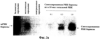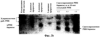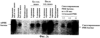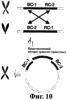Патент на изобретение №2270866
|
||||||||||||||||||||||||||
(54) МОЛЕКУЛЯРНЫЙ КОНТРОЛЬ СЕГРЕГАЦИИ ТРАНСГЕНОВ И ИХ УТЕЧКИ С ИСПОЛЬЗОВАНИЕМ СИСТЕМЫ ВОЗМЕЩАЕМОГО БЛОКИРОВАНИЯ ФУНКЦИИ
(57) Реферат:
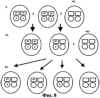
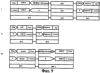
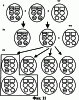
Изобретение относится к генной инженерии. Комплекс ДНК-конструкций содержит трансгенный инсерт, включающий представляющий интерес трансген, одну или более блокирующих конструкций и одну или более необязательных восстанавливающих конструкций, помещенных вблизи блокирующих конструкций. Включение указанного комплекса ДНК-конструкций в геном растения позволяет повторно использовать трансгенную сельскохозяйственную культуру без риска утечки трансгенов в окружающую среду. Это особенно подходит для растений, склонных к аутокроссингу с их дикими типами или культивируемыми родственными формами. 5 н. и 40 з.п. ф-лы, 11 ил.
Область техники, к которой относится изобретение Данное изобретение относится к усовершенствованному способу и комплексу ДНК-конструкций, называемых здесь системами возмещаемого блокирования функции (RBF), для получения увеличенного уровня безопасности в контроле сегрегации трансгенов и “утечки” трансгенов, в том числе интрогрессии. Эти способ и система позволяют также фермерам повторно использовать их трансгенную сельскохозяйственную культуру без риска утечки трансгенов в окружающую среду. Повышенный уровень надежности достигается с использованием систем возмещаемого блокирования функции (RBF), включающих в себя одну или более блокирующих нуклеотидных конструкций (ВС), тесно связанных с представляющим интерес геном (TGI), причем эту блокирующую конструкцию встраивают в интрон представляющего интерес гена (TGI), если используют одну блокирующую конструкцию (ВС), или она находится между двумя блокирующими конструкциями (ВС), если используют более чем одну блокирующую конструкцию, а также одну или более восстанавливающих конструкций (RC), т.е. нуклеотидных конструкций для восстановления блокированной функции при контролируемом пользователем химическом, физическом или механическом вмешательстве. Данное изобретение относится не только к способу, но также к системам возмещаемого блокирования функции (RBF), a также к их применению в получении векторов, клеток, клеточных линий и/или трансгенных размножающихся половым путем многоклеточных организмов (SRMO), в частности растений и некоторых животных, таких как рыба, мелкие ракообразные, моллюски и т.д. Уровень техники На протяжении нескольких последних лет безопасность получения трансгенных сельскохозяйственных культур была причиной большой озабоченности среди общественности в целом и она привлекала большое внимание также и сообщества ученых. В нескольких публикациях сообщалось, что пыльца трансгеных растений распространяется с полевых участков. Наиболее проблематичными являются трансгеные сельскохозяйственные виды, которые имеют дикие родственные формы, с которыми они могут гибридизоваться в природе, если им удалось ускользнуть, но важно также предупреждать и контролировать скрещивание между различными, но родственными трансгенными или нетрансгенными сельскохозяйственными растениями или родственными дикими типами, т.е. сохранять чистоту зародышевых линий. Группы риска были найдены среди определенных сельскохозяйственных видов, например кукурузы, некоторых масличных растений, таких как Brassicae, но также среди деревьев, которые являются специфической проблемой вследствие их большой продолжительности жизни и большого продуцирования пыльцы. Даже если эта проблема ограничена в настоящее время растениями, существует вероятность, что подобные проблемы встретятся, когда животные станут более серьезными мишенями в трансгенных способах получения. Рыба, мелкие ракообразные, домашняя птица, овцы и т.д. являются уже возможными в качестве трансгенных животных. Вероятно также, что большой диапазон модификаций будет применен к трансгенным растениям в будущем, что усиливает потребность в предупреждении утечки трансгенов. Хотя опасности для здоровья человека или животного конкретного трансгена и его продукта могут быть испытаны и измерены, влияние утечки генов оценить более сложно. С другой стороны, потенциальные возможности улучшения сельскохозяйственных культур с использованием способов рекомбинантных ДНК являются настолько огромными, что более продуктивно и выгодно с точки зрения расходов найти решения для предупреждения утечки или ускользания генов, чем запрещать применение трансгенеза. Вследствие большого влияния трансгенеза и беспокойств, возникающих в отношении утечки трансгенов, несколько подходов для решения этой проблемы были представлены в предшествующей литературе. Эти первые подходы включали в себя растения со сконструированной стерильностью и контролем прорастания семян, называемых также «терминаторной технологией», предпринятой фирмой Monsanto (Патент США 5723765). Были опубликованы также несколько сообщений о применении условных летальных факторов, также называемых суицидными генами. Гены ослабления, т.е. гены, ослабляющие селективные преимущества трансгенов в сорняках, были предложены для использования в контроле утечки трансгенов (Gressel, J., (1999), Tibtech. 17: 361-366). Применение полученной генной инженерией мужской или женской стерильности исследовалось несколькими группами. В способе мужской стерильности (MS) предотвращалось опыление, например, остановкой синтеза мРНК. Опыление может быть восстановлено экспрессией ингибитора РНКазы. В указанном случае фактор восстановления (RF) переносится опыляющей линией. Эти два элемента, MS и RF, присутствуют в двух различных индивидуальных растениях. Только гибрид этих двух растений-индивидуумов несет как блокирующий, так и восстанавливающий факторы и посредством этого является фертильным. Таким образом, способ мужской стерильности направлен на поддержание образования гибридных семян. Однако мужская стерильность не предотвращает утечки трансгена в окружающую среду, так как опыленные женские (MS) растения все еще способны к продуцированию гибридных семян, которые при осыпании остаются в поле после сбора урожая. Полученный генной инженерией контроль фертильности достигался также в трансгенном Brassica napus экспрессией гена РНКазы под контролем специфического для выстилающего слоя (тапетума) пыльника промотора табака (De Block and Debrouwer, (1993), Planta, 189: 218-225). Экспрессия гена РНКазы в клетках тапетума пыльника убивает пыльцу на ранних стадиях развития. Опыление цветков с мужской стерильностью пыльцой трансгенных растений В. napus, экспрессирующей ген РНКазы под контролем того же самого промотора, восстанавливает признак мужской стерильности (MS), и эти гибридные растения не могут продуцировать нормальную фертильную пыльцу с экспрессией этих двух генов. Патент США 5750867 описывает сохранение растений с мужской стерильностью. Патент США 5767374 описывает сходный способ для растений с женской стерильностью (FM), в котором ген рибонуклеазы экспрессируется в клетках тычинок женских родительских растений, и экспрессия этого гена-убийцы в гибридных растениях восстанавливается экспрессией гена-восстановителя, поступающего из опыляющей родительской линии. Этот блокирующий ген экспрессируется в женских органах родительского растения, тогда как пыльца остается фертильной. Предполагается, что растения с женской стерильностью (FS) должны опыляться растениями с мужской стерильностью (MS) для получения гибридных семян. В патенте США 572892 описан способ на основе экспрессии антисмысловой мРНК гена, жизненно важного для развития пыльников. Эта антисмысловая молекула экспрессируется одновременно со смысловой молекулой, причем экспрессия обеих из них запускаются одним и тем же промотором. Одновременная экспрессия гена со смысловой и антисмысловой ориентацией обеспечивает механизм молчания, который предотвращает развитие пыльников. В патенте США 6013859 описан способ гибридизации. Блокирование обеспечивается двумя последовательными ферментативными реакциями в пыльниках или микроспорах. Опыляющая родительская линия сообщает устойчивость к селективному маркеру (гербициду). Патент США 6005167, в свою очередь, описывает способ, который использует антисмысловой способ для обеспечения мужской или женской стерильности на основе блокирования экспрессии халконсинтазы в развивающемся пыльнике или другой части цветка. Халконсинтаза является ключевым ферментом в синтезе флаваноидов. Блокирование экспрессии этого гена приводит к необратимому блокированию оплодотворения. В патенте США 5723765 описан способ прекращения прорастания или функции семян. Этот способ предусматривает активацию функции ингибированного блокирующего гена посредством вырезания специфической последовательности ДНК между промотором и блокирующим или терминаторным геном специфическим ферментом Cre-рекомбиназой, кодируемым другим геном, помещенным под контроль Tet-репрессируемого промотора. Семена трансгенных растений, не обработанные тетрациклином, способны прорастать в природных условиях. Если эти трансгенные семена обрабатывают тетрациклином, ген, кодирующий Cre-рекомбиназу, активируется и вырезает ДНК-инсерт между промотором Lea и геном токсина. Посредством этого блок удаляется и токсин активируется. Этот токсин не убивает растение сразу же, так как экспрессия должна инициироваться только во время поздней стадии эмбриогенеза под действием промотора Lea. Идея, описанная в патенте США 5723765, предполагает ингибирование развития семян во втором поколении. Без механизма супрессии ген-«убийца» активируется во время поздних стадий развития зародыша потомства, и, следовательно, семена следующего поколения не прорастают. Фундаментальной проблемой указанного способа является то, что как только эти растения обрабатывали тетрациклином, т.е. ген-убийца активировался, они не могли быть спасены. Кроме того, если указанное трансгенное растение, несущее восстанавливаемую тетрациклином конструкцию, ускользает в окружающую среду, оно способно прорастать, вырастать до зрелого состояния, цвести и размножаться половым путем. Другими словами, не предупреждается утечка трансгена из трансгенных растений. Указанная так называемая «терминаторная технология» встретила негативное внимание общественности, так как она дает производящим эти семена компаниям возможность контролировать рынок производства трансгенных семян. Международная патентная заявка WO 94/03619 описывает контроль роста трансгенного растения посредством химической активации или «генного переключения». Переключенный ген продуцирует репрессор, который может инактивировать экспрессию «гена-разрушителя». При применении химической индукции гена-репрессора ген-разрушитель репрессируется. Если ген репрессора не активируется, ген-разрушитель разрушает данную клетку и функции данного растения. Механизм репрессии может включать в себя репрессорный белок, в том числе систему промотора и оператора. Альтернативно, репрессор может присутствовать в виде гена, кодирующего ингибитор гена, кодирующего белок-разрушитель. «Химический переключатель», описанный в WO 94/03619, может быть также использован для контроля активности гена рекомбиназы, который может удалять последовательность ДНК, фланкированную свитами узнавания рекомбиназы. Фермент рекомбиназа удаляет часть гена данного признака и блокирует функцию ценного добавленного признака. Международная патентная заявка WO 00/37660 описывает способы и генетические композиции для ограничения ауткроссинга и перетекания нежелательного гена в сельскохозяйственные растения. WO 00/37660 описывает генетическую систему, содержащую две ДНК-конструкции, репрессируемый доминантный летальный ген и репрессорный ген, который локализован в локусе, сегрегирующемся независимо от репрессируемого доминантного летального гена. Описана также сложная система клонирования и селекции для сконструированных растений, содержащих репрессируемый летальный ген и репрессорный ген в различных сестринских хромосомах. Патент США 5498533 описывает регуляцию развития картофеля экспрессией смысловых и антисмысловых конструкций гена кальмодулина. Экспрессия гена кальмодулина в смысловой ориентации увеличивает рост побегов и клубней, тогда как растения, несущие антисмысловые конструкции, проявляют уменьшенный рост побегов и клубней, в то время как экспрессия антисмыслового гена кальмодулина может быть использована в качестве фактора, блокирующего физиологическую функцию. Gressel (1999) в его обзоре (Tibtech, 17: 361-366) описывает несколько применений существующих и гипотетических систем на основе способа тандемного трансгенетического ослабления (ТМ), который может быть использован для предупреждения появления суперсорняков. ТМ-гены являются позитивными или нейтральными в отношении сельскохозяйственной культуры, но вредными для сорняка. ТМ-гены могут изменять морфологию или физиологию сорняка обеспечением покоя семян, раннего или позднего созревания семян, карликовости растений, а также других признаков, которые ослабляют эволюционную адаптированность произвольных сорняков в сравнении с родительским штаммом. Однако гены ослабления не могут полностью предотвращать утечку трансгенов, так как они не являются летальными и даже не являются вредными для сельскохозяйственных растений. Таким образом, технология трангенетического ослабления не предотвращает скрещивания между различными линиями одних и тех же сельскохозяйственных культур. Эффект ослабления генов обычно достигается только после пролонгированных периодов негативного отбора, во время которых возможные образования молчащих генов или мутации в ослабляющих генах уменьшают их действие и вследствие этого действуют против предотвращения утечки трансгенов. В обзорной статье Gressel (1999) предлагает применение тандемных конструкций, содержащих два гена ослабления, для уменьшения возможности утечки трансгенов. Тандемы генов ослабления не решают проблемы скрещивания между различными линиями родственных сельскохозяйственных растений. Молчание или мутации в одном гене ослабления уменьшают действие давления негативного отбора. Большинство систем, предназначенных для обеспечения контроля утечки трансгенных растений, были описаны как невозмещаемые. Другими словами, они не могут быть повторно использованы после применения системы восстановления. Дополнительным недостатком в некоторых из вышеописанных систем является то, что они требуют вегетативного размножения, поскольку прекращается оплодотворение или развитие зародыша. Таким образом, ситуация контроля далека от идеальной, несмотря на множество различных подходов, предложенных в предшествующем уровне знаний. Требуются другие и более эффективные способы и системы для решения данной проблемы. При использовании ослабляющих генов можно сделать вывод, что с ослабляющими генами не достигается достаточная безопасность. Требуются летальные или вызывающие стерильность гены. Прежние системы, применяющие летальные или вызывающие стерильность гены, были описаны и использованы, но они не могут гарантировать достаточные уровни надежности, в частности, при необходимости более продолжительных периодов селекции. Например, предыдущие способы и системы не отвечают на следующий вопрос: Что происходит в часто встречающихся случаях, когда инактивированы летальные гены, гены-разрушители или блокирующие гены? Можно ожидать, что летальные или вызывающие стерильность гены становятся молчащими или мутированными с частотой около 10-6, что делает возможной небольшую, но допускаемую утечку трансгенов. Кроме того, некоторые из прежних систем, использующих летальные или вызывающие стерильность гены, требуют нежелательных стадий, таких как распространение химикалиев в окружающую среду. Это не очень хорошо воспринимается пользователями или общественностью в целом. Можно упомянуть также отсутствие возможности повторного использования сельскохозяйственной культуры. Основной целью данного изобретения является обеспечение повышенного уровня безопасности для контроля интрогрессии или сегрегации (расщепления) и утечки трансгенов в окружающую среду. Дополнительной целью данного изобретения является предоставление фермеру точных инструкций по повторному использованию его сельскохозяйственной культуры. Следующей целью данного изобретения является обеспечение системы, в которой действия генов, имеющих блокирующий эффект, могут быть восстановлены без применения химикалиев, и, если химикалии должны быть использованы, это восстановление может быть проведено в ограниченных условиях (в закрытом помещении). Сущность изобретения Повышенный уровень безопасности для контроля сегрегации трансгенов и утечки в окружающую среду достигается посредством способа и/или системы, включающих комплекс конструкций ДНК, содержащих одну или несколько блокирующих конструкций (ВС), содержащих гены, способные блокировать одну или несколько функций, которые являются существенными для выживания и полового размножения трансгенного организма. Повышенный уровень безопасности достигается включением в реципиентный организм или размножающийся половым способом многоклеточный организм (SRMO) в форме одного или нескольких трансгенных инсертов (TI) или ДНК-кассет, одной или нескольких блокирующих конструкций (ВС), которые могут быть помещены в интрон представляющего интерес гена (TGI) или могут быть помещены в тесной близости с трансгенным инсертом (TI), содержащим один или несколько представляющих интерес трансгенов (TGI). Функция, блокированная этой блокирующей конструкцией (ВС), предпочтительно восстанавливается контролируемым пользователем вмешательством, которое не присутствует в природных условиях. Если происходит ауткроссинг трансгенного организма, функция, которая является существенной для выживания или полового размножения, блокируется. Контролируемое пользователем вмешательство является таким, что оно может быть повторено не только один раз, но и несколько раз, и предпочтительно это вмешательство не требует химических вмешательств, которые должны проводиться в окружающей среде. Повышенный уровень безопасности, в сравнении с предыдущими способами и системами, достигается конкретно с использованием двух или нескольких блокирующих конструкций (ВС), помещенных на каждой стороне трансгенного инсерта (TI), содержащего один или несколько представляющих интерес трансгенов (TGI), или помещением блокирующей конструкции (ВС) в интроне представляющего интерес трансгена (TGI). Таким образом, данное изобретение относится к способу увеличения уровня надежности для контроля сегрегации трансгенов и предупреждения утечки трансгенов в окружающую среду с использованием молекулярного механизма контроля, предусматривающего стадии конструирования одного или нескольких комплексов ДНК-конструкций, называемых здесь системами возмещаемого блокирования функции (RBF), содержащих, кроме одного или нескольких представляющих интерес трансгенов (TGI), одну или несколько блокирующих конструкций (ВС), расположенных в тесной близости с представляющим интерес трансгеном (TGI), предпочтительно в интроне представляющего интерес трансгена (TGI), или при использовании более чем одной блокирующей конструкции (ВС), между указанными блокирующими конструкциями (ВС), и по меньшей мере одно средство для восстановления, или восстанавливающий инструмент, т.е. контролируемое пользователем средство или вмешательство для восстановления блокированных функций. Указанная блокирующая конструкция (ВС) обладает способностью блокировать по меньшей мере одну функцию, существенную для выживания и/или полового размножения размножающегося половым способом многоклеточного организма (SRMO). Нуклеотидная последовательность или ген, обеспечивающие блокирующий эффект в блокирующей конструкции (ВС), либо является представляющим интерес трансгеном (TGI), либо является тесно связанным с по меньшей мере одним представляющим интерес трансгеном (TGI), кодирующим желаемый трансгеный продукт. Восстановление блокированных функций предусматривает по меньшей мере одно контролируемое пользователем вмешательство, необязательно объединенное с одной или несколькими восстанавливающими конструкциями (RC). Этот контроль имеет место автоматически посредством негативного отбора в поколениях после первого ускользнувшего гибридного поколения, содержащего представляющий интерес ген (TGI). Присутствие восстанавливающей конструкции (RC) в технологии возмещаемого блокирования функции (RBF) данного изобретения позволяет использовать в блокирующей конструкции (ВС) трансгены, которые могут быть летальными или вызывать стерильность в растении-хозяине. Применение таких генов делает негативный отбор, требуемый для того, чтобы этот эффект имел место, абсолютным и укорачивает необходимое время для негативного отбора в сравнении с негативным отбором в технологии ослабления генов. Уровень безопасности систем возмещаемого блокирования функции (RBF) с использованием двойных блокирующих конструкций (ВС) увеличивается, так как молчание или мутация в одной из блокирующих конструкций (ВС) не уменьшает действие второй блокирующей конструкции (ВС). Альтернативно, уровень безопасности систем возмещаемого блокирования функции (RBF) данного изобретения может быть увеличен введением блокирующей конструкции (ВС) в интрон представляющего интерес трансгена (TGI). Это уменьшает возможности того, что блокирующая конструкция (ВС) также инактивируется посредством кроссинговера или мутирует, без вредных эффектов в отношении представляющего интерес трансгена. Таким образом, комбинирование концепции систем возмещаемого блокирования функции (RBF) с системой множественной, т.е. двойной или тройной и/или введенной в интрон, блокирующей конструкции (ВС) обеспечивает существенно увеличенный уровень надежности в контроле сегрегации (расщепления) трансгенов и утечки трансгенов, чтобы сделать эту систему выполнимой в ДНК-технологии трансгенных растений и животных на практике. Трансгенный размножающийся половым способом многоклеточный организм (SRMO) в конечном счете предохраняется от полового размножения или продуцирования жизнеспособного потомства блокирующей конструкцией (ВС), которая в природных, неконтролируемых условиях прекращает функцию, которая является существенной для выживания или размножения указанного размножающегося половым способом многоклеточного организма (SRMO). Эта блокированная функция может быть восстановлена для того, чтобы сделать возможными нормальное развитие, пролиферацию, рост и половое размножение для целей сельского хозяйства, садоводства, лесоводства или любых других возможных целей с использованием по меньшей мере одного контролируемого пользователем вмешательства, которое в чувствительный для этого момент в цикле развития размножающегося половым способом многоклеточного организма (SRMO) восстанавливает блокированную функцию. Указанное средство для восстановления функции предусматривает внешнюю, контролируемую пользователем стадию обработки или манипуляции, в том числе химическое, физическое или механическое вмешательство, которое восстанавливает или разблокирует блокированную функцию. Трансген предохраняется от утечки в окружающую среду посредством гибридизации или ауткроссинга родительского трансгенного размножающегося половым способом многоклеточного организма (SRMO) с его родственными формами дикого типа или другими культивируемыми нетрансгенными или трансгенными родственными формами с использованием функции комплекса блокирующих конструкций (ВС), которая в природных условиях прекращает существенную функцию трансгенного размножающегося половым способом многоклеточного организма (SRMO) или любых гибридов, несущих указанную блокирующую конструкцию (ВС) до тех пор, пока не будет обеспечено контролируемое пользователем вмешательство извне. Этот контроль сегрегации быстро приводит к исчезновению трансгена в природе. Данное изобретение обеспечивает комплекс ДНК-конструкций для предотвращения сегрегации и утечки трансгенов в размножающемся половым способом многоклеточном организме (SRMO) в окружающую среду. Указанный комплекс ДНК-конструкций, называемый здесь системой возмещаемого блокирования функции (RBF), содержит одну или несколько единиц блокирующих конструкций (ВС), содержащих по меньшей мере одну нуклеотидную последовательность, способную блокировать молекулярную или физиологическую функцию, которая является существенной для выживания и/или размножения трансгенного размножающегося половым способом многоклеточного организма (SRMO). Ген указанной нуклеотидной последовательности в блокирующей конструкции (ВС) может быть альтернативно представляющим интерес трансгеном (TGI), т.е. он может быть нуклеотидной последовательностью, образующей самоконтролирующее образование. Предпочтительно он может быть помещен в тесно связанном виде в непосредственной близости или в соседстве с представляющим интерес трансгеном (TGI), либо в интроне, либо между двумя блокирующими конструкциями (ВС). Система возмещаемого блокирования функции (RBF) может также содержать одну или несколько необязательных восстанавливающих конструкций (RC), которые помещены вблизи от блокирующих конструкций (ВС) или в неаллельных хромосомах. Восстанавливающие конструкции (RC) не должны помещаться в сестринской хромосоме хромосомы, содержащей представляющий интерес трансген (TGI), фланкированный блокирующими конструкциями (ВС), или с блокирующими конструкциями (ВС), встроенными в интрон представляющего интерес трансгена (TGI). Восстанавливающие конструкции (RC) являются регулируемыми внешним, контролируемым пользователем, искусственным вмешательством. Другими словами, система возмещаемого блокирования функции (RBF) может функционировать с восстанавливающей конструкцией (RC) или без восстанавливающей конструкции до тех пор, пока некоторые внешние средства или вмешательства, контролируемая пользователем обработка для восстановления не станут доступными и не смогут быть применены. Трансгенный размножающийся половым способом многоклеточный организм (SRMO) предохраняется от размножения прекращением прилагаемого пользователем вмешательства, которое предотвращает действие блокирующей конструкции (ВС). Это действие, например ауткроссинг, прекращает и/или изменяет существенную функцию, действуя на выживание, развитие и/или половое размножение трансгенного размножающегося половым способом многоклеточного организма (SRMO). Блокированная функция восстанавливается в чувствительный для этого момент роста или цикла развития размножающегося половым способом многоклеточного организма (SRMO), чтобы позволить нормальный рост и размножение указанного организма для целей продуцирования, в том числе для применений в сельском хозяйстве, плодоовощном хозяйстве, лесоводстве и/или промышленности, путем применения по меньшей мере одного применимого извне искусственного и контролируемого пользователем вмешательства. Если трансгенный размножающийся половым способом многоклеточный организм (SRMO) склонен к интербридингу или ауткроссингу с его культивируемыми родственными формами дикого типа, сегрегация трансгена контролируется и/или утечка указанного трансгена в природу предотвращается тем фактом, что в природе не обеспечивается вмешательство или обработка, которая устраняла бы негативные эффекты блокирующей конструкции (ВС). Таким образом, в природных условиях прекращается по меньшей мере одна существенная молекулярная или физиологическая функция трансгенного размножающегося половым способом многоклеточного организма (SRMO), которая предотвращает половое размножение. В способе данного изобретения функция, влияющая на выживание размножающегося половым способом многоклеточного организма-хозяина (SRMO) или его цикл развития или репродуктивный цикл, характеризуется прекращением развития, изменением фенотипа или морфологии трансгенного размножающегося половым способом многоклеточного организма (SRMO) органоспецифическим, пространственно-временным или конститутивным образом так, что предотвращается выживание или половое размножение. Подходящим моментом для удаления блокирующего эффекта или восстановления от блокирующего эффекта посредством стадии внешнего, искусственного и контролируемого пользователем вмешательства или обработки является чувствительные для этого момент или стадия в цикле развития размножающегося половым способом многоклеточного организма (SRMO), причем в этот момент органоспецифическое, пространственно-временное или конститутивное морфологическое изменение предохраняет размножающийся половым способом многоклеточный организм (SRMO) от свободной гибридизации или скрещивания и тем самым от полового размножения в природных условиях. Такими чувствительными моментами являются, например, раннее или позднее прорастание семян, цветение, неспособность образования соцветия, цветков или плодов и/или образование карликового фенотипа, в частности, когда блокирующая конструкция (ВС) содержит нуклеотидную последовательность, экспрессия которой блокирует существенную молекулярную или физиологическую функцию или вызывает морфологическое изменение, предотвращающее свободную гибридизацию и/или размножение. Блокирование имеет место на уровне ДНК, мРНК, белка или метаболита и приводит к некрозу, фенотипическому или физиологическому изменению, такому как раннее или позднее прорастание семян, отсутствие роста и/или вегетации, неспособность образования цветков, соцветий, цветков или плодов, образование карликового фенотипа и т.д. Блокирующая конструкция (ВС) может быть помещена в интроне нуклеотидной последовательности представляющего интерес трансгена (TGI) или две или несколько блокирующих конструкций (ВС) помещают на каждой стороне трансгенного инсерта (TI), содержащего один или несколько представляющих интерес трансгенов (TGI). Если две блокирующие конструкции (ВС) являются одинаковыми или различными и восстанавливаемыми одинаковыми или различными механизмами восстановления, их помещают на обеих сторонах представляющей интерес нуклеотидной последовательности (TGI) в системе множественного, т.е. двойного или тройного возмещаемого блокирования функции (M-RBF, D-RBF или T-RBF). В противоположность этому, восстанавливающая конструкция (RC) должна быть помещена в один и тот же SRMO, но предпочтительно в различных неаллельных хромосомах, что приводит к системе сегрегирующего возмещаемого блокирования функции (S-RBF). В системах множественного возмещаемого блокирования функции (M-RBF) первую восстанавливающую конструкцию (RC1) помещают вблизи от второй блокирующей конструкции (BC2) в первом трансгенном инсерте (TI). Помещенный во второй неаллельной хромосоме комплекс вторая восстанавливающая конструкция – первая блокирующая конструкция (RC2-BC1) контролирует данный трансгенный инсерт (TI) и этот первый трансгенный инсерт (TI) контролирует второй трансгенный инсерт (TI), соответственно. Комплекс представляющий интерес трансген-блокирующие конструкции (BC1-TGI-BC2) контролирует другой комплекс представляющий интерес трансген-блокирующая конструкция (TGI-BC) с увеличенной надежностью и негативным отбором представляющего интерес трансгена (TGI). В данном изобретении рассматриваются также клонирующие векторы, т.е. ДНК-кассеты, и клетки или клеточные линии для удобного получения трансгенных растений или трансгенных животных (за исключением человека), несущих один или несколько трансгенных инсертов (TI), контролируемых системами возмещаемого блокирования функции (RBF) данного изобретения. Также представлен новый сайт поликлонирования, показанный в SEQ ID NO:4, для удобного инсертирования представляющих интерес трансгенов (TGI) и других блокирующих и восстанавливающих конструкций (ВС и RC). Описаны также синтетические нуклеотидные последовательности, которые адаптированы либо для инсертирования в интроны представляющих интерес трансгенов (TGI), либо для фланкирования представляющих интерес трансгенов (TGI) и для инсертирования в модельные растения и/или в модельных животных. Характерные признаки данного изобретения определены более подробно в формуле изобретения. Подробное описание чертежей Фиг.1 является схематическим представлением молекулярных конструкций, используемых в системах простого возмещаемого блокирования функции (RBF). Эти конструкции представлены в деталях (верхний ряд) для демонстрации принципов помещения генов и на более общем уровне (средний и нижний ряды). Блоки промоторов, представленные остриями стрелок, показывают направление этих генных последовательностей. Аббревиатуры: р – промотор, 3′-конец – 3′-конец гена или сайта сигнала полиаденилирования; as – антисмысловая. Фиг.1а представляет конструкцию, описанную в примере 1. Фиг.1b представляет конструкцию, описанную в примере 2. Фиг.2 представляет молекулярные конструкции, используемые в системе возмещаемого блокирования функции (RBF) и описанные в примере 3. Эти конструкции представлены в деталях (верхний ряд) для демонстрации принципов помещения генов и на более общем уровне (средний и нижний ряды). Блоки промоторов, представленные остриями стрелок, показывают направление этих генных последовательностей. Аббревиатуры: р – промотор, 3′-конец – 3′-конец гена или сайта сигнала полиаденилирования. Фиг.3. Нозерн-блот-анализ экспрессии барназы (бактериальной рибонуклеазы) и barstar (ингибитора барназы) в трансгенном табаке во время созревания зародыша, прорастания семян и с обработкой тепловым шоком и без обработки тепловым шоком. Гены барназы и barstar используются в качестве блокирущего и восстанавливающего генов, соответственно, в системе возмещаемого блокирования функции (RBF). Фиг.3а показывает экспрессию мРНК барназы в зародышах табака на средней стадии (беловатые зародыши) и поздней стадии (желтоватые зародыши) эмбриогенеза. 10 мг тотальной РНК наносили на каждую дорожку. Тотальную РНК из нетрансгенных зародышей и 0,1, 0,3 и 1,0 пг синтезированной РНК барназы, смешанной с 10 мг тотальной РНК из нетрансформированных зародышей табака, использовали в качестве контролей. Экспрессия мРНК барназы составляла на средней стадии эмбриогенеза 0,1 пг/мг тотальной РНК, а на поздней стадии эмбриогенеза 0,03 пг/мг тотальной РНК. Фиг.3b показывает экспрессию мРНК барназы в проростках табака. 10 мг тотальной РНК наносили на каждую дорожку. Тотальную РНК из нетрансгенных зародышей и 0,2, 0,5 и 1,0 пг синтезированной РНК барназы, смешанной с 10 мг тотальной РНК из нетрансформированных зародышей табака, использовали в качестве контролей. Экспрессия мРНК барназы начиналась на третий день прорастания, достигала максимума 0,04 пг/мг тотальной РНК на четвертый день и угасала на пятый день прорастания. На блоте были обнаружены также две неспецифические полосы хлоропластной рибосомной РНК. Фиг.3с показывает экспрессию мРНК barstar после теплового шока (h.s.) и без теплового шока. 10 мг тотальной РНК наносили на каждую дорожку. 0,5, 2,0 и 10,0 пг синтезированной РНК barstar, смешанной с 10 мг тотальной РНК из нетрансгенных контрольных зародышей табака, использовали в качестве положительного контроля. Обработка тепловым шоком активировала экспрессию мРНК барназы до 1-3 пг/мг тотальной РНК как в стадии эмбриогенеза, так и в стадии прорастания во время развития. Экспрессия мРНК barstar под контролем промотора теплового шока HSp была растекающейся без температурной обработки, на уровне 0,05-0,2 пг/мг тотальной РНК. Фиг.4 представляет молекулярные конструкции, используемые в системе возмещаемого блокирования функции (RBF) и описанные в примере 4. Эти конструкции представлены в деталях (верхний ряд) для демонстрации принципов помещения генов и на более общем уровне (средний и нижний ряды). Блоки промоторов, представленные остриями стрелок, показывают направление этих генных последовательностей. Аббревиатуры: р – промотор, 3′-конец – 3′-конец гена или сайта сигнала полиаденилирования. Конструкции I и II являются инсертами в различных неаллельных хромосомах и показаны раздельно. Фиг.5 представляет молекулярные конструкции, используемые в системе возмещаемого блокирования функции (RBF) и описанные в примере 5. Эти конструкции представлены в деталях (верхний ряд) для демонстрации принципов помещения генов и на более общем уровне (средний и нижний ряды). Блоки промоторов, представленные остриями стрелок, показывают направление этих генных последовательностей. Конструкции I и II являются инсертами в различных неаллельных хромосомах и показаны раздельно. Аббревиатуры: р – промотор, 3′-конец – 3′-конец гена или сайта сигнала полиаденилирования. Фиг.5а показывает конструкцию в хромосоме I в деталях и на более общем уровне. Фиг.5b показывает конструкцию в хромосоме II в деталях и на более общем уровне. Фиг.6 представляет последовательность гена барназы, помещенную внутри интрона гена uidA: Конструкция гена барназы показана по ходу транскрипции (справа) и содержит промотор SH-EP (показаны приблизительно последние триста нуклеотидов), кодирующую последовательность и сайты сигнала полиаденилирования. Конструкция гена uidA помещена против хода транскрипции (слева) и начинается от нуклеотида 1735 кодирующей последовательности. Конструкция гена uidA показывает последовательности экзон-интрон-экзон-сайт полиаденилирования. Аббревиатуры: nt – нуклеотиды; FUE – дальний элемент против хода транскрипции сайта полиаденилирования; NUE – ближний элемент против хода транскрипции сайта полиаденилирования; SphI, Spel, PstI, Bcll – сайты рестрикции; > и < – направления сигнальных сайтов, СААТ и ТАТА, сигнальных блоков промотора. Фиг.7 представляет молекулярные конструкции, используемые в системе возмещаемого блокирования функции (RBF) и описанные в примере 6. Эти конструкции представлены в деталях (верхний ряд) для демонстрации принципов помещения генов и на более общем уровне (средний и нижний ряды). Блоки промоторов, представленные остриями стрелок, показывают направление этих генных последовательностей. Конструкции I и II являются инсертами в различных неаллельных хромосомах и показаны раздельно. Аббревиатуры: р – промотор, 3′-конец – 3′-конец гена или сайта сигнала полиаденилирования. Фиг.7а показывает конструкцию с одним инсертом системы двойного индуцирующего возмещаемого блокирования функции (RBF). Фиг.7b показывает конструкцию с двумя инсертами двойного сегрегирующего возмещаемого блокирования функции (RBF). Фиг.8 является схематическим изображением, показывающим гибридизацию трансгенных растений, несущих трансгенные инсерты (TI) системы сегрегирующего возмещаемого блокирования функции (S-RBF) (описанной в примерах 4, 5 и 6 и представленной на фиг.4, 5 и 7b). Показаны хромосомы, несущие блокирующую конструкцию (В), связанную с представляющим интерес трансгеном (Т), а также восстанавливающую конструкцию (R), помещенную в другой неаллельной хромосоме. Хромосомы дикого типа (WT) показаны без маркировок ВТ или R. Родительские растения (Р), участвующие в гибридизации, показаны в первом ряду. Состояние генов, участвующих в восстанавливаемом блоке функций (RBF), не изменяется в результате межлинейного скрещивания. Внутрилинейное потомство F1 имеет гомозиготный генотип для возмещаемого блокирования функции (RBF) и представляющего интерес трансгена (TGI). Внешняя регуляция возмещаемого блокирования функции подразумевает поддержание гомозиготного состояния трансгенов посредством внутрилинейной гибридизации. В случае внешней гибридизации генотипы первого гибридного потомства F1 являются гетерозиготными в отношении всех трансгенных конструкций. Растения остаются живыми. Сегрегирующий восстанавливаемый блокирование функции (RBF) начинает действовать с гибридного потомства F2 в случае внешней гибридизации. В гибридах F2 только половина растений несет блокирующую конструкцию (В), связанную с представляющим интерес трансгеном (Т). Половина из них будет не способна к размножению вследствие отсутствия восстанавливающей конструкции (R). Таким образом, начиная со второго внешнего гибридного потомства, 50% негативный отбор исключает представляющий интерес трансген из природных популяций. В случае сегрегирующего возмещаемого блокирования функции (RBF) восстанавливающая конструкция (R) может свободно расщепляться в геномах растений дикой популяции. Фиг.9 представляет молекулярные конструкции, используемые в системе возмещаемого блокирования функции (RBF) и описанные в примере 7. Эти конструкции представлены в деталях (верхний ряд) для демонстрации принципов помещения генов и на более общем уровне (средний и нижний ряды). Блоки промоторов, представленные остриями стрелок, показывают направление этих генных последовательностей. Конструкции I и II и III являются инсертами в различных неаллельных хромосомах и показаны раздельно. Аббревиатуры: р – промотор, 3′-конец – 3′-конец гена или сайта сигнала полиаденилирования; ВС1 – первая блокирующая конструкция; ВС2 – вторая блокирующая конструкция; RC1 – первая восстанавливающая конструкция; RC2 – вторая восстанавливающая конструкция. Фиг.10 представляет систему тройного возмещаемого блокирования функции (T-RBF). Готовый к использованию реципиентный размножающийся половым способом многоклеточный организм (SRMO) несет три трансгенных инсерта (TI) или ДНК-кассеты, которые локализованы (каждая) в отдельных неаллельных хромосомах в реципиентной линии размножающегося половым способом многоклеточного организма (SRMO). В первом трансгенном инсерте (TI-I) находятся первая блокирующая конструкция (ВС-1) и вторая восстанавливающая конструкция (RC-1), во втором трансгенном инсерте (TI-II) – вторая блокирующая конструкция (ВС-2) и первая восстанавливающая конструкция (RC-1). Указанные первый и второй трансгеные инсерты (TI-I и TI-II) помещены в двух различных неаллельных хромомсомах. Представляющий интерес трансген (трансгены) (TGI) клонируют в поликлонирующий сайт между двумя блокирующими конструкциями (ВС-1 и ВС-2) с образованием трансгенного инсерта III, который трансформируют в третью неаллельную хромосому реципиентной линии размножающегося половым способом многоклеточного организма (SRMO). Стрелки и острия стрелок указывают взаимодействия между блокирующими и восстанавливающими конструкциями (ВС & RC). Первая восстанавливающая конструкция (RC-1) контролирует блокирующую конструкцию (ВС-1) в первом и третьем трансгенном инсертах (TI-I и TI-III), а вторая восстанавливающая конструкция (RC-II) контролирует вторую блокирующую конструкцию (ВС-II) во втором трансгенном инсерте (TI-II) и в третьем трансгенном инсерте (TI-III). Если полученный размножающийся половым способом многоклеточный организм (SRMO) гибридизуется с дикими родственными формами, хромосомы будут разделяться в соответствии с законом Менделя и частотой 7/8 в каждом поколении. Фиг.11 является схематическим изображением гибридизации трансгенных растений/размножающихся половым способом многоклеточных организмов (SRMO), несущих трансгенные инсерты (TI) системы тройного возмещаемого блокирования функции (T-RBF). Конструкции первого возмещаемого блокирования функции (RBF) изображены как В1 и R1 для блокирующей и восстанавливающей конструкций, соответственно. Конструкции В2 и R2 принадлежат второму возмещаемому блокированию функции (RBF). Блокирующая и восстанавливающая конструкции помещены в различных неаллельных хромосомах в противоположном порядке и связаны в парах R1+B2 и R2+B1. Представляющий интерес трансген (Т) помещен в третьей неаллельной хромосоме между первой (В1) и второй (В2) блокирующими конструкциями. Внутрилинейное скрещивание поддерживает гомозиготный генотип обеих конструкций восстанавливаемых блокирований функции (RBF) и представляющего интерес трансгена. Растения этого потомства способны размножаться. Сегрегация конструкций в потомстве внешних гибридов F2 приводит к сильному негативному отбору размножающегося половым способом многоклеточного организма (SRMO), несущего представляющий интерес трансген (Т). Ни одна из трансгенных конструкций не может свободно гибридизоваться с дикими родственными формами. Потомство F2, нарисованное в блоке, способно к половому размножению. Подробное описание изобретения Термины, используемые в изобретении. В данном изобретении большинство используемых терминов имеют то же самое значение, которое они обычно имеют в областях способов рекомбинантных ДНК, молекулярной биологии и в селекции растений и животных. Однако некоторые термины используются несколько отличающимся образом и объясняются более подробно ниже. Термин «система возмещаемого блокирования функции (RBF)» означает концептуальную молекулярную систему или молекулярный механизм контроля, включающий в себя одну или несколько ДНК-конструкций или трансгенных инсертов (TI), которые содержат одну или несколько блокирующих конструкций (ВС), имеющих нуклеотидную или ДНК-последовательность, способную блокировать функцию, которая является существенной для выживания или размножения трансгенного размножающегося половым способом многоклеточного организма (SRMO), а также контролируемые пользователем средства для восстановления блокированной функции, т.е. инструмента восстановления. Система возмещаемого блокирования функции (RBF) выполняет контроль сегрегации (расщепления) и предотвращает утечку в окружающую среду представляющего интерес трансгена (трансгенов) (TGI) в размножающихся половым способом многоклеточных организмах, которые включают в себя как растения, так и животных (кроме человека), к которым применима эта система возмещаемого блокирования функции (RBF). Трансгенный инсерт (TI) системы возмещаемого блокирования функции (RBF) вводят в размножающиеся половым способом многоклеточные организмы (SRMO) вместе с представляющими интерес трансгенами (TGI) известными per se способами трансформации. Следует отметить, что некоторые системы возмещаемого блокирования функции (RBF), определенные ниже, не являются настолько новыми, как таковые, без включения нового признака данного изобретения, которым является либо помещение блокирующих конструкций (ВС) в интроне представляющего интерес трансгена (TGI), либо помещение двух блокирующих конструкций (ВС) на каждой стороне представляющего интерес трансгена (TGI), либо в системе множественного возмещаемого блокирования функции (M-RBF). Термин “система возмещаемого блокирования функции с задержанным действием (D-RBF) Модельные системы возмещаемого блокирования функции (RBF) могут быть подразделены на следующие типы в соответствии с механизмом действия и построением структуры этих конструкций. Если система возмещаемого блокирования функции (RBF) содержит только блокирующую конструкцию (ВС) и ее действие компенсируется прилагаемым извне восстановлением, то эта система является системой простого возмещаемого блокирования функции (RBF). Если восстанавливающая конструкция (RC) активируется в ответ на внешний стимул и высвобождает функцию, прекращенную блокирующей конструкцией (ВС), экспрессируемой под контролем отличающегося промотора, эта модель является общей и полной, называемой здесь системой индуцируемого возмещаемого блокирования функции (RBF). Если восстанавливающая конструкция (RC) помещена в другую неаллельную хромосому, эта модель является системой сегрегирующего возмещаемого блокирования функции (RBF) (S-RBF). Система обращенного сегрегирующего возмещаемого блокирования функции (RS-RBF) содержит два различных трансгенных инсерта (TI) системы возмещаемого блокирования функции (RBF). Они расположены в различных неаллельных хромосомах напротив друг друга, так что блокирующая конструкция (ВС) первого возмещаемого блокирования функции (RBF) связана с восстанавливающей конструкцией второй системы возмещаемого блокирования функции (RBF), a блокирующая конструкция (ВС) второй системы возмещаемого блокирования функции (RBF) связана с восстанавливающей конструкцией (RC) первой системы возмещаемого блокирования функции (RBF). Термин «компенсируемое извне или простое возмещаемое блокирование функции (RBF)» означает систему возмещаемого блокирования функции (RBF), которая состоит только из блокирующей конструкции (ВС). Блокирующая конструкция (ВС) может быть конститутивно активной, как описано в примере 1, специфической для стадии развития, как описано в примере 2, или органоспецифической. Этот инструмент восстановления функции включает в себя компенсацию извне требуемого метаболита, например аминокислоты, как описано в примере 1, гормона, как описано в примере 2, или какого-либо другого метаболита. В этом случае восстанавливающая конструкция (RC) отсутствует. Термин «индуцируемое возмещаемое блокирование функции (I-RBF)» означает систему возмещаемого блокирования функции (RBF), которая состоит как из блокирующей конструкции (ВС), так и восстанавливающей конструкции (RC). Экспрессия блокирующей конструкции (ВС) может быть конститутивной или органоспецифической или специфической для стадии развития, как описано в примере 3. Восстанавливающая конструкция (RC) может быть отвечающей на внешний физический стимул, как показано в примере 3, или на химический стимул, как описано в WO 94/03619. Физически индуцируемое возмещаемое блокирование функции (I-RBF) данного изобретения отличается от химически индуцируемого возмещаемого блокирования функции (I-RBF), описанного в WO 94/03619, также и тем фактом, что блокирующая конструкция помещена в интроне представляющего интерес трансгена (TGI). «Восстанавливающая конструкция (RC)» может быть помещена в той же самой хромосоме, что и блокирующая конструкция (ВС), с образованием «системы индуцируемого возмещаемого блокирования функции (I-RBF)». Восстанавливающая конструкция (RC) может быть помещена в той же самой хромосоме или в хромосоме, которая является аллельной с представляющим интерес трансгеном (TGI) и/или блокирующей конструкцией (ВС), как описано в WO 00/37660, или она может быть помещена в другой, неаллельной хромосоме, что приводит к системе сегрегирующего возмещаемого блокирования функции (S-RBF). Термин «возмещаемое блокирование функции (RBF) с задержанным действием» или «сегрегирующее возмещаемое блокирование функции (S-RBF)» означает систему возмещаемого блокирования функции (RBF), содержащую блокирующую конструкцию (ВС) и восстанавливающую конструкцию (RC), которые расположены в различных неаллельных хромосомах в одном и том же индивидуальном организме. Процесс восстановления функции или вмешательства в эту систему предусматривает внутрилинейное скрещивание для поддержания гомозиготных состояний генов возмещаемого блокирования функции (RBF) и для предотвращения разделения блокирующей и восстанавливающей конструкций (RC&BC). Термин «система обращенного возмещаемого блокирования функции с задержанным действием (RD-RBF)» или «система обращенного сегрегирующего возмещаемого блокирования функции с задержанным действием (RS-RBF)» означает, что дополнительная блокирующая конструкция (ВС) связана с первой восстанавливающей конструкцией (RC1) в первом трансгенном инсерте (TI) и эта дополнительная блокирующая конструкция (ВС) восстанавливается второй восстанавливающей конструкцией (RC2), связанной с представляющим интерес трансгеном (TGI), и первой блокирующей конструкцией (BC1) во втором трансгенном инсерте (TI). Эта система обращенного сегрегирующего возмещаемого блокирования функции с задержанным действием (RS-RBF) контролирует высвобождение как блокирующих, так и восстанавливающих конструкций (BC&RC) в окружающую среду. Система сегрегирующего возмещаемого блокирования функции (S-RBF) не работает в первом поколении внелинейной гибридизации (ауткроссинга), так как она будет присутствовать в гетерозиготном состоянии, показанном как BrRr (где b и r являются рецессивными аллелями, которые не содержат блокирующей конструкции (ВС) или восстанавливающей конструкции (RC), соответственно). Таким образом, обе конструкции действуют, как в гомозиготной родительской линии. Система сегрегирующего возмещаемого блокирования функции (S-RBF) начинает действовать со второго поколения ауткроссинга, когда все гибриды Bbrr будут погибать или имеют измененное свойство вследствие отсутствия восстанавливающей конструкции (RC). Эта система возмещаемого блокирования функции (RBF) предполагает 50% негативный отбор представляющего интерес трансгена (TGI), связанного с блокирующей конструкцией (ВС), в каждом гибридном поколении после первого гибридного потомства. Система сегрегирующего возмещаемого блокирования функции (S-RBF) предполагает признание того факта, что восстанавливающая конструкция (RC) может высвобождаться в окружающую среду. Термин «блокирующая конструкция (ВС)» означает ДНК- или нуклеотидную последовательность, которая является жизненно важной или незаменимой для выживания и/или полового размножения трансгенного организма-хозяина. Блокирующая конструкция (ВС) может быть одновременно представляющим интерес трансгеном (TGI), образуя самоподдерживающую организацию. Предпочтительно, блокирующие конструкции (Вс) являются другими, чем представляющие интерес трансгены (TGI), но они должны быть тесно связаны с представляющими интерес трансгенами (TGI). Блокирующая ДНК- или нуклеотидная последовательность в блокирующей конструкции (ВС) ответственна за блокирование конкретной, молекулярной или физиологической функции в организме-хозяине. Она содержит нуклеотидную (ДНК или РНК) последовательность, например ген, действие которого приводит к исчезновению или стерильности организма-хозяина. Таким образом, размножающийся половым способом многоклеточный организм (SRMO), т.е. растение или животное, становится неспособным к половому размножению при неманипулированных, природных условиях, если он ускользает в окружающую среду. Указанные нуклеотидные последовательности этих блокирующих конструкций (ВС) вместе со средствами для восстановления, т.е. восстанавливающими инструментами, контролируют перенос представляющих интерес трансгенов (TGI) в следующие поколения и, следовательно, распространение этого гена (этих генов) в окружающие природные или культивируемые популяции. Блокирующая конструкция (ВС) предпочтительно связана с представляющим интерес трансгеном (TGI), в крайнем случае она помещена в интроне представляющего интерес трансгена (TGI), фланкирующем его. Необходимо, чтобы по меньшей мере одна, предпочтительно две, блокирующие конструкции (ВС) были расположены в одном и том же трансгенном инсерте (TI) в качестве представляющего интерес трансгена (TGI). Все эти конструкции должны присутствовать в одном и том же индивидууме. Блокирующая конструкция (ВС) должна экспрессироваться в организме-хозяине либо конститутивно, либо органоспецифическим, специфическим для стадии развития или пространственно-временным образом. Термин “блокирующая конструкция” является синонимом терминов “репрессируемый летальный ген”, “ген-разрушитель” или “ген-терминатор”, которые использовались в предыдущих публикациях WO 94/03619, WO 00/37660 или US 5723765. Термин “обращенное возмещаемое блокирование функции с задержанным действием (RD-RBF)” или “обращенное сегрегирующее возмещаемое блокирование функции с задержанным действием (RS-RBF)” означает систему, которая контролирует высвобождение как блокирующей конструкции (ВС) (связанной с представляющим интерес трансгеном (TGI)), так и восстанавливающей конструкции (RC). Сегрегация (расщепление) системы возмещаемого блокирования функции (RBF) показана схематически на фиг.11. Восстанавливающая конструкция (RC) содержит другой блокирующий ген, который контролирует высвобождение восстанавливающей конструкции (RC). Действие этого блокирующего гена восстанавливается второй восстанавливающей конструкцией (RC), которая связана с первой блокирующей конструкцией (ВС) и представляющим интерес трансгеном (TGI). На фиг.11 трансгенный инсерт системы возмещаемого блокирования функции (RBF), во этих аллелях, для удобства обозначен как Т для представляющего интерес трансгена, В1 используют для первой блокирующей конструкции, действующей в комбинации с R1, которая является первой восстанавливающей конструкцией, и В2 для второй (отличающейся от первой) блокирующей конструкции, действующей в комбинации с R2, которая является второй восстанавливающей конструкцией. Аллельные хромосомы B1B1TTR2R2 расположены в одной паре аллельных (сестринских) хромосом, a R1R1B2B2 расположены в другой паре аллельных хромосом. Первый гибрид ауткроссинга будет нести генотипы B1b1TtR2r2 и R1r1B2b2, соответственно. Таким образом, начиная со второго поколения ауткроссинга блокирующая конструкция В1 будет контролировать высвобождение представляющего интерес трансгена Т, а блокирующая конструкция В2 будет контролировать высвобождение восстанавливающей конструкции R1. Таким образом, система обращенного возмещаемого блокирования функции (RS-RBF) контролирует высвобождение всех трансгенных конструкций из данного растения. Внешний контроль или искусственная обработка включает в себя контролируемое пользователем скрещивание для поддержания гомозиготного состояния трансгенных растений (способом, подобным используемому в обычной системе сегрегирующего возмещаемого блокирования функции (S-RBF)). Вторая восстанавливающая или блокирующая конструкция может быть слита с первой блокирующей или восстанавливающей конструкцией в одних и тех же генных последовательностях следующим образом: В1, слитая с R2, и В2, слитая с R1. Некоторые типы сегрегирующего и обращенного возмещаемого блокирования функции (RBF) описаны в WO 00/376600. Однако не описываются блокирующая конструкция (ВС), инсертированная в интрон представляющего интерес трансгена (TGI), и множественное, в том числе двойное или тройное, возмещаемое блокирование функции (RBF). Термин «внутрилинейное скрещивание» означает скрещивание между двумя гомозиготными организмами, которое обеспечивает гомозиготный организм и, другими словами, поддерживает гомозиготные состояния и является безопасным для окружающей среды вмешательством, обеспечиваемым данным изобретением. Термин «интрогрессия» означает поток генов, которые проходят через репродуктивные барьеры и перемещаются между популяциями посредством скрещивания фертильных гибридов с родительскими популяциями. Как описано выше, целью данного изобретения является увеличение безопасности известных систем возмещаемого блокирования функции (RBF). Эта задача достигнута развитием концепции систем множественного, в том числе двойного и тройного возмещаемого блокирования функции (RBF), а также систем введенных в интрон блокирующих конструкций (ВС). Термин «увеличенный уровень надежности» означает уровень надежности, при котором частота инактивации представляющего интерес трансгена (TGI) является более низкой, чем 10-6, предпочтительно является более низкой, чем 10-8, более предпочтительно является более низкой, чем 10-10, и наиболее предпочтительно является более низкой, чем 10-12, в случаях, когда инактивация блокирующего гена равна 10-6. Двойное возмещаемое блокирование функции (RBF) снижает частоту до 10-12. Двойное возмещаемое блокирование функции (RBF) с индуцируемой восстанавливающей конструкцией (RC) имеет 100% негативный отбор, если не имеет место инактивация. Следует ожидать, что размножающийся половым способом многоклеточный организм (SRMO) теряет представляющий интерес трансген (TGI), тесно связанный с блокирующей конструкцией, со скоростью, которая является более высокой, чем скорость, обычно упоминаемая в предыдущей литературе. Термин «двойное возмещаемое блокирование функции (D-RBF)» означает систему возмещаемого блокирования функции (RBF), которая содержит две блокирующие конструкции (ВС). Предпочтительно две различные блокирующие конструкции (ВС) экспрессируются под контролем различных промоторов. Блокирующие гены связаны с представляющим интерес трансгеном (TGI) с обеих сторон. Данная блокирующая конструкция (ВС) может быть даже инсертирована внутри гена в подходящем интроне (фиг.5). Уменьшается возможность, что действие указанного возмещаемого блокирования функции (RBF) разрушается вследствие кроссинговера, делеции ДНК, механизма с образованием молчащего промотора или других подобных событий. Структурно двойные блокирующие конструкции (ВС) связаны с представляющим интерес трансгеном (TGI), предпочтительно с обеих сторон этого гена. Блокирующие конструкции (ВС) могут состоять из одинаковых или различных генов, запускаемых одним и тем же или различными промоторами. Предпочтительно построение комплексных конструкций ДНК получают при запуске двух различных блокирующих генов различными промоторами и помещении их в дистальных положениях относительно трансгенного инсерта (TI), содержащего один или несколько представляющих интерес трансгенов (TGI). Если блокирующие конструкции (ВС) содержат один и тот же функциональный ген, достаточно одной восстанавливающей конструкции (пример 6). Двойное возмещаемое блокирование функции (D-RBF) может содержать две различные восстанавливающие конструкции (RC), если блокирующие конструкции (ВС) содержат различные функциональные гены. Восстанавливающие конструкции (RC) могут быть помещены в один и тот же или в различные трансгенные инсерты (TI). Таким образом, двойное возмещаемое блокирование функции (D-RBF) может быть основано на индуцируемом или сегрегирующем восстанавливаемом блокировании функции (RBF). Индуцируемое двойное возмещаемое блокирование функции (D-RBF) с единственной восстанавливающей конструкцией (RC) описано в примере 6. Термин “система множественного возмещаемого блокирования функции (M-RBF) Представляющий интерес трансген (Т) помещают между различными блокирующими конструкциями (В1 и В2) таким же образом, как в двойном восстанавливаемом блокировании функции (D-RBF), что приводит к системе, обозначаемой В1ТВ2. Две различные восстанавливающие конструкции (R1 и R2) помещают раздельно в две различные неаллельные (несестринские) хромосомы. Дополнительные блокирующие конструкции связывают с восстанавливающими конструкциями в следующем порядке; R1+B2 и R2+B1. Эти инсерты клонируют в реципиентной линии растения в гомозиготном состоянии в неаллельных (несестринских) хромосомах. Дополнительно, обращенное возмещаемое блокирование функции (RBF), не содержащее представляющий интерес трансген (TGI), включают в линию реципиентного размножающегося половым способом многоклеточного организма (SRMO) и трансформирующий вектор с двойным блокированием размножающегося половым способом многоклеточного организма (SRMO) несет сайт поликлонирования, расположенный между указанными двумя блокирующими конструкциями (ВС). Этот вектор может быть использован для клонирования различных представляющих интерес трансгенов в реципиентную линию размножающегося половым способом многоклеточного организма (SRMO) в третьей неаллельной хромосоме (фиг.10). Механизм тройного возмещаемого блокирования функции (RBF) действует таким же образом, как сегрегирующее возмещаемое блокирование функции (S-RBF). Он начинает работать со второго гибридного потомства. Растения в первом гибридном потомстве, содержащие конструкцию представляющего интерес трансгена (TGI) и способные к половому размножению, составляют только 12,5% от всего гибридного потомства или 25% гибридов от гибридов, несущих представляющий интерес трансген (TGI). Гибридизация этих растений показана схематически на фиг.11. Предпочтительным вариантом системы множественного возмещаемого блокирования функции (M-RBF) является “система тройного возмещаемого блокирования функции (RBF)”, которая описана в примере 7, но квалифицированным специалистом в данной области могут быть сконструированы даже более сложные системы возмещаемого блокирования функции (RBF) с использованием общих принципов, изложенных в описании данного изобретения. Термин “средства восстановления” или “инструмент восстановления” означают способ восстановления размножающегося половым способом многоклеточного организма-хозяина (SRMO) с освобождением его от последствий действия блокирующих конструкций (ВС). Инструмент восстановления может предусматривать одно или несколько вмешательств или обработок. Указанные контролируемые пользователем стадии манипуляции приводят к прямой компенсации извне блокированной функции, которая может возмещать недостаточность конкретной функции. Инструмент восстановления может также быть внешним стимулом, активирующим одну или несколько восстанавливающих конструкций (RC), которые были инсертированы в геном размножающегося половым способом многоклеточного организма (SRMO). Таким образом, инструмент восстановления функции предусматривает по меньшей мере одно внешнее контролируемое пользователем вмешательство, комбинированное с одной или несколькими необязательными “восстанавливающими конструкциями (RC) Термин “восстанавливающая конструкция (RC) Восстанавливающую конструкцию (RC) вводят в геном размножающегося половым способом многоклеточного организма (SRMO) отдельно или вместе с блокирующей конструкцией (ВС) и представляющим интерес трансгеном (TGI) или представляющими интерес трансгенами (TGI). Действие восстанавливающей конструкции (RC) всегда является контролируемым пользователем и регулируемым извне. Восстанавливающая функцию конструкция (RC) не действует, другими словами, эта функция отсутствует во время жизни размножающегося половым способом многоклеточного организма (SRMO) при не подвергнутых обработке или манипуляции природных условиях. Термин “контролируемое пользователем вмешательство” означает, что внешний контроль функции “инструмента восстановления” может быть обеспечен внешним стимулом на чувствительном к нему промоторе. В случае использования системы возмещаемого блокирования функции (RBF), называемой здесь “системой сегрегирующего возмещаемого блокирования функции (S-RBF)”, внешняя регуляция подразумевает внутрилинейное скрещивание трансгенных размножающихся половым способом многоклеточных организмов (SRMO) для поддержания гомозиготного состояния системы возмещаемого блокирования функции (RBF). Термин “SRMO” означает размножающиеся половым способом многоклеточные организмы, включающие как растения, так и животных. Предпочтительными растениями являются цветущие растения, которые согласно таксономической классификации включают в себя как покрытосемянные, так и голосемянные растения, в частности сельскохозяйственные растения, например зерновые. В частности, размножающимися половым способом многоклеточными организмами (SRMO) являются организмы, которые могут самоопыляться или скрещиваться с другими родственными формами дикого типа или культивируемыми как трансгенными, так и нетрансгенными организмами. Предпочтительными животными являются, например, рыба, мелкие ракообразные, улитки, домашняя птица, крупный рогатый скот и т.д. Термин “представляющий интерес трансген (генная или нуклеотидная последовательность) или представляющий интерес трансген (TGI)” означает ДНК- или нуклеотидную последовательность, в том числе последовательности РНК, которые кодируют желаемый генный продукт, т.е. белок или фермент или другие вещества, которые могут включать в себя метаболиты, гормоны, токсины, антибиотики и т.д. и которые могут быть получены в качестве конечных продуктов посредством действия прямых генных продуктов. Указанные представляющие интерес трансгены (TGI) вводят в геном размножающегося половым способом организма (SRMO) с использованием доступных генетической трансформации и/или систем переноса. ДНК-последовательности или представляющие интерес трансгены (TGI) обычно кодируют продукты или молекулы, применимые в сельском хозяйстве, плодоовощном хозяйстве, лесоводстве и/или других промышленных приложениях. Альтернативно, генные продукты имеют некоторые другие возможные применения. Обычно эта ДНК-последовательность является чужеродной или гетерологичной для размножающегося половым способом многоклеточного организма (SRMO), но иногда могут использоваться и вводиться гомологичные последовательности нуклеиновых кислот или ДНК, например, в виде множественных копий, например, в виде тандемных инсертов, для получения необязательных количеств желаемого продукта. Другими словами, ДНК-последовательность может состоять из одного гена или одной нуклеотидной последовательности или их множества. Альтернативно, несколько различных генов могут быть введены в виде мультикопий или тандемных инсертов для облегчения экспрессии сложных метаболических путей. В конкретном варианте данного изобретения сама блокирующая конструкция (ВС) служит в качестве представляющего интерес трансгена (TGI). Термин “блокирование” обозначает механизм молекулярного контроля, например, нуклеотидную последовательность, экспрессия которой блокирует, прекращает или ингибирует функцию, существенную для выживания, роста, развития и/или полового размножения, и способна прекращать развитие молекулярного аппарата данного организма-хозяина на уровне ДНК, мРНК, белка или метаболита. Если этот молекулярный аппарат применяется в чувствительный или уязвимый момент в цикле развития, это приводит к некрозу, изменению фенотипа, раннему или позднему образованию семян или зацветанию, неспособности образования соцветий, цветков или плодоношения, образованию карликового фенотипа или некоторых морфологических изменений, которые предотвращают свободную гибридизацию или интербридинг трансгенных размножающихся половым способом многоклеточных организмов (SRMO) в природных условиях. Термин “природные условия” означает условия выращивания, которые являются обычными параметрами окружающей среды, включающими в себя температуру, влажность, освещение, химический состав почвы во время роста или получения размножающегося половым способом многоклеточного организма (SRMO), в том числе растения или животного в сельском хозяйстве, плодоовощном хозяйстве, лесоводстве, а также в природе. Термин “утечка трансгена в окружающую среду” означает предотвращение утечки трансгена в природу через гибридизацию или скрещивание родительского трансгенного организма с его нетрансгенными родственными формами дикого типа или в культуре. Другими словами, предотвращается нежелательный перенос гена или генов посредством полового скрещивания и высвобождение этого трансгена. Трансгенный организм предохраняется от роста, полового размножения или развития жизнеспособного потомства. Это достигается разрешением блокирующей конструкции (ВС) функционировать при нормальных, природных, неманипулируемых, не подвергнутых обработке или вмешательству условиях. Указанные природные, неманипулируемые условия блокируют, прекращают, разрушают или нарушают функцию, которая является существенной для выживания или размножения размножающегося половым способом многоклеточного организма (SRMO). Посредством расщепления указанная существенная функция размножающегося половым способом многоклеточного организма (SRMO) приводит к исчезновению трансгена в природе. “Восстановление (возмещение) блокированной функции” означает устранение блокирования данной блокированной функции для того, чтобы сделать возможными нормальный рост и половое размножение размножающегося половым способом многоклеточного организма (SRMO) для целей сельского хозяйства, плодоовощного хозяйства, лесоводства, промышленных или любых других целей. Удаление блокирования функции получают предпочтительно искусственными, контролируемыми пользователем средствами, применяемыми извне. Указанные применяемые извне средства включают в себя стадии манипуляции, т.е. обработки или вмешательства, которые применяются сами по себе или через восстанавливающую конструкцию (RC). Термины “внешняя обработка” и “внешняя манипуляция” означают некоторые искусственные вмешательства или действия, которые отличаются от действий, встречающихся природно в местах выращивания сельскохозяйственных культур или в природе. Предпочтительно, они применяются в ограниченных условиях (в закрытом помещении). ОБЩЕЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ Данное изобретение относится к молекулярной биологии и обеспечивает способ и систему для улучшения контроля сегрегации трансгена и распространения его в природную популяцию или в другую популяцию трансгенной или нетрансгенной сельскохозяйственной культуры в окружающей среде. Комплекс ДНК-конструкций, называемый здесь системой возмещаемого блокирования функции (RBF), в противоположность невозмещаемым системам блокирования, известным из предыдущего уровня техники, является ключевым предметом данного изобретения. Блокирование включает в себя применение нуклеотидной последовательности, способной блокировать определенные жизненно важные физиологические функции или связанные с развитием функции размножающихся половым способом многоклеточных организмов (SRMO), в том числе растений и/или животных. Блокированная функция приводит к исчезновению ускользнувшего трансгена, т.е. это блокирование является существенным для выживания данного трансгена. Альтернативно, она приводит к значимому изменению в физиологии растения или животного, приводя к неспособности размножаться данного растения или животного. Блокированная функция растения или животного может быть молекулярной, биохимической или физиологической по природе. Возмещение блокированной функции предусматривает внешнее, искусственное контролируемое пользователем вмешательство, включающее в себя химическую, физическую или механическую обработку, которую выполняют с ДНК-конструкцией или без ДНК-конструкции, позволяющей трансгенному растению или животному преодолеть блокирование данной функции. Одним из применений этой технологии является предохранение трансгена от ускользания (утечки) посредством гибридизации в природных условиях. Данное изобретение относится к областям молекулярной биологии, трансгенной технологии, биохимии и физиологии растений, а также конкретным областям синтеза ДНК и антисмысловой технологии. Таким образом, данное изобретение относится к системе молекулярного контроля с увеличенным уровнем надежности для контроля экспрессии трансгенов или неумышленной утечки трансгенов в окружающую среду. Система молекулярного контроля действует через систему возмещаемого блокирования функции (RBF), заявляемую в данном изобретении, которая предлагает идею связывания представляющих интерес трансгенов (TGI) в вектор или ДНК-кассету, содержащую одну или несколько ДНК-конструкций, способных блокировать конкретную функцию в растении или в животном. Это блокирование приводит к исчезновению или гибели растения или животного или прекращению развития или изменяет фенотип или физиологию растения/животного в природных условиях таким образом, что предотвращается половое размножение. Это блокирование функции является возмещаемым посредством действия восстанавливающего функцию инструмента, который включает в себя внешнюю контролируемую пользователем обработку и необязательно включает в себя по меньшей мере одну ДНК-конструкцию, кодирующую мРНК, белок или ферментный продукт, индуцируемую внешним химическим или физическим стимулом или экспрессирующуюся конститутивно. Таким образом, любой размножающийся половым способом многоклеточный организм (SRMO), в том числе родительская линия или любой ее последующий гибрид, несущие представляющий интерес трансген (TGI), связанный с системой возмещаемого блокирования функции (RBF), будет погибать или останется стерильным в природных условиях, если не будет применено внешнее контролируемое пользователем вмешательство, которое имеет место, например, в случае неумышленного ускользания (утечки) или ауткроссинга. Растения или животные, обеспеченные системой возмещаемого блокирования функции (RBF), могут быть возмещены, т.е. блокирование может быть удалено, посредством конкретной внешней процедуры на определенной стадии развития. Система возмещаемого блокирования функции (RBF) является не только конкретной молекулярной конструкцией, но в широком смысле инструментом, который оперирует с тремя концептуальными конструкциями: представляющим интерес трансгеном (TGI), блокирующими конструкциями (ВС) и восстанавливающими функцию конструкциями (RC) и контролируемым пользователем вмешательством. Представляющий интерес трансген (TGI) является ДНК-последовательностью, которая контролируется системой возмещаемого блокирования функции (RBF). Она сообщает организму-хозяину один или несколько признаков, полезных в сельском хозяйстве, промышленном производстве, лесоводстве или плодоовощном хозяйстве. Блокирующие конструкции (ВС) и восстанавливающие функцию конструкции (RC) относятся к системе возмещаемого блокирования функции (RBF) и служат для контроля представляющего интерес трансгена (TGI) в трансгенных размножающихся половым способом многоклеточных организмах (SRMO). Блокирующая конструкция (ВС) блокирует конкретную физиологическую или молекулярную функцию и предотвращает гибридизацию или ауткроссинг растений-хозяев или животных-хозяев, несущих по меньшей мере один представляющий интерес трансген (TGI), связанный с данной блокирующей конструкцией (ВС). Блокирующая конструкция (ВС) содержит ДНК-конструкцию, связанную с представляющим интерес трансгеном (TGI), и кодирует фактор, способный блокировать некоторую молекулярную или физиологическую функцию. Это блокирование приводит к гибели или изменению фенотипа или физиологии трансгенного размножающегося половым способом многоклеточного организма (SRMO), приводя к неспособности к половому размножению. Блокирующая конструкция (ВС) может действовать через ДНК-рекомбиназу, антисмысловую мРНК, рибонуклеазу, токсин, продуцирование гормонов, ферментативное действие или некоторые другие молекулярные механизмы. Блокирование конкретной функции может также включать в себя любое изменение в молекулярном аппарате, приводящее к некоторым фенотипическим или физиологическим изменениям, предотвращающим размножение размножающегося половым способом многоклеточного организма (SRMO) в природных условиях. Это может быть, например, раннее или позднее прорастание семян или цветение, неспособность образования соцветия или плодов, образование карликового фенотипа или некоторое другое морфологическое изменение. Предпочтительными примерами молекулярных механизмов, действующих в блокирующих конструкциях (ВС) являются экспрессия: – фермента необычной органоспецифичности или специфичности в отношении стадии развития; – антисмысловой мРНК фермента, важного для развития или функционирования размножающегося половым способом многоклеточного организма (SRMO); – токсина; – фермента, продуцирующего токсичный метаболит; или – фермента, рекомбинирующего молекулы ДНК, РНК или белка таким образом, который являются нежелательным для размножающегося половым способом многоклеточного организма (SRMO). Последовательность нуклеиновой кислоты, присутствующая в блокирующей конструкции (ВС), должна предпочтительно экспрессировать фермент, который характеризуется разрушением или сверхпродуцированием химического вещества, такого как метаболит с необычной органоспецифичностью или специфичностью в отношении стадии развития. Такими ферментами являются, например, L-аспарагиназа или L-глутаминаза. Блокирующая конструкция (ВС) может также содержать последовательность нуклеиновой кислоты, кодирующую антисмысловую мРНК фермента, который является существенным для развития и/или функционирования организма-хозяина, нуклеотидную последовательность, кодирующую нуклеазу или рекомбиназу нуклеиновых кислот. Предпочтительным примером является нуклеотидная последовательность, кодирующая энт-кауренсинтазу А, которая является важной в развитии или плодоношении (образовании плодов) растения-хозяина. Блокирующая конструкция (ВС) может содержать нуклеотидную последовательность, кодирующую протеазу или токсин, например протеинкиназу NPK15, или фермент, продуцирующий токсичное вещество или метаболит, например антибиотик. Альтернативно, блокирующая конструкция (ВС) может содержать последовательность нуклеиновой кислоты, кодирующую нуклеазу или рекомбиназу нуклеиновых кислот. Нуклеотидные последовательности, используемые в блокирующих конструкциях (ВС), предпочтительно являются синтетическими и адаптированными к предпочтительности (преферентивности) в отношении хозяина или к предпочтительности (преферентивности) в отношении интрона, использующими интрон подходящей нуклеотидной последовательности или гена, кодирующего желаемый продукт в организме-хозяине. Предпочтительной синтетической последовательностью в некоторых вариантах данного изобретения является синтетическая нуклеотидная последовательность барназы, содержащая SEQ ID NO:1. Она адаптирована к предпочтительности в отношении хозяина, в данном случае в отношении растения, тогда как SEQ ID NO:3 адаптирована к предпочтительности в отношении интрона гена растения. Некоторые полезные конструкции и/или способы, применимые для возмещаемого блокирования функции (RBF), могут быть разработаны на основе рассматриваемой ниже информации. Антисмысловой способ может быть использован для блокирования ключевой функции растения посредством экспрессии антисмысловой мРНК конкретного фермента, ответственного за данную ключевую функцию. Антисмысловая мРНК может блокировать экспрессию конкретного гена или генов на определенной стадии развития. Например, блокирование экспрессии энт-кауренсинтазы A Arabidopsis приводит к карликовому фенотипу этого растения (Sun and Kamiya, 1994, Plant Cell, 6: 1509-1518). Энт-каурен является ключевой молекулой в биосинтетическом пути гиббереллинов, которые являются важными гормонами растений. Энт-кауренсинтаза А кодируется однокопийным геном и может быть блокирована при помощи антисмыслового способа. Если молекула антисмысловой мРНК экспрессируется в стадии прорастания, отсутствие гиббереллина может компенсироваться наружным применением GА3 (пример 2). Другим примером является блокирование антисмысловой мРНК экспрессии глутаминсинтетазы в прорастающих семенах. Глутаминсинтетаза является критическим ферментом во всех превращениях аминокислот в прорастающих семенах (Temple, et al., 1993, Mol. Gen. Genet. 236: 315-325). Блокирование развития размножающегося половым способом многоклеточного организма (SRMO), в частности растения, может быть достигнуто экспрессией некоторых ферментов, происходящих из бактерий, например L-аспарагиназы (Filpula, et al., 1988, Nucl. Acid Res. 16: 10385) или L-глутаминазы (Ramakrishnan and Joseph, 1996, Canadisn J. Microbiol. 42: 316-325), которые могут быть компенсированы наружным введением этой аминокислоты (пример 1). Та же самая технология может быть применена в ряде других случаев. Примерами применимых технологий являются блокирование синтеза гормонов растений, в том числе блокирование синтеза ауксина, цитокинина или АБК, блокирование образования аминокислоты (аминокислот) или блокирование метаболических путей. Несколько различных типов невозмещаемых блокирований функции, предназначенных для обеспечения контроля ускользания (утечки) трансгенных растений, были описаны различными группами в различных патентных заявках и публикациях. Многие из описанных ранее молекулярных механизмов применимы в данном изобретении. Например, невозмещаемое блокирование развития зародышей может быть достигнуто в Brassica napus экспрессией модифицированного экзотоксина А Pseudomonas aeruginosa под контролем промотора белка напина (Koning et al., 1992, Plant Mol. Biol., 18: 247-258). Стерильность пыльцы может быть достигнута с использованием экспрессии А-цепи дифтерийного токсина под контролем промотора lat52. Экспрессия токсина может применяться для достижения экстирпации клеток в развивающейся пыльце (Twell, 1995, Protoplasma, 187: 144-154). В патенте США US 5498533 описана регуляция развития картофеля экспрессией смысловых и антисмысловых конструкций гена кальмодулина. Экспрессия гена кальмодулина со смысловой ориентацией увеличивала рост побегов и клубней, тогда как растения, несущие антисмысловые конструкции, проявляли уменьшенный рост побегов и клубней. Таким образом, экспрессия антисмыслового гена кальмодулина может быть использована как фактор, блокирующий физиологическую функцию. Имеется ряд генов токсинов или летальных генов, которые способны блокировать определенные функции в растении. Некоторые нуклеазы могут разрушать аппарат синтеза мРНК. Предметами интенсивных исследований были ферменты барназа и рибонуклеаза А. Барназа, происходящая из Bacillus amyloliquefaciens (Hartly, 1989, Trends Biochem. Sci. 14; 450-454), является наиболее известным летальным геном с тех пор, как Mariani, et al. (1990, Nature 347: 737-741) использовал ее для конструирования контроля фертильности в трансгенных растениях. Превосходным признаком системы барназы является ее специфический ингибитор barstar, который эффективно блокирует действие РНКазы в растениях (Mariani, et al., 1992, Nature, 357: 384-387). Таким образом, летальное блокирование функции в растениях, достигаемое экспрессией барназы, может быть устранено экспрессией гена barstar (WO 94/03619, WO 00/37660). Некоторые каталитические или цитолитические летальные белки могут блокировать определенные функции. Рибосомный ингибиторный белок (RIP) из гена RIP сапорина 6 (Barthelemy, et al., 1993, J. Biol. Chem. 268: 6541-6548; GenBank ID SOSAPG, accession No. X15655) Saponaria officinalis непосредственно препятствует экспрессии белков в клетке растения, но не является токсичным для других организмов. Широко обсуждается в данной области дифтерийный токсин А, который является другим эффективном агентом блокирования. Он использовался несколько раз в качестве условного летального маркерного гена в растениях (Czako and An, 1991, Plant Physiol. (Bethesda) 95: 687-692; Nillson, et al., 1998, Plant J. 15: 799-804), так как он приводит к эксцирпации клеток (Van Der Geest, et al., 1996, Plant Physiol. (Rockville) 109: 1151-1158). Удаление действия RIP или дифтерийного токсина А является возможным с использованием антисмыслового способа; имеется также сообщение об эффективном ингибировании дифтерийного токсина мансононом-D (Madhusoodana, et al., 1998, J. Cell. Physiol. 176: 40-49). Мансонон-D, сесквитерпеноид ортонафтохинон, ингибировал цитотоксичность рицина, модекцина, токсина Pseudomonas и токсина Diphtheria и может быть предположительно использован для устранения блокирования, осуществляемого экспрессией этих токсинов в растениях. Было показано, что экзотоксин A Pseudomonas aeruginosa может специфически прекращать развитие зародышей табака (Koning, et al., 1992, Plant Mol. Biol. 18: 247-258). Имеются и другие примеры генов, потенциально применимых в данной области. Патент США US 6022720 описывает белок Вах, который регулирует запрограммированную смерть (апоптоз) клеток млекопитающих. Харпин, продукт гена hrpN Erwinia amylophlora, индуцирует HR в табаке и блокируется обработкой К52а, ингибитором протеинкиназы (Popham, et al., 1995, Physiol. Mol. Plant Pathology, 47; 39-50). NPK15, протеин-серин/треонин-киназа табака, действует как суицидный ген и блокирует пролиферацию клеток-хозяев (Ito, et al. 1994, Mol. Gen. Genet. 245: 1-10). Гены из Т-ДНК Agrobacteria, ответственные за сверхпродуцирование цитокинина или ауксина, могли быть также использованы для изменения фенотипа растений, т.е. ген ipt (Redig, et al., 1997, Physiol. Plantarum, 99: 89-96) или гены auxl и aux2 (Beclin, et al., 1993, Transgenic Res. 2: 48-55; Hamza, et al., 1993, Theor. Appi. Gen. 86: 657-664). Упомянутые выше гены используются в качестве летальных маркеров или суицидных генов, но не было описано возмещение (устранение) блокирования функции. В данном изобретении несколько генов могут быть использованы для получения трансгенных инсертов (TI) для сборки систем возмещаемого блокирования функции (RBF), когда возмещение блокирования проводят антисмысловым способом или экспрессией гена блокирующей конструкции (ВС) под контролем промотора, репрессируемого белком, кодируемым восстанавливаемым геном. Все эти способы являются доступными для специалиста с квалификацией в данной области. Нуклеотидные последовательности, получаемые для применения в блокирующих конструкциях (ВС), предпочтительно являются синтетическими и адаптированы в отношении предпочтительности (преферентивности) хозяина, например предпочтительности растения, и/или в отношении предпочтительности интрона, в частности, подходящего, достаточно большого интрона, присутствующего в представляющем интерес трансгене (TGI), кодирующем желаемый продукт. Восстанавливающая функцию конструкция (RC), которая необязательно зависит от системы возмещаемого блокирования функции (RBF), используемой в данном изобретении, восстанавливает блокированную функцию в том случае, если применяется обработка извне, и не работает в природных условиях. Восстанавливающая конструкция (RC) содержит нуклеотидную последовательность, обеспечивающую активирующий или гасящий механизм экспрессии гена или нуклеотидной последовательности в восстанавливающей конструкции (RC). Эти нуклеотидные последовательности способны экспрессировать, например, антисмысловую РНК блокирующей нуклеотидной последовательности. Нуклеотидная последовательность восстанавливающей конструкции (RC) может быть, например, антисмысловой РНК, которая по существу похожа на ген NPK15, или нуклеотидной последовательностью, способной экспрессировать белок, который может инактивировать белок, экспрессируемый блокирующей конструкцией (ВС). В типичном примере инактивирующим белком является белок barstar, который инактивирует нуклеазу барназу, связываясь с ней. Восстанавливающая функцию конструкция (RC) может содержать нуклеотидную последовательность, способную экспрессировать фермент, продуцирующий метаболит, способный компенсировать недостаточность, произведенную действием блокирующей конструкции (ВС). Альтернативно, восстанавливающая конструкция (RC) может содержать нуклеотидную последовательность, способную экспрессировать фермент, который превращает токсическое вещество или токсический метаболит, продуцируемый блокирующей конструкцией (ВС), в безвредный продукт. Такой пример обеспечивается нуклеотидной последовательностью, кодирующей фосфаттрансферазу, способную превращать антибиотическое вещество, такое как гигромицин. Кроме необязательной восстанавливающей конструкции (RC) средства для восстановления функции или восстанавливающий функцию инструмент включают в себя контролируемое пользователем вмешательство с использованием, например, наружной компенсации при помощи химического вещества, например метаболита, репрессии промотора блокирующей конструкции, способной погасить экспрессию мРНК блокирующего гена с использованием способа антисмысловой РНК, экспрессии белка, способного инактивировать белок блокирующей конструкции (ВС) специфическим связыванием, протеолизом или другими взаимодействиями белок-белок, экспрессии фермента, способного превращать токсическое вещество в безвредное вещество, экспрессии вещества или метаболита, способного компенсировать или инактивировать блокирующее токсическое вещество или взаимодействовать с блокирующим токсическим веществом, метаболитом, а также других молекулярных механизмов, которые могут регулироваться извне. Восстанавливающий функцию фактор или средства, удаляющие блокирование функции, могут быть наружным контролируемым пользователем вмешательством, включающим в себя химические, физические или механические средства. Применимыми химическими средствами являются, например, аминокислоты, гормоны, салициловая кислота, тетрациклин и т.д. Физические средства или факторы включают в себя температуру, свет, осмос, гравитацию, влажность. Механические средства включают в себя, например, межлинейную гибридизацию или поддержание гомозиготных состояний. Восстанавливающий функцию фактор или средства с устраняющей блокирование функцией, применимые под контролем пользователя, могут включать в себя регулируемые извне трансгенные последовательности, такие как рецепторы, антисмысловые РНК, границы последовательностей, репрессоры или протеолитические белки, включающие в себя ферменты или их продукты. Наружный контроль может быть стимулом, активирующим чувствительный промотор или межлинейное скрещивание для поддержания гомозиготного состояния представляющих интерес трансгенов (TGI) в случае системы сегрегирующего возмещаемого блокирования функции (S-RBF). Восстановление блокированной функции может выполняться различными путями и зависит от конкретной модели системы возмещаемого блокирования функции (RBF). Для удобства эти инструменты или средства подразделяют на основе типа восстановления функции на несколько групп следующим образом: компенсация извне блокированной функции, восстановление посредством репрессии действия блокирующей конструкции (ВС) на уровне ДНК, восстановление с использованием способа антисмысловой РНК, восстановление на уровне взаимодействия белок-белок и на уровне ферментов или других метаболитов. Наружные контролируемые пользователем средства включают в себя следующие вмешательства: – индукцию извне чувствительного восстанавливающего функцию гена/белка; – экспрессию конститутивно нуклеотидной последовательности восстанавливающей функцию конструкции; – нанесение наружно вещества или метаболита для компенсации недостаточности, вызванной блокирующей конструкцией (ВС); поддержание гомозиготного состояния восстанавливающей конструкции (RC), блокирующей конструкции (ВС) и представляющего интерес трансгена (TGI) при помощи внутрилинейного скрещивания; – индукцию восстанавливающей конструкции (RC) с использованием химикалия, активирующего индуцируемый промотор восстанавливающей конструкции (RC); и/или – индукцию восстанавливающей конструкции (RC) с использованием физического фактора (тепла, холода, осмотика и т.д.), активирующего индуцируемый промотор восстанавливающей конструкции (RC). Средства для компенсации блокирования включают в себя наружное применение вещества, метаболита или гормона для компенсации недостаточности, вызываемой блокирующей конструкцией (ВС). Такими веществами являются, например, гиббереллины, которые могут предоставляться растениям извне, или антисмысловая мРНК энт-кауренсинтазы А. В качестве средства для восстановления блокированной функции может быть также использована экспрессия репрессорного пептида или белка, связывающегося с последовательностью оперона, который контролирует экспрессию блокирующей нуклеотидной последовательности. Примеры применимых репрессоров и операторов включают в себя tet-репрессорный белок Tn10 и tet-операторная ДНК-последовательность. Экспрессия антисмысловой РНК блокирующей нуклеотидной последовательности может быть использована для активации гасящего механизма экспрессии блокирующей конструкции (ВС). Эта экспрессия предусматривает, например, экспрессию антисмысловой РНК гена NPK15. Далее, экспрессия белка, инактивирующего белок, экспрессируемый блокирующей конструкцией (ВС), также применима в качестве средства для восстановления блокированной функции. Белком, инактивирующим белок, экспрессируемый блокирующей конструкцией (ВС), является, например, белок barstar, который инактивирует нуклеазу барназу связыванием с ней. Экспрессия фермента, продуцирующего метаболит, способный компенсировать недостаточность, вызываемую действием блокирующей Km и (ВС), является применимым средством для восстановления функции. Экспрессия фермента, способного превращать токсическое вещество или токсический метаболит, продуцируемый блокирующей конструкцией (ВС), в безвредный продукт, является возможным средством для восстановления функции. Применимый пример обеспечивается ферментом, который преобразует свойства антибиотика. В качестве примера может быть упомянута фосфаттрансфераза антибиотика гигромицина. Компенсация извне блокированной функции может быть использована в случае возмещаемого блокирования функции (RBF), приводящего к отсутствию вещества, метаболита или гормона. Отсутствие этого вещества или этой молекулы может быть компенсировано добавлением извне той же самой или подобной молекулы, например, блокированием экспрессии энт-кауренсинтазы в прорастающих семенах (пример 2). Экспрессия L-аспарагиназы или L-глутаминазы приводит к отсутствию аспарагина/глутамина (Temple, et al., 1993, Mol. Gen. Genet. 236: 315-325), но это отсутствие может быть компенсировано добавлением извне глутамина во время проращивания семян (пример 1). Восстановление функции репрессией действия блокирующей конструкции (ВС) на уровне ДНК может быть применено к любому блокирующему гену. Репрессия этой экспрессии выполняется конкретной ДНК-последовательностью, имеющей пограничный репрессорный элемент. Репрессорный элемент может быть белком, РНК или ДНК. Он может быть любой ДНК-последовательностью, экспрессия которой предотвращает экспрессию блокирующей конструкции (ВС). Эта репрессорная молекула должна экспрессироваться из восстанавливающей конструкции (RC), помещенной под контроль индуцируемого промотора. Например, известный tet-репрессорный белок Tn10 (Gatz and Quail, 1988, Proc. Natl. Acad. Sci. USA, 85: 1394-1397; Gatz, et al., 1991, Mol. Gen. Genet. 227: 229-237) может служить в качестве восстанавливающего функцию элемента в случае, когда ДНК-последовательность tet-оперона, ответственная за связывание с этим репрессорным белком, находится внутри промотора данной блокирующей конструкции (ВС) (пример 7). Восстанавливающая функцию конструкция (RC), экспрессирующая репрессорный белок, может быть интегрирована в другую неаллельную хромосому и служить в качестве системы сегрегирующего возмещаемого блокирования функции (S-RBF) (пример 7). Восстановление функции с использованием способа антисмысловой РНК применяется к большинству сообщенных условных летальных маркерных генов, действующих в качестве системы возмещаемого блокирования функции (RBF), где функцию восстанавливают при помощи антисмыслового способа. Экспрессируемая мРНК минус-цепи блокирующего гена запускает механизм гашения (образования молчащего гена), который спасает размножающийся половым способом многоклеточный организм-хозяин от ингибирования определенной функции. Если белок токсина имеет ингибиторный белок или продуцируемый ферментативно ингибиторный метаболит, восстановление функции обеспечивается на уровне белка. Этим ингибитором может быть белок, связывающийся с токсином или инактивирующий его протеолитически. Наиболее известным примером этого взаимодействия является механизм барназа-barstar (примеры 3-7). Ингибитор, согласно данному изобретению, может быть некой ферментативно продуцируемой молекулой, способной инактивировать действие токсина. Например, если блокирующим фактором является сверхпродуцирование вещества или метаболита, восстановление может выполняться молекулой-аналогом, которая способна блокировать фермент, сверхпродуцирующий это вещество или метаболит. Если токсин является веществом или метаболитом, продуцируемым ферментом, кодируемым блокирующим геном, ингибитором может быть фермент, инактивирующий данное токсическое вещество. Например, блокирование могло бы быть экспрессией антибиотика, тогда как ингибирование его действия, устраняющее блокирование, могло бы достигаться, например, действием специфической фосфаттрансферазы данного антибиотика. Средства для восстановления блокированной функции обеспечиваются наружным применением вещества для компенсации недостаточности, вызываемой блокирующей конструкцией. Предпочтительным вариантом является обеспечение гиббереллинов для ДНК-конструкций, экспрессирующих антисмысловую мРНК энт-кауренсинтазы А, или экспрессии репрессорного пептида/белка, связывающегося с последовательностью оперона, который контролирует экспрессию блокирующего гена, например tet-репрессорного белка Tn10, связывающегося с ДНК-последовательностью tet-оператора. Взаимосвязь между тремя концептуальными конструкциями данного изобретения, т.е. представляющим интерес трансгеном (TGI), блокирующей конструкцией (ВС) и восстанавливающей функцию конструкцией (RC), является очень важной для обеспечения увеличенного уровня надежности контроля, который является основной целью данного изобретения. Блокирующая конструкция (ВС) и представляющий интерес трансген (TGI) должны быть тесно связаны вместе. Поскольку блокирующая конструкция (ВС) контролирует утечку представляющего интерес трансгена (TGI), эти конструкции должны быть связаны вместе так тесно, как это возможно, для минимизации или полного исключения возможности кроссинговера. Эта конструкция должна быть локализована по меньшей мере в одном и том же трансгенном инсерте (TI) и в одном и том же индивидуальном организме, предпочтительно в интроне представляющего интерес трансгена (TGI). Если используются более чем одна блокирующая конструкция, они должны быть помещены непосредственно на каждой стороне трансгенного инсерта (TI), содержащего представляющие интерес трансгены (TGI), помещенные между двумя блокирующими конструкциями (ВС). Они должны быть чрезвычайно близко связаны, как только это возможно. Для минимизации возможности разделения блокирующей конструкции (ВС) и представляющего интерес трансгена (TGI) блокирующий ген может быть инсертирован в интрон представляющего интерес трансгена (TGI). Это было описано в примере 5. В этом случае большие мутации блокирующей конструкции (ВС) являются почти невозможными без одновременного разрушения представляющего интерес трансгена (TGI). Инсертирование гена барназы (блокирующей конструкции (ВС)) в интрон представляющего интерес трансгена (TGI) (uidA или GUS) в противоположной ориентации показано на фиг.5 и 6. Промотор блокирующей конструкции (ВС) совпадает с сигналом полиаденилирования представляющего интерес трансгена (TGI). Второй экзон гена uidA совпадает с 3′-UTP барназы. Кодирующая последовательность барназы локализована в интроне гена uidA. Сигналы полиаденилирования барназы частично совпадают с этим интроном и частично с экзоном гена uidA. Встроенная в интрон блокирующая конструкция (ВС) служит в качестве средства увеличения надежности молекулярного контроля ускользания трансгена. ДНК-конструкция может содержать, например, синтетическую нуклеотидную последовательность, адаптированную для инсертирования в блокирующую конструкцию (ВС), встроенную в интрон представляющего интерес трансгена (TGI). Интрон представляющего интерес трансгена (TGI) является, например, адаптированным для нуклеотидной последовательности блокирующей конструкции (ВС), которую помещают в интрон в противоположной ориентации относительно представляющей интерес нуклеотидной последовательности. Пример этого обеспечивается при инсертировании синтетической нуклеотидной последовательности, по существу идентичной последовательности гена барназы и имеющей SEQ ID NO:3. Предпочтительным примером комплексной ДНК-конструкции, используемой в данном изобретении, является синтетическая последовательность генов барназы и barstar, которые адаптированы для экспрессии в растении, SEQ ID NO:1 и SEQ ID NO:2, соответственно. Указанная комплексная ДНК-конструкция дополнительно характеризуется тем, что она имеет сайт поликлонирования, показанный в SEQ ID NO:4, сконструированный с открытой рамкой считывания (ORF), адаптированной для экспрессии В предпочтительном варианте данного изобретения, двойном возмещаемом блокировании функции (D-RBF) и тройном возмещаемом блокировании функции (T-RBF), трансгенный инсерт (TI), содержащий один или несколько представляющих интерес трансгенов (TGI), помещают между двумя блокирующими конструкциями (ВС), снабженными сайтом поликлонирования для удобного введения различных представляющих интерес трансгенов (TGI) для инсертирования в подходящие реципиентные клетки. Блокирующая конструкция (ВС) или фактор, вызывающий блокирование, может содержать любую нуклеотидную последовательность, кодирующую рецептор, антисмысловую РНК, нуклеазу, токсин, фермент или другой продукт. Блокирующая нуклеотидная последовательность может быть в уникальных ситуациях, когда блокирующий ген имеет промышленно ценное свойство, заключающееся в том, что он является представляющим интерес геном (TGI), образуя самоподдерживающую организацию. Обычно блокирующая конструкция (ВС) и представляющие интерес трансгены (TGI) не являются одними и теми же. Восстанавливающую конструкцию (RC) предпочтительно помещают в тот же самый организм или размножающийся половым способом многоклеточный организм (SRMO), а не в другой индивидуальный организм, как это описано в некоторых вариантах предыдущего уровня. Восстанавливающая конструкция (RC) и блокирующая конструкция (ВС) могут быть локализованы тесно связанными вместе или находиться в транс-положении, даже в разных хромосомах, в зависимости от типа возмещаемого блокирования функции (RBF). В том случае, когда восстановление функции при возмещаемом блокировании функции (RBF) предусматривает наружную компенсацию недостатка конкретного метаболита, нет необходимости в восстанавливающей конструкции (RC). В системе сегрегирующего возмещаемого блокирования функции (RBF) восстанавливающая конструкция (RC) должна находиться в неаллельной хромосоме, отличающейся от неаллельной хромосомы, несущей блокирующую конструкцию (ВС). Экспрессия блокирующей конструкции (ВС) может быть конститутивной или временной со специфичностью в отношении органа или стадии развития. Экспрессия восстанавливающей конструкции (RC) может быть чувствительной к наружному стимулу или эта блокирующая конструкция может быть конститутивно экспрессируемой, как в системе сегрегирующего возмещаемого блокирования функции (RBF). Конститутивная экспрессия блокирующей конструкции (ВС) приводит к наивысшему контролю экспрессии трансгена. Механизм восстановления функции зависит от наружной обработки или стимула. Если данное ингибирование включает в себя отсутствие вещества, метаболита или гормона, восстановление функции такого конститутивно экспрессируемого блокирования может выполняться наружной компенсацией недостатка данного соединения (пример 1). Если это восстановление предусматривает экспрессию продукта под контролем индуцируемого промотора, индукция этого промотора должна быть непрерывной. Это означает, что искусственный фактор восстановления функции должен применяться во время всего жизненного цикла трансгенного растения. Сразу же после прекращения обработки растение погибнет вследствие действия блокирующей конструкции (ВС). Экспрессия блокирующего фактора может быть конститутивной или пространственно-временной, т.е. органоспецифической или специфической в отношении стадии развития. Конститутивная экспрессия восстанавливающего функцию фактора может использоваться в системе сегрегирующего возмещаемого блокирования функции (RBF). Экспрессия восстанавливающего фактора может быть также индуцируемой определенным стимулом в зависимости от механизма конкретной используемой системы возмещаемого блокирования функции (RBF). Блокирующая конструкция (ВС) может экспрессироваться в организме-хозяине конститутивно или органоспецифическим, специфическим для стадии развития, или пространственно-временным образом, и восстановление функции производится наружной компенсацией, вызываемой блокированием недостаточности или наружной индукцией чувствительного восстанавливающего функцию гена/белка или конститутивной, или временной экспрессией восстанавливающего функцию гена. Конститутивная, а также специфическая в отношении стадии развития или органоспецифическая экспрессия как блокирующей конструкции (ВС), так и восстанавливающей конструкции (RC), локализованных в разных неаллельных хромосомах, приводит к сегрегирующему возмещаемому блокированию функции (S-RBF), предусматривающему наиболее удобный контроль экспрессии трансгена (примеры 5-7). Наиболее очевидным способом конструирования этого контроля является множественная, т.е. двойная или тройная, трансформация. Сначала конститутивно экспрессирующуюся восстанавливающую конструкцию (RC) трансформируют в размножающийся половым способом многоклеточный организм (SRMO). Затем конструкцию, содержащую представляющий интерес трансген (TGI), связанный с геном, блокирующим определенную функцию данного растения, трансформируют в другую неаллельную хромосому. Трансформанты или их гибриды с нетрансгенными растениями проверяют для подтверждения, что они имеют единственную копию данного гена и что эти конструкции находятся на разных хромосомах. Гомозиготы для обеих конструкций выращивают в поле. Если трансгенный размножающийся половым способом многоклеточный организм (SRMO) или растение, гибридизуется с любым другим размножающимся половым способом многоклеточным организмом (SRMO) или растением, первое гибридное потомство будет выживать, поскольку все эти растения будут гетерозиготными в отношении обоих генов возмещаемого блокирования функции (RBF). Начиная со второго гибридного поколения, каждое второе гибридное растение, несущее блокирующий ген, будет погибать, так как восстанавливающий и блокирующий гены возмещаемого блокирования функции (RBF) будут расщепляться в различных клетках во время мейоза. Таким образом, восстановление сегрегирующего возмещаемого блокирования функции (S-RBF) включает в себя подтверждение межлинейного скрещивания размножающегося половым способом многоклеточного организма (SRMO) (WO 00/37660). Блокирующая конструкция (ВС) начинает работать только во втором гибридном потомстве и приводит к 50% негативному отбору трансгенной конструкции. Промоторами, применимыми для конститутивной экспрессии являются 35S из CaMV (Condit, et al., 1983, J. Mol. Appi. Gen. (USA), 2: 301-314; Zaitlin, et al., 1985, Biotechnology in plant science: relevance to agriculture in the eighties. Orlando, Fla. (USA). Academic press, 227-235; Williamson, et al., 1989, Plant Physiol., 90: 1570-1576), NOS (Depicker, et al., 1982, J. Mol. Appi. Gen. (USA), 1: 561-573, OCS (De Greve, et al., 1985, J. Mol. Appi. Gen. (USA), 1: 499-511) и другие. Промоторы для органоспецифической и специфической для стадии развития экспрессии могут быть получены из очень широкой группы. Для сегрегирующего возмещаемого блокирования функции (S-RBF) не имеет значения, является ли промотор “растекающимся” (“ликовым”) или нет. Важно, чтобы как блокирующая конструкция, так и восстанавливающая конструкция (ВС и RC) экспрессировались под контролем одного и того же промотора, что приводит к одновременной экспрессии этих конструкций в одних и тех же тканях растения-хозяина. Органоспецифическая, специфическая для стадии развития или специфическая в отношении индукции экспрессия генов возмещаемого блокирования функции (RBF) приводит к временному контролю критических стадий в жизненном цикле трансгенных растений и требуется наружный стимул для возмещения блокирования конкретной функции (RBF). Стадиями развития для временной экспрессии могут быть прорастание семян, цветение, созревание зародышей или плодов, образование соцветий, удлинение стебля или корня и т.д. Экспрессия блокирующей конструкции (ВС) только на конкретной стадии развития трансгенного растения делает процедуру восстановления функции более короткой и более легкой, чем в случае конститутивной экспрессии (пример 3). В то время как экспрессия блокирующего гена должна регулироваться органоспецифическим или специфическим для стадии развития промотором, экспрессия восстанавливающего функцию гена должна регулироваться индуцируемым промотором, в частности, если восстановление не основано на наружной компенсации отсутствия конкретного метаболита. Таким образом, сельскохозяйственное растение, несущее временно экспрессируемое возмещаемое блокирование функции (RBF), требует искусственного восстанавливания функции только в конкретной стадии развития или в конкретном органе. Без восстановления функции растение будет погибать или будет иметь измененный фенотип во время этой конкретной стадии развития в природных условиях. Важным свойством органоспецифического или специфического для определенной стадии развития промотора является его высокая специфичность, за исключением сегрегирующего возмещаемого блокирования функции (S-RBF). Другими словами, промотор должен корректно экспрессироваться, а не быть “ликовым” (растекающимся) промотором, так как блокирующая конструкция может убивать растение или нарушать развитие растения при экспрессии на нежелательных стадиях. Конститутивная репрессия блокирующей конструкции (ВС) в природных условиях является возможным путем организации возмещаемого блокирования функции (RBF). Если блокирующая конструкция (ВС) экспрессируется на конкретной стадии размножающегося половым способом многоклеточного организма (SRMO) или развития растения под контролем специфического промотора, восстанавливающая конструкция (RC) находится под контролем репрессируемого промотора, например модифицированного 35Sp CaMV или NOSp с tet-репрессором Tn10 (Gatz and Quail, 1988, Proc. Natl. Acad. Sci. USA, 85: 1394-1397; Gatz, et al., 1991, Mol. Gen. Genet. 227: 229-237). Этот промотор является репрессированным на протяжении жизненного цикла с белком, экспрессируемым из третьего гена, расположенного под контролем конститутивного промотора (пример 4). Промотор восстанавливающего элемента может быть запущен в действие, только когда внешний стимул (химический или физический) удаляет этот репрессорный элемент. В случае tet-репрессора связывающий белок изменяет его собственную конформацию и активирует экспрессию этой репрессированной конструкции в присутствии тетрациклина. В этом случае восстановление функции является обработкой тетрациклином трансгенного растения во время экспрессии блокирующей конструкции (ВС). Другим возможным сценарием является экспрессия всех трех генов, находящихся под контролем органоспецифического промотора. В этом случае tet-оперон Tn10 должен быть введен в промотор восстанавливающего гена. Существуют также другие кандидаты на роль репрессируемых промоторов (Lanzer, et al., 1988, Proc. Natl. Acad. Sci. USA, 85: 8973-8977). Для развития или конструирования системы возмещаемого блокирования функции (RBF) с временной экспрессией органоспецифические или специфические в отношении стадии развития промоторы должны быть исследованы на возможную экспрессию блокирующей конструкции (ВС). Несколько промоторов обнаруживают высокую специфичность для прорастания семян в экспрессии. К сожалению, каждый промотор имеет также дополнительную ‘star’-экспрессию (основную экспрессию) на других стадиях развития. Цистеин-эндопептидазы SH-EP или ЕР-С1 специфически деградируют запасные белки. Они являются белками, экспрессируемыми на высоком уровне в прорастающих семенах. Дополнительная экспрессия может наблюдаться в стареющих тканях: семядолях, бобах и даже в стеблях. Эти гены имеют большие промоторные районы около 1200-1700 п.н. Цистеин-эндопептидаза ЕР-С1 была клонирована из Phaseolus vulgaris (Ogushi et al., 1992, Plant Mol. Biol. 19: 705-706) и экспрессирована в ряде делеционных анализов в проростках трансгенного табака (Yamauchi et al., 1996, Plant Mol. Biol. 30: 321-329). Экспрессия в созревших бобах делает применение этого промотора проблематичным для экспрессии этой блокирующей конструкции (ВС) в прорастающих семенах. Промотор SH-EP сульфгидрил-эндопептидазы (1676 п.н.) был клонирован из Vigna mungo (Akasofu et al., 1990, Nucl. Acid Res. 18: 1892) и исследован в качестве SH-EP-цистеин-эндопептидазы (Yamauchi et al., 1996, Plant Mol. Biol. 30: 321-329). Он экспрессируется исключительно в прорастающих проростках. Только низкая дополнительная экспрессия была обнаружена в созревших бобах. Экспрессия начинается на второй день после начала прорастания. Максимум экспрессии в прорастающих семенах имеет место на третий день на уровне мРНК и на четвертый день на уровне белка. Ферментативная активность проявляется до пятого-шестого дня и затем падает. Клонированная генная конструкция с ее собственным промотором продуцирует в трансгенном табаке даже больше белка, чем в Vigna mungo. Было также обнаружено действие этого промотора в зародыше табака. Таким образом, согласно данному изобретению этот промотор может быть использован, в частности, в системе двойного возмещаемого блокирования функции (D-RBF) во время развития семян и прорастания семян. Промоторы LEA экспрессируются на поздних стадиях развития зародыша (Huges and Galau, 1989; Galau, et al., 1992, Plant Physiol. 99: 783-788; Devis, et al., 1996, Plant J. 9: 2005-2015). Промоторы LEA не являются «ликовыми» (растекающимися) и являются высоко специфическими для позднего эмбриогенеза и, следовательно, являются применимыми промоторами для временной экспрессии RBF. Промотор гена, кодирующего фермент O-метилтрансферазу кофеиновой кислоты (СОМТ), экспрессирует белок чрезвычайно специфическим образом в ткани цветоножек райграсса пастбищного (английского) Lolium perenne (McAlister, et al., 1998, Australian J. Plant Physiol. 25: 225-235). Экспрессия блокирующей конструкции (ВС) под контролем этого типа промотора может предотвращать образование цветоножек и, следовательно, размножение растений райграсса в природных условиях. Фрагмент 620 п.н. промотора из гена МТ1-А (металлотионеин-подобного) является достаточным для управления экспрессией в корнях трансформированного хлопчатника (Hudspeth, et al., 1996, Plant Mol. Biol. 31: 701-705). В соответствии с данным изобретением он может быть использован для блокирования развития корней. Примерами применимых промоторов являются промоторы малатсинтазы (MS) и изоцитратлиазы (ICL), которые являются активными во время прорастания и в проростках подавляются сахарозой и активируются GA3. Известно, что дополнительная экспрессия происходит в стареющих органах и в прорастающей пыльце. Пространственная и временная экспрессия под контролем промоторов огурцов (Reynolds and Smith, 1995, Plant Mol. Biol. 27: 487-497), масличного рапса (Zhang et al., 1994, Plant Physiol. 104: 875-864) и томатов (Janssen, 1995, Plant Physiol. 108: 1339) интенсивно исследовалась в трансгеннных табаке и масличном рапсе. Малатсинтаза (MS) имеет более широкий спектр экспрессии, но является менее идеальной для специфической для прорастания экспрессии (Sarah et al., 1996, Mol. Gen. Genet. 250: 153-161). Промоторы изоцитратлиазы (ICL) и малатсинтазы (MS) являются активными в пыльце и применимы в системах двойного возмещаемого блокирования функции (D-RBF). Промоторы MS и ICL В. napus имеют среднюю 1% активность в пыльце в сравнении с экспрессией в проростках (Zhang et al., 1994, Plant Physiol. 104: 875-864). Эти промоторы экспрессируются слабо в стареющих органах, таких как семядоли (Graham et al., 1992, Plant Cell 4: 349-357). Они могут активироваться в темноте и подавляться сахарозой и другими сахарами (Graham et al., 1992, Plant Cell 6: 761-772). Экспрессия изоцитратлиазы (ICL) в зрелых семенах может достигать значительных уровней (5-10%) в проростках (Turley and Trelease, 1990, Plant Mol. Biol. 14: 137-146). Промоторы MS и ICL интенсивно исследовались в качестве цис-действующих элементов (Sarah et al., 1996, Mol. Gen. Genet. 250: 153-161). Сообщался фрагмент 17 п.н., чувствительный к гиббереллину, из промотора катепсин В-подобного белка пшеницы (Cejudo et al., 1992, Plant Mol. Biol. 20: 849-856). AMY (высокий P1)-альфа-амилаза является активной в эндосперме во время прорастания. Все известные промоторы амилаз являются также активными в различных органах растений Phaseolus vulgaris и Vigna mungo (Minamikava et al., 1992, Plant Cell Physiol. 33: 253-258). Имеется известная индуцируемая GA3 P1-амилаза из ячменя (Rahmatullah et al., 1989, Plant Vol. Biol. 12: 119-121). Бета-1,3-глюканаза является активной в эндосперме во время прорастания. Она является противогрибковым белком, индуцируемым GA3. Она является активной также и в листьях и других органах и индуцируется поранением (Vogeli-Lange et al., 1994, Plant J. 5: 273-278). Имеется ряд других кандидатов на промоторы для временной экспрессии и это число увеличивается с каждым годом. Экспрессия гена, ответственного за восстановление действия блокирующей конструкции (ВС), требует промоторов, отвечающих на внешний стимул. Внешний стимул может быть химическим или физическим. Химическим стимулом может быть любая молекула, способная регулировать активность конкретного промотора. Физическим стимулом может быть температура, осмос, свет, гравитация или что-то еще. Следующие два типа промоторов служат в качестве примеров ответов на внешний стимул: индуцируемые салицилатом и температурным шоком промоторы. Промотор гена, связанного с патогенезом белка PA1a, исследовали в экспериментах с 5′-делецией в трансгенном табаке (Ohshima, et al., 1990, Plant Cell 2: 95-106). В другом делеционном эксперименте были обнаружены несколько регуляторных элементов в промоторе 902 п.н. PR-1a (Van de Rhee, et al., 1990, Plant Mol. Biol. 21; 451-461; Payne, et al., 1988, Plant Mol. Biol. 11: 89-94; Pfitzner, et al., 1988, Mol. Gen. Genet. 211: 290-295). Было показано, что активность этих промоторов повышалась после 24-48 часов индукции салицилатом и была близкой к индукции, вызываемой инфекцией TMV. Промоторы белков теплового шока исследовали тщательно в различных растениях. Их активность часто была в несколько раз более высокой, чем активность промотора 35S. Индукция HS-промоторов обычно происходит при повышении температуры окружающей среды до 35-45°С (Ainley and Key, 1990, Plant Mol. Biol. 14: 949-967). Отрицательным признаком промоторов белков температурного шока является их экспрессия в проростках и их активация также физическим стимулом, иным, чем нагрев. Другим применимым промотором является промотор гена глутатион-S-трансферазы кукурузы (GSTII-27), который может активироваться гербицидом Safeners R-25788 (WO 94/03619). В патенте США US 5880333 сообщалась химически регулируемая экспрессия промотора. Tet-репрессорная система Tn10 (Gatz and Quail, 1988, Proc. Natl. Acad. Sci. USA, 85: 1394-1397) активируется химическим агентом – тетрациклином, но ее механизм является противоположным стандартной индукции. Эта система требует двух генов: во-первых, восстанавливающего гена под контролем промотора, содержащего tet-оператор, во-вторых, другого гена, расположенного под контролем конститутивного или органоспецифического и/или отвечающего на стадию развития промотора гена, кодирующего репрессорный белок, связывающийся с сигнальной последовательностью ДНК tet-оператора. Эта система является репрессированной, пока наружно применяемый тетрациклин не активирует восстанавливающий ген. Данное изобретение относится к векторам, плазмидам, содержащим один или несколько трансгенных инсертов (TI), которые могут быть собраны с образованием комплекса ДНК-конструкций или систем возмещаемого блокирования функции (RBF) данного изобретения, и к применению указанных систем возмещаемого блокирования функции (RBF) для обеспечения увеличенного уровня надежности в контроле сегрегации и утечки генов при переносе в размножающийся половым способом многоклеточный организм (SRMO), в том числе в растениях и в животных (кроме человека). Эти векторы, плазмиды и ДНК-кассеты могут переноситься в клетки размножающегося половым способом многоклеточного организма с образованием трансформированных трансгенных клеток растений или животных и в конечном счете растений или животных. Уровень контроля расщепления и/или утечки в окружающую среду увеличивается блокирующей конструкцией, которая может быть извлечена и может не только использоваться, но также повторно использоваться фермером или другим конечным пользователем при обеспечении его соответствующей инструкцией. В практическом применении трансгенные растения, несущие систему возмещаемого блокирования функции (RBF), могут выращиваться в открытом поле и не отличаться от нетрансгенных растений. Сельскохозяйственные растения, несущие систему возмещаемого блокирования функции (RBF), требуют внешней обработки для восстановления блокированной функции обычно в конкретной стадии развития. Этот способ описан подробно только для применения в растениях, но он также вполне применим для животных. Описанный выше анализ позволяет сравнивать кандидатные конструкции и механизмы на диапазон действия, надежность и пригодность различных систем возмещаемого блокирования функции (RBF) в контроле сегрегации и утечки генов. Превосходство систем возмещаемого блокирования функции (RBF) данного изобретения основано на их способности элиминировать действие блокирующей конструкции (ВС) с одновременным действием представляющего интерес трансгена (TGI). Иногда это блокирование может не действовать после кроссинговера или мутаций блокирующей конструкции (ВС) или в результате некоторых гасящих механизмов. Кроссинговер может разделять блокирующую конструкцию (ВС) и контролируемый представляющий интерес трансген (TGI) в том случае, если генотип растений-хозяев является гетерозиготным в отношении этих генов. Чем ближе вместе находятся блокирующая конструкция (ВС) и контролируемые представляющие интерес трансгены (TGI), тем меньше возможность разделения этих конструкций. Помещение блокирующей конструкции (ВС) в интрон представляющего интерес трансгена (TGI) существенно уменьшает возможность отделения или мутации блокирующей конструкции (ВС) без повреждения представляющего интерес трансгена (TGI) вследствие совпадения сигнальной и кодирующей последовательностей обоих этих генов. Конститутивная экспрессия блокирующей конструкции (ВС) может приводить к непрерывному давлению для разрыва этого блокирования. Таким образом, предпочтительно, чтобы блокирующая конструкция (ВС) экспрессировалась под контролем органоспецифических или специфических для определенной стадии развития промоторов. Система сегрегирующего возмещаемого блокирования функции (S-RBF) может увеличить возможность разделения или раздельной репрессии блокирующей конструкции (ВС) и представляющего интерес трансгена (TGI) вследствие нескольких гибридизаций с другими растениями, тогда как две различные блокирующие конструкции (ВС) на обеих сторонах представляющего интерес трансгена (TGI), т.е. система двойного возмещаемого блокирования функции (RBF), сильно уменьшает возможность неудачи этого блокирования. Применение этих предупредительных средств элиминирует возможность функциональной неудачи в блокирующих конструкциях (ВС). Система множественного или тройного возмещаемого блокирования функции (M-RBF или T-RBF) объединяет все самые лучшие признаки сегрегирующего, двойного и обращенного возмещаемого блокирования функции (RBF). Она сообщает более сильный негативный отбор, чем система сегрегирующего (одиночного или обращенного) возмещаемого блокирования функции (SR-RBF). Тройное возмещаемое блокирование функции (T-RBF) не требует специальных обработок, кроме внутрилинейной гибридизации или скрещивания, подобных системе сегрегирующего возмещаемого блокирования функции (S-RBF). Она предохраняет любую трансгенную конструкцию от перетекания в окружающую среду. Она имеет двойную защиту и, следовательно, является высоконадежной, как и система двойного возмещаемого блокирования функции (D-RBF). Она может быть легко сконструирована, так как различные нуклеотидные последовательности, необходимые для трансгенных инсертов (TI), используемые в сборке системы возмещаемого блокирования функции (RBF), помещают в три различных трансгенных инсерта (TI), которые помещают в различные неаллельные хромосомы. Это позволяет использовать готовые к применению реципиентные линии размножающегося половым способом многоклеточного организма (SRMO) для клонирования различных представляющих интерес трансгенов (TGI). В заключение можно сделать вывод, что из рассмотренных моделей наиболее удобной, надежной и безопасной системой возмещаемого блокирования функции (RBF), доступной в настоящее время, является система тройного возмещаемого блокирования функции (T-RBF). В качестве блокирующих генов могут быть использованы гены, имеющие различные функции и экспрессируемые под контролем различных промоторов. Они должны быть помещены на обеих сторонах представляющего интерес трансгена (TGI) или представляющих интерес трансгенов (TGI). Конструирование и применение различных систем возмещаемого блокирования функции (RBF) описаны в следующих ниже примерах, которые включены только для иллюстративных целей и не должны быть использованы для ограничения заявленного формулой изобретения объема защиты. ПРИМЕР 1 Непрерывная компенсация метаболита, элиминированного конститутивной экспрессией блокирующей конструкции (ВС) фермента L-аспарагиназы/L-глутаминазы. Возмещение с использованием вводимого извне L-аспарагина/L-глутамина. Схематические положения этих генов показаны на фиг.1а. Эта система возмещаемого блокирования функции (RBF) является примером, как работает эта система на уровне метаболита – в этом случае на уровне аминокислоты или азотного метаболизма. Она является наиболее контролируемой системой возмещаемого блокирования функции (RBF). Блокирование азотного метаболизма приводит к летальным эффектам, приводя к отсутствию аминокислот, в частности, во время прорастания семян. L-аспарагиназа является активной в тканях “стока”, богатых аспарагином. Экспрессия этого фермента в физиологически активных тканях может ингибировать нормальный метаболизм азота. Блокирующую конструкцию (ВС) помещают вблизи представляющего интерес гена или между представляющими интерес генами. L-аспарагиназа из Lupinus angustifolius (Dickson, et al., 1992, Plant Mol. Biol. 20: 333-336), Erwinia Chrysanthemi (Filpula, et al., 1988, Nucl. Acid Res. 16: 10385), Arabidopsis thaliana (Casado, et al., 1995, Plant Physiol. 108: 1321-1322) или другого источника может экспрессироваться под контролем NOS, ОСТ, 35S или других конститутивных промоторов. Конститутивная экспрессия этого фермента делает растение-хозяина непрерывно зависимым от наружного предоставления аспарагина. Такой жесткий контроль этого трансгена может быть необходим, когда представляющий интерес ген создает реальную угрозу для людей или окружающей среды. Молекулярные конструкции представлены на фиг.1а. ПРИМЕР 2 Антисмысловая мРНК энт-кауренсинтазы А, временно экспрессируемая в прорастающих семенах репы огородной (турнепса) Возмещение с использованием вводимых извне гиббереллинов. Схематические положения этих генов показаны на фиг.1b. Эта система возмещаемого блокирования функции (RBF) работает на уровне “гашения” РНК с возмещением блокирования, работающим на уровне гормона. Мутация в малокопийном гене, кодирующем энт-кауренсинтазу А, приводит к карликовому фенотипу Arabidopsis thaliana. Экспрессия антисмысловой РНК этого фермента приводит к тому же самому изменению фенотипа вследствие блокирования синтеза гиббереллина в растении репе огородной (турнепсе). Это является временным блокированием функции, так как фермент находится под контролем промотора SH-EP (цистеин-эндопептидазы ЕР) из Vigna mungo. Экспрессия антисмысловой энт-кауренсинтазы А начинается во время позднего эмбриогенеза и усиливается на стадии прорастания семян. Растение-хозяин, несущее систему возмещаемого блокирования функции (RBF), развивает карликовый фенотип и является неспособным к размножению в природных условиях. Возмещение этого блокирования функции предусматривает наружное применение гиббереллинов. Было показано, что с использованием наружного нанесения гиббереллинов из карликового фенотипа Arabidopsis могут быть вновь получены нормальные растения (Xu, et al., 1997, Plant Physiol. (Rockville) 114: 1471-1476; Phillips and Huttly, 1994, Plant Mol. Biol. 24: 603-615). На практике семена трансгенных сельскохозяйственных растений должны быть покрыты содержащей гиббереллин смесью для получения растений с нормальным фенотипом. Молекулярная конструкция показана на фиг.1b. ПРИМЕР 3 Возмещаемое блокирование функции (RBF) с барназой, контролируемой промотором эндопептидазы SH-EP. Восстановление функции с использованием гена барназы, находящегося под контролем промотора теплового шока HS. Схематические положения этих генов показаны на фиг.2. Эта система возмещаемого блокирования функции (RBF) работает на уровне взаимодействия белков. Она состоит из генов барназы и barstar, происходящих из Bacillus amyloliquefaciens (Hartly, 1989, Trends Biochem. Sci. 14: 450-454). Барназа (бактериальная рибонуклеаза) является наиболее известным летальным геном, с тех пор как Mariani, et al. (1990, Nature 347: 737-741) использовал ее для конструирования контроля фертильности в трансгенных растениях. При экспрессии в растении барназа блокирует экспрессию всех генов посредством разрушения молекул РНК. Для действия в качестве блокирующей конструкции (ВС) ген барназы экспрессировали под контролем промотора цистеин-эндопептидазы (SH-EPp), клонированного из Vigna mungo и прежде называемого промотором сульфгидрил-эндопептидазы (Akasofu et al., 1990, Nucl. Acid Res. 18: 1892; Yamauchi et al., 1996, Plant Mol. Biol. 30: 321-329). Восстанавливающая конструкция (RC) содержит ген barstar, помещенный под контроль промотора теплового шока (HSp) сои (Ainley and Key, 1990, Plant Mol. Biol. 14: 949-967). Синтетическую барназу (ген) помещали под контроль промотора SH-EP и давали ей экспрессироваться во время эмбриогенеза и прорастания семян в трансформированных растениях табака. Схематическая структура этой конструкции, несущей четыре гена, показана на фиг.2. Эта ДНК-конструкция содержит также uidA (GUS) в качестве представляющего интерес гена и ген hpt в качестве селектируемого маркера. Ген uidA клонировали под контролем промотора 35S CaMV, а ген hpt под контролем промотора NOS Agrobacterium. Все четыре гена расположены в одной Т-ДНК вектора на основе pBIN19 (pGPTV-HPT) (Bevan М., 1984, Nucl. Acid Res. 12: 871-8717; Frish et al., 1995, Plant Mol. Biol. 27: 405-409). Для исследования экспрессии HSp и SH-EPp растения табака трансформировали геном uidA, находящимся под контролем этих промоторов. Исследования на этих трансгенных растениях табака показали, что промотор HS экспрессировал ликовый фермент GUS иногда на низком уровне в генеративных органах. Он демонстрировал слабый пик экспрессии на третий день прорастания и постепенно исчезал во время нескольких следующих дней без стимуляции извне. Он активировался в ответ на высокую температуру в диапазоне 37-45°С, применяемую в течение 0,5-3 часов. Для промотора HS использовали активирующую обработку 40°С в течение одного часа. Экспрессия GUS под контролем промотора HS возрастала во время первых двух дней и продолжалась в течение по меньшей мере 3-4 дней после теплового шока. Промотор SH-EP экспрессировал фермент GUS в зародышах и в проростках. Пик активности GUS под контролем SH-EPp достигался в средней или поздней стадиях развития зародыша и он снова появлялся на третий-пятый день прорастания. В других органах растений табака не было экспрессии промотора SH-EP. Несколько трансгенных растений табака были получены после трансформации ДНК-конструкцией, содержащей представляющий интерес ген uidA, ген селектируемого маркера hpt и систему возмещаемого блокирования функции (RBF) с генами барназы и barstar. Трансгенные растения табака, несущие конструкцию возмещаемого блокирования функции (RBF), демонстрировали нормальный фенотип. Они вполне нормально росли, цвели и производили коробочки (плоды) после самоопыления. Семена из этих растений имели нормальный размер. Однако они не прорастали. Применение теплового шока во время проращивания не восстанавливало блокированную функцию прорастания этих трансгенных семян. Как предсказывалось, развитие зародыша было остановлено экспрессией барназы во время созревания коробочки. Таким образом, зародыши этих семян были фактически мертвыми. Все соцветия срезали из растений табака, после чего растениям давали снова образовать соцветия. Растения с новыми цветками и зелеными коробочками подвергали действию теплового шока 40°С в течение 1 часа в каждый второй день. Из этих растений табака собирали созревшие коробочки. Семена проращивали и подвергали действию теплового шока по меньшей мере один раз в течение одного часа при 40°С во второй и третий день прорастания. Обработанные нагреванием семена образовали нормальные проростки, которые вырастали до нормальных растений табака. Эти семена начинали прорастать также без теплового шока. Однако большинство из них не могли распускать их семядоли. Они проявляли этиолированный фенотип и погибали. Однако часть этих проростков распускала семядоли и преодолевала блокирование развития. Наиболее очевидно, успешно прораставшие семена накапливали существенное количество белка barstar для ингибирования барназы, экспрессируемой во время прорастания семян. Только повторяемая несколько раз обработка тепловым шоком несущих коробочки растений снимала блокирование функции прорастания семян. Одноразовая обработка тепловым шоком в течение 2 часов при 40°С, применяемая на различных стадиях эмбриогенеза от зацветания до образования коричневой коробочки, не удаляла блокированное прорастание. Непрерывная обработка тепловым шоком, примененная к растениям табака с коробочками и цветками, также не привела к успеху. Коробочки страдали от засухи и семена не развивались должным образом. Хотя контрольные нетрансгенные семена прорастали после непрерывного теплового шока, несущие конструкцию возмещаемого блокирования функции (RBF) семена не прорастали. Возможно непрерывный тепловой шок высушивал коробочки и промотор HS не продуцировал достаточного количества барназы в период времени до высыхания коробочек. Хотя часть проросших трансгенных проростков F1 не была положительной в GUS-анализе в отношении расщепления трансгена, все семена той же самой трансгенной линии, не обработанные тепловым шоком, не прорастали. Это можно объяснить тем фактом, что экспрессия GUS под контролем промотора SH-EP регистрировалась на низком уровне также и в других частях коробочки. Это означает, что барназа могла блокировать синтез мРНК также и в зародышах, которые не несли конструкции возмещаемого блокирования функции (RBF). Действие блокирующих и восстанавливающих генов подтверждали Нозерн-анализом экспрессии (фиг.3). Экспрессия мРНК барназы под контролем промотора SH-EP имела пик в средней стадии эмбриогенеза (беловатые зародыши) на уровне 0,1 пг/1 мкг тотальной РНК и уменьшалась до 0,03 пг/мкг тотальной РНК в позднем эмбриогенезе (желтоватые зародыши). Второй пик экспрессии начинался на третий день прорастания семян, достигал 0,04 пг/1 мкг тотальной РНК на четвертый день и исчезал на пятый день прорастания. Промотор HS демонстрировал растекающуюся экспрессию мРНК barstar на двух стадиях развития, т.е. прорастания зародыша и семени, на уровне 0,05-0,2 пг/1 мкг тотальной РНК. Обработка высокой температурой активировала экспрессию barstar под контролем промотора HS до 1-3 пг/1 мкг тотальной РНК. В частности, активность промотора HS возрастала после нескольких последовательных одноразовых обработок высокой температурой в течение одного-двух дней. Хотя Нозерн-анализ показал, что промотор HS имел высокую растекающуюся активность, экспрессия barstar не была достаточной для того, чтобы ингибировать действие барназы без обработки высокой температурой, особенно на стадии эмбриогенеза. В целом, если учитывать, что экспрессия белка часто следует после экспрессии мРНК с задержкой от нескольких часов до одного дня, результаты Нозерн-анализа и GUS-анализа экспрессии промоторов SH-EP и HS были сравнимыми, и конструкция возмещаемого блокирования функции (RBF) работала как предсказывалось. На практике система возмещаемого блокирования функции (RBF) предусматривает применение одночасовой обработки тепловым шоком при 40°С один раз в два последовательных дня во время развития коробочек и по меньшей мере один раз во время прорастания. Любые другие использованные температурные обработки не смогли снять блокированное прорастание семян. Система возмещаемого блокирования функции (RBF) надежно контролирует расщепление и ускользание (утечку) представляющего интерес трансгена (TGI) в случае uidA. Если эти трансгеные растения случайно нагревались бы таким же образом в природе, это, вероятно, не могло бы повторяться в нескольких последующих сезонах выращивания. Трансгенные растения, продуцирующие семена, могут выращиваться в условиях оранжерей, где легко выполнять обработку тепловым шоком. В открытом поле трансгенное растение, несущее систему возмещаемого блокирования функции (RBF), продуцирует семена, которые не прорастают. Пыльца, несущая конструкцию возмещаемого блокирования функции (RBF), может опылять в открытом поле родственные растения. Однако семена, продуцируемые после этого опыления, не прорастают, таким образом, система возмещаемого блокирования функции (RBF) предотвращает распространение трансгенов в природной окружающей среде. ПРИМЕР 4 Сегрегирующее возмещаемое блокирование функции (S-RBF) с геном cry9Aa в качестве представляющего интерес трансгена (TGI) и геном барназы в качестве блокирующей конструкции (ВС) в одном инсерте и геном barstar в качестве восстанавливающей конструкции в другом трансгенном инсерте (TI). Схематические положения этих генов показаны на фиг.4. Эта система возмещаемого блокирования функции (RBF) включает в себя в качестве блокирующей конструкции (ВС) ген барназы под контролем промотора эндопептидазы SH-EP (SH-ЕРр), клонированного из Vigna mungo. Ген cry9Aa использовали в этой конструкции в качестве представляющего интерес трансгена (TGI). Вектор трансформации растений pVK112 конструировали на основе вектора pGPTV-HPT (Becker, D. et al., 1992. Новые бинарные векторы с селектируемыми маркерами, локализованными проксимально относительно левой границы Т-ДНК. Plant Mol. Biol. 20, 1195-1197). Восстанавливающую конструкцию (RC), ген barstar под контролем промотора 35Sp или HSp клонировали в pGPTV-KAN (Becker, D. et al., 1992, Plant Mol. Biol. 20, 1195-1197) и трансформировали в растения табака. Растения, экспрессирующие мРНК barstar ожидаемым образом, использовали для второй трансформации вектором pGPTV-HPT, несущим блокирующую конструкцию (ВС) и представляющий интерес инсектицидный ген cry9Aa (Kuvshinov V., et al., 2001, Plant Sci. 160: 341-353). Растения, несущие два инсерта, включающих HSp-barstar-восстанавливающую конструкцию (RC), проявляли те же самые признаки, что и растения с одним инсертом возмещаемого блокирования функции (RBF), описанным в примере 3. Трансгенные растения табака с восстанавливающей конструкцией (RC), регулируемой 35Sp, проявляют нормальный фенотип. Летальный генотип в семенах, содержащих инсерт только с блокирующей конструкцией (ВС) и геном cry9Aa, имеет место во время гибридизации с табаком дикого типа после сегрегации (расщепления) в соответствии с фиг.8. Живые семена содержат обе конструкции (ВС и RC) или только одну восстанавливающую конструкцию (RC) или не содержат трансгенных инсертов. Негативный отбор против трансгенной инсерции, содержащей блокирующую конструкцию (ВС) и ген cry9Aa, составлял 50% в каждом потомстве. ПРИМЕР 5 Сегрегирующее возмещаемое блокирование функции (S-RBF), когда блокирующий ген, barnase, локализован под контролем органоспецифического промотора в интроне представляющего интерес трансгена (TGI) и регулируется органоспецифическим промотором (SH-EP). Восстанавливающей конструкцией (RC) является ген barstar, расположенный в отличающейся неаллельной хромосоме под контролем конститутивного (35Sp) или индуцируемого тепловым шоком (HSp) промотора. Схематические положения этих генов показаны на фиг.5а и 5b. Возмещаемое блокирование функции (RBF) действует на уровне белок-белкового взаимодействия. Блокирующий ген, ген барназы, расположен в интроне представляющего интерес гена uidA (GUS) для предотвращения разделения блокирующей конструкции (ВС) и представляющего интерес трансгена (TGI) во время кроссинговера. Ген барназы регулируется промотором SH-EP из Vigna mungo. Восстанавливающий ген, barstar, расположен в отличающейся неаллельной хромосоме под контролем того же самого промотора. Растения табака, несущие восстанавливающую конструкцию (RC) с геном barstar, находящимся под контролем HSp или 35Sp (описанными в примере 4), повторно трансформировали трансгенной конструкцией, содержащей ген GUS с введенной в интрон кодирующей последовательностью барназы (фиг.5). Это трансгенное растение не отличалось от нетрансгенного растения вследствие непрерывного отсутствия экспрессии барназы. Восстановление трансгенных растений обеспечивается, когда гомозиготные (BBRR) трансгенные растения скрещиваются друг с другом и, таким образом, в потомстве поддерживается гомозиготный генотип. Когда такое трансгенное растение скрещивается с посторонним растением, гибридное растение первого поколения также проявляет нормальный фенотип, но несет гетерогенный (BbRr) генотип. Сегрегирующее возмещаемое блокирование функции (S-RBF) становится эффективным со второго гибридного поколения, когда блокирующая и восстанавливающая конструкции (ВС и RC) будут расщепляться в различные клетки-хозяева. Каждый второй гибридный зародыш, содержащий блокирующую конструкцию (ВС), будет нести генотип Bbrr, приводящий к гибели этого растения во время эмбриогенеза или прорастания, на стадии, зависящей от вида растения. Посредством этого блокирующая конструкция (ВС) выполняет эффективный негативный отбор во время гибридизации с посторонними растениями. Этот негативный отбор будет устранять трансген из популяции во время нескольких поколений. Число растений, несущих этот фактор негативного отбора (X), будет уменьшаться с числом поколений (n) в следующей пропорции: Х=Y·2-n, где Y является начальным числом растений, несущих негативный отбор в большой популяции. Например, если начальное число гибридных растений было Y=1000, после n=10 поколений число растений X, несущих конструкцию возмещаемого блокирования функции (RBF), падает ниже 1, Х=1000·2-10 или Х<1. Поскольку блокирующая конструкция (ВС) находится внутри представляющего интерес трансгена (TGI), кроссинговер не может разделить представляющий интерес трансген (TGI) и блокирующую конструкцию (ВС) в аллельные хромосомы. Конструкция с геном barstar, регулируемая промотором теплового шока, объединяет признаки систем сегрегирующего и индуцируемого возмещаемого блокирования функции (RBF). Для выживания и размножения эти трансгенные растения требуют обработки тепловым шоком, как описано в примере 3. Последовательности интрона и сигнала полиаденилирования представляющего интерес трансгена (uidA) рассматривались с промотором, UTR, кодирующей последовательностью и сигналом полиаденилирования блокирующего гена (гена барназы). Сигнальные последовательности интрона, используемые в этой конструкции: доля ТА 55-70%, сайт границы экзон/интрон AG/GUAUGU (SEQ ID NO:5), последовательность точки ветвления (UA) CUAAC (SEQ ID NO:6), расположенная на 20-50 нуклеотидов против хода транскрипции от 3′-сайта границы интрон/экзон GCAG/G (SEQ ID NO:7). U-обогашенные последовательности стимулируют сплайсинг независимо от их положения в интроне. Одиночные или двойные замены U на G сильно ингибируют сплайсинг. Высокое содержание AU и особенно U необходимо в первых и последних 50 нуклеотидах этого интрона. Экзоны являются приблизительно на 15% богаче в содержании GC, чем интроны. Терминация транскрипции РНК-полимеразой II требует присутствия функционального сигнала полиаденилирования в транскрипционной единице ближнего элемента против хода транскрипции (NUE) AAUAAA (SEQ ID NO:8), однако растения имеют много модификаций этого сайта. Дальний элемент против хода транскрипции (FUE) является еще более неконсервативным. Общими признаками FUE являются U- или UG-богатые последовательности, расположенные в 10-40 нуклеотидах перед NUE. Наиболее часто встречающимся является UUUGUA (SEQ ID NO:9), но также UGUGUUUUUU (SEQ ID NO:10) или UGUUGUG (SEQ ID NO:11). Сайт расщепления также не является высококонсервативным. Обычно он состоит из Y нуклеотидов А и расположен в 10-30 нуклеотидах после сайта NUE. В промоторном районе имеются два различных блока: ТАТА-блок в консенсусной последовательности TCACTATATATAG (SEQ ID NO:12), локализованной в 15-60 нуклеотидах от начала транскрипции, и СААТ- (SEQ ID NO:13) или AGGA-блок (SEQ ID NO:14), локализованный в 30-100 нуклеотидах против хода транскрипции (слева) от ТАТА-блока. Трансляция начинается в 40-80 нуклеотидах по ходу транскрипции (справа) от стартовой точки трансляции от кодона AUG в предпочтительной последовательности ААСАА TGGCT (SEQ ID NO:12) (нуклеотиды, напечатанные жирным шрифтом, являются высококонсервативными). В качестве примера, последовательность гена барназы, расположенная в интроне гена uidA под контролем промотора SH-EP, показана на фиг.5а. Трансгенное растение табака, содержащее инсертированную в интрон блокирующую конструкцию (ВС), а также контрольные растения, содержащие ген GUS под контролем промотора салициловой кислоты, экспрессировали ген GUS и дали положительные результаты в GUS-анализе. Однако контрольные растения, содержащие укороченный ген GUS, не обнаружили никакой GUS-активности. Этот результат предполагает, что интрон гена GUS был сплайсирован правильно. ПРИМЕР 6 Двойная блокирующая конструкция (ВС) и одна восстанавливающая конструкция (RC), помещенные в один трансгенный инсерт с представляющим интерес геном (GUS) и раздельно в различные неаллельные хромосомы. Схематические положения этих генов показаны на фиг.7. Система возмещаемого блокирования функции (RBF) включает в себя в качестве первой блокирующей конструкции (ВС) ген барназы, находящийся под контролем промотора эндопептидазы SH-EP (SH-EPp), клонированного из Vigna mungo, и вторую блокирующую конструкцию (ВС), содержащую тот же самый ген барназы, находящийся под контролем промотора круциферина (CRUp) из Brassica napus. Трансгенный инсерт содержит только одну восстанавливающую конструкцию (RC), содержащую ген barstar, находящийся под контролем промотора теплового шока (HSp). Этот трансгенный инсерт нес также ген GUS в качестве представляющего интерес трансгена (TGI) и ген гигромицин-фосфат-трансферазы (hpt) в качестве селектируемого маркера. Эта конструкция была введена в вектор, используемый в примере 3. Вторая блокирующая конструкция (ВС), ген барназы под контролем CRUp, была введена в другую сторону представляющего интерес трансгена (GUS) между генами GUS и hpt с получением вектора pVK30 (фиг.7а). Предварительные исследования экспрессии CRUp с геном GUS показали, что этот промотор экспрессируется в стадии развития зародыша таким же образом, как SH-EPp. Эту конструкцию из pVK30 трансформировали в растения табака. GUS-положительные трансгенные растения табака собирали для дополнительных исследований. В этих экспериментах двойное возмещаемое блокирование функции (D-RBF) действовало, как и в одиночном возмещаемом блокировании функции (RBF), описанном в примере 3. Единственное различие заключалось в том, что возможность инактивации двойного возмещаемого блокирования функции (D-RBF) в результате мутаций или молчания промотора была оценена как в 106 раз более низкая, чем в случае одиночного возмещаемого блокирования функции (RBF). Второй подход выполняли с конечной конструкцией в pVK30. Восстанавливающий ген barstar удаляли из содержащего эту конструкцию вектора pVK30 (фиг.7b). Эту двойную блокирующую конструкцию (ВС) без гена barstar трансформировали в растения табака, несущие ген barstar (экспрессируемый под контролем промоторов 35Sp или HSp), которые были описаны в примерах 4 и 5. Трансформированные регенерированные проростки отбирали с использованием отбора с гигромицином. Экспрессию гена uidA детектировали в GUS-анализе. Обе блокирующие конструкции (ВС) экспрессировались во время созревания зародыша. Ген barstar экспрессировался конститутивно под контролем промотора 35S и индуцировался тепловым шоком под контролем промотора HSp. Конструкции возмещаемого блокирования функции (RBF) с одним инсертом и двумя инсертами действуют таким же образом, как описано в примере 3. Для успешного прорастания семян трансгенные растения табака должны быть обработаны тепловым шоком на стадии созревания семенных коробочек. В случае 353-регулируемого гена barstar блокирующая конструкция (ВС) убивает зародыши после отделения содержащего представляющий интерес трансген (TGI) инсерта от содержащего восстанавливающую конструкцию (RC) инсерта, как показано на фиг.8. ПРИМЕР 7 Тройное возмещаемое блокирование функции (RBF) Две системы возмещаемого блокирования функции (RBF): barnase/barstar и NPK15/tet-репрессор с геном GUS в качестве представляющего интерес трансгена, помещенным между блокирующими системами barnase и NPK15. Схематические положения этих генов показаны на фиг.9. Первая конструкция возмещаемого блокирования функции (RBF) состоит из блокирующего гена NPK15 или иногда онкогена, кодирующего индолацетамидсинтазу (IAMS), как в Van Onchelen et al., 1986, FEBS Lett. 198, 357-360 (B1), находящихся под контролем химерного промотора SH-EPp с инсертированной последовательностью tet-оперона, и восстанавливающего гена (R1), кодирующего репрессорный белок Tet10 и находящегося под контролем промотора 35S из CaMV. Вторая конструкция возмещаемого блокирования функции (RBF) состоит из блокирующего гена барназы (В2), находящегося под контролем промотора CRUp, и восстанавливающего гена (R2), кодирующего ген barstar и находящегося под контролем промотора 35Sp. Это трансгенное растение несет три различные трансгенные конструкции, находящиеся в различных неаллельных хромосомах. Первая конструкция содержит представляющий интерес трансген (I), помещенный между двумя различными блокирующими конструкциями (B1 и B2). Вторая конструкция содержит первую восстанавливающую конструкцию (R1), связанную со второй блокирующей конструкцией (В2). Третья конструкция содержит вторую восстанавливающую конструкцию (R2), связанную с первой блокирующей конструкцией (B1). Эти блокирующие конструкции действуют в основном на стадии эмбриогенеза и прорастания. Фиг.10 изображает реципиентную трансгенную линию растения и трансформирующий вектор растения с двойной блокирующей конструкцией для клонирования различных представляющих интерес трансгенов (TGI). Расщепление этих конструкций во время гибридизации и интербридинга показано и описано более подробно на фиг.11.
Формула изобретения
1. Способ обеспечения увеличенного уровня надежности для контроля сегрегации трансгенов в размножающемся половым способом растении и/или для обеспечения автоматического предотвращения утечки трансгенов, отличающийся тем, что этот способ предусматривает молекулярный механизм, включающий в себя стадии (а) конструирования одной или более систем возмещаемого блокирования функции (RBF) или комплексов ДНК-конструкций для инсертирования в реципиентную клетку или клеточную линию, содержащих трансгенный инсерт (TI), включающий один или более представляющих интерес трансгенов (TGI), кодирующих один или более желаемых генных продуктов, одну или более блокирующих конструкций (ВС), и обеспечения, по меньшей мере, одного контролируемого пользователем средства для восстановления блокированных функций; (i) причем указанная блокирующая конструкция (ВС) обладает способностью блокировать, по меньшей мере, одну молекулярную или физиологическую функцию, существенную для выживания или размножения размножающегося половым способом растения, причем блокирующая конструкция (ВС) локализована в той же самой хромосоме, что и трансгенный инсерт (TI), и в тесной близости от трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI), предпочтительно в интроне представляющего интерес трансгена (TGI) или фланкируя каждую сторону трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI); (ii) причем указанное средство для восстановления блокированной функции предусматривает одно или несколько контролируемых пользователем вмешательств, которые могут быть зависимыми или могут быть независимыми от одной или нескольких восстанавливающих конструкций (RC), которые помещены вблизи блокирующей конструкции (ВС) в трансгенном инсерте (TI), содержащем один или более представляющих интерес трансгенов (TGI), и/или в неаллельных хромосомах; и (b) восстановления блокированной функции, для того чтобы обеспечить возможность нормального развития во время культивирования и размножения указанного размножающегося половым способом растения для продукционных целей посредством применения на чувствительной стадии цикла развития или репродуктивного цикла указанного размножающегося половым способом растения одного или нескольких контролируемых пользователем вмешательств. 2. Способ по п.1, характеризующийся тем, что система возмещаемого блокирования функции (RBF) является системой двойного возмещаемого блокирования функции (D-RBF), содержащей, по меньшей мере, две блокирующие конструкции (ВС), которые являются одинаковыми или различными и которые возмещаемы одинаковыми или различными восстанавливающими функцию механизмами, фланкирующие каждую сторону трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI). 3. Способ по п.1, характеризующийся тем, что система возмещаемого блокирования функции (RBF) является системой множественного, предпочтительно тройного возмещаемого блокирования функции (M-RBF, T-RBF), содержащей два, предпочтительно три или более трансгенных инсертов (TI), помещенных в трех или более неаллельных хромосомах, в которой первый трансгенный инсерт (TI) содержит, по меньшей мере, две различные блокирующие конструкции (ВС), второй трансгенный инсерт (TI) содержит одну или более первых восстанавливающих конструкций (RC), связанных с необязательной блокирующей конструкцией (ВС), являющейся такой же или отличающейся от одной из блокирующих конструкций (ВС) первого трансгенного инсерта (TI), и третий трансгенный инсерт (TI) содержит одну или более вторых восстанавливающих конструкций (RC), которые отличаются от первых восстанавливающих конструкций (RC) и которые связаны с блокирующей конструкцией (ВС), соответствующей одному из первых трансгенных инсертов (TI), которая не является той же самой, что и необязательная блокирующая конструкция (ВС) второго трансгенного инсерта (TI). 4. Способ по п.1, характеризующийся тем, что для получения системы множественного возмещаемого блокирования функции (M-RBF) реципиентное трансгенное размножающееся половым способом растение обеспечивают двумя трансгенными инсертами (TI) системы обращенного сегрегирующего возмещаемого блокирования функции (RS-RBF) в двух неаллельных хромосомах без представляющего интерес трансгена (TGI) и трансформирующим вектором, содержащим две блокирующие конструкции (ВС), расположенные на каждой стороне клонирующего сайта для желаемых представляющих интерес трансгенов (TGI), предназначенным для трансформации реципиентной трансгенной линии размножающегося половым способом растения в третью неаллельную хромосому. 5. Способ по п.1, характеризующийся тем, что система возмещаемого блокирования функции (RBF) является системой сегрегирующего возмещаемого блокирования функции (S-RBF), содержащей восстанавливающую конструкцию (RC) и блокирующую конструкцию (ВС), помещенные в различные неаллельные хромосомы, причем блокирующая конструкция (ВС) находится в интроне трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI). 6. Способ по п.1, характеризующийся тем, что система индуцируемого возмещаемого блокирования функции (I-RBF) содержит восстанавливающую конструкцию (RC), помещенную в ту же самую хромосому, в которую помещена блокирующая конструкция (ВС), но экспрессируемую под контролем отличающегося промотора, причем блокирующая конструкция (ВС) или ее часть помещена в интрон трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI). 7. Способ по п.1, характеризующийся тем, что экспрессия, по меньшей мере, одной нуклеотидной последовательности в блокирующей конструкции (ВС) блокирует конститутивным, органоспецифическим или пространственно-временным образом молекулярную или физиологическую функцию, существенную для выживания или размножения трансгенного размножающегося половым способом растения. 8. Способ по п.1, характеризующийся тем, что удаление действия блокирующей конструкции (ВС) достигается применением контролируемого пользователем вмешательства, которое на чувствительной стадии в цикле развития или репродуктивного цикла трансгенного размножающегося половым способом растения снимает конститутивное, достигнутое органоспецифическим или пространственно-временным образом морфологическое или физиологическое блокирование, предотвращающее скрещивание и/или половое размножение этого трансгенного размножающегося половым способом растения. 9. Способ по п.8, характеризующийся тем, что эта чувствительная стадия, существенная для выживания и/или размножения, выбрана из группы, состоящей из раннего или позднего прорастания семян, цветения, образования карликового фенотипа, неспособности образования цветков в соцветии и/или плодов, неспособности к прорастанию, вегетации, корнеобразованию и фотосинтезу. 10. Способ по п.8, характеризующийся тем, что чувствительная стадия, существенная для выживания и/или размножения, выбрана из группы, состоящей из раннего или позднего прорастания семян. 11. Способ по п.1, характеризующийся тем, что удаление эффекта блокированной функции предусматривает контролируемый пользователем регулируемый извне или изнутри молекулярный механизм, необязательно применяемый в ограниченных условиях (в закрытом помещении). 12. Способ по п.11, характеризующийся тем, что контролируемый пользователем, регулируемый извне и/или изнутри молекулярный механизм для удаления эффекта блокированной функции выбран из группы, состоящей из (a) добавления, по меньшей мере, одного вводимого извне вещества; (b) применения, по меньшей мере, одного внешнего химического или физического стимула, способного активировать необязательную восстанавливающую конструкцию (RC); (c) репрессии промотора блокирующей конструкции (ВС); (d) подавления экспрессии мРНК нуклеотидной последовательности блокирующей конструкции (ВС) технологией антисмысловых РНК; (e) разрешения экспрессии белка, способного инактивировать белок, кодируемый нуклеотидной последовательностью блокирующей конструкции (ВС), посредством применения активируемых извне молекулярных механизмов; (f) разрешения экспрессии фермента, продуцирующего вещество, способное инактивировать белок, экспрессируемый нуклеотидной последовательностью блокирующей конструкции (ВС); (g) разрешения экспрессии фермента, способного инактивировать токсическое вещество, продуцируемое ферментом; (h) разрешения экспрессии фермента, способного продуцировать вещество, компенсирующее недостаточность, вызываемую действием блокирующей конструкции (ВС); (i) разрешения экспрессии фермента, способного продуцировать вещество, изменяющее действие, вызываемое блокирующей конструкцией (ВС); (j) разрешения специфического связывания нуклеиновой кислоты и/или белка; (k) применения протеолиза; (l) разрешения белок-белкового взаимодействия; (m) экспрессии фермента, способного к детоксикации токсического вещества; (n) экспрессии вещества, способного к компенсации или инактивации сверхпродуцируемого токсического вещества; (о) экспрессии вещества, способного к взаимодействию с токсическим веществом; (р) обеспечения фермента, способного превращать токсическое или сверхпродуцируемое вещество с неправильной пространственно-временной специфичностью; (q) компенсации извне недостаточности, вызываемой блокирующей конструкцией (ВС); (r) индуцирования извне реагирующего на внешний фактор возмещения ДНК, РНК, белка или метаболита; (s) конститутивной экспрессии нуклеотидной последовательности восстанавливающей функцию конструкции (RC); (t) пространственно-временной экспрессии нуклеотидной последовательности восстанавливающей конструкции (RC) и (u) разрешения внутрилинейного скрещивания в том случае, когда система возмещаемого блокирования функции (RBF) является системой сегрегирующего двойного или множественного возмещаемого блокирования функции (RBF). 13. Способ по п.1, характеризующийся тем, что контролируемый пользователем, регулируемый извне молекулярный механизм для возмещения блокированной функции предусматривает разрешение внутрилинейного скрещивания, поддерживающего гомозиготные состояния. 14. Комплекс ДНК-конструкций или система возмещаемого блокирования функции (RBF) для обеспечения увеличенного уровня надежности в контроле сегрегации трансгенов в размножающемся половым способом растении и/или обеспечения автоматического предотвращения интрогрессии, характеризующийся тем, что указанный комплекс ДНК-конструкций содержит: (a) трансгенный инсерт (TI), содержащий, по меньшей мере, один представляющий интерес трансген (TGI), кодирующий один или более желаемых генных продуктов; (b) одну или более блокирующих конструкций (ВС), которые являются возмещаемыми при помощи контролируемых пользователем средств для восстановления функции, причем указанная блокирующая конструкция (ВС) включает в себя, по меньшей мере, одну нуклеотидную последовательность, способную блокировать, по меньшей мере, одну молекулярную или физиологическую функцию, существенную для выживания или размножения размножающегося половым способом растения, и локализована в той же самой хромосоме, в которой находится трансгенный инсерт (TI), и в тесной близости от трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI), предпочтительно в интроне представляющего интерес трансгена (TGI) или фланкируя каждую сторону трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI); (с) одну или более необязательных восстанавливающих конструкций (RC), которые помещены вблизи блокирующих конструкций (ВС) в трансгенном инсерте (TI), содержащем один или более представляющих интерес трансгенов (TGI), помещенных в одной или более неаллельных хромосомах. 15. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что система возмещаемого блокирования функции (RBF) является системой двойного возмещаемого блокирования функции (D-RBF), содержащей блокирующие конструкции (ВС), которые являются, по меньшей мере, двумя одинаковыми или различными блокирующими конструкциями и расположены на обеих сторонах трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI), и могут возмещаться одинаковыми или различными механизмами возмещения. 16. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что система возмещаемого блокирования функции (RBF) является системой множественного возмещаемого блокирования функции (M-RBF), содержащей два, предпочтительно три или более трансгенных инсертов (TI), помещенных в трех или более неаллельных хромосомах, где первый трансгенный инсерт (TI) содержит, по меньшей мере, две различные блокирующие конструкции (ВС), второй трансгенный инсерт (TI) содержит одну или более первых восстанавливающих конструкций (RC), связанных с необязательной блокирующей конструкцией, соответствующей одной из блокирующих конструкций (ВС) первого трансгенного инсерта (TI), и третий трансгенный инсерт (TI) содержит одну или более вторых восстанавливающих конструкций (RC), которые являются одинаковыми или различными относительно первых восстанавливающих конструкций (RC) и которые связаны с блокирующей конструкцией (ВС), являющейся такой же, что и блокирующая конструкция (ВС) первого трансгенного инсерта, которая не является такой же, что и необязательная блокирующая конструкция (ВС) второго трансгенного инсерта (TI). 17. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что в системе множественного возмещаемого блокирования функции (M-RBF) реципиентное трансгенное размножающееся половым способом растение обеспечено двумя трансгенными инсертами (TI) системы обращенного сегрегирующего возмещаемого блокирования функции (RS-RBF) в двух неаллельных хромосомах без представляющего интерес трансгена (TGI) и трансформирующим вектором, содержащим блокирующие конструкции (ВС), помещенные на каждой стороне клонирующего сайта для желаемых представляющих интерес трансгенов (TGI), предназначенным для трансформации реципиентной трансгенной линии размножающегося половым способом растения в третью неаллельную хромосому. 18. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что система возмещаемого блокирования функции (RBF) является системой индуцируемого возмещаемого блокирования функции (I-RBF), содержащей восстанавливающую конструкцию (RC), помещенную в той же самой хромосоме, что и блокирующая конструкция (ВС), причем блокирующая конструкция (ВС) находится в интроне трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI). 19. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что система возмещаемого блокирования функции (RBF) является системой сегрегирующего возмещаемого блокирования функции (S-RBF), содержащей восстанавливающую конструкцию (RC) и блокирующую конструкцию (ВС), помещенные в различные неаллельные хромосомы, причем блокирующая конструкция (ВС) или ее часть находится в интроне трансгенного инсерта (TI), содержащего один или более представляющих интерес трансгенов (TGI). 20. Комплекс ДНК конструкций по п.14, характеризующийся тем, что индуцируемая восстанавливающая конструкция (RC) может активироваться в ответ на контролируемое пользователем вмешательство, что восстанавливает функцию, существенную для выживания и полового размножения трансгенного размножающегося половым способом растения, причем указанная функция блокирована блокирующей конструкцией (ВС), экспрессируемой под контролем того же самого или отличающегося типа промотора. 21. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что две нуклеотидные последовательности блокирующей конструкции (ВС), образующие систему двойного возмещаемого блокирования функции (D-RBF), возмещаются одинаковыми или различными средствами для возмещения. 22. Комплекс ДНК-конструкций по п.15, характеризующийся тем, что индуцируемая восстанавливающая конструкция (RC) может активироваться в ответ на контролируемое пользователем вмешательство, что восстанавливает функцию, блокированную блокирующей конструкцией (ВС), экспрессируемой под контролем того же самого или отличающегося промотора. 23. Комплекс ДНК-конструкций по п.13, характеризующийся тем, что блокирующая конструкция (ВС) содержит, по меньшей мере, одну нуклеотидную последовательность, конститутивная, органоспецифическая или пространственно-временная экспрессия которой блокирует существенную молекулярную или физиологическую функцию на уровне ДНК, мРНК, белка или метаболита и/или вызывает фенотипическое, физиологическое или морфологическое изменение, предотвращающее свободное скрещивание и/или размножение подвергаемого ауткроссингу трансгенного размножающегося половым способом растения в природных, неконтролируемых условиях, приводя к его исчезновению. 24. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что контролируемое пользователем средство для восстановления функции представляет собой контролируемый пользователем регулируемый извне или изнутри молекулярный механизм, необязательно применяемый в ограниченных условиях. 25. Комплекс ДНК-конструкций по п.24, характеризующийся тем, что контролируемый пользователем регулируемый извне или изнутри молекулярный механизм включает в себя механизм, выбранный из группы, состоящей из: (a) добавления, по меньшей мере, одного вводимого извне вещества; (b) применения, по меньшей мере, одного наружного химического или физического стимула, способного активировать необязательную восстанавливающую конструкцию (RC); (c) репрессии промотора блокирующей конструкции (ВС); (d) подавления экспрессии мРНК блокирующего гена технологией антисмысловых РНК; (e) разрешения экспрессии белка, способного инактивировать блокирующую конструкцию (ВС), кодирующую белок, посредством применения активируемых извне молекулярных механизмов; (f) разрешения экспрессии фермента, способного продуцировать вещество, инактивирующее белок, экспрессируемый блокирующей конструкцией (ВС); (g) разрешения экспрессии фермента, способного инактивировать токсическое вещество, продуцируемое ферментом; (h) разрешения экспрессии фермента, способного продуцировать вещество, компенсирующее недостаточность, вызываемую действием блокирующей конструкции (ВС); (i) разрешения экспрессии фермента, способного продуцировать вещество, компенсирующее изменение, вызываемое действием блокирующей конструкции (ВС); (j) разрешения специфического связывания нуклеиновой кислоты и/или белка; (k) применения протеолиза; (l) разрешения белок-белкового взаимодействия; (m) экспрессии фермента, способного к детоксикации токсического вещества; (n) экспрессии вещества, способного к компенсации или инактивации сверхпродуцируемого токсического вещества; (о) экспрессии вещества, способного к взаимодействию с токсическим веществом; (р) обеспечения фермента, продуцирующего токсическое или сверхпродуцируемое вещество; (q) компенсации извне недостаточности, вызываемой блокирующей конструкцией (ВС); (r) индуцирования извне реагирующего на внешний фактор возмещения ДНК, РНК, белка или метаболита; (s) конститутивной экспрессии нуклеотидной последовательности восстанавливающей функцию конструкции (RC); (t) пространственно-временной экспрессии нуклеотидной последовательности восстанавливающей конструкции (RC) и (u) разрешения внутрилинейного скрещивания в том случае, когда система возмещаемого блокирования функции (RBF) является системой множественного возмещаемого блокирования функции (RBF). 26. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что контролируемое пользователем средство для восстановления функции представляет собой контролируемый пользователем регулируемый извне молекулярный механизм, который включает разрешение внутрилинейного скрещивания. 27. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что блокирующая конструкция (ВС) содержит нуклеотидную последовательность, экспрессирующуюся в организме-хозяине конститутивным, органоспецифическим или пространственно-временным образом, а восстанавливающая конструкция (RC) содержит нуклеотидную последовательность, способную возмещаться посредством контролируемого пользователем вмешательства, регулирующего отвечающую на обработку нуклеотидную последовательность или ее генный продукт, или посредством конститутивной экспрессии этой нуклеотидной последовательности в восстанавливающей конструкции (RC) и/или ее генного продукта. 28. Комплекс ДНК-конструкций по п.27, характеризующийся тем, что блокирующая конструкция (ВС) содержит нуклеотидную последовательность, которая экспрессирует фермент органоспецифическим, зависящим от стадии развития или пространственно-временным образом, антисмысловую мРНК фермента, существенного для выживания или полового размножения организма-хозяина. 29. Комплекс ДНК-конструкций по п.28, характеризующийся тем, что блокирующая конструкция (ВС) содержит синтетическую нуклеотидную последовательность, адаптированную к предпочтениям организма-хозяина и/или предпочтениям интрона в отношении представляющего интерес трансгена (TGI), кодирующего желаемый продукт. 30. Комплекс ДНК-конструкций по п.29, характеризующийся тем, что синтетическая нуклеотидная последовательность кодирует фермент барназу, имеющую SEQ ID NO:1, и имеет последовательность нуклеотидов, адаптированную к предпочтениям растения и/или адаптированную к интрону гена растения, причем указанная последовательность является SEQ ID NO:3. 31. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что восстанавливающая конструкция (RC) содержит репрессорный пептид/белок, связывающийся с последовательностью оперона, которая регулирует экспрессию блокирующей нуклеотидной последовательности. 32. Комплекс ДНК-конструкций по п.31, характеризующийся тем, что репрессорный пептид/белок является репрессорным белком tet Tn10, a последовательность оперона является ДНК-последовательностью tet-оператора. 33. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что восстанавливающая конструкция (RC) содержит нуклеотидную последовательность, экспрессирующую антисмысловую РНК блокирующей нуклеотидной последовательности и способную активировать механизм подавления экспрессии блокирующей конструкции (ВС), компенсируя недостаточность, вызываемую действием блокирующей конструкции (ВС). 34. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что восстанавливающая конструкция (RC) содержит нуклеотидную последовательность, экспрессирующую белок, инактивирующий белок, экспрессируемый блокирующей конструкцией (ВС). 35. Комплекс ДНК-конструкций по п.34, характеризующийся тем, что нуклеотидная последовательность восстанавливающей конструкции (RC) экспрессирует белок Barstar, инактивирующий нуклеазу барназу путем связывания с ней. 36. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что блокирующая конструкция (ВС) содержит синтетическую нуклеотидную последовательность, которая адаптирована для того, чтобы быть инсертированной в интрон представляющего интерес трансгена (TGI), путем обеспечения предпочтения интрона в отношении нуклеотидной последовательности. 37. Комплекс ДНК-конструкций по п.36, характеризующийся тем, что синтетическая нуклеотидная последовательность, которая адаптирована в отношении инсертирования, является нуклеотидной последовательностью, по существу идентичной последовательности гена барназы, и имеет последовательность SEQ ID NO:3. 38. Комплекс ДНК-конструкций по п.36, характеризующийся тем, что синтетические последовательности генов барназы и Barstar, которые адаптированы для экспрессии в растениях, содержат SEQ ID NO:1 и SEQ ID NO:2, соответственно. 39. Комплекс ДНК-конструкций по п.36, характеризующийся тем, что он имеет синтетический сайт поликлонирования (SEQ ID NO:4), сконструированный с открытой рамкой считывания (ORF) и адаптированный для экспрессии 40. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что увеличенный уровень надежности означает, что частота инактивации блокирующей конструкции (ВС) является более низкой, чем 10-6, предпочтительно более низкой, чем 10-8, более предпочтительно более низкой, чем 10-10, наиболее предпочтительно более низкой, чем 10-12. 41. Комплекс ДНК-конструкций по п.14, характеризующийся тем, что увеличенный уровень надежности означает, что достигается 100% предотвращение утечки трансгена. 42. Клонирующий вектор, содержащий одну или более блокирующих конструкций (ВС), применимых для сборки комплекса ДНК-конструкций по п.14, характеризующийся тем, что этот вектор обеспечен сайтом поликлонирования для удобного переноса представляющего интерес трансгена (TGI) в подходящее положение в хромосоме размножающегося половым способом растения. 43. Клонирующий вектор по п.42, характеризующийся тем, что сайт поликлонирования представляет собой SEQ ID NO:4. 44. Способ получения клеток или клеточных линий размножающегося половым способом растения с увеличенным уровнем надежности в контроле сегрегации трансгенов и/или автоматическим предотвращением интрогрессии, предусматривающий введение одного или более трансгенных инсертов (TI) для сборки комплекса ДНК-конструкций по п.14 для удобной комплементации с вектором, обеспеченным представляющими интерес трансгенами (TGI), содержащим один или более представляющих интерес трансгенов (TGI) в клонирующем векторе (векторах) по любому из пп.42 и 43. 45. Способ получения трансгенного размножающегося половым способом растения с увеличенным уровнем надежности в контроле сегрегации трансгенов и/или автоматическим предотвращением интрогрессии, предусматривающий введение одного или более представляющих интерес трансгенных инсертов (TI), выбранных из группы, состоящей из блокирующих конструкций (ВС), необязательно объединенных с одной или более восстанавливающими конструкциями (RC), с, по меньшей мере, одним трансгенным инсертом (TI), имеющим один или более представляющих интерес трансгенов (TGI) или клонирующий сайт для инсертирования одного или более представляющих интерес трансгенов (TGI), для сборки комплекса ДНК-конструкций или системы возмещаемого блокирования функции (RBF) по п.14.
РИСУНКИ
MM4A Досрочное прекращение действия патента из-за неуплаты в установленный срок пошлины за
Дата прекращения действия патента: 17.07.2007
Дата публикации: 10.01.2011
|
||||||||||||||||||||||||||
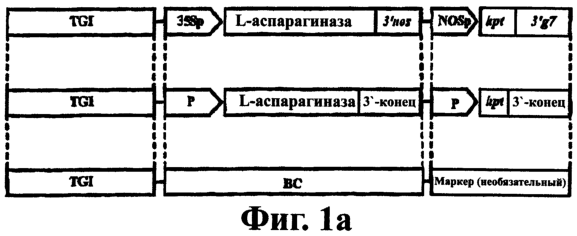
 или “система сегрегирующего возмещаемого блокирования функции (S-RBF)” означает тип системы возмещаемого блокирования функции (RBF), в котором восстанавливающая конструкция или конструкции (RC) помещены в одни и те же индивидуальные организмы в различных неаллельных хромосомах отдельно от блокирующей конструкции (ВС), которая тесно связана с представляющим интерес трансгеном (трансгенами) (TGI), т.е. комплексной TGI-BC-конструкции. Таким образом, сегрегация происходит независимо от комплексной TGI-BC-конструкции. Это означает, что блокирующий фактор не начинает действовать до второго гетерозиготного гибридного поколения (F2), когда блокирующая и восстанавливающая конструкции (ВС и RC) сегрегируют (расщепляются) в разные генеративные клетки, как описано ниже.
или “система сегрегирующего возмещаемого блокирования функции (S-RBF)” означает тип системы возмещаемого блокирования функции (RBF), в котором восстанавливающая конструкция или конструкции (RC) помещены в одни и те же индивидуальные организмы в различных неаллельных хромосомах отдельно от блокирующей конструкции (ВС), которая тесно связана с представляющим интерес трансгеном (трансгенами) (TGI), т.е. комплексной TGI-BC-конструкции. Таким образом, сегрегация происходит независимо от комплексной TGI-BC-конструкции. Это означает, что блокирующий фактор не начинает действовать до второго гетерозиготного гибридного поколения (F2), когда блокирующая и восстанавливающая конструкции (ВС и RC) сегрегируют (расщепляются) в разные генеративные клетки, как описано ниже. -галактозидазы в клонирующем векторе pUC19.
-галактозидазы в клонирующем векторе pUC19.