Плазмида pSE297 содержит 4 нуклеотидные мутации, включающие замену остатка G остатком Т в положении нуклеотида 377, замену остатка G остатком А в положении нуклеотида 588, замену остатка А остатком G в положении нуклеотида 633 и замену остатка А остатком Т в положении нуклеотида 1067. Замена нуклеотида в положении 588 вызывает замену остатка А остатком Т в положении аминокислоты 139, замена нуклеотида в положении 633 вызывает замену остатка К остатком Е в положении аминокислоты 154, и замена нуклеотида в положении 1067 вызывает замену остатка Q остатком Н в положении аминокислоты 298. Соотношение В2:В1, продуцируемое клетками, несущими указанную плазмиду, составило 0,67:1 (таблица 4).
Описанным ниже способом была выполнена дополнительная серия перестановок ДНК для дальнейшего уменьшения количества продуцируемого авермектина циклогексил В2 по отношению к авермектину циклогексил В1. Полусинтетическая перестановка В лучшем клоне была произведена перестановка способом, описанным в публикации международной заявки на патент РСТ WO 97/20078 компании Maxygen Inc. Отдельные олигонуклеотиды, кодирующие замены, лучше всего соответствующие уменьшению соотношению В2:В1, вводили в реакционную смесь для выполнения реакции перестановки в пятимолярном избытке по отношению к цепям матрицы aveC. Каждое ошибочное спаривание нуклеотидов в олигонуклеотиде было фланкировано 20 полностью идентичными нуклеотидами для обеспечения правильного введения во время реакции перестановки. Олигонуклеотиды были приобретены в компании Operon Technologies (Alameda, CA). Выращивание НТР S. avermitilis Независимые клоны удаляли с трансформационных планшетов и инокулировали в 200 мкл среды R5 (Kieser, T. et al., “Practical Streptomyces Genetics”, 2000, Norwich, U.K., John Innes Foundation) в глубоких 96-луночных планшетах для посева и выращивали, встряхивая, при 30°С. В каждую лунку помещали стеклянный шарик для диспергирования мицелия и перемешивания культуры. В течение указанного времени наблюдался равномерный и плотный рост культур. Через 4-5 дней 20 мкл культуры из среды с посевным материалом переносили на продуцирующие планшеты, к остальному объему добавляли глицерин до конечной концентрации, равной 20%, и замораживали в качестве контрольных планшетов при -80°С. Продуцирующие планшеты инкубировали во влажных условиях при 30°С в течение 12-14 дней. Споруляция штаммов происходила после инкубации в течение 5-8 дней. Продуцирующие планшеты были аналогичны описанным в публикации международной заявки на патент РСТ WO 99/41389 компании Pfizer Inc. за исключением того, чтобы добавляли 1% агарозы для создания твердой поверхности. Экстракция и скрининг методом ESI-MC/MC В каждую лунку добавляли в общей сложности 1 мл этилацетата и инкубировали, встряхивая, при комнатной температуре в течение 20 минут. Было получено примерно 750 мкл этилацетатной фазы, которую переносили на 96-луночный планшет и упаривали в течение ночи. Осадок вновь суспендировали в 100 мкл метанола и 1 мМ NaCl, 5 мкл полученного раствора инъецировали в масс-спектрометр при помощи автоматического пробоотборника в 96-луночном формате и анализировали непосредственно в текучей фазе инъекции без очистки при помощи жидкостной хроматографии или другого метода отделения. Соединения ионизировали методом электрораспылительной ионизации и два отдельных канала контролировали в двух переходах MC/MC. Переход MC/MC для натриевого иона В1 составляет от m/z 921 до m/z 777 и для натриевого иона В2 составляет от m/z 939 до m/z 795 в положительном режиме. Для выполнения данного высокопроизводительного скрининга использовали масс-спектрометр Finnigan TSQ-7000 компании Micromass Quattro-LC и автоматический пробоотборник Twin-Pal компании Leap Technology. Объединение отдельных хроматограмм В1 и В2 для каждой лунки позволило идентифицировать удачные результаты. Было идентифицировано пятьдесят семь (57) новых комбинаций замен аминокислот, которые позволяют получить соотношения авермектинов циклогексил В2 : циклогексил В1, равные 0,35:1 или меньше (фигура 6). Некоторые из указанных новых мутаций позволяют получить соотношения авермектинов циклогексил В2 : циклогексил В1 около 0,30:1 или меньше; некоторые мутации позволяют получить соотношения авермектинов циклогексил В2 : циклогексил В1 около 0,25:1, и некоторые мутации позволяют получить соотношения авермектинов циклогексил В2 : циклогексил В1 около 0,20:1 или меньше. Были идентифицированы две (2) новые мутации, позволяющие продуцировать авермектины циклогексил В2:циклогексил В1 в соотношениях, равных 0,37 или 0,38. Депонирование биологических материалов Указанные ниже плазмиды были депонированы в Американскую коллекцию типовых культур (АТСС) по адресу: 12301 Parklawn Drive, Rockville, MD, 20852, USA, 29 января 1998 г., при этом указанным плаз-мидам были присвоены следующие номера доступа.
Американская коллекция типовых культур в настоящее время расположена по адресу: 108101 University Blvd, Manassas, VA. 20110, USA. Все патенты, заявки на патент и приведенные выше публикации полностью включены в данное описание изобретения. Объем настоящего изобретения не ограничен конкретными вариантами осуществления, рассмотренными в данном описании изобретения, которые служат только для иллюстрации отдельных объектов данного изобретения, и функционально эквивалентные способы и компоненты также входят в объем данного изобретения. Из приведенного выше описания изобретения и прилагаемых чертежей специалисту в данной области должны быть очевидны другие модификации изобретения помимо представленных в данном описании изобретения. Такие модификации входят в объем прилагаемой формулы изобретения.
Формула изобретения
1. Молекула полинуклеотида, содержащая нуклеотидную последовательность, которая, по существу, аналогична последовательности аллеля aveC Streptomyces avermitilis, последовательности плазмиды pSE186 (АТСС 209604), кодирующей продукт гена AveC S. avermitilis, нуклеотидной последовательности открытой рамки считывания (ORF) aveC S. avermitilis, показанной в SEQ ID No.l, или их вырожденному варианту, но при этом нуклеотидная последовательность дополнительно содержит мутации, кодирующие комбинацию замен аминокислот в положении аминокислотных остатков, соответствующих положениям аминокислот SEQ ID No.2, благодаря чему клетки штамма S.avermitilis АТСС 53692, в которых инактивирован аллель aveC дикого типа и которые экспрессируют молекулу полинуклеотида, содержащую мутированную нуклеотидную последовательность, способны продуцировать авермектины класса 2:1 с меньшим соотношением по сравнению с соотношением, продуцируемым клетками штамма S.avermitilis АТСС 53692, экспрессирующими только аллель aveC дикого типа, причем, когда авермектины класса 2:1 являются авермектинами циклогексил В2: циклогексил В1, соотношение авермектинов класса 2:1 составляет 0,35:1 или меньше, где комбинация замен аминокислот выбрана из группы, состоящей из (a) D48E, А61Т, А89Т, S138T, А139Т, G179S, A198G, P289L; (b) G40S, D48E, L136P, G179S, E238D; (с) D48E, L136P, R163Q, G179S; (d) D48E, L136P, R163Q, G179S, E238D; (e) D48E, L136P, R163Q, G179S, A200G, E238D; (f) D48E, L136P, G179S, E238D; (g) D48E, A61T, L136P, G179S, E238D; (h) D48E, A61T, L136P, G179S; (i) D48E, A89T, S138T, A139T, G179S; (j) D48E, A61T, L136P, G179S, A198G, P202S, E238D, P289L; (k) D48E, A61T, L136P, S138T, A139F, G179S, E238D, P289L; (l) D48E, L136P, G179S, A198C, E238D, P289L; (m) D48E, A61T, S138T, A139F, G179S, A198G, P289L; (n) D48E, L84P, G111V, S138T, A139T, G179S, A198G, P289L; (o) Y28C, D48E, A61T, A89T, S138T, A139T, G179S, E238D; (p) D48E, A61T, A107T, S108G, L136P, G179S, S192A, E238D, P289L; (q) D48E, L136P, G179S, R250W; (r) D48E, A89T, S138T, A139T, R163Q, G179S; (s) D48E, L136P, G179S, A198G, P289L; (t) D48E, F78L, A89T, L136P, G179S; (u) D48E, A89T, S138T, A139T, G179S, E238D, F278L; (v) D48E, A89T, L136P, R163Q, G179S; (w) D48E, A61T, A89T, C111V, S138T, A139F, G179S, E238D, P289L; (x) D25G, D48E, А89Т, L136P, S138T, А139Т, V141A, I159T, R163Q, G179S; (у) D48E, А89Т, S90G, L136P, R163Q, G179S, E238D; (z) D48E, А61Т, А89Т, Gil IV, S138T, A139T, G179S, E238D, P289L; (aa) D48E, A89T, S138T, A139T, G179S; (ab) D48E, L136P, R163Q, G179S, S231L; (ac) D48E, L136P, S138T, A139F, G179S, V196A, E238D; (ad) D48E, A61T, A89T, F99S, S138T, A139T, G179S, E238D; (ae) G35S, D48E, A89T, S138T, A139T, G179S, P289L; (af) D48E, A61T, A89T, S138T, A139T, G179S, V196A, E238D; (ag) D48E, A89T, G111V, S138T, A139T, G179S, A198G, E238D; (ah) S41G, D48E, A89T, L136P, G179S; (ai) D48E, A89T, L136P, R163Q, G179S, P252S; (aj) D48E, A89T, L136P, G179S, F234S; (ak) D48E, A89T, L138P, R163Q, G179S, E238D; (al) Q36R, D48E, A89T, L136P, G179S, E238D; (am) D48E, A89T, L136P, R163Q, G179S; (an) D48E, A89T, S138T, G179S; (ao) D48E, A89T, L136P, G179S, E238D; (ap) D48E, A89T, L136P, K154E, G179S, E238D; (aq) D48E, A89T, S138T, A139T, K154R, G179S, V196A, P289L; (ar) D48E, A89T, S138T, A139F, G179S, V196A, E238D; (as) D48E, A61T, A89T, L136P, G179S, V196A, A198G, P289L; (at) D48E, A61T, S138T, A139F, G179S, G196A, E238D, P289L; (au) D48E, A89T, L136P, G179S; (av) D48E, A89T, V120A, L136P, G179S; (aw) D48E, A61T, A89T, S138T, A139F, G179S, V196A, A198G, E238D; (ax) D48E, А61Т, А89Т, G111V, S138T, A139F, G179S, V196A, E238D; (ау) D48E, A61T, A89T, S138T, A139T, G179S, V196A, E238D, P289L; (az) D48E, A61T, A89T, L136P, S138T, A139F, G179S, A198G, E238D; (ba) D48E, A89T, S138T, A139F, G179S, A198G, V220A; (bb) D48E, A61T, A89T, S138T, A139T, G179S, V196A, E238D, R239H, P289L; (be) D48E, A61T, A89T, L136P, G179S, P289L; (bd) D48E, A89T, S138T, A139T, G179S, V196A, E238D, P289L; (be) D48E, A61T, A89T, S138T, A139F, G179S, V196A, E238D; (da) S41G, D48E, A61T, R71L, A89T, L136M, S138T, A139T, T149S, G179S, V196A, E238D, F278L, P289L; (db) S41G, D48E, A61T, R71L, A89T, L136M, S138T, A139T, T149S, F176C, G179S, V196A, E238D, P289L; (dc) D48E, R71L, A89T, L136P, T149S, F176C, G179S, E238D, I280V; (dd) D48E, A61T, R71L, W110L, T149S, G179S, V196A, L206M, E238D, V271A, I280V; (de) D48E, A61T, R71L, A89T, L136M, S138T, A139T, T149S, G179S, V196A, E238D, H279Q, P289L; (df) D48E, A61T, R71L, A89T, L136M, S138T, A139T, T149S, G179S, V196A, E238D, G287E, P289Q; (dg) D48E, A61T, R71L, A89T, L136M, S138T, A139T, Т149S G179S, V196A, E238D, P289L; (dh) D48E, A61T, R71L, A89T, A139T, T149S, F176C, G179S, V196A, E238D, V285G, P289L; (di) Q38R, D48E, А61Т, R71L, L87V, А89Т, L136M, S138T, А139Т, T149S, G179S, V196A, E238D, P289L; (dj) D48E, A61T, L87V, A89T, W11OL, S138T, A139T, T149S, G179S, V196A, E238D, P289L; (dk) D48E, A61T, R71L, A89T, L136M, S138T, A139T, T149S, G179S, V196A, E238D, P289L; (dl) D48E, A89T, L136P, K154E, G179S, S231L, E238D; (ea) D48E, A61T, R71 L, A89T, L136P, T149S, F176C, G179S, V196A, E238D, I280V; (eb) D48E, R71L, A89T, L136P, T149S, F176C, G179S, E238D, I280V; (ec) Q36P, D48E, A61T, R71L, A89T, L136P, T149S, F176C, G179S, V196A, E238D, I280V; (ed) D48E, A61T, R71L, A89T, A139T, T149S, F176C, G179S, V196A, E238D, I280V; (ee) V2M, D48E, A61T, R71L, A89T, L136P, T149S, F176C, G179S, V196A, E238D, I280V; (ef) D48E, A61T, R71L, A89T, L136P, T149S, F176C, G179S, V196A, E238D, I280V, A302T; (eg) D48E, R71L, A89T, L136P, T149S, F176C, G179S, E238D, P289L; (eh) D48E, R71L, A89T, L136P, T149S, F176C, G179S, E238D, A302T; (ei) D48E, R71L, A89T, L136Р, T149S, F176C, G179S, V196A, E238D, I280V; (ej) D48E, A61T, R71L, A89T, L136P, T149S, F176C, G179S, V196A, E238D; (ek) V2M, D48E, R71L, A89T, L136P, T149S, F176C, G179S, V196A, E238D, I280V; (el) D48E, A61T, R71L, A89T, L136P, T149S, R162H, F176C, G179S, V196A, E238D, I280V; (em) D48E, R71L, A89T, V120A, L136P, T149S, K154E, G179S, S231L, E238D; (en) D48E, R71L, A89T, V120A, L136P, T149S, F176C, G179S, S231L, E238D, I280V; (eo) D48E, A61T, R71L, L87V, A89T, A139T, T149S, F176C, G179S, V196A, E238D, V285G, P289L; (ep) D48E, A61T, R71L, L87V, A89T, S90N, A139T, T149S, F176C, G179S, V196A, E238D, V285G, P289L; (eq) D48E, R71L, A89T, L136P, K154E, G179S, S231L, E238D; (er) D48E, R71L, A89T, V120A, L136P, K154E, F176C, G179S, S231L, E238D; (es) D48E, R71L, A89T, V120A, L136P, T149S, K154E, F176C, G179S, S231L, E238D. 2. Молекула полинуклеотида по п.1, где комбинация замен аминокислот содержит комбинацию группы (еа): D48E, A61T, R71L, A89T, L136P, T149S, F176C, G179S, V196A, E238D, I280V. 3. Молекула полинуклеотида по любому из пп.1 и 2, где соотношение авермектинов циклогексил В2:циклогексил В1 составляет примерно 0,20:1 или меньше. 4. Молекула полинуклеотида по любому из пп.1 и 2, где соотношение авермектинов циклогексил В2:циклогексил В1 составляет примерно 0,10:1 или меньше. 5. Рекомбинантный вектор, предназначенный для продуцирования авермектинов класса 2:1 в уменьшенном соотношении, содержащий молекулу полинуклеотида по любому из пп.1-4. 6. Клетка-хозяин, предназначенная для продуцирования авермектинов класса 2:1 в уменьшенном соотношении, содержащая рекомбинантный вектор по п.5. 7. Клетка-хозяин по п.6, которая является клеткой рода Streptomyces. 8. Клетка-хозяин по п.7, которая является клеткой Streptomyces avermitilis. 9. Способ получения мутированного штамма Streptomyces avermitilis, включающий (i) мутирование аллеля aveC в клетке штамма S. avermitilis, вызывающее комбинацию замен аминокислот в продукте гена AveC, или (ii) введение в клетку штамма S. avermitilis мутированного аллеля aveC или его вырожденного варианта, кодирующего продукт гена AveC, содержащий комбинацию замен аминокислот, где клетки, содержащие мутированный аллель aveC или его вырожденный вариант, кодирующий продукт гена AveC, способны продуцировать авермектины циклогексил В2:циклогексил В1 в соотношении 0,35:1 или меньше, при этом комбинация замен аминокислот выбрана из группы, приведенной в п.1. 10. Способ по п.9, в котором комбинация замен аминокислот содержит комбинацию группы (еа): D48E, А61Т, R71L, А89Т, L136P, T149S, F176C, G179S, V196A, E238D, I280V. 11. Способ по п.10, в котором соотношение авермектинов циклогексил В2: циклогексил В1 составляет примерно 0,20:1 или меньше. 12. Способ по п.10, в котором соотношение авермектинов циклогексил В2: циклогексил В1 составляет примерно 0,10:1 или меньше. 13. Клетка рода Streptomyces, предназначенная для продуцирования авермектинов класса 2:1 в уменьшенном соотношении, которая содержит мутированный аллель aveC S.avermitilis или его вырожденный вариант, кодирующий продукт гена AveC, включающий комбинацию замен аминокислот, выбранную из группы, приведенной в п.1. 14. Клетка по п.13, в которой комбинация замен аминокислот содержит комбинацию группы (еа): D48E, А61Т, R71L, А89Т, L136P, T149S, F176C, G179S, V196A, E238D, I280V. 15. Клетка по любому из пп.13 и 14, которая является клеткой Streptomyces avermitilis. 16. Способ получения авермектинов класса 2:1 в уменьшенном соотношении, включающий культивирование штамма клеток Streptomyces avermitilis по п.15 в культуральной среде в условиях, позволяющих или индуцирующих продуцирование им авермектинов, и выделение указанных авермектинов из культуры. 17. Композиция авермектинов циклогексил В2:циклогексил В1, обладающая противопаразитарными и пестицидными свойствами, представляющая собой культуральную среду после культивирования клеток Streptomyces avermitilis, полученных по любому из пп.9-12, содержащую авермектины циклогексил В2: циклогексил В1 в соотношении 0,35:1 или меньше.
РИСУНКИ
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



 -gal, промотор Т7, промотор ТАС, верхний и нижний
-gal, промотор Т7, промотор ТАС, верхний и нижний  -промоторы, промоторы trp и lac, слитые промоторы trp-lac и специально для Streptomyces промоторы ermE, melC, tipA и т.д. В соответствии с конкретным вариантом осуществления изобретения может быть создан экспрессирующий вектор, содержащий открытую рамку считывания aveC или мутированную открытую рамку считывания, клонированную рядом с сильным структурным промотором, таким как промотор ermE, выделенный из Saccharopolyspora erythraea. Как описано в патенте США № 6248579, вектор, содержащий промотор ermE, был введен в штамм S. avermitilis, и результаты последующего анализа продуктов ферментации методом ВЭЖХ свидетельствовали об увеличении титра продуцированных авермектинов по сравнению с количеством, продуцированным тем же штаммом, экспрессирующим только аллель aveC дикого типа.
-промоторы, промоторы trp и lac, слитые промоторы trp-lac и специально для Streptomyces промоторы ermE, melC, tipA и т.д. В соответствии с конкретным вариантом осуществления изобретения может быть создан экспрессирующий вектор, содержащий открытую рамку считывания aveC или мутированную открытую рамку считывания, клонированную рядом с сильным структурным промотором, таким как промотор ermE, выделенный из Saccharopolyspora erythraea. Как описано в патенте США № 6248579, вектор, содержащий промотор ermE, был введен в штамм S. avermitilis, и результаты последующего анализа продуктов ферментации методом ВЭЖХ свидетельствовали об увеличении титра продуцированных авермектинов по сравнению с количеством, продуцированным тем же штаммом, экспрессирующим только аллель aveC дикого типа. (International Biotechnologies Inc.), являющийся гидрофильным маркерным пептидом, может быть встроена стандартными методами в экспрессирующий вектор на участке, соответствующем, например, карбоксильному концу полипептида гена AveC. Экспрессированный продукт слияния полипептида AveC и эпитопа FLAG
(International Biotechnologies Inc.), являющийся гидрофильным маркерным пептидом, может быть встроена стандартными методами в экспрессирующий вектор на участке, соответствующем, например, карбоксильному концу полипептида гена AveC. Экспрессированный продукт слияния полипептида AveC и эпитопа FLAG -фактора, иммуноглобулинов, белков наружных оболочки, пенициллиназы и рецепторов Т-клеток.
-фактора, иммуноглобулинов, белков наружных оболочки, пенициллиназы и рецепторов Т-клеток.