Патент на изобретение №2155811
|
||||||||||||||||||||||||||
(54) ГЕН, КОДИРУЮЩИЙ КАРМИНОМИЦИН-4-0-МЕТИЛТРАНСФЕРАЗУ, ВЕКТОР, ХОЗЯЙСКАЯ КЛЕТКА И СПОСОБ ПОЛУЧЕНИЯ ДАУНОРУБИЦИНА
(57) Реферат: Изобретение относится к биотехнологии. Получена нуклеотидная последовательность фрагмента ДНК, кодирующего карминомицин – 4-0-метилтрансферазу. Сконструированы плазмиды pWHM901, pWHM902 и pWHM903, содержащие указанную последовательность. Получен рекомбинантный штамм S.Lividans ТК 24/pWHM902 и рекомбинантный штамм Е.соli pGP1 – 2/ pWHM903 – продуценты карминомицин-4-0-метилтрансферазы. В способе получения даунорубцина культивируют рекомбинантные штаммы, продуцирующие карминомицин-4-0-метилтрансферазу в среде, содержащей карминомицин с последующим выделением целевого продукта. Изобретение позволяет получить даунорубцин путем биоконверсии карминомицина в даунорубцин, применяемый в противораковой терапии. 7 с.п.ф-лы, 4 ил. Изобретение относится к гену, кодирующему карминомицин-4-О-метилтрансферазу, вектору, хозяйской клетке и способу получения даунорубицина рекомбинантным путем. Предшествующий уровень техники Антрациклины группы даунорубицинов, такие, как доксорубицин, карминомицин и аклациномицин, являются наиболее широко используемыми лекарственными средствами, применяемыми в противораковой терапии [F. Arcamone, Doxorubicin, Academic Press, New Jork, 1981, pp. 12-25; A. Grein, Process Biochem. 16:34, 1981; T. Kaneko, Chimicaoggi May: 11 (1988)]. Улучшенные производные даунорубицина и доксорубицина были получены путем химического синтеза в целях усиления их противоопухолевой активности, особенно, при пероральном способе введения, а также в целях устранения острой токсичности и хронической кардиотоксичности, связанной с использованием этих лекарственных средств при лечении рака [Penco, Process Biochem. 15:12 (1980); T. Kaneko, Chimicaggi May: 11 (1988)]. Примерами таких аналогов являются 4′-эпидоксиорубицин (Epirubicin  ) и 4-деметоксидаунорубицин (Idarubicin ).
Указанные природные соединения продуцируются различными штаммами стрептомицетов (S. peucetius, S. coeruleorubidus, S. griseus, S. griseoruber, S. insignis, S. viridochromogenes, S. bifurcus и streptomyces sp. штамм C5), а также Actinmyces carminata.
Доксорубицин продуцируется только S. peucetius подвидом caesius, а даунорубицин продуцируется как S. peucetius так и другими штаммами стрептомицетов, указанными выше. Штаммы S. peucetius подвид caesins 1MRU 3920 (этот штамм идентичен ATCC 27952, и далее он будет сокращенно обозначаться “S. peucetius ATCC 3920), S. peucetius ATCC 29050 (“S. pencetius 29050), и S. pencetius подвид caesins ATCC 27952 (“S. pencetius 27952) являются известными и описаны в USA-359028. S. pencetius 29050 и 27952 были депонированы в Американской коллекции типовых культур, Роквилл, MD, США, под номерами допуска ATCC 29050 и 27952.
Антрациклин доксорубицин (2) продуцируется S. pencetius 27952 из малоновой кислоты, пропионовой кислоты и глюкозы в соответствии с путем метаболизма, показанном на фиг. 1 /см. рисунки, прилагаемые в конце описания/. Как видно из этого чертежа, ) и 4-деметоксидаунорубицин (Idarubicin ).
Указанные природные соединения продуцируются различными штаммами стрептомицетов (S. peucetius, S. coeruleorubidus, S. griseus, S. griseoruber, S. insignis, S. viridochromogenes, S. bifurcus и streptomyces sp. штамм C5), а также Actinmyces carminata.
Доксорубицин продуцируется только S. peucetius подвидом caesius, а даунорубицин продуцируется как S. peucetius так и другими штаммами стрептомицетов, указанными выше. Штаммы S. peucetius подвид caesins 1MRU 3920 (этот штамм идентичен ATCC 27952, и далее он будет сокращенно обозначаться “S. peucetius ATCC 3920), S. peucetius ATCC 29050 (“S. pencetius 29050), и S. pencetius подвид caesins ATCC 27952 (“S. pencetius 27952) являются известными и описаны в USA-359028. S. pencetius 29050 и 27952 были депонированы в Американской коллекции типовых культур, Роквилл, MD, США, под номерами допуска ATCC 29050 и 27952.
Антрациклин доксорубицин (2) продуцируется S. pencetius 27952 из малоновой кислоты, пропионовой кислоты и глюкозы в соответствии с путем метаболизма, показанном на фиг. 1 /см. рисунки, прилагаемые в конце описания/. Как видно из этого чертежа,  – родомицинон (4), карминомицин (3) и даунорубицин (1) являются промежуточными соединениями в данном процессе метаболизма [Grein Advan. Appl. Microbiol. 32:203 (1987), Eckardt & Wagner J. basic Microbiol. 28:137 (1988)].
Две стадии указанного пути метаболизма включают в себя О-метилирование дискретных промежуточных соединений, а именно: превращение акланоновой кислоты в метилакланонат, и превращение карминомицина /3/ в даунорубицин /1/. Было показано, что бесклеточные экстракты S. pencetins 29050, S. insignic ATCC 31913, S. coeruleorubidus ATCC 31276, и Streptomyces sp. C5 катализуют последнюю из двух вышеуказанных стадий в присутствии S-аденозил-L-метионина [Connors и др., J. Gen. Microbiol. 136:1895 (1990)], что позволяет предположить, что все указанные штаммы содержат специфическую карминомицин-4-О-метилтрансферазу (COMT-белок).
Гены, кодирующие биосинтез даунорубицина и резистентность к даунорубицину, были получены из S. pencetins 29050 и S. pencetins 27952 путем экспериментального клонирования [Stutzman-Engwall и Hutchinson, Proc. Natl. Acad. Sci. USA 86: 3135 (1988); Otten и др., J. Bacteriol. 172-3427 (1990)].
Эти исследования показали, что при введении в Streptomyces lividans 1326, указанные клонированные гены приобретают способность к продуцированию -родомицинона и резистентность к даунорубицину и доксорубицину для этого хозяина. В последующей работе, мы провели исследования с целью проверки, могут ли указанные клоны приобретать способность к превращению карминомицина в даунорубицин при введении их в S. lividans. В этой работе был выделен ДНК-сегмент в 1,6 тысяч оснований (kb), который вводит ген карминомицин-4-О-метилтрансферазы, сокращенно обозначаемый далее “dnrK”.
Краткое описание изобретения – родомицинон (4), карминомицин (3) и даунорубицин (1) являются промежуточными соединениями в данном процессе метаболизма [Grein Advan. Appl. Microbiol. 32:203 (1987), Eckardt & Wagner J. basic Microbiol. 28:137 (1988)].
Две стадии указанного пути метаболизма включают в себя О-метилирование дискретных промежуточных соединений, а именно: превращение акланоновой кислоты в метилакланонат, и превращение карминомицина /3/ в даунорубицин /1/. Было показано, что бесклеточные экстракты S. pencetins 29050, S. insignic ATCC 31913, S. coeruleorubidus ATCC 31276, и Streptomyces sp. C5 катализуют последнюю из двух вышеуказанных стадий в присутствии S-аденозил-L-метионина [Connors и др., J. Gen. Microbiol. 136:1895 (1990)], что позволяет предположить, что все указанные штаммы содержат специфическую карминомицин-4-О-метилтрансферазу (COMT-белок).
Гены, кодирующие биосинтез даунорубицина и резистентность к даунорубицину, были получены из S. pencetins 29050 и S. pencetins 27952 путем экспериментального клонирования [Stutzman-Engwall и Hutchinson, Proc. Natl. Acad. Sci. USA 86: 3135 (1988); Otten и др., J. Bacteriol. 172-3427 (1990)].
Эти исследования показали, что при введении в Streptomyces lividans 1326, указанные клонированные гены приобретают способность к продуцированию -родомицинона и резистентность к даунорубицину и доксорубицину для этого хозяина. В последующей работе, мы провели исследования с целью проверки, могут ли указанные клоны приобретать способность к превращению карминомицина в даунорубицин при введении их в S. lividans. В этой работе был выделен ДНК-сегмент в 1,6 тысяч оснований (kb), который вводит ген карминомицин-4-О-метилтрансферазы, сокращенно обозначаемый далее “dnrK”.
Краткое описание изобретенияНастоящее изобретение относится к ДНК, расположение сайтов рестрикции которой показано на фиг 2 /см. приложение/, или к фрагменту, происходящему от указанной ДНК, и содержащему ген dnrK, кодирующий карминомицин-4-О-метилтрансферазу. Для удобства, показанный на фиг 2 ДНК-сегмент далее будет именоваться “ДНК-вставкой”, а ДНК-последовательность указанного гена dnrK представлена на фиг 3 /см. приложение/. Кроме того, настоящее изобретение относится: /1/ к рекомбинантным векторам, которые обладают способностью к трансформации клетки-хозяина, и содержат ДНК-вставку или происходящий от нее рестрикционный фрагмент, содержащий ген dnrK; /2/ к рекомбинантным векторам, которые обладают способностью повышать число копий гена dnrK и количество его продукта в штамме Streptomyces spp., продуцирующем даунорубицин. Краткое описание чертежей На фиг. 1 схематически проиллюстрирован биосинтетический путь метаболизма доксорубицина. На фиг. 2 проиллюстрирован рестрикционный анализ путем картирования первой ДНК настоящего изобретения. Эта ДНК представляет собой вставку в рекомбинантной плазмиде pWHM 902, сконструированной путем инсерции SphI/PvuII – /1,6 kb/ фрагмента ДНК, содержащего ген карминомицин-4-О-трансферазы /dnrK/, который был получен из рекомбинантной плазмиды pWHM 901 путем переваривания ферментами SphI и PvuII, в SphI/SmaI-сайты плазмиды pWHM 3, представляющей собой E. coli-Streptomyces-челночный вектор [Vara и др., J. Bacteriol 171: 5872 (1989)]. Карта, показанная на фиг. 2, не должна рассматриваться как исчерпывающая иллюстрация всех рестрикционных сайтов, присутствующих в ДНК-сегменте. Однако сайты, показанные на фиг.2, являются достаточными для точного распознавания сегментов. На фиг. 3 схематически проиллюстрирована нуклеотидная последовательность ДНК-сегмента dnrK, кодирующего карминомицин-4-О-трансферазу. Этот сегмент соответствует области, расположенной между SphI- и PvuII-сайтами рестрикции плазмиды pWHM 902 и представляет собой кодирующую нить в направлении 5′—>3′. Выведенная аминокислотная последовательность транслированной открытой рамки считывания, кодирующей карминомицин-4-О-метилтрансферазу, показана после нуклеотидной последовательности гена dnrK (SEQ 1D N 1, SEQ 1D N 2). На фиг. 4 проиллюстрировано рестрикционное картирование второй ДНК настоящего изобретения. Эта ДНК представляет собой вставку в рекомбинантной плазмиде pWHM 903, сконструированной путем инсерции NdeI/EcoR- (1,4 kb) фрагмента ДНК, полученного из 5,8 kb – Sphl-фрагмента ДНК плазмиды pWHM 901 с помощью сайт-направленного мутагенеза, в NdeI- и EcoRI-сайты плазмидного вектора экспрессии pT7-7 E. coli [Tabor и Richardson, Proc. Natl. Acad. Sci. USA 82:1074 (1986)]. Карта, показанная на фиг. 4, не должна рассматриваться как исчерпывающая информация относительно всех рестрикционных сайтов, присутствующих в данном ДНК-сегменте. Однако показанные на фиг.4 сайты являются достаточными для однозначного распознавания сегментов. Подробное описание изобретения ДНК-вставка и рестрикционные фрагменты настоящего изобретения содержат ген (dnrK), кодирующий карминомицин-4-О-метилтрансферазу. Для данного экспрессируемого гена ДНК может иметь свою собственную последовательность, регулирующую транскрипцию, а в частности, свой собственный промотор, который соответствующим образом соединен с данным геном и который распознается РНК-плимеразой хозяйской клетки. Альтернативно, ДНК-вставка или рестрикционный фрагмент могут быть лигированы соответствующим образом с другой регулирующей транскрипцию последовательностью, либо они могут быть клонированы в вектор в соответствующем рестрикционном сайте, надлежащим образом расположенном возле регулирующей транскрипцию последовательности в указанном векторе. ДНК-вставка или рестрикционный фрагмент, несущие ген карминомицин-4-О-метилтрансферазы, могут быть клонированы в вектор для клонирования рекомбинантных ДНК. Для этого может быть использован любой автономно реплицирующийся и/или интергирующий агент, содержащий ДНК-молекулу, к которой могут быть добавлены один или несколько дополнительных ДНК-сегментов. Однако, обычно, в качестве такого вектора используется плазмида. Предпочтительной плазмидой является многокопийная плазмида pWHM 3 или pIJ702 [Katz и др. J. Gen. Microbiol. 129:2703 (1983)]. Другими подходящими плазмидами являются pIJ385 [Mayeri и др., J. Bacteriol. 172:6061 (1990)], pIJ680 [Hopwood и др., Genetic Manipulation of Streptomyces. A Laboratory Manual, John Innes Foundation, Norwich, UK, 1985], pWHM 601 [Guilfoile и Hutchinson, Proc. Natl. Acad. Sci. USA 88:8553 (1991)] или pPM 927 [Smokina и др., Gene 94:52 (1990)]. Для введения ДНК-вставки или рестрикционного фрагмента в вектор может быть использована любая подходящая техника. Указанная инсерция может быть осуществлена путем лигирования ДНК в линеаризованный вектор в соответствующем сайте рестрикции. Для этих целей может быть осуществлено прямое присоединение “липких” или “тупых” концов, присоединение гомополимера, либо могут быть использованы линкерная или адаптерная молекулы. Рекомбинантный вектор используется для трансформации соответствующей клетки-хозяина. Эти клетки-хозяева могут быть карминомицин- или даунорубицин-восприимчивыми, то есть, они не могут расти в присутствии определенного количества карминомицина или даунорубицина; либо эти клетки могут быть карминомицин- или даунорубицин-резистентными. Таким хозяином может быть микроорганизм. Поэтому могут быть трансформированы штаммы S. peucetius, в частности, S. peucetius 29050, и другие штаммы видов Streptomyces, которые продуцируют, либо не продуцируют антрациклины. Трансформанты штаммов стрептомицетов обычно получают путем трансформации протопластов. Ген dnrK может быть также введен в другие векторы и экспрессирован в бактериях, не относящихся к стрептомицетам, например в E. coli COMT-белок, полученный с помощью трансформированного хозяина, может быть использован для биоконверсии карминомицина в даунорубицин. Этот способ дает возможность получать даунорубицин высокой чистоты из клеточного экстракта, продуцированного путем ферментации и содержащего помимо даунорубицина нежелательный побочный продукт в виде карминомицина. Процесс биоконверсии может быть осуществлен либо путем непосредственного использования свободных или иммобилизованных трансформированных клеток, либо путем выделения COMT-белка, который может быть использован в свободной форме, иммобилизован стандартными способами на полимере, стекле, целлюлозе или на аналогичных субстратах посредством ионной или ковалентной связи, или привит к волокнам, проницаемым для субстрата, либо он может быть иммобилизован посредством перекрестного сшивания, COMT-белок может быть также использован в неочищенном клеточном экстракте. Рекомбинантный вектор настоящего изобретения может быть также использован для трансформации соответствующей хозяйской клетки, которая продуцирует даунорубицин, в целях усиления биоконверсии карминомицина и минимизации количества нежелательного промежуточного соединения, присутствующего в конечном клеточном экстракте. Эти хозяйские клетки могут быть карминомицин-даунорубицин- или доксорубицин-резистентными, т.е., они могут расти в присутствии любого количества карминомицина, даунорубицина или доксорубицина. Поэтому могут быть трансформированы штаммы S. peucetius, в частности S. peucetius 29050, и другие штаммы стрептомицетов, продуцирующие антрациклины. Трансформанты штаммов стрептомицетов обычно получают путем трансформации протопластов. Даунорубицин может быть получен путем культивирования трансформированного штамма S. peucetius или других видов Streptomyces, которые не содержат гена dnrK с последующим выделением продуцированного таким образом даунорубицина или родственных антрациклинов. ДНК-вставки получают из геномной ДНК S. peucetius 29050. Этот штамм был депонирован в Американской коллекции типовых культур [Роквилл, Мэриленд (MD), США] под номером ATCC 29050. Для этих целей может быть также использован штамм S. peucetius 27952, который происходит от S. peucetius 29050, и который также способен превращать карминомицин в даунорубицин. ДНК-вставка может быть получена путем: /a/ получения библиотеки геномной ДНК штамма S. peucetius 29050 или происходящего от него штамма; /b/ скрининга полученной библиотеки на наличие в ней клонов, способных превращать карминомицин в даунорубицин; /c/ выделения ДНК-вставки из рекомбинантного вектора, который является частью библиотеки и который дал положительный результат при скрининге на способность к превращению карминомицина в даунорубицин; и /d/ необязательного выделения из указанной ДНК-вставки рестрикционного фрагмента, содержащего ген, который кодирует карминомицин-4-О-метилтрансферазу. Указанная в стадии /a/ библиотека может быть получена путем частичного переваривания геномной ДНК штамма S. peucetius 29050 или происходящего от него штамма. Для этого предпочтительно использовать рестриктирующий фермент Mbol. Полученные таким образом ДНК-фрагменты могут быть подвергнуты фракционированию по размерам; причем, предпочтительными являются размеры от 3 до 5 kb. Эти фрагменты лигируют в линеаризованный вектор, такой как pWHM3 или pIJ702. Хозяйские клетки трансформируют лигирующей смесью. Обычно хозяйские клетки могут не продуцировать карминомицин или даунорубицин и могут быть карминомицин- или даунорубицин-восприимчивыми; например, они могут быть восприимчивы к 10 микрограммам или менее карминомицина или дауномицина на мл. Например, могут быть трансформированы протопласты штамма S. lividans JI623 (Hopwood и др., Genetic Manipulation of Streptomyces. A Laboratory Manual, John Innes Foundation, Norwich, UK, 1985). В стадии /b/ полученные трансформанты скринируют на способность превращать карминомицин в даунорубицин и секретировать даунорубицин. Клоны, способные превращать карминомицин в даунорубицин, идентифицируют с помощью хроматографического анализа экстрактов культуральной среды, содержащей карминомицин, проводимого в целях обнаружения даунорубицина в указанных экстрактах. Эти клоны выделяют, а содержащиеся в них рекомбинантные векторы экстрагируют. После переваривания рекомбинантных векторов соответствующими рестриктирующими ферментами в стадии /c/, ДНК S. peucetius 29050, инсерцированная в каждый вектор, может быть идентифицирована с определением ее размера и картирована. Таким образом, может быть установлено, что данный вектор содержит ДНК-вставку настоящего изобретения. Кроме того, могут быть выделены две или несколько перекрывающихся вставок, которые полностью или частично включены в ДНК настоящего изобретения. Эти вставки могут быть объединены вместе путем расщепления в их общем рестрикционном сайте с последующим лигированием в целях получения ДНК настоящего изобретения, которую, если это необходимо, урезают по длине соответствующими рестриктирующими ферментами. Рестрикционные фрагменты ДНК-вставки, которые содержат ген, кодирующий COMT-белок, могут быть получены в стадии /d/ также путем расщепления ДНК-вставки соответствующим рестриктирующим ферментом. ДНК настоящего изобретения может быть мутирована с использованием методов, не влияющих на ее способность сообщать признак, ответственный за превращение карминомицина в даунорубицин. Это может быть достигнуто, например, с помощью сайт-специфичного мутагенеза. Мутированная таким образом ДНК является частью настоящего изобретения. ДНК настоящего изобретения может быть также введена в векторы, подходящие для экспрессии гена dnrK в хозяине, не относящемся к стрептомицетам, например, в E. coli. Представленные ниже примеры иллюстрируют настоящее изобретение. Материалы и методы Бактериальные штаммы и плазмиды: Штамм E. coli DH5  , который является восприимчивым к ампициллину и апрамицину, был использован для субклонирования ДНК-фрагментов, а штамм E. coli K38 (Russel & Model, J. Bacteriol. 159: 1034 (1984) был использован для экспрессии гена dnrK S. peucetius. Для получения однонитевой ДНК для секвенирования dnrK гена был использован штамм E. coli JM105.
Среды и буферы: E. coli DH5a выдерживали на LB-агаре (Sambrook и др., Molecular Cloning. A Laboratory Manual 2-ое изд. Cold Spring Harbor-Press, Cold Spring Harbor, NY, 1989). При селекции на присутствие трансформантов добавляли ампициллин или апрамицин в концентрациях 50 мкг/мл и 100 мкг/мл, соответственно. E. coli JM105 выдерживали на агаровой минимальной среде M9 (Sambrook и др. , Molecular Cloning. A Laboratory Manual 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989), а колонии переносили на LB-среду и в течение ночи культивировали при 37oC в целях получения бактерий, используемых для продуцирования однонитевой ДНК. H-агар (Sambrook и др. , Molecular Cloning. A Laboratory Manual 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989) использовали для культивирования E. coli DH5a, трансформированных репликативной формой ДНК M3 [(Yansch-Perron и др., Gene 33: 103 (1985)]. S. lividans выдерживали на агаре R2YE (Hopwood и др., Genetic Manupulation of Streptomyces. A Laboratory Manual, John Innes Foundation, Norwich. UK, 1985) для получения спор, а также для регенерации протопластов.
Субклонирование ДНК-фрагментов: ДНК-образцы переваривали соответствующими рестриктирующими ферментами и подвергали разделению на агарозном геле стандартными методами (Sambrook и др., Molecular Cloning. A Laboratory Manual, 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989). Агарозные срезы, содержащие нужные ДНК-фрагменты, вырезали из геля, и из этих срезов, используя устройство GENECLEAN (Bio 101, La Jolla, CA), выделяли ДНК. С помощью традиционной техники (Sambrook и др., Molecular Cloning. A Laboratory Manual, 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989), выделенные ДНК-фрагменты субклонировали в E. coli и E. coli/Streptomyces челночные векторы для экспериментальной биотрансформации и экспрессии, соответственно, и в векторы M13 [(Yansch-Perron и др., Gene 33: 103 (1985)] для секвенирования.
Секвенирование ДНК: После субклонирования нужных ДНК-фрагментов в вектор M13, получали однонитевую ДНК с использованием стандартной техники (Sambrook и др., Molecular Cloning. A Laboratory Manual, 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989) и использовали при секвенировании. Данные о ДНК-последовательности были получены с использованием набора для секвенирования с секвеназой версии 2.0 (US Biochemicals, Cleveland, OH) в соответствии с инструкцией изготовителей. Во избежание компрессии, вместо dGTP использовали 7-Deaza dGTP. Сначала, для получения последовательности первых 200-250 оснований использовали универсальный праймер вектора M13, а затем из этой последовательности, используя ДНК-синтезатор Applied Biosystems 391 в соответствии с инструкцией изготовителей, синтезировали 17-мерный олигонуклеотид, который использовали в качестве праймера для дальнейшего секвенирования ДНК до тех пор, пока не будут получены данные о полной ДНК-последовательности.
Трансформация видов Streptomyces и E. coli: , который является восприимчивым к ампициллину и апрамицину, был использован для субклонирования ДНК-фрагментов, а штамм E. coli K38 (Russel & Model, J. Bacteriol. 159: 1034 (1984) был использован для экспрессии гена dnrK S. peucetius. Для получения однонитевой ДНК для секвенирования dnrK гена был использован штамм E. coli JM105.
Среды и буферы: E. coli DH5a выдерживали на LB-агаре (Sambrook и др., Molecular Cloning. A Laboratory Manual 2-ое изд. Cold Spring Harbor-Press, Cold Spring Harbor, NY, 1989). При селекции на присутствие трансформантов добавляли ампициллин или апрамицин в концентрациях 50 мкг/мл и 100 мкг/мл, соответственно. E. coli JM105 выдерживали на агаровой минимальной среде M9 (Sambrook и др. , Molecular Cloning. A Laboratory Manual 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989), а колонии переносили на LB-среду и в течение ночи культивировали при 37oC в целях получения бактерий, используемых для продуцирования однонитевой ДНК. H-агар (Sambrook и др. , Molecular Cloning. A Laboratory Manual 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989) использовали для культивирования E. coli DH5a, трансформированных репликативной формой ДНК M3 [(Yansch-Perron и др., Gene 33: 103 (1985)]. S. lividans выдерживали на агаре R2YE (Hopwood и др., Genetic Manupulation of Streptomyces. A Laboratory Manual, John Innes Foundation, Norwich. UK, 1985) для получения спор, а также для регенерации протопластов.
Субклонирование ДНК-фрагментов: ДНК-образцы переваривали соответствующими рестриктирующими ферментами и подвергали разделению на агарозном геле стандартными методами (Sambrook и др., Molecular Cloning. A Laboratory Manual, 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989). Агарозные срезы, содержащие нужные ДНК-фрагменты, вырезали из геля, и из этих срезов, используя устройство GENECLEAN (Bio 101, La Jolla, CA), выделяли ДНК. С помощью традиционной техники (Sambrook и др., Molecular Cloning. A Laboratory Manual, 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989), выделенные ДНК-фрагменты субклонировали в E. coli и E. coli/Streptomyces челночные векторы для экспериментальной биотрансформации и экспрессии, соответственно, и в векторы M13 [(Yansch-Perron и др., Gene 33: 103 (1985)] для секвенирования.
Секвенирование ДНК: После субклонирования нужных ДНК-фрагментов в вектор M13, получали однонитевую ДНК с использованием стандартной техники (Sambrook и др., Molecular Cloning. A Laboratory Manual, 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989) и использовали при секвенировании. Данные о ДНК-последовательности были получены с использованием набора для секвенирования с секвеназой версии 2.0 (US Biochemicals, Cleveland, OH) в соответствии с инструкцией изготовителей. Во избежание компрессии, вместо dGTP использовали 7-Deaza dGTP. Сначала, для получения последовательности первых 200-250 оснований использовали универсальный праймер вектора M13, а затем из этой последовательности, используя ДНК-синтезатор Applied Biosystems 391 в соответствии с инструкцией изготовителей, синтезировали 17-мерный олигонуклеотид, который использовали в качестве праймера для дальнейшего секвенирования ДНК до тех пор, пока не будут получены данные о полной ДНК-последовательности.
Трансформация видов Streptomyces и E. coli:Компетентные клетки E. coli получали методом с использованием хлорида кальция (Sambrook и др., Molecular Cloning. A Laboratory Manual. 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989) и трансформировали стандартными способами (Sambrook и др. , Molecular Cloning. A Laboratory Manual. 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989). Мицелий S. lividans TK24 культивировали в среде YEME (Hopwood и др., Genetic Manipulation of Streptomyces. A Laboratory Manual. John Innes Foundation, Norwich, UK, 1985) и через 48 часов собирали. Мицелярные пеллеты дважды промывали 10,3% раствором сахарозы, и использовали для получения протопластов способом, описанным в руководстве Hopwood (Hopwood и др., Genetic Manipulation of Streptomyces. A Laboratory Manual, John Innes Foundation, Norwich, UK, 1985). Пеллеты протопластов суспендировали примерно в 300 микролитрах P-буфера (Hopwood и др. , Genetic Manipulation of Streptomyces. A Laboratory Manual John Innes Foundation, Norwich, UK, 1985), и 50-микролитровую аликвоту этой суспензии использовали для каждой трансформации. Протопласты трансформировали с помощью плазмидной ДНК в соответствии с методом мелкомасштабной трансформации Hopwood и др. (Hopwood и др., Genetic Manipulation of Streptomyces. A Laboratory Manual, John Innes Foundation, Norwich, UK, 1985). После 17-часовой регенерации на среде R2YE при 30oC, чашки покрывали 50 мкг/мл тиострептона, и культивировали при 30oC до тех пор, пока не будут образовываться споры. Биоконверсия карминомицина в даунорубицин: Трансформанты S. lividans, включающие в себя различные плазмиды, были инокулированы в жидкую среду R2YE (Hopwood и др., Genetic Manipulation of Streptomyces. A Laboratory Manual, John Innes Foundation, Norwich, UK, 1985), содержащую 5 мкг/мл тиострептона. После 2-х дневного культивирования при 30oC, 2,5 мл этой культуры переносили в 25 мл среды Strohl [(Dekleva и др., Can J. Microbiol. 31:287 (1985)] содержащей 20 мкг/мл. Культуры выращивали в колбе Эрленмейера с перегородкой на роторном шейкере при 300 об/мин и при 30oC в течение 72 часов, после чего к культурам добавляли карминомицин /в виде водного раствора при концентрации 10 млг/мл/ до получения конечной концентрации 5 мкг/мл. После дополнительного 24-часового инкубирования на шейкере, культуры инкубировали в водяной бане при 60oC в течение 45 минут после добавления 150 млг/мл щавелевой кислоты для гидролиза гликозидных форм антрациклиновых метаболитов. Эти метаболиты экстрагировали из культур с использованием 15 мл хлороформа после того, как pH культур доводили до 8,4-8,6. Затем раствор хлороформа фильтровали через 0,45 мкм-Acrodisc CR-фильтр (Gelman Sciences, Ann Arbor, M1) и 100 мкл этого фильтрата анализировали с помощью ВЭЖХ на кассете /8 мм х 10 см/ с Water Nova-Pak C18 /подвижная фаза: метанол : вода, 85:15, доведенная до pH 2,5 путем добавления фосфорной кислоты, скорость потока 3 мл/мин/. Выход колонки контролировали с помощью системы подачи, содержащей водный растворитель Water 6000 УФ-детектора 441, работающего при 254 нм, и модуля обработки данных 740. Для определения количеств указанных метаболитов, выделенных из культур, в качестве внешних стандартов использовали карминомицин и даунорубицин /10 мкг/мл в метаноле/. Пример 1. Клонирование гена dnrK, кодирующего карминомицин-4-О-метилтрансферазу Несколько космидных клонов, описанных Stutzman-Engwall и Hutchinson [(Proc. Natl. Acad. Sci. USA 86: 3135 (1989)] и представляющих собой геномную ДНК S. peucetius 29050 /приблизительно 96 kb/, трансформировали в S. liwidans TK24, и полученные трансформанты анализировали на биоконверсию карминомицина в даунорубицин в соответствии с методом, описанным в главе “Материалы и методы” настоящей заявки. Космидный клон pWHM339 [Otten и др., J. Bacteriol 172:3427 (1990)] осуществлял биоконверсию 22% добавленного карминомицина в даунорубицин. EcoRI-фрагмент /11,2 kb/ из вставки в pWHM339 субклонировали в космидный вектор pKC505 [Richardson и др., Gene 61:231 (1987)], в результате чего получали плазмиду pWHM534. S. liwidans TK24, трансформированный с использованием плазмиды pWHM534, обнаруживал 25-60%-ную биоконверсию добавленного карминомицина в даунорубицин. SphI-фрагмент /5,8 kb/ из pWHM534 субклонировали SphI-сайте высококопийной плазмиды pWHM3, и получали плазмиду pWHM901. S. liwidans трансформированный с помощью плазмиды pWHM901, обнаруживал 50-85%-ную биоконверсию карминомицина в даунорубицин. SphI/PvuII-фрагмент /1,6 kb/ клонировали из pWHM901 сначала в SphI/SmaI-сайты pUC19 [Yansch-Perron и др. Gene 33:103 (1985)], а затем этот 1,6 kb-ДНК-фрагмент субклонировали из последней плазмиды в виде HindIII/EcoRI-фрагмента в HindIII/EcoRI-сайты pWHM3, в результате чего получали плазмиду pWHM902 /фиг. 2/. S. liwidans трансформированный с помощью pWHM902, обнаруживал 100%-ную биоконверсию добавленного в культуру карминомицина в даунорубицин. Область ДНК-последовательности, содержащая ген dnrK Секвенирование 2,5 kb-ДНК-сегмента из 5,8 kb-Sp1-фрагмента в pWHM901 осуществляли путем субклонирования 0,4 kb-SphI/XhoI-, 0,7 kb-XhoI/SstI-, 0,6 kb-SstI/Sall-, и 0,8 kb-Sall/XhoI-фрагментов из pWHM902 в M13mp18 и -mp19-векторы [Yanich-Perron и др., Gene 33:103 (1985)] с получением обеих ориентаций каждого ДНК-сегмента. ДНК-секвенирование полученных четырех клонов осуществляли как описано в главе “Материалы и методы” настоящей заявки. Полученная ДНК-последовательность 1,6 kb-фрагмента ДНК, содержащего dnrK-ген, и аминокислотная последовательность COMT-белка, выведенная путем анализа ДНК-последовательности с использованием программы CODONPREFEPENCE, описанной Derevaux и др. [Nucl. Acids Res. 12:387 (1984)], показаны на фиг. 3. Открытая рамка считывания dnrK, идентифицированная с помощью анализов CODONPRERENCE и TRANSLATE [Derevaux и др., Nucl. Acids Res. 12:387 (1984)], кодирует COMP-белок. Пример 2. Конструирование вектора экспрессии гена dnrK в E. coli SphI/PvuII-фрагмент ДНК, составляющий приблизительно 1,6 kb и содержащий полную открытую рамку считывания dnrK вместе с одной фланкирующей последовательностью /фиг. 3/, субклонировали в SphII- и SmaI-переваренную pUC19 [Yansch-Perron и др., Gene 33:103 (1985)], в результате чего получали плазмиду pWHM904 /не показана/. Затем, используя ДНК-синтезатор (Applied Biosystems 391) в соответствии с инструкциями изготовителей, синтезировали два олигодезоксинуклеотидных праймера, соответствующих последовательностям на обеих сторонах амплифицированного фрагмента, содержащего ген dnrK и представляющего собой: 5′-GGG TCTAGA GGATCC AGGAG CAG CATATG ACC GCT GAA CCG ACC GTC GCG GCC CGG CCG CAG CAG AT-3′: Праймер N 1 (SEQ 1D N 3) 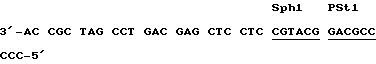 : Праймер N 2 (SEQ ID N 4).
3-е положение второго, третьего и шестого кодонов /показанных буквами жирным шрифтом/ гена dnrK изменяли с использованием прамера N 1 для восстановления наиболее часто используемого кодона в высокоэкспрессируемых генах E. coli, что способствует усилению экспрессии гена dnrK в E. coli.
ATG ACC GCT GAA CCG ACC GTC GCG GCC CGG CCG GAG CAGA: мутированная последовательность (SEQ 1D N) : Праймер N 2 (SEQ ID N 4).
3-е положение второго, третьего и шестого кодонов /показанных буквами жирным шрифтом/ гена dnrK изменяли с использованием прамера N 1 для восстановления наиболее часто используемого кодона в высокоэкспрессируемых генах E. coli, что способствует усилению экспрессии гена dnrK в E. coli.
ATG ACC GCT GAA CCG ACC GTC GCG GCC CGG CCG GAG CAGA: мутированная последовательность (SEQ 1D N)ATG ACA GCC GAA CCG ACG GTC GCG GCC CGG CCG GAG CAGA: последовательность дикого типа (SEQ 1D N) Эти праймеры использовали для амплификации dnrK последовательности pWHM904 от нуклеотида 205 /начало отк. рамки счит. dnrK до нуклеотида 445/ (фиг. 3) с помощью стандартных методов, использующих полимеразно-цепную реакцию с ДНК Streptomyces (см. например, Guilfoill и Hutchinson, J. Bacteriol, 174:3659 (1992)). Из амплифицированного продукта вырезали Ndel/Ncol-фрагмент (88 п.о.) и лигировали с Ncol/EcoRI-фрагментом /1,3 kb/, полученным из pWHM902, содержащим оставшийся ген dnrK (фиг. 2 и 3), и с Ndel/EcoRI -переваренным вектором pT7-7 [Tabor и Richardson Proc. Natl. Acad. Sci. USA 82:1074 (1985)], в результате чего происходило слияние открытой рамки считывания гена dnrK с промотором гена 10T7 указанного вектора экспрессии E. coli. Компетентные клетки DH5 E. coli трансформировали лигированной ДНК, и полученные трансформанты скринировали на pT7-7 с dnrK. Сконструированную в результате плазмиду обозначали pWHM903 /фиг. 4/.
Экспрессия гена dnrK в E. coliКомпетентные клетки E. coli, содержащие плазмиду pGpI-2 [Tabor и Richardson, Proc. Natl. Acad. Sci. USA] , подвергали селекции на LB-агаре (Sambrook и др. , Molecular Cloning. A Laboratory Manual, 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989), содержащем ампициллин /100 мкг/мл/ и канамицин (50 мкг/мл), после их культивирования при 30oC. Процедура получения компетентных клеток E. coli, содержащих pGPI-2, в основном такая же, как и для любого другого штамма E. coli за исключением того, что культуры клеток выдерживают при 30oC, а не при 37oC. Компетентные клетки E. coli содержащие pGPI-2, получали из клеток, культивированных при 30oC до ОП550 = 0,5 – 0,6 в LB-среде, содержащей канамицин. Во избежание чрезмерной экспрессии РНК-полимеразы T7, что может привести к получению мутантной плазмиды, очень важно при обычном хранении и при предварительном культивировании штаммов, содержащих pGPI-2, поддерживать температуру 30oC. Один трансформант, содержащий pGPI-2 и pWHM903 инокулировали в 25 мл 2 х YT-среды (Sambrook и др., Molecular Cloning. A Laboratory Manual, 2-ое изд., Cold Spring Harbor Press, Cold Spring Harbor, NY, 1989), содержащую 100 мкг/мл ампициллина и 50 мкг/мл канамицина, и культивировали при 30oC в течение ночи, энергично размешивая, при этом. На следующее утро культуры подвергали резкому нагреванию при 42oC в течение 30 минут в водяной бане-шейкере, а затем снова переносили в условия температуры 30oC. После последующей 90-минутной инкубации 1 мл культуры центрифугировали 1 минуту в микроцентрифуге при 14000 об/мин., супернатант отбрасывали, а клеточный дебрис ресуспендировали в 100 мкл буфера Лэммли (Laemmli, Nature (Лондон), 227:680 (1970) и кипятили в течение 5 минут. Белки, содержащиеся в прокипяченом образце, анализировали в 10% ДСН-полиакриламидном геле, используя стандартную технику [Laemmli, Nature (Лондон), 227:680 (1970)], путем сравнения с белками, полученными из клеточного экстракта E. coli, трансформированной с помощью вектора pT7-7, не содержащего ген dnrK, COMT-белок мигрирует при Mr 38700. Пример 3. Превращение карминомицина в даунорубицин клетками, содержащими COMT-белок Один трансформант E. coli, содержащий pGPI-2 и pWHM903, инокулировали в 25 мл 2 х YT-среды, содержащей 100 мкг/мл и 50 мкг/мл канамицина, и культивировали при 30oC в течение ночи, энергично размешивая при этом. На следующее утро культуры подвергали резкому нагреванию при 42oC в течение 30 минут в водяной бане-шейкере, а затем, после добавления 5 мкг/мл карминомицина, снова переносили в условия 30oC. После этого культуры культивировали еще 90 минут, затем антрациклиновые метаболиты выделяли стандартными методами и анализировали с помощью ВЭЖХ. Сравнение соответствующих областей пиков сигналов для карминомицина и даунорубицина на ВЭЖХ-хроматограмме показало, что 7580 карминомицина, добавленного в культуральную среду, превратилось в даунорубицин. Формула изобретения
РИСУНКИ
MM4A – Досрочное прекращение действия патента СССР или патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе
Дата прекращения действия патента: 18.11.2007
Извещение опубликовано: 20.07.2010 БИ: 20/2010
|
||||||||||||||||||||||||||