|
|
(21), (22) Заявка: 2003120859/15, 10.02.1997
(24) Дата начала отсчета срока действия патента:
10.02.1997
(30) Конвенционный приоритет:
09.02.1996 US 08/599,226
(43) Дата публикации заявки: 10.01.2005
(45) Опубликовано: 20.02.2006
(56) Список документов, цитированных в отчете о
поиске:
JP 2109994 А, 23.04.1990. JP 6319587 А, 22.11.1994. JP 4364133 А, 16.12.1992. NF 260226 А, 27.04.1995.
(62) Номер и дата подачи первоначальной заявки, из которой данная заявка выделена: 98117079 10.02.1997
Адрес для переписки:
127055, Москва, а/я 11, пат.пов. Н.К.Попеленскому
|
(72) Автор(ы):
АЛЛЕН Дебора Дж. (GB),
ВОГЕН Тристан Дж. (GB),
КАЙМАКСАЛАН Зера (TR),
МАКГИНЕСС Брайен Т. (GB),
МАНКОВИЧ Джон Э. (US),
РОБЕРТС Эндрю Дж. (GB),
САКОРАФАШ Поль (US),
СЭЛФЕЛД Джохен Г. (DE),
УАЙТ Майкл (US),
УИЛТОН Элисон Дж. (GB),
ХУГЕНБУМ Хендрикус Р. Дж. М. (NL),
ШОЭНХАУТ Дэвид (US),
ЛАБКОВСКИ Борис (US)
(73) Патентообладатель(и):
Абботт Байотекнолоджи эЛтиди. (BM)
|
(54) СПОСОБ ИНГИБИРОВАНИЯ АКТИВНОСТИ ЧЕЛОВЕЧЕСКОГО TNF (ВАРИАНТЫ), ПРИМЕНЕНИЕ ВЫДЕЛЕННОГО АНТИТЕЛА ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА В КАЧЕСТВЕ КОМПОНЕНТА ДЛЯ ПРОИЗВОДСТВА ЛЕКАРСТВЕННОГО СРЕДСТВА (ВАРИАНТЫ) И ВЫДЕЛЕННОЕ ЧЕЛОВЕЧЕСКОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (ВАРИАНТЫ), ПРИМЕНЕНИЕ ВЫДЕЛЕННОГО АНТИТЕЛА ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА В КАЧЕСТВЕ КОМПОНЕНТА ДЛЯ ПРОИЗВОДСТВА ЛЕКАРСТВЕННОГО СРЕДСТВА (ВАРИАНТЫ) И ВЫДЕЛЕННОЕ ЧЕЛОВЕЧЕСКОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ
(57) Реферат:
Изобретение относится к области медицины и касается применения выделенных антител человека или его фрагментов в качестве компонента для производства лекарственного средства, которое специфически связывает фактор некроза опухоли (hTNF). Эти антитела обладают высокой аффинностью в отношении hTNFa (например Kd=10-8 М или менее), низкой скоростью диссоциации (например Koff=10-3 с-1 или менее) и нейтрализуют активность hTNF in vitro и in vivo. Антитела, соответствующие изобретению, могут быть антителом полной длины или его антиген-связывающим фрагментом. Антитела или фрагменты антител, соответствующие изобретению, используют для детекции hTNF и для ингибрования активности hTNF, например, у человека, страдающего от нарушения, при котором активность hTNF является вредной. Преимущество изобретения заключается в разработке новых высокоаффинных рекомбинантных антител к hTNF или их фрагментов с низкой кинетикой диссоциацией. 8 н. и 7 з.п. ф-лы, 17 табл., 11 ил. 
Область техники, к которой относится изобретение
Изобретение относится к медицинской фармакологии и касается, в частности, применения антител человека, которые специфически связывают или нейтрализуют hTNF, при производстве лекарственных препаратов для лечения нарушений, при которых активность TNF является вредной.
Уровень техники
Фактор некроза опухоли (TNF) является цитокином, образуемым множеством типов клеток, включая моноциты и макрофаги, который был исходно идентифицирован на основе своей способности индуцировать некроз некоторых мышиных опухолей (см., например, Old L. (1985) Science 230:630-632). Впоследствии было показано, что фактор, называемый кахектин, ассоциированный с кахексией, является той же молекулой, что TNF. TNF участвовал в генерации шока (см., например, Beutler В. and Cerami A. (1988) Annu. Rev. Biochem. 57:505-518; Beutler В. and Cerami A. (1989) Annu. Rev. Immunol. 7:625-655). Более того, TNF участвовал в патофизиологии ряда других заболеваний и нарушений человека, включая сепсис, инфекции, аутоиммунные заболевания, отторжение трансплантата и болезнь трансплантат против хозяина (см..например, MoellerA. et al., (1990) Cytokine 2:162-169; Патент США No 5231024 Moeller et al.; Европейский Патент No 260610 B1 Moeller A. et al.; Vasilli P. (1992) Annu. Rev. Immunol 10:411-452; Tracey K.J. and Cerami A. (1994) Annu. Rev. Med. 45:491-503).
Вследствие вредной роли человеческого TNF (hTNF) в ряде нарушений у человека были разработаны терапевтические способы ингибирования или противодействия активности hTNF. В частности, антитела, которые связывают или нейтрализуют hTNF, рассматривались как средства ингибирования активности hTNF. Одними из самых ранних таких антител были мышиные моноклональные антитела (mAbs), секретируемые гибридомами, полученными из лимфоцитов мышей, иммунизированных hTNF (см., например, Hahn Т. et al. (1985) Proc. Natl. Acad. Sci. USA 82: 3814-3818; Liang C-M. et al. (1986) Biochem. Biophys. Res. Commun. 137: 847-854; Hirai M. et al.(1987) J. Immunol. Methods 96: 57-62; Fendly B.M. et al.(1987) Hybridoma 6:359-370; Moeller A. et al. (1990) Cytokine 2:162-169; Патент США No 5231024 Moeller et al.; Европейский Патент No 186833 B1 Wallach D.; Европейская Патентная Заявка No 218868 А1 Old et al.; Европейский Патент No 260610 B1 Moeller A. et al.). Поскольку эти мышиные антитела против hTNF часто проявляли высокую аффинность к hTNF (например, Kd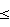 10-9 M) и были способны нейтрализовать активность hTNF, их применение in vivo могло ограничиваться проблемами, связанными с введением мышиных антител человеку, такими как короткий период полужизни в сывортке, неспособность включать определенные функции эффекторов человека и появление нежелательного иммунного ответа против мышиных антител у человека (реакция “человеческих антител против мыши” (НАМА)). 10-9 M) и были способны нейтрализовать активность hTNF, их применение in vivo могло ограничиваться проблемами, связанными с введением мышиных антител человеку, такими как короткий период полужизни в сывортке, неспособность включать определенные функции эффекторов человека и появление нежелательного иммунного ответа против мышиных антител у человека (реакция “человеческих антител против мыши” (НАМА)).
При попытке преодолеть проблемы, связанные с использованием полностью мышиных антител у человека, мышиные антитела против hTNF были с помощью генетической инженерии сделаны “человекоподобными”. Например, были получены химерные антитела, в которых вариабельные участки цепей антител имеют мышиную природу, а константные участки цепей антител – человеческую (Knight D.M. et al. (1993) Mol. Immunol. 30:1443-1453; Публикация РСТ No WO 92/16553 Daddona P.E. et al.). Кроме того, были также получены человекоподобные антитела, в которых гипервариабельные домены вариабельных участков антитела имеют мышиную природу, но остальные вариабельные участки и константные участки антител имеют человеческую природу (Публикация РСТ No WO 92/11383 Adair J.R. et al.). Однако поскольку эти химерные и гуманизированные антитела еще сохраняют некоторые мышиные последовательности, они еще могут вызвать нежелательную иммунную реакцию, реакцию человеческих антихимерных антител (НАСА), особенно при введении в течение длительных периодов, например при хронических показаниях, таких как ревматоидный артрит (см,, например, Elliott M.J. et al. (1994) Lancet 344:1125-1127; Elliott M.J. et al. (1994) Lancet 344: 1105-1110).
Предпочтительным ингибирующим hTNF агентом по отношению к мышиным mAbs и их производным (например, химерным или человекоподобным антителам) было бы полностью человеческое антитело против hTNF, поскольку такой агент не вызывал бы реакции НАМА, даже если бы применялся в течение длительных периодов. Человеческие моноклональные аутоантитела против hTNF были получены способами с использованием человеческой гибридомы (Boyle P. Et al. (1993) Cell. Immunol. 152:556-568; Boyle P. et al. (1993) Cell. Immunol. 152:569-581; Европейская Патентная Заявка No 614984 А2 Boyle et al.). Однако, как сообщалось, эти моноклональные аутоантитела гибридомной природы обладали настолько низкой аффинностью в отношении hTNF, что ее трудно было подсчитать известными способами, не были способны к связыванию растворимого hTNF и не были способны нейтрализовать hhTNF-индуцированную цитотоксичность (см. Boyle et al., supra). Более того, успех способа с использованием человеческой гибридомы зависит от естественного присутствия в периферической крови человека лимфоцитов, продуцирующих аутоантитела, специфичные для hTNF. В ряде исследований у человека обнаружены сывороточные аутоантитела против hTNF (Fomsgaard A. et al. (1989) Scand. J. Immunol. 30: 219-223; Bendtzen К. et al. (1990) Prog. Leukocyte Biol. 10B: 447-452), тогда как в других исследованиях они не были обнаружены (Leusch H-G. et al. (1991) J.Immunol. Methods 139: 145-147).
Альтернативой существующим в естественных условиях человеческим антителам против hTNF были бы рекомбинантные антитела против hhTNF. Описаны рекомбинантные человеческие антитела, которые связывают hTNF с относительно низкой аффинностью (т.е. Кd 10-7 М) и быстрой скоростью диссоциации (т.е. Кoff 10-2 с-1). Однако ввиду относительно быстрой кинетики диссоциации эти антитела не могут быть применены для терапевтических нужд. Кроме того, были описаны рекомбинантные человеческие антитела против hTNF, которые не нейтрализуют активность DTNF, но значительно усиливают связывание hTNF с поверхностью клеток и усиливают внедрение в цитоплазму hTNF (Lidbury A. et al. (1994) Biotechnol. Ther. 5:27-45; Публикация РСТ No WO 92/03145 Aston R. et al.).
Соответственно человеческие антитела, такие как рекомбинантные человеческие антитела, которые связывают растворимый hTNF с высокой аффинностью и низкой кинетикой диссоциации и которые способны нейтрализовать активность hTNF, включая hTNF-индуцированную цитотоксичность (in vitro и in vivo) и hTNF-индуцированную активацию клеток, все еще являются необходимыми.
Сущность изобретения
Изобретение относится к применению человеческих антител, в частности, рекомбинантных человеческих антител, которые специфически связывают человеческий TNF. Антитела, соответствующие изобретению, характеризуются связыванием hTNF с высокой аффинностью и медленной кинетикой диссоциации, а также нейтрализацией активности hTNF, включая hTNF-индуцированную цитотоксичность (in vitro и in vivo) и hTNF-индуцированную активацию клеток. Антитела, соответствующие изобретению, далее характеризуются связыванием hTNF, но не hTNF (лимфотоксина) и наличием способности связывать кроме человеческого TNF также факторы некроза опухоли (TNF) других приматов и TNF неприматов.
Антитела, соответствующие изобретению, могут быть полной длины (например, антитело lgG1 или lgG4) или могут содержать только антиген-связывающий фрагмент (например, фрагмент Fab, F(ab’)2 или scFv). Наиболее предпочтительное рекомбинантное антитело, соответствующее изобретению, обозначаемое D2E7, имеет домен CDR3 легкой цепи, содержащий аминокислотную последовательность SEQ ID No 3 и домен CDR3 тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No 4. Предпочтительно антитело D2E7 имеет вариабельный участок легкой цепи (LCVR), содержащий аминокислотную последовательность SEQ ID No 1 и вариабельный участок тяжелой цепи (HCVR), содержащий аминокислотную последовательность SEQ ID No 2.
В изобретении предлагаются способ ингибирования активности человеческого TNF, в котором осуществляют контактирование человеческого TNF с выделенным антителом человека или его антиген-связывающим фрагментом и при этом ингибируют активность человеческого TNF, а также применение выделенного антитела человека или его антиген-связывающего фрагмента в качестве компонента для производства средства (лекарственного препарата) для лечения нарушений, при которых активность TNF является вредной, посредством введения антитела субъекту. В одном из вариантов осуществления изобретения упомянутое выделенное человеческое антитело или его антиген-связывающий фрагмента диссоциирует из человеческого TNF с Кd равной 1×10-8 М или менее и c константой скорости Кoff 1×10-3 с-1 или менее (оба значения определены с помощью поверхностного плазменного резонанса) и нейтрализуют цитотоксичность человеческого hTNF в стандартном анализе с использованием L929 in vitro с IC50 равной 1×10-7 М или меньше. Более предпочтительно, если выделенное человеческое антитело или его антиген-связывающий фрагмент диссоциируют из ассоциации (комплекса) с человеческим TNF с Кoff равной 5×10-4 с-1 или менее и даже более предпочтительно с Кoff 1×10-4 с-1 или менее. Более предпочтительно, если выделенное человеческое антитело или его антиген-связывающий фрагмент нейтрализуют цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50 1×10-8 М или менее и еще более предпочтительно с IC50 5×10-10 М или менее.
В другом варианте осуществления изобретения используется человеческое антитело или его антиген-связывающий фрагмент со следующими характеристиками:
a) диссоциирует из hTNF с Кoff 1×10-3 с-1 или менее, что определено с помощью поверхностного плазменного резонанса;
b) имеет по меньшей мере домен CDR3 легкой цепи, содержащий аминокислотную последовательность SEQ ID No 3 или SEQ ID No 3, модифицированную посредством замены одним аланином в положении 1, 4, 5, 7 или 8 или заменой от одной до пяти консервативных аминокислот в положениях 1, 3, 4, 6, 7, 8 и/или 9;
c) имеет по меньшей мере домен CDR3 тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No 4 или SEQ ID No 4, модифицированную посредством замены одним аланином в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11 или заменой от одной до пяти консервативных аминокислот в положениях 2, 3, 4, 5, 6, 8, 9, 10, 11 и/или 12.
Более предпочтительно, если антитело или его антиген-связывающий фрагмент диссоциируют из человеческого TNF с Кoff 5×10-4 с-1 или менее. Еще более предпочтительно, если антитело или его антиген-связывающий фрагмент диссоциируют из человеческого TNF с Кoff 1×10-4 с-1 или менее.
В еще одном варианте осуществления изобретения используется выделенное человеческое антитело или его антиген-связывающий фрагмент с LCVR, содержащим аминокислотную последовательность SEQ ID No 1, и с HCVR, содержащим аминокислотную последовательность SEQ ID No 2. В некоторых вариантах осуществления антитело может иметь константный участок тяжелой цепи lgG1 или константный участок тяжелой цепи lgG4. В других вариантах осуществления антитело является фрагментом Fab, фрагментом F(ab’)2 или фрагментом Fv одной цепи.
В следующем варианте осуществления изобретение в качестве компонента для производства упомянутого средства для лечения нарушений предлагается применение антитела D2E7.
В других вариантах осуществления изобретение представляет антитела или его антиген-связывающие фрагменты с LCVR, имеющим домен CDR3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No3, SEQ ID No 11, SEQ ID No 12, SEQ ID No 13, SEQ ID No 14, SEQ ID No 15, SEQ ID No 16, SEQ ID No 17, SEQ ID No 18, SEQ ID No 19, SEQ ID No 20, SEQ ID No 21, SEQ ID No 22, SEQ ID No 23, SEQ ID No 24, SEQ ID No 25, SEQ ID No 26, или с HCVR, имеющим домен CDR3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No 4, SEQ ID No 27, SEQ ID No 28, SEQ ID No 29, SEQ ID No 30, SEQ ID No 31, SEQ ID No 32, SEQ ID No 33, SEQ ID No 34 и SEQ ID No 35.
В еще одном варианте осуществления изобретение представляет выделенное человеческое антитело или его антиген-связывающий фрагмент, которые нейтрализуют активность человеческого TNF, но не человеческого TNF (лимфотоксина). В предпочтительном варианте осуществления человеческое антитело или его антиген-связывающий фрагмент нейтрализуют активность человеческого TNF, TNF шимпанзе и TNF по меньшей мере еще одного примата, выбранного из группы, состоящей из hTNF павиана, TNF игрунки, hTNF циномолгуса и TNF резуса. Предпочтительно, если антитело также нейтрализует активность TNF по меньшей мере одного непримата. Например, в одном варианте осуществления выделенное человеческое антитело или его антиген-связывающий фрагмент нейтрализуют также активность собачьего TNF. А в другом варианте выделенное человеческое антитело или его антиген-связывающий фрагмент нейтрализуют также активность свиного TNF. В еще одном варианте осуществления выделенное человеческое антитело или его антиген-связывающий фрагмент нейтрализуют также активность мышиного TNF.
Другой аспект изобретения относится к молекулам нуклеиновых кислот, кодирующих соответствующие изобретению антитела или их антиген-связывающие фрагменты. Предпочтительная нуклеиновая кислота, соответствующая изобретению, кодирующая LCVR D2E7, имеет нуклеотидную последовательность, представленную на Фигуре 7, и SEQ ID No 36. Другая предпочтительная нуклеиновая кислота, соответствующая изобретению, кодирующая HCVR D2E7, имеет нуклеотидную последовательность, представленную на Фигуре 8, и SEQ ID No 37. Рекомбинантные экспрессирующие векторы, несущие кодирующие антитела нуклеиновые кислоты, соответствующие изобретению, и клетки-хозяева, в которые интродуцированы такие векторы, также охватываются изобретением, как и способы получения антител, соответствующих изобретению, посредством культивирования клеток-хозяев, соответствующих изобретению.
Еще один аспект изобретения относится к способам ингибирования активности человеческого TNF с использованием антитела или его антиген-связывающегофрагмента, соответствующего изобретению. В одном варианте осуществления способ предусматривает контактирование человеческого ТМР с антителом, соответствующим изобретению, или его антиген-связывающим фрагментом, при этом ингибируется активность человеческого TNF. В другом варианте осуществления способ предусматривает введение антитела, соответствующего изобретению, или его антиген-связывающего фрагмента человеку, страдающему от нарушения, при котором активность TNF является вредной, при этом у человека ингибируется активность TNF. Нарушением может быть, например, сепсис, аутоиммунное заболевание (например, ревматоидный артрит, аллергия, рассеянный склероз, аутоиммунный диабет, аутоиммунный увеит и почечный синдром), инфекционное заболевание, злокачественное новообразование, отторжение трансплантата или болезнь трансплантат против хозяина, легочное нарушение, костное нарушение, кишечное нарушение или сердечное нарушение.
Упомянутые выделенные антитела человека или его антиген-связывающего фрагмента согласно изобретению могут применяться для производства средств для лечения, в частности, следующих нарушений: аутоиммунные заболевания, инфекционные болезни, болезни, связанные с отторжением трансплантата (болезни трансплантат против хозяина), нарушения, связанные со злокачественным образованием, легочные нарушения, кишечные нарушения, сердечные нарушения, сепсис, ревматоидный артрит, ревматоидный спондилит, остеоартрит, подагрический артрит, аллергия, рассеянный склероз, аутоиммунный диабет, аутоиммунный увеит, почечный синдром, воспалительные костные заболевания, заболевания, связанные с резорбцией кости, алкогольный гепатит, вирусный гепатит, нарушения свертывания крови, ожоги, реперфузионные повреждения, нарушения, связанные с образованием келоида, нарушения, связанные с формированием рубцовой ткани, гипертермия, септический шок, эндотоксический шок, сепсис, вызываемый грамотрицательными бактериями, токсический шоковый синдром, малярия, менингит, кахексия, СПИД, бактериальный менингит, СПИД-ассоциированный комплекс (ARC), вторичная инфекция цитомегаловируса при трансплантации, нарушения, связанные с лихорадкой и миалгией, обусловленные инфекцией, и вторичная кахексии при инфекции, нарушения, связанные с отторжением аллотрансплантата, нарушения, связанные со стимуляцией роста опухоли, нарушения, связанные с усилением метастатического потенциала и появлением цитотоксичности при злокачественных образованиях, нарушения, связанные с ингибированием роста опухоли или метастазов, нарушения, связанные с респираторным дистресс-синдромом взрослых, легочный шок, хронические легочные воспалительные заболевания, легочный саркоидоз, легочный фиброз, силикоз, воспалительные кишечные нарушения, идиопатические воспалительные заболевания кишечника, болезнь Крона, язвенный колит, ишемия сердца, сердечная недостаточность, гепатит.
Упомянутые выделенные антитела человека или его антиген-связывающего фрагмента согласно изобретению могут также применяться для введения антитела человеку вместе с цитокином интерлейкином-6 (IL-6) или введения человеку, у которого концентрация IL-6 в сыворотке или плазме превышает 500 пг/мл, а также для производства средства, в котором выделенное антитело используется в комбинации с фармацевтически приемлемым носителем, или для производства средства для введения выделенного антитела с по меньшей мере одним дополнительным терапевтическим агентом.
В предпочтительном варианте осуществления изобретение в качестве упомянутого субъекта выступает человек и применение упомянутых выделенных антител человека или его антиген-связывающего фрагмента согласно изобретению предназначается для производства лекарственного препарата для лечения нарушений, при которых активность TNF является вредной, посредством введения антитела человеку для ингибирования активности человеческого TNF.
Перечень фигур чертежей
На Фигурах 1 и 2 представлены последовательности нуклеиновых кислот вариабельного участка легкой цепи D2E7 (D2E7 VL; показано также в SEQ ID No 1), аланин-сканмутанты D2E7VL (LD2E7*.A1, LD2E7*.A3, LD2E7*.A4, LD2E7*.A5, LD2E7*.A7 и LD2E7*.A8), вариабельный участок легкой цепи подобного D2E7 антитела 2SD4 (2SD4 VL; показано также в SEQ ID No 9) и другие вариабельные участки легкой цепи, подобные D2E7 (ЕР В12, VL10E4, VL100А9, VL100D2. VL10F4, LOE5, VLLOF9, VLLOF10, VLLOG7, VLLOG9, VLLOH1, VLLOH10, VL1B7, VL1С1, VL1C7, VL0.1F4.VL0.1H8, LOE7, LOE7.A и LOE7.T). На Фигуре 1А представлены домены FR1, CDR1, FR2 и CDR2. На Фигуре 1В представлены домены FR3, CDR3 и FR4. Домены легкой цепи CDR1 (“CDR L1”), CDR2 (“CDR L2”) и CDR3 (“CDR L3”) находятся в прямоугольнике.
На Фигурах 3 и 4 представлены аминокислотные последовательности вариабельного участка D2E7 (D2E7 VH; показано также в SEQ ID No 2), аланин-сканмутанты D2E7VH (HD2E7*.A1, HD2E7*.A2, HD2E7*.A3, HD2E7*.A4, HD2E7*.A5, HD2E7*.A6, HD2E7*.A7, HD2E7*.A8 и HD2E7*.A9), вариабельный участок тяжелой цепи, подобного D2E7 антитела 2SD4 (2SD4 VH, показано также в SEQ ID No 10) и другие вариабельные участки тяжелой цепи, подобные D2E7 (VH1B11, VH1D8, VH1A11, VH1B12, VH1-D2, VH1E4, VH1F6, VH1G1, 3С-Н2, VH1-D2.N и VH1-D2.Y). На Фигуре 2А представлены домены FR1, CDR1, FR2 и CDR2. На Фигуре 2В представлены домены FR3, CDR3 и FR4. Домены тяжелой цепи CDR1 (“CDR H1”), CDR2 (“CDR H2”) и CDR3 (“CDR Н3”) находятся в прямоугольнике.
На Фигуре 5 графически представлено ингибирование TNF-индуцированной цитотоксичности L929 человеческим антителом D2E7 против hTNF по сравнению с мышиным антителом МАК 195 против hTNF.
На Фигуре 6 графически представлено ингибирование связывания rhTNF с рецепторами hTNF на клетках U-937 с помощью человеческого антитела D2E7 против hTNF по сравнению с мышиным антителом МАК 195 против hTNF.
На Фигуре 7 графически представлено ингибирование индуцированной TNF экспрессии ELAM-1 на HUVEC с помощью человеческого антитела D2E7 против hTNF по сравнению с мышиным антителом МАК 195 против hTNF.
На Фигуре 8 представлен график в виде столбиков защиты от TNF-индуцированной гибели сенсибилизированных D-галактозамином мышей посредством введения человеческого антитела против hTNF D2E7 (черные столбики) по сравнению с мышиным антителом МАК 195 против hTNF (заштрихованные столбики).
На Фигуре 9 представлена нуклеотидная последовательность вариабельного участка легкой цепи D2E7c прогнозированной аминокислотной последовательностью, находящейся ниже нуклеотидной последовательности. Участки CDR L1, CDR L2 и CDR L3 подчеркнуты.
На Фигуре 10 представлена нуклеотидная последовательность вариабельного участка тяжелой цепи D2E7 с прогнозированной аминокислотной последовательностью, находящейся ниже нуклеотидной последовательности. Участки CDR H1, CDR H2 и CDR H3 подчеркнуты.
На Фигуре 10 приведен график, представляющий эффект лечения антителами D2E7 трансгенных мышей Тg197 со средним размером суставов в качестве модели полиартрита.
Сведения, подтверждающие возможность осуществления изобретения
Данное изобретение относится к выделенным человеческим антителам или их антиген-связывающим фрагментам, которые связывают TNF с высокой аффинностью, низкой скоростью диссоциации и высокой нейтрализующей активностью, и применению этих антител для ингибирования активности человеческого TNF и в качестве компонентов лекарственных средств для лечения различных нарушений, при которых активность TNF является вредной. В частности в изобретении предлагаются способы применения антител, соответствующих изобретению, для детекции человеческого TNF или для ингибирования активности человеческого TNF либо in vitro, либо in vivo.
С целью более легкого понимания изобретения сначала определяются некоторые термины.
Термин “человеческий TNF” (сокращаемый здесь как hTNF или просто hTNF), используемый здесь, предназначен для определения человеческого цитокина, который существует в виде секретируемой формы с молекулярной массой 17 кД и ассоциированной с мембаной формы с молекулярной массой 26 кД, биологически активная форма которого содержит тример нековалентно связанных молекул с массой 17 кД. Структура hTNF описана далее в, например, Pennica D. et al., (1984) Nature 312:724-729; Davis J.M. et al. (1987) Biochemistry 26:1322-1326; Jones E.Y. et al,(1989) Nature 338:225-228. Термин человеческий TNF предполагает включение рекомбинантного человеческого TNF (rhTNF), который может быть получен стандартными способами рекомбинантной экспрессии или закуплен (R & D Systems, Catalog No. 210-TA, Minneapolis, MN).
Термин “антитело”, используемый здесь, предназначен для определения молекул иммуноглобулина, состоящего из четырех полипептидных цепей (две тяжелые (Н) цепи и две легкие (L) цепи), связанных дисульфидными связями. Каждая тяжелая цепь содержит вариабельный участок тяжелой цепи (сокращенный здесь как HCVR или VH) и константный участок тяжелой цепи. Костантный участок тяжелой цепи содержит три домена СН1, СН2 и СН3. Каждая легкая цепь содержит вариабельный участок легкой цепи (сокращенный здесь как LCVR или VL) и константный участок легкой цепи. Константный участок легкой цепи содержит один домен CL Участки VH и VL могут далее подразделяться на участки гипервариабельности, называемые определяющими комплементарность участками (CDR), окруженные участками, которые являются более консервативными, называемыми скелетными участками (FR). Каждая из VH и VL состоит из трех CDR и четырех FR участков, расположенных от амино- до карбоксильного конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4.
Термин “антиген-связывающий фрагмент” антитела (или просто “фрагмент антитела”), используемый здесь, относится к одному или более фрагментам антитела, которые сохраняют способность специфически связывать антиген (например, hTNF). Показано, что антиген-связывающая функция антитела может быть осуществлена фрагментами антитела полной длины. Примеры связывающих фрагментов, охватываемые термином “антиген-связывающий фрагмент” антитела, включают (i) фрагмент Fab, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CHl; (ii) фрагмент Р(ab’)2, бивалентный фрагмент, содержащий два фрагмента Fab, связанные дисульфидным мостиком в районе петли; (iii) фрагмент Fd, состоящий из доменов VH и СН1; (iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела; (v) фрагмент dAb (Ward et al., (1989) Nature 341:544-546), который состоит из домена VH, и (vi) выделенный участок (CDR), определяющий комплементарность. Более того, хотя два домена фрагмента Fv, VL и VH, кодируются отдельными генами, они могут быть рекомбинантными способами связаны с помощью синтетического линкера, который обеспечивает их получение в виде одной белковой цепи, в которой участки VL и VH спарены с образованием моновалентных молекул (известных как Fv одной цепи (scFv); см., например, Bird et al. (1988) Science 242:423-426; Huston et al.,(1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Предполагается, что такие антитела из одной цепи также охватываются термином “антиген-связывающий фрагмент” антитела. К ним относятся также другие формы антител из одной цепи, такие как диатела. Диатела являются бивалентными, биспецифическими антителами, в которых домены VH и VL экспрессируются на одной полипептидной цепи, но с использованием линкера, который является слишком коротким, чтобы позволять спаривание двух доменов на одной и той же цепи, что заставляет домены спариваться с комплементарными доменами другой цепи и создавать два антиген-связывающих сайта (см., например, Holliger Р. et al. (1993) Proc. Natl. Acad. Sci. USA 90:6444-6448; Poljak R.J. et al. (1994) Structure 2:1121-1123).
Далее, антитело или его антиген-связывающий фрагмент могут быть частью более крупных молекул иммуноадгезии, образованных ковалентной или нековалентной связью антитела или фрагмента антитела с одним или более белком или пептидом. Примеры таких молекул иммуноадгезии включают использование участка ядра стрептавидина для получения тетрамерной молекулы scFv (Kipriyanov S.M. et al (1995) Human Antibodies and Hybridomas 6:93-101) и использование остатка цистеина, маркерного пептида и С-концевой полигистидиновой метки для получения бивалентных и уменьшенных биомолекул scFv (Kipriyanov S.M. et al (1994) Mol. Immunol., 31:1047-1058). Фрагменты антител, такие как Fab F(ab’)2, могут быть получены из целых антител с использованием принятых способов, таких как разложение папаином или пепсином соответственно целых антител. Более того, антитела, фрагменты антител и молекулы иммуноадгезии могут быть получены с использованием стандартных способов с применением рекомбинантной ДНК, как описано здесь.
Термин “человеческое антитело”, используемый здесь, включает антитела, имеющие вариабельные и константные участки, выделенные из последовательностей зародышевого иммуноглобулина человека. Человеческие антитела, соответствующие изобретению, могут включать остатки аминокислот, не кодируемые последовательностями зародышевого иммуноглобулина человека (например, мутации, интродуцированные ненаправленным или сайт-специфическим мутагенезом in vitro или соматической мутацией in vivo), например, в CDR и особенно в CDR3. Однако термин “человеческое антитело”, используемый здесь, не включает антитела, в которых последовательности CDR, выделенные из эмбрионов других видов млекопитающих, таких как мышь, были пересажены на человеческие скелетные последовательности.
Термин “рекомбинантное человеческое антитело”, используемый здесь, включает все человеческие антитела, которые получены, экспрессированы, созданы или выделены рекомбинантными средствами, такие как антитела, экспрессированные с использованием рекомбинантного экспрессирующего вектора, введенного в клетку-хозяин (описано далее в Разделе II ниже), антитела, выделенные из набора известных рекомбинантных комбинаторных человеческих антител (описано далее в Разделе III ниже), антитела, выделенные из животного (например, мыши), которое является трансгенным в отношении генов человеческого иммуноглобулина (см., например, Taylor L.D. et al. (1992) Nucl. Acids Res. 20:6287-6295) или антитела, полученные, экспрессированные, созданные или выделенные любыми другими средствами, которые включают сплайсинг последовательности гена человеческого иммуноглобулина до других последовательностей ДНК. Такие рекомбинантные человеческие антитела имеют вариабельные и константные участки, выделенные из последовательностей человеческого зародышевого иммуноглобулина. В некоторых вариантах осуществления, однако, такие рекомбинантные человеческие антитела подвергают мутагенезу in vitro (или, если используют животное, трансгенное по последовательностям Ig человека, соматическому мутагенезу in vivo) и, таким образом, аминокислотные последовательности участков VH и VL рекомбинантных антител являются последовательностями, которые, поскольку они выделены из последовательностей зародышевых VH и VL человека и близки к ним, не могут в естественных условиях существовать в зародышевом наборе антител человека in vivo.
“Выделенное антитело”, как используют здесь, предназначено для определения антитела, которое практически не содержит других антител, имеющих различные антигенные специфичности (например, выделенное антитело, которое специфически связывает hTNF, практически не содержит антител, которые специфически связывают антигены, отличные от hTNF). Выделенное антитело, которое специфически связывает hTNF, может, однако, иметь перекрестную реактивность с другими антигенами, такими как молекулы TNF из других видов (детально обсуждается ниже). Более того, выделенное антитело может практически не содержать иной клеточный материал и/или химические соединения.
“Нейтрализующее антитело”, как используют здесь, (или “антитело, которое нейтрализовало активность hTNF) подразумевает антитело, связывание которого с hTNF приводит к ингибированию биологической активности hTNF. Это ингибирование биологической активности hTNF может быть оценено измерением одного или более индикаторов биологической активности hTNF, таких как hTNF-индуцированной активации клетки и связывания hTNF с рецепторами hTNF. Эти индикаторы биологической активности hTNF могут быть оценены одним или более методом различных стандартных анализов in vitro и in vivo, известных в уровне техники (см. Пример 4). Предпочтительно способность антитела нейтрализовать активность hTNF оценивается по ингибированию hTNF-индуцированной цитотоксичности клеток L929. В качестве дополнительного или альтернативного параметра активности hTNF может быть оценена способность антитела ингибировать hTNF-индуцированную экспрессию ELAM-1 на HUVEC как степень hTNF-индуцированной активации клеток.
Термин “поверхностный плазменный резонанс”, используемый здесь, относится к оптическому явлению, положенному в основу проведения анализа протекающих в данный момент биоспецифических взаимодействий по детекции изменений концентраций белка в матриксе биосенсора, например, с использованием системы BIAcore (Pharmacia Biosensor AB, Uppsala, Sweden and Piscataway, NJ). Дальнейшее описание см. в Примере 1 и Jönsson U. et al. (1993) Ann. Biol. Clin. 51:19-26; Jönsson U. et al. (1991) Biotechniques 11:620-627; Johnson B. et al. (1995) J. Mol. Recognit. 8:125-131 и Johnson В. et al. (1991) Anal. Biochem. 198:268-277.
Термин “Koff“, используемый здесь, предназначен для определения константы скорости диссоциации антитела из комплекса антитело/антиген.
Термин “Кd“, используемый здесь, предназначен для обозначения константы диссоциации определенного взаимодействия антитело-антиген.
Термин “молекула нуклеиновой кислоты”, используемый здесь, включает молекулы ДНК и молекулы РНК. Молекула нуклеиновой кислоты может быть однонитиевой или двунитевой, но предпочтительно двунитевой ДНК.
Термин “выделенная молекула нуклеиновой кислоты”, используемый здесь в отношении нуклеиновых кислот, кодирующих антитела или фрагменты антител (например, VH, VL, CDR3), которые связывают hTNF, предназначен для определения молекулы нуклеиновой кислоты, в которой нуклеотидные последовательности, кодирующие антитела или фрагменты антител, не содержат другие нуклеотидные последовательности, кодирующие антитела или фрагменты антител, которые связывают антигены, отличные от hTNF, эти другие последовательности могут в естественных условиях примыкать к нуклеиновой кислоте в геномной ДНК человека. Таким образом, например, выделенная нуклеиновая кислота, соответствующая изобретению, кодирующая участок VH антитела против TNF, не содержит другие последовательности, кодирующие другие участки VH, которые связывают антигены, отличные от TNF.
Термин “вектор” в контексте описания обозначает молекулу нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой он был связан. Одним типом вектора является “плазмида”, которая означает кольцевую петлю двунитевой ДНК, в которую могут быть лигированы дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, в котором дополнительные сегменты ДНК могут быть лигированы в геном вируса. Определенные векторы способны к автономной репликации в клетке-хозяине, в которую они интродуцированы (например, бактериальные векторы, имеющие бактериальный ориджин (точку начала) репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина введением в клетку-хозяин и вследствие этого реплицируются вместе с геномом хозяина. Более того, определенные векторы могут направлять экспрессию генов, с которыми они оперативно связаны. Такие векторы называются в контексте заявки “рекомбинантные экспрессирующие векторы” (или просто “экспрессирующие векторы”). В основном экспрессирующие векторы для использования в способах с применением рекомбинантной ДНК часто находятся в форме плазмид. В настоящем описании “плазмида” и “вектор” могут быть использованы взаимозаменяемо, поскольку плазмида является наиболее часто применяемой формой вектора. Однако изобретение включает другие формы экспрессирующих векторов, такие как вирусные векторы (например, репликационно-дефектные ретровирусы, аденовирусы и аденоассоциированные вирусы), которые выполняют эквивалентные функции.
Термин “рекомбинантная клетка-хозяин” (или просто “клетка-хозяин”) в контексте заявки предназначен для определения клетки, в которую интродуцирован рекомбинантный экспрессионный вектор. Следует понимать, что такие термины обозначают только определенную данную клетку, но не потомство такой клетки. Поскольку определенные модификации в последующих поколениях могут происходить вследствие мутаций или воздействия окружающей среды, такое потомство может на самом деле не быть идентичными родительской клетке, но также подпадает под определение термина “клетка-хозяин” в контексте заявки.
Различные аспекты изобретения подробно описываются в следующих подразделах.
1. Человеческие антитела, которые связывают TNF человека
В изобретении представлены выделенные человеческие антитела или их антиген-связывающие фрагменты, которые связывают человеческий TNF с высокой аффинностью, низкой скоростью диссоциации и высокой нейтрализующей способностью. Предпочтительно человеческие антитела, соответствующие изобретению, являются рекомбинантными нейтрализующими человеческими антителами против hTNF. Наиболее предпочтительным рекомбинантным нейтрализующим антителом согласно изобретению является D2E7, которое имеет последовательности VL и VH, как показано на Фигурах 1, 2 и Фигурах 3 4, соответственно (аминокислотная последовательность участка VL D1E7 также представлена как SEQ ID No 1; аминокислотная последовательность участка VH D1 Е7 также представлена как SEQ ID No 2). Связывающие свойства D2E7 по сравнению с мышиными mAB МАК 195 против hTNF, которые обладают высокой аффинностью и низкой кинетикой диссоциации, и другими человеческими антителами против hTNF 2SD4 с близкой D2E7 последовательностью сведены ниже в таблицу:
| Антитело |
Koff c-1 |
kon М-1 сек-1 |
Кd М |
Стехиометрия |
| D2E7 lgG1 |
8,81×10-5 |
1,91×105 |
6,09×10-10 |
1,2 |
| 2SD4 lgG4 |
8,4×10-3 |
4,20×105 |
2,00×10-8 |
0,8 |
| МАК 195 |
8,70×10-5 |
1,90×105 |
4,60×10-10 |
1,4 |
| F(ab’)2 |
|
|
|
|
Антитела D2E7 и близкие антитела также обладают высокой способностью к нейтрализации активности hTNF, что было определено в ряде анализов in vitro и in vivo (см. Пример 4). Например, эти антитела нейтрализуют hhTNF-индуцированную цитотоксичность клеток L929 со значениями IC50 в интервале от приблизительно 10-7 М до приблизительно 10-10 М. D2E7 при экспрессии в виде антитела lgG1 полной длины нейтрализует hTNF-индуцированную цитотоксичность клеток L929 с 1650 приблизительно 1,25х10-10 M. Более того, нейтрализующая активность D2E7 поддерживается, если антитело экспрессируется как Fab, F(ab’)2 или фрагмент scFv. D2E7 также ингибирует hTNF-индуцированную активацию клеток, что измеряют по hTNF-индуцированной экспрессии ELAM-1 на HUVEC (IC50=приблизительно 1,85×10-10 М) и по связыванию hTNF рецепторами hTNF на клетках U-937 (IC50=приблизительно 1,56×10-10 М). Относительно последнего случая D2E7 ингибирует связывание hTNF с обоими рецепторами hTNF – р55 и р75. Более того, антитело ингибирует hTNF-индуцированную гибель in vivo у мышей (ED50=1-2,5 мкг/мышь).
Что касается специфичности связывания D2E7, то это антитело связывает человеческий TNF в различных формах, включая растворимый hTNF, трансмембранный hTNF и hTNF, связанный с клеточными рецепторами. D2E7 не связывается специфически с другими цитокинами, такими как лимфокины (hTNF ), 1L(интерлейкин)-1, IL-1, IL-2, IL-4, IL-6, IL-8, IFN ), 1L(интерлейкин)-1, IL-1, IL-2, IL-4, IL-6, IL-8, IFN и TGF. Однако, D2E7 действительно обладает перекрестной реактивностью с факторами некроза опухоли из других видов. Например, антитело нейтрализует активность TNF по меньшей мере пяти приматов (шимпанзе, павиана, игрунки, циномолгуса и резуса) с приблизительно такими же значениями IC50, что и для нейтрализации активности мышиного TNF, хотя приблизительно в 1000 раз слабее, чем человеческого TNF (см. Пример 4, подраздел Е). D2E7 связывает также собачий и свиной hTNF. и TGF. Однако, D2E7 действительно обладает перекрестной реактивностью с факторами некроза опухоли из других видов. Например, антитело нейтрализует активность TNF по меньшей мере пяти приматов (шимпанзе, павиана, игрунки, циномолгуса и резуса) с приблизительно такими же значениями IC50, что и для нейтрализации активности мышиного TNF, хотя приблизительно в 1000 раз слабее, чем человеческого TNF (см. Пример 4, подраздел Е). D2E7 связывает также собачий и свиной hTNF.
В одном аспекте изобретение касается антител и фрагментов антител D2E7, антител и фрагментов антител, близких D2E7, и других человеческих антител и фрагментов антител с эквивалентными D2E7 свойствами, такими как высокая аффинность связывания hTNF при низкой кинетике диссоциации и высокой нейтрализующей активности. В одном варианте осуществления изобретение представляет выделенное человеческое антитело или его антиген-связывающий фрагмент, которые диссоциируют из человеческого TNF с Кd 1×10-8 М или менее и константе скорости диссоциации Кoff 1×10-3 сек-1 М или менее, измеренными с помощью поверхностного плазменного резонанса, и нейтрализуют цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50 1×10-7 М или менее. Более предпочтительно, если выделенное человеческое антитело или его антиген-связывающий фрагмент диссоциируют из человеческого TNF с Koff 5×10-4 сек-1 М или менее или даже более предпочтительно с Кoff 1×10-4 сек-1 М или менее. Более предпочтительно, если выделенное человеческое антитело или его антиген-связывающий фрагмент нейтрализуют цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50 1×10-8 М или менее, даже более предпочтительно с IC50 1×10-9 М или менее и еще более предпочтительно – IC50 5×10-10 М или менее. В предпочтительном варианте осуществления антитело является выделенным человеческим рекомбинантным антителом или его антиген-связывающим фрагментом. В другом предпочтительном варианте осуществления антитело также нейтрализует hhTNF-индуцированную активацию клеток, что оценивают стандартным анализом in vitro hTNF-индуцированной экспрессии ELAM-1 на эндотелиальных клетках пупочной вены человека (HUVEC).
Анализ с помощью поверхностного плазменного резонанса для определения Кd и Кoff может быть выполнен, как описано в Примере 1. Стандартный анализ in vitro с использованием L929 для определения значений IC50 описан в Примере 4, подразделе А. Стандартный анализ in vitro hTNF-индуцированной экспрессии ELAM-1 на эндотелиальных клетках пупочной вены человека (HUVEC) описан в Примере 4, подразделе А. Примеры рекомбинантных человеческих антител, которые соответствуют или, как предполагают, соответствуют вышеописанным критериям кинетики и нейтрализации, включают антитела, имеющие следующие [VH/VL] пары, последовательности которых представлены на Фигурах 1, 2, 3 и 4 (см. также анализы кинетики и нейтрализации в Примерах 2, 3 и 4): [D2E7 VH/D2E7 VL]; [HD2E7*.A1/D2E7 VL]; [HD2E7*.A2/D2E7 VL]; [HD2E7*.A3/D2E7 VL]; [HD2E7*.A4/D2E7 VL]; [HD2E7*.A5/D2E7 VL]; [HD2E7*.A6/D2E7 VL]; [HD2E7*.A7/D2E7 VL]; [HD2E7*.A8/D2E7 VL]; [HD2E7*.A9/D2E7 VL]; [D2E7 VH/LD2E7*.A1]; [D2E7 VH/LD2E7*.A4]; [D2E7 VH/LD2E7*.A5]; [D2E7 VH/LD2E7*.A7]; [D2E7VH/LD2E7*.A8]; [HD2E7*.A9/LD2E7*.A1]; [VH1-D2/LOE7]; [VH1-D2.N/LOE7.T]; [VH1-D2.Y/LOE7.A]; [VH1-D2.N/LOE7.A]; [VH1-D2/EP B12] и [3C-H2/LOE7].
Из уровня техники хорошо известно, что домены CDR3 тяжелой и легкой цепей антитела играют важную роль в специфичности/аффинности при связывании антитела с антигеном. Соответственно в другом аспекте изобретение касается человеческих антител, которые имеют низкую кинетику диссоциации для ассоциации с hTNF и которые имеют домены CDR3 легкой и тяжелой цепей, являющиеся структурно идентичными или близкими таковым D2E7. Как показано в Примере 3, положение 9 D2E7 VL CDR3 может быть занято Ala или Thr без существенного влияния на Кoff. Соответственно консенсусный участок D2E7 VL CDR3 содержит аминокислотную последовательность Q-R-Y-N-R-A-P-Y-(T/A) (SEQ ID No 3). Кроме того, положение 12 D2E7 VH CDR3 может быть занято Tyr или Asn без существенного влияния на Кoff. Соответственно консенсусный участок D2E7 VHCDRS содержит аминокислотную последовательность V-S-Y-L-S-T-A-S-S-L-D-(Y/N) (SEQ ID No 4). Более того, как продемонстрировано в Примере 2, в домене CDR3 тяжелой и легкой цепей D2E7 легко провести замену одним остатком аланина (в положении 1, 4, 5, 7 или 8 в VL CDR3 или в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11 в VH CDR3) без существенного влияния на Кoff. Более того, специалист оценит, что при возможности легкой замены аланином в доменах D2E7 VL и VH CDR3 возможны замены другими аминокислотами в доменах CDR3 при сохранении низкого значения константы скорости диссоциации антитела, в частности, замены консервативных аминокислот. Под “заменой консервативной аминокислоты” в контексте заявки подразумевается замена, при которой остаток аминокислоты замещается другим остатком аминокислоты, имеющим близкую боковую цепь. Семейства остатков аминокислот, имеющих близкие боковые цепи, были определены в уровне техники, в том числе основные боковые цепи (например, лизин, аргинин, гистидин), кислые боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), -разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Предпочтительно в доменах D2E7 VL и/или VH CDR3 делают не более чем от одной до пяти замен консервативных аминокислот. Более предпочтительно в доменах D2E7 VL и/или VH CDR3 делают не более чем от одной до трех замен консервативных аминокислот. Кроме того, замены консервативных аминокислот не следует делать в положениях аминокислот, критических для связывания DTNF. Как показано в Примере 3, положения 2 и 5 D2E7 VL CDR3 и положения 1 и 7 D2E7 VH CDR3 оказались критическими для взаимодействия с hhTNF и, таким образом, предпочтительно не делать замен консервативных аминокислот в этих положениях (хотя замена аланином в положении 5 D2E7 VL CDR3 является приемлемой, как описано выше).
Соответственно в другом варианте осуществления изобретения используется выделенное человеческое антитело или его антиген-связывающий фрагмент со следующими характеристиками:
a) диссоциирует из ассоциации (комплекса) с hTNF с константой скорости Кoff 1×10-3 с-1 или менее, что определено с помощью поверхностного плазменного резонанса;
b) имеет домен CDR3 легкой цепи, содержащий аминокислотную последовательность SEQ ID No 3 или SEQ ID No 3, модифицированную посредством замены одним аланином в положении 1, 4, 5, 7 или 8 или заменой от одной до пяти консервативных аминокислот в положениях 1,3,4, 6, 7, 8 и/или 9;
c) имеет домен CDR3 тяжелой цепи, содержащий аминокислотную последовательность SEQ ID No 4 или SEQ ID No 4, модифицированную посредством замены одним аланином в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11 или заменой от одной до пяти консервативных аминокислот в положениях 2, 3, 4, 5, 6, 8, 9, 10, 11 и/или 12.
Более предпочтительно, если антитело или его антиген-связывающий фрагмент диссоциируют из человеческого hhTNF с Кoff 5×10-4 с-1 или менее.
Еще более предпочтительно, если антитело или его антиген-связывающий фрагмент диссоциируют из человеческого TNF с Кoff 1×10-4 с-1 или менее.
В еще одном варианте осуществления изобретения используется выделенное человеческое антитело или его антиген-связывающий фрагмент с вариабельным участком легкой цепи (LCVR), имеющим домен CDR3, содержащий аминокислотную последовательность SEQ ID No 3 или SEQ ID No 3. модифицированную посредством замены одним аланином в положении 1, 4, 5, 7 или 8 или с вариабельным участком тяжелой цепи (HCVR), имеющим домен CDR3, содержащий аминокислотную последовательность SEQ ID No 4 или SEQ ID No 4, модифицированную посредством замены одним аланином в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11. Предпочтительно, если LCVR далее имеет домен CDR2, содержащий аминокислотную последовательность SEQ ID No 5 (т.е. D2E7 VLCDR2), и HCVR далее имеет домен CDR2, содержащий аминокислотную последовательность SEQ ID No 6 (т.е. D2E7 VH CDR2). Еще более предпочтительно, если LCVR далее имеет домен CDR1, содержащий аминокислотную последовательность SEQ ID No 7 (т.е. D2E7 VL CDR1) и HCVR далее имеет домен CDR1, содержащий аминокислотную последовательность SEQ ID No 8 (т.е. D2E7 VH CDR1). Предпочтительны скелетные участки VL из семейства зародышевой линии человека VkI, более предпочтительны – из гена Vk зародышевой линии человека А20 и наиболее предпочтительны – из скелетных последовательностей D2E7 VL, представленных на Фигурах 1 и 2. Предпочтительны скелетные участки VH из семейства зародышевой линии человека VH3, более предпочтительны – из гена VH зародышевой линии человека DP-31 и наиболее предпочтительны – из скелетных последовательностей D2E7 VH, представленных на Фигурах 3 и 4.
В еще одном варианте осуществления изобретения используется выделенное человеческое антитело или его антиген-связывающий фрагмент с вариабельным участком легкой цепи (LCVR), содержащим аминокислотную последовательность SEQ ID No 1 (т.е. D2E7 VL) и вариабельным участком тяжелой цепи (HCVR), содержащим аминокислотную последовательность SEQ ID No 2 (т.е. D2E7 VH). В некоторых вариантах осуществления антитело содержит константный участок тяжелой цепи, такой как константный участок lgG1, lgG2, lgG3, lgG4, IgA, IgE, IgM, IgD. Предпочтительно константным участком тяжелой цепи является константный участок тяжелой цепи lgG1 или константный участок тяжелой цепи lgG4. Более того, антитело может содержать как константный участок легкой цепи либо каппа-константный участок легкой цепи, либо лямбда-константный участок легкой цепи. Предпочтительно антитело содержит каппа-константный участок легкой цепи. С другой стороны, фрагментом антитела может быть, например, фрагмент Fab или фрагмент Fv из одной цепи.
Выделенное человеческое антитело или его антиген-связывающие фрагменты может иметь домены CDR3 VL и VH, близкие D2E7, например, антитела или их антиген-связывающие фрагменты с вариабельным участком легкой цепи (LCVR), имеющим домен CDR3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No 3, SEQ ID No 11, SEQ ID No 12, SEQ ID No 13, SEQ ID No 14, SEQ ID No 15, SEQ ID No 16, SEQ ID No 17, SEQ ID No 18, SEQ ID No 19, SEQ ID No 20, SEQ ID No 21, SEQ ID No 22, SEQ ID No 23, SEQ ID No 24, SEQ ID No 25, SEQ ID No 26 или с вариабельным участком тяжелой цепи (HCVR), имеющим домен CDR3, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No 4, SEQ ID No 27, SEQ ID No 28, SEQ ID No 29, SEQ ID No 30, SEQ ID No 31, SEQ ID No 32, SEQ ID No 33, SEQ ID No 34 и SEQ ID No 35.
В качестве еще одного примера осуществления изобретения может быть рассмотрено рекомбинантное человеческое антитело или его антиген-связывающий фрагмент, которые нейтрализуют активность человеческого TNF, но не человеческого TNF. Предпочтительно, если антитело или его антиген-связывающий фрагмент также нейтрализуют активность hTNF шимпанзе и по меньшей мере одного дополнительного hTNF приматов, выбранного из группы, состоящей из hTNF павиана, TNF игрунки, hTNF циномолгуса и TNF резуса. Предпочтительно, если антитело или его антиген-связывающий фрагмент нейтрализуют TNF человека, шимпанзе и/или дополнительного примата в стандартном анализе in vitro с использованием L929 с IC50 1×10-8 М или менее, более предпочтительно 1×10-9 М, еще более предпочтительно 5×10-10 М или менее. В одном субварианте осуществления антитело нейтрализует также активность собачьего TNF, предпочтительно в стандартном анализе in vitro с использованием L929 с IC50 1×10-7 М или менее, более предпочтительно 1×10-8 М или менее и даже более предпочтительно 5×10-9 М или менее. В другом субварианте осуществления антитело нейтрализует также активность свиного TNF, предпочтительно с 1050 1×10-5 М или менее, более предпочтительно 1×10-6 М или менее и даже более предпочтительно 5×10-7 М или менее. В еще одном субварианте осуществления антитело нейтрализует также активность мышиного TNF предпочтительно с IC50 1×10-4 М или менее, более предпочтительно 1×10-5 М или менее и даже более предпочтительно 5×10-6 М или менее.
Антитело или фрагмент антитела, соответствующие изобретению, могут образовывать производные или быть связаны с другой активной молекулой (например, другим пептидом или белком). Соответственно антитела или фрагменты антител, соответствующие изобретению, предусматривают включение производных или иным образом модифицированных форм человеческих антител против hTNF, описанных здесь, включая молекулы иммуноадгезии. Например, антитело или фрагмент антитела, соответствующие изобретению, могут быть функционально связаны (химическим соединением, генетическим слиянием, нековалентной ассоциацией или иным способом) с одной или более молекулярных структур, таких как другое антитело (например, биспецифическое антитело или диатело), обнаруживаемый агент (метка), цитотоксический агент, фармацевтический агент и/или белок или пептид, которые могут влиять на ассоциацию антитела или фрагмента антитела с другой молекулой (такой как участок ядра стрептавидина или полигистидиновой метки).
Один тип производного антитела получают перекрестным сшиванием двух или более антител (одного и того же типа или различных типов, например, для создания биспецифичных антител). Подходящие перекрестно сшивающие агенты включают агенты, которые являются гетеробифункциональными, имеющими две различные реактивные группы, разделенные подходящим спейсером (например, m-малеимидобензоил-N-гидроксисукцинимидный эфир) или гомобифункциональными (например, дисукцинимидил суберат). Такие линкеры предлагаются Pierce Chemical Company, Rockford, IL.
Подходящие обнаруживаемыми агентами, с помощью которых могут быть образованы производные антитела или фрагмента антитела, соответствующие изобретению, включают флуоресцентные соединения. Примеры флуоресцентных меток включают флуоресцеин, изотиоцианат флуоресцеина, родамин, хлорид 5-диметиламин-1-нафталинсульфонила, фикоэритрин и т.п. Антитело также может быть дериватизировано обнаруживаемыми ферментами, такими как щелочная фосфатаза, пероксидаза хрена, глюкозооксидаза и т.п. Когда антитело образует производное с обнаруживаемым ферментом, его выявляют добавлением дополнительных реагентов, которые фермент использует с образованием выявляемого продукта реакции. Например, когда присутствует обнаруживаемый агент пероксидаза хрена, добавление пероксида водорода и диаминобензидина приводит к образованию окрашенного продукта реакции, который является определяемым. Антитело может также образовать производное с биотином и быть выявленным посредством непрямого измерения связывания авидина или стрептавидина.
II. Экспрессия антител
Антитело или фрагмент антитела, соответствующие изобретению, могут быть получены рекомбинантной экспрессией генов легкой и тяжелой цепей иммуноглобулина в клетке-хозяине. Для рекомбинантной экспрессии антитела клетку-хозяин трансфицируют одним или более рекомбинантных экспрессионных векторов, несущих фрагменты ДНК, кодирующие легкую и тяжелую цепи иммуноглобулина антитела таким образом, что легкая и тяжелая цепи экспрессируются в клетке-хозяине, и предпочтительно выделяются в среду, в которой культивируют клетки-хозяина, из этой среды могут быть выделены антитела. Стандартные способы с применением рекомбинантной ДНК используются для получения генов тяжелой и легкой цепей, инкорпорации этих генов в рекомбинантные экспрессионные векторы и интродукции векторов в клетки-хозяина так, как описано Sambrook, Fritsch and Maniatis (eds) Molecular Cloning; A Laboratory Manual, Second Edition, Cold Spring Harbor, N.Y. (1989; Ausubel F.M. et al. (eds.) Current Protocols in Molecular Biology, Green Publishing Associates (1989) и Патент США No 4816397 Boss et al.
Для экспрессии D2E7 или родственного D2E7 антитела сначала получают фрагменты ДНК, кодирующие легкую и тяжелую цепи вариабельных участков. Эти ДНК могут быть получены амплификацией и модификацией последовательностей легкой и тяжелой цепей зародышевой линии с использованием полимеразной цепной реакции (ПЦР). Последовательности ДНК зародышевой линии генов вариабельных участков тяжелой и легкой цепей человека известны из уровня техники (см., например, “Vbase” – базу данных последовательностей зародышевой линии человека; см. также Kabat Е.А et al.(1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No 91-3242; Tomlinson I.M. et al. (1992) “The Repertoire of Human Germline VH Sequences Reveals about Fifty Groups of VH Segments with Different Hypervariable Loops” J. Mol. Biol. 227:776-798; Cox J.P.L et al.(1994) “A Directory of Human Germ-line VH Segments Reveals a Strong Bias in their Usage” Eur. J. Immunol. 24:827-836; содержание каждого из которых специально введено здесь в виде ссылки). Для получения фрагмента ДНК, кодирующего вариабельный участок тяжелой цепи D2E7 или близкого D2E7 антитела, член семейства VH3 генов VH зародышевой линии человека амплифицируют с помощью стандартной ПЦР. Наиболее предпочтительной является амплификация последовательности зародышевой линии VH DP-31. Для получения фрагмента ДНК, кодирующего вариабельный участок легкой цепи D2E7 или близкого D2E7 антитела, член семейства VkI генов VL зародышевой линии человека амплифицируют стандартной ПЦР. Наиболее предпочтительной является амплификация последовательности зародышевой линии VLA20. ПЦР-праймеры, пригодные для использования при амплификации последовательностей зародышевой линии VH DP-31 и зародышевой линии VL A20, могут быть определены на основе нуклеотидных последовательностей, приведенных в ссылках, цитированных supra, с использованием стандартных способов.
После получения фрагментов VH и VL зародышевой линии эти последовательности могут быть подвергнуты мутагенезу, чтобы они кодировали описанные здесь аминокислотные последовательности D2E7 или близкие D2E7. Аминокислотные последовательности, кодируемые последовательностями ДНК зародышевых линий VH и VL, сначала сравнивают с аминокислотными последовательностями VH и VL D2E7 или близкими D2E7 для идентификации остатков аминокислот в последовательности D2E7 или близкой D2E7, которые отличаются от зародышевой линии. Затем соответствующие нуклеотиды последовательностей ДНК зародышевой линии подвергают мутагенезу таким образом, что мутантная последовательность зародышевой линии кодирует последовательность аминокислот D2E7 или близкую D2E7, используя при этом генетический код для того, чтобы установить, какие изменения нуклеотидов следует произвести. Мутагенез последовательностей зародышевой линии проводят стандартными способами, такими как ПЦР-обусловленный мутагенез (в котором мутантные нуклеотиды вводятся в ПЦР-праймеры таким образом, что продукт ПЦР содержит мутации) или сайт-направленный мутагенез.
Более того, следует отметить, что, если последовательности “зародышевой линии”, полученные ПЦР-амплификацией, кодируют аминокислотные отличия скелетных участков от правильной конфигурации зародышевой линии (т.е. отличия в амплифицированной последовательности по сравнению с правильной последовательностью зародышевой линии, например, как результат соматической мутации), то может быть желательным изменить эти отличия аминокислот в обратном направлении – к правильным последовательностям зародышевой линии (т.е. “обратная мутация” скелетных остатков в конфигурации зародышевой линии).
После получения фрагментов ДНК, кодирующих сегменты VH и VL D2E7 или близких D2E7 (путем амплификации и мутагенеза генов VH и VL зародышевой линии, как описано выше), могут быть проведены дальнейшие манипуляции с этими фрагментами ДНК стандартными способами с использованием рекомбинантной ДНК, например превращение генов вариабельного участка в гены цепи антитела полной длины, в гены фрагмента Fab или в ген scFv. В этих манипуляциях фрагмент ДНК, кодирующий VL или VH, оперативно связан с другим фрагментом ДНК, кодирующим другой белок, такой как константный участок антитела или гибкий линкер. Термин “оперативно связан”, используемый в данном контексте, предназначен для обозначения того, что два фрагмента ДНК связаны так, что аминокислотные последовательности, кодируемые двумя фрагментами ДНК, остаются в рамке считывания.
Выделенная ДНК, кодирующая участок VH, может быть превращена в ген тяжелой цепи полной длины посредством оперативного связывания VH-кодирующей ДНК с другой молекулой ДНК, кодирующей константные участки тяжелой цепи (СН1, СН2 и СН3). Последовательности генов константных участков тяжелой цепи человека известны из уровня техники (см., например, Kabat E.A et al.(1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No 91-3242), а фрагменты ДНК, содержащие эти участки, могут быть получены стандартной ПЦР-амплификацией. Константный участок тяжелой цепи может быть константным участком lgG1, lgG2, lgG3, lgG4, IgA, IgE, IgM, IgD, но наиболее предпочтительным является константный участк lgG1 или lgG4. Для гена фрагмента Fab тяжелой цепи VH-кодирующая ДНК может быть оперативно связана с другой молекулой ДНК, кодирующей только константный участок СН1 тяжелой цепи.
Выделенная ДНК, кодирующая участок VL, может быть превращена в ген легкой цепи полной длины (так же, как ген легкой цепи Fab) посредством оперативного связывания VL-кодирующей ДНК с другой молекулой ДНК, кодирующей константный участок легкой цепи CL. Последовательности генов константных участков легкой цепи человека известны из уровня техники (см., например, Kabat E.A. et al.(1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No 91-3242) и фрагменты ДНК, содержащие эти участки, могут быть получены стандартной ПЦР-амплификацией. Константный участок легкой цепи может быть константным участком каппа- или лямбда-, но наиболее предпочтительно является константным участком каппа-.
Для получения гена scFv VH- и VL-кодирующие фрагменты ДНК оперативно связывают с другим фрагментом, кодирующим гибкий линкер, например, кодирующим последовательность аминокислот (Gly4-Ser)3, таким образом, что последовательности VL и VH могут быть экспрессированы в виде прилегающего белка из одной нити, в котором участки VL и VH связаны гибким линкером (см., например, Bird et al. (1988) Science 242:423-426; Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883; McCafferty et al., Nature (1990) 348:552-554).
Для экспрессии антител или фрагментов антител, соответствующих изобретению, кодирующие легкие и тяжелые цепи полной длины или их фрагменты ДНК, полученные, как описано выше, вводят в экспрессионные векторы таким образом, что гены становятся оперативно связанными с последовательностями контроля транскрипции и трансляции. В данном контексте термин “оперативно связанные” предназначен для обозначения того, что ген антитела лигируют в вектор так, что последовательности контроля транскрипции и трансляции в векторе выполняют предназначенную им функцию регуляции транскрипции и трансляции гена антитела. Экспрессионный вектор и последовательности контроля экспрессии выбирают так, что они совместимы с используемой экспрессирующей клеткой-хозяином. Ген легкой цепи антитела и ген тяжелой цепи антитела могут быть введены в раздельные векторы или, что более типично, оба гена вводят в один и тот же экспрессионный вектор. Ген антитела вводят в экспрессионный вектор стандартными способами (например, лигированием комплементарных рестрикционных сайтов на фрагменте гена антитела и вектора или лигированием тупых концов, если рестрикционные сайты отсутствуют). Перед введением последовательностей легкой и тяжелой цепей D2E7 или близких D2E7 экспрессионный вектор может уже нести последовательности константных участков антитела. Например, одним из подходов к превращению последовательностей VH и VL D2E7 или близких D2E7 в гены антитела полной длины является их инсерция в экспрессионный вектор, уже кодирующий константный участок тяжелой цепи и константный участок легкой цепи соответственно таким образом, что сегмент VH оперативно связан с сегментом(ами) СН в векторе, а сегмент VL оперативно связан с сегментом CL в векторе. Дополнительно или альтернативно рекомбинантный экспрессионный вектор может кодировать сигнальный пептид, который облегчает выделение цепи антитела из клетки-хозяина. Ген цепи антитела может быть клонирован в вектор таким образом, что сигнальный пептид связан в рамке считывания с геном амино-конца цепи антитела. Сигнальный пептид может быть сигнальным пептидом иммуноглобулина или гетерологичным сигнальным пептидом (т.е. сигнальным пептидом из белка неиммуноглобулина).
Дополнительно к генам цепи антитела рекомбинантные экспрессирующие векторы, соответствующие изобретению, несут регуляторные последовательности, которые контролируют экспрессию генов цепи антитела в клетке-хозяине. Термин “регуляторная последовательность” предусматривает включение промоторов, энхансеров и других элементов контроля экспрессии (например, сигналов полиаденилирования), которые контролируют транскрипцию или трансляцию генов цепи антитела. Такие регуляторные последовательности описаны, например, см. Goeddel Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Любой специалист оценит, что конструирование экспрессионного вектора, включая выбор регуляторных последовательностей, может зависеть от таких факторов, как выбор клетки-хозяина для трансформации, желательный уровень экспрессии белка и т.д. Предпочтительные регуляторные последовательности для экспрессии в клетке-хозяине млекопитающих включают вирусные элементы, которые направляют экспрессию белка в клетках млекопитающих на высоком уровне, такие как промоторы и/или энхансеры, выделенные из цитомегаловируса (ЦМВ) (такие как промотор/энхансер ЦМВ), вирус Simian 40 (SV40) (такие как промотор/энхансер SV40), аденовирус (например, аденовирусный главный поздний промотор (AdMLP)) и полиома. Для дальнейшего описания вирусных регуляторных элементов и их последовательностей см., например, Патент США No 5168062 Stinski; Патент США No 4510245 Bell et al. и Патент США No 4968615 Schaffner et al.
Дополнительно к генам цепи антитела и регуляторным последовательностям рекомбинантные экспрессирующие векторы, соответствующие изобретению, могут нести дополнительные последовательности, такие как последовательности, которые регулируют репликацию вектора в клетках хозяина (например, ориджины репликации) и гены селектируемых маркеров. Ген селектируемого маркера облегчает селекцию клеток-хозяев, в которые был интродуцирован вектор (см., например, Патенты США NoNo 4399216, 4634665 и 5179017 все Axel et al.). Например, в типичном случае ген селектируемого маркера обусловливает устойчивость к лекарственным веществам, таким как G418, гигромицин или метотрексат, клетки-хозяина, в которую был инродуцирован вектор. Предпочтительные гены селектируемых маркеров включают ген дигидрофолатредуктазы (DHFR) (для использования клеток-хозяев dhfr– с селекцией/амплификацией на фоне метотрексата) и ген neo (для селекции на фоне G418).
Для экспрессии легкой и тяжелой цепей экспрессионным вектором(ами), кодирующим тяжелую и легкую цепи, трансфицируют клетку-хозяин стандартными способами. Различные трактовки термина “трансфекция” преднаначены для включения широкого ряда способов, обычно используемых при интродукции экзогенной ДНК в прокариотную или эукариотную клетку-хозяин, например, электропорацию, преципитацию фосфатом кальция, трансфекцию с помощью DEAE-декстрана и др. Хотя теоретически является возможным экспрессировать антитела, соответствующие изобретению, либо в прокариотных, либо в эукариотных клетках-хозяевах, экспрессия антител в эукариотных клетках, и наиболее предпочтительно, в клетках-хозяевах млекопитающих, является наиболее предпочтительной, поскольку такие эукариотные клетки и особенно клетки млекопитающих, более вероятно, чем прокариотные клетки собирают и секретируют правильно образованное и иммунологически активное антитело. Прокариотная экспрессия генов антител, как было описано, является неэффективной для получения активных антител с высоким выходом (Boss М.А. and Wood C.R. (1985) Immunology Today 6:12-13).
Предпочтительные клетки-хозяева млекопитающих для экспрессии рекомбинантных антител, соответствующих изобретению, включают клетки яичников китайского хомячка (клетки СНО) (включая клетки СНО dhfr–, описанные Uriaub and Chasin (1980) Proc. Natl. Acad. Sci. USA 77: 4216-4220, используемые с селективным маркером DHFR, например, как описано R.J.Kaufman and P.A.Sharp (1982) Mol. Biol. 159: 601-621), клетки миеломы NSO, клетки COS и клетки SP2. Когда рекомбинантные экспрессирующие векторы, кодирующие гены антитела, интродуцируют в клетки-хозяева млекопитающих, антитела образуются при культивировании клеток-хозяев в течение периода времени, который достаточен для экспрессии антитела в клетке-хозяине, или (более предпочтительно) секреции антитела в культуральную среду, в которой выращивают клетки-хозяина. Антитела могут быть выделены из культуральной среды с использованием стандартных способов очистки белков.
Клетки-хозяина также могут быть использованы для получения фрагментов интактных антител, таких как фрагменты Fab и молекулы scFv. Следует понимать, что варианты вышеописанной процедуры входят в объем изобретения. Например, может быть желательно трансфицировать клетку-хозяин ДНК, кодирующей либо легкую цепь, либо тяжелую цепь (но не обе) антитела, соответствующего изобретению. Способ с использованием рекомбинатной ДНК может также быть использован для удаления некоторых или всех ДНК, кодирующих отдельно или обе легкую и тяжелую цепи, которые не являются необходимыми для связывания hTNF. Молекулы, экспрессируемые с таких усеченных молекул ДНК, также охватываются антителами, соответствующими изобретению. Кроме того, перекрестным сшиванием антитела, соответствующего изобретению, со вторым антителом с помощью стандартных химических способов перекрестного сшивания также могут быть получены бифункциональные антитела, в которых одна тяжелая и одна легкая цепь являются антителом, соответствующими изобретению, а другая тяжелая и другая легкая цепь специфичны к антигену, отличному от hTNF.
В предпочтительной системе рекомбинантной экспрессии антитела или его антиген-связывающего фрагмента, соответствующих изобретению, рекомбинатный экспрессионный вектор, кодирующий обе тяжелую цепь антитела и легкую цепь антитела, интродуцируют в клетки СНО dhfr– посредством трансфекции с помощью фосфата кальция. В рекомбинатном экспрессионном векторе каждый из генов тяжелой и легкой цепей антитела оперативно связан с регуляторными элементами энхансера/промотора (например, выделенными из SV40, ЦМВ, аденовируса и т.п., такими как регуляторный элемент энхансера ЦМВ/промотора AdMLP или регуляторный элемент энхансера SV40/промотора AdMLP) для того, чтобы направлять транскрипцию генов на высоком уровне. Рекомбинантный экспрессионный вектор несет также ген DHFR, который обеспечивает селекцию клеток СНО, которые были трансфицированы вектором с использованием селекции/амплификации на фоне метотрексата. Отобранные клетки-хозяева-трансформанты являются культурой, обеспечивающей экспрессию тяжелой и легкой цепей антитела, и интактное антитело выделяют из культуральной среды. Для получения рекомбинантного экспрессионного вектора, трансфекции клеток-хозяев, селекции трансформантов, культивирования клеток-хозяев и выделения антитела из культуральной среды используют стандартные молекулярно-биологические способы.
В свете вышеописанного изобретение также касается композиций нуклеиновых кислот, векторов и клеток-хозяев, которые могут быть использованы для рекомбинантной экспрессии антител и фрагментов антител, соответствующих изобретению. Нуклеотидная последовательность, кодирующая вариабельный участоклегкой цепи D2E7, представлена на Фигуре 9 и SEQ ID No 36. Домен CDR1 LCVR охватывает нуклеотиды 70-102, домен CDR2 охватывает нуклеотиды 148-168 и домен CDR3 охватывает нуклеотиды 265-291. Нуклеотидная последовательность, кодирующая вариабельный участок тяжелой цепи D2E7, представлена на Фигуре 10 и SEQ ID No 37. Домен CDR1 HCVR охватывает нуклеотиды 91-105, домен CDR2 охватывает нуклеотиды 148-198 и домен CDR3 охватывает нуклеотиды 295-330. Специалист оценит, что нуклеотидные последовательности, кодирующие антитела, близкие D2E7, или их фрагменты (например, домен CDR, такой как домен CDR3) могут быть выделены из нуклеотидных последовательностей, кодирующих LCVR и HCVR D2E7 с использованием генетического кода и стандартных молекулярно-биологических способов.
В одном варианте осуществления изобретения используется выделенная нуклеиновая кислота, кодирующая домен CDR3 легкой цепи, с аминокислотной последовательностью SEQ ID No 3 (т.е. D2E7 VL CDR3) или SEQ ID No 3, модифицированной посредством замены одним остатком аланина в положении 1, 4, 5, 7 или 8 или посредством замены от одной до пяти консервативных аминокислот в положениях 1, 3, 4, 6, 7, 8 и/или 9. Эта нуклеиновая кислота может кодировать только участок CDR3 или, более предпочтительно, кодирует вариабельный участок легкой цепи целого антитела (LCVR). Например, нуклеиновая кислота может кодировать LCVR, имеющий домен CDR2, содержащий аминокислотную последовательность SEQ ID No 5 (т.е. D2E7 VL CDR2) и домен CDR1, содержащий аминокислотную последовательность SEQ ID No 7 (т.е. D2E7 VL CDR1).
В еще одном варианте осуществления изобретении используется выделенная нуклеиновая кислота, кодирующая домен CDR3 тяжелой цепи, с аминокислотной последовательнсотью SEQ ID No 4 (т.е. D2E7 VH CDR3) или SEQ ID No 4, модифицированной посредством замены одного остатка аланина в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11 или посредством замены от одной до пяти консервативных аминокислот в положениях 2, 3, 4, 5, 6, 8, 9, 10, 11 и/или 12. Эта нуклеиновая кислота может кодировать только участок CDR3 или, более предпочтительно, кодирует вариабельный участок тяжелой цепи целого антитела (HCVR). Например, нуклеиновая кислота может кодировать HCVR, имеющий домен CDR2, содержащий аминокислотную последовательность SEQ ID No 6 (т.е. D2E7 VH CDR2) и домен CDR1, содержащий аминокислотную последовательность SEQ ID No 8 (т.е. D2E7 VH CDR1).
В изобретении могут также использоваться нуклеиновые кислоты, кодирующие домен CDR3, близкий D2E7, например, содержащий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID No 3, SEQ ID No 4, SEQ ID No 11, SEQ ID No 12, SEQ ID No 13, SEQ ID No 14, SEQ ID No 15, SEQ ID No 16, SEQ ID No 17, SEQ ID No 18, SEQ ID No 19, SEQ ID No 20, SEQ ID No 21, SEQ ID No 22, SEQ ID No 23, SEQ ID No 24, SEQ ID No 25, SEQ ID No 26, SEQ ID No 27, SEQ ID No 28, SEQ ID No 29, SEQ ID No 30, SEQ ID No 31, SEQ ID No 32, SEQ ID No 33, SEQ ID No 34 и SEQ ID No 35.
В еще одном варианте осуществления изобретения используется выделенная нуклеиновая кислота, кодирующая вариабельный участок легкой цепи антитела, содержащий аминокислотную последовательность SEQ ID No 1 (т.е. LCVR D2E7). Предпочтительно данная нуклеиновая кислота содержит нуклеотидную последовательность SEQ ID No 36, хотя специалист оценит, что вследствие дегенерации генетического кода другие нуклеотидные последовательности могут кодировать аминокислотную последовательность SEQ ID No 1. Нуклеиновая кислота может кодировать только LCVR или может также кодировать константный участок легкой цепи антитела, оперативно связанный с LCVR. В одном варианте осуществления данная нуклеиновая кислота находится в рекомбинантном экспрессионном векторе.
В еще одном варианте осущствления изобретения используется выделенная нуклеиновая кислота, кодирующая вариабельный участок тяжелой цепи антитела, содержащий аминокислотную последовательность SEQ ID No 2 (т.е. HCVR D2E7). Предпочтительно данная нуклеиновая кислота содержит нуклеотидную последовательность SEQ ID No 37, хотя специалист оценит, что вследствие дегенерации генетического кода другие нуклеотидные последовательности могут кодировать аминокислотную последовательность SEQ ID No 2. Нуклеиновая кислота может кодировать только HCVR или может также кодировать константный участок тяжелой цепи антитела, оперативно связанный с HCVR. Например, нуклеиновая кислота может содержать константный участок lgG1 или lgG4. В одном варианте осуществления данная нуклеиновая кислота находится в рекомбинантном экспрессирующем векторе.
Также могут использоваться рекомбинантные экспрессирующие векторы, кодирующие обе тяжелую цепь антитела и легкую цепь антитела. Например, рекомбинантный экспрессионный вектор, кодирующий:
a) легкую цепь антитела, имеющую вариабельный участок, с аминокислотной последовательностью SEQ ID No 1 (т.е. LCVR D2E7), и
b) тяжелую цепь антитела, имеющую вариабельный участок, с аминокислотной последовательностью SEQ ID No 2 (т.е. HCVR D2E7).
Один или более рекомбинантынй экспрессионный вектор, соответствующий изобретению, может быть интродуцирован в клетки-хозяина. Предпочтительно клетка-хозяин является клеткой-хозяином млекопитающего, более предпочтительно клетка-хозяин является клеткой СНО, клеткой NSO или клеткой COS.
Синтез рекомбинантного человеческого антитела, соответствующего изобретению, может быть произведен посредством культивирования клетки-хозяина, соответствующей изобретению, в подходящей среде культивирования до того времени, пока не синтезируется рекомбинантное человеческое антитело, соответствующее изобретению. Способ может далее предусматривать выделение рекомбинантного человеческого антитела из культуральной среды.
III. Селекция рекомбинантных человеческих антител
Рекомбинантные человеческие антитела, соответствующие изобретению, дополнительно к описанным здесь антителам D2E7 или близким D2E7 могут быть выбраны просмотром рекомбинантной комбинаторной библиотеки антител, предпочтительно библиотеки scFv отображающей фаги, полученных с использованием кДНК человеческих VL и VH, полученных с мРНК, выделенной из лимфоцитов человека. Методологии получения и скрининга таких библиотек известны из уровня техники. В дополнение к имеющимся в продаже наборам для получения библиотек с демонстрацией фагов (например, Pharmacia Recombinant Phage Antibody System, каталог No 27-9400-01; набор демонстрации фагов Stratagene SurfZAP , каталог No 240612), примеры способов и реагентов, особенно пригодных для применения при создании и скрининге библиотек, отображающих антитела, могут быть обнаружены, например, Ladner et al. Патент США No 5223409; Kang et al. Публикация PCT No WO 92/18619; Dower et al. Публикация РСТ No WO 91/17271; Winter et al. Публикация PCT No WO 92/20791; Markland et al. Публикация PCT No WO 92/15679; Breitling et al. Публикация PCT No WO 93/01288; McCafferty et al. Публикация РСТ No WO 92/01047; Garrard et al. Публикация РСТ No WO 92/09690; Fuchs et al. (1991) Bio/Technology 9: 1370-1372; Hay et al. (1992) Hum. Antibod. Hybridomas 3: 81-85; Huse et al. (1989) Science 246: 1275-1281; McCafferty et al., Nature (1990) 348: 552-554; Griffiths et al. (1993) EMBO J. 12: 725-734; Hawkins et al. (1992) J. Mol. Biol. 226: 889-896; CClckson et al. (1991) Nature 352: 624-628; Gram et al. (1992) PNAS 89:3576-33580; Garrad et al. (1991) Bio/Technology 9: 1373-1377; Hoogenboom et al. ((1991) Nuc. Acid Res. 19: 4133-4137; Barbas et al. (1991) PNAS 88:7978-7982. , каталог No 240612), примеры способов и реагентов, особенно пригодных для применения при создании и скрининге библиотек, отображающих антитела, могут быть обнаружены, например, Ladner et al. Патент США No 5223409; Kang et al. Публикация PCT No WO 92/18619; Dower et al. Публикация РСТ No WO 91/17271; Winter et al. Публикация PCT No WO 92/20791; Markland et al. Публикация PCT No WO 92/15679; Breitling et al. Публикация PCT No WO 93/01288; McCafferty et al. Публикация РСТ No WO 92/01047; Garrard et al. Публикация РСТ No WO 92/09690; Fuchs et al. (1991) Bio/Technology 9: 1370-1372; Hay et al. (1992) Hum. Antibod. Hybridomas 3: 81-85; Huse et al. (1989) Science 246: 1275-1281; McCafferty et al., Nature (1990) 348: 552-554; Griffiths et al. (1993) EMBO J. 12: 725-734; Hawkins et al. (1992) J. Mol. Biol. 226: 889-896; CClckson et al. (1991) Nature 352: 624-628; Gram et al. (1992) PNAS 89:3576-33580; Garrad et al. (1991) Bio/Technology 9: 1373-1377; Hoogenboom et al. ((1991) Nuc. Acid Res. 19: 4133-4137; Barbas et al. (1991) PNAS 88:7978-7982.
В предпочтительном варианте для выделения человеческих антител с высокой аффинностью и низкой константой скорости диссоциации в отношении hTNF мышиные антитела против hTNF, имеющие высокую аффинность и низкую константу скорости диссоциации в отношении hTNF (например, МАК 195, гибридома для получения которых имеет номер депонента ЕСАСС 87 050801), впервые используют для селекции последовательностей человеческих тяжелой и легкой цепей, имеющих близкую активность связывания в отношении hTNF, с применением эпитопного импринтинга или направленной селекции, данные способы описаны Hoogenboom et al., Публикация РСТ No WO 93/06213. Библиотеками антител, использованными в данном способе, являются предпочтительно библиотеки scFv, полученные и исследованные, как описано McCafferty et al.; Публикация РСТ No WO 92/01047; McCafferty et al., Nature (1990)348:552-554; Griffiths et al. (1993) EMBO J. 12:725-734. Скрининг библиотек антител scFv предпочтительно проводят, используя в качестве антигена рекомбинантный человеческий hTNF.
После выбора исходных сегментов человеческих VL и VH проводят эксперименты “смешивания и определения соответствия”, в которых проводят скрининг различных пар исходно выбранных сегментов VL и VH в отношении связывания hTNF с целью селекции предпочтительных комбинаций пар VL/VH. Кроме того, для дальнейшего улучшения аффинности и/или снижения константы скорости диссоциации связывания hTNF, сегменты VL и VH предпочтительной пары (пар) VL/VH могут быть подвергнуты ненаправленному мутагенезу, предпочтительно в области CDR3 VH и/или VL, способом, аналогичным способу получения соматической мутации in vivo, обусловливающим созревание аффинности антител в течение естественного иммунного ответа. Данное созревание аффинности in vitro может быть осуществлено амплификацией участков VH и VL с использованием ПЦР-праймеров, комплементарных VH CDR3 или VL CDR3 соответственно, данные праймеры “вбивают” со случайной смесью четырех нуклеотидных оснований в определенные положения, так что полученные продукты ПЦР кодируют сегменты VH и VL, в которых случайные мутации были интродуцированы в участки CDR3 VH и/или VL. Данные сегменты VH и VL со случайными мутациями могут быть снова подвергнуты скринингу в отношении связывания hTNF, и могут быть отобраны последовательности, которые проявляют высокую аффинность и низкую константу скорости диссоциации в отношении hTNF.
Аминокислотные последовательности отобранных тяжелой и легкой цепей антитела можно сравнить с аминокислотными последовательностями тяжелой и легкой цепей зародышевой линии. В случаях, когда определенные скелетные остатки отобранных цепей VL и/или VH отличаются от конфигурации зародышевой линии (например, в результате соматической мутации генов иммуноглобулина, использованной для получения фаговой библиотеки), может быть желательным “обратная мутация” измененных скелетных остатков отобранных антител с возвращением к конфигурации зародышевой линии (т.е. изменение скелетных аминокислотных последовательностей отобранных антител, чтобы они стали такими же, как скелетные аминокислотные последовательности зародышевой линии). Такая “обратная мутация” (или “приведение к зародышевой линии”) скелетных остатков может быть выполнена стандартными молекулярно-биологическими способами интродукции специфических мутаций (например, сайт-направленным мутагенезом, ПЦР-обусловленным мутагенезом и др.).
После скрининга и выделения антитела против hTNF, соответствующего изобретению, из библиотеки рекомбинантных иммуноглобулинов нуклеиновая кислота, кодирующая отобранное антитело, может быть выделена из демонстрационной упаковки (например, из генома фага) и субклонирована в другие экспрессирующие векторы стандартными способами с использованием рекомбинантной ДНК. Если желательно, возможны дальнейшие манипуляции с нуклеиновой кислотой для создания других форм антител, соответствующих изобретению (например, связывание с нуклеиновой кислотой, кодирующей дополнительные иммуноглобулиновые домены, такие как дополнительные константные участки). Для экспрессии рекомбинантного человеческого антитела, выделенного при скрининге комбинаторной библиотеки, ДНК, кодирующую антитело, клонируют в рекомбинантный экспрессионный вектор и интродуцируют в клетки-хозяина млекопитающего, как подробно описано выше в Разделе II.
IV. Фармацевтические композиции и фармацевтическое применение
Антитела и фрагменты антител, соответствующие изобретению, предлагаются для использования в производстве средств для лечения нарушений, при которых активность TNF является вредной, и могут быть введены в фармацевтические композиции, пригодные для введения субъекту. В типичном случае фармацевтическая композиция содержит антитело или фрагмент антитела, соответствующие изобретению, и фармацевтически приемлемый носитель. Используемый здесь термин “фармацевтически приемлемый носитель” включает любые возможные растворители, дисперсионные среды, оболочки, антибактериальные и противогрибковые агенты, изотонические и задерживающие абсорбцию агенты и др., которые являются физиологически совместимыми. Примеры фармацевтически приемлемых носителей включают один или более из перечисленных далее носителей: вода, солевой раствор, забуференный фосфатом солевой раствор, декстроза, глицерин, этанол и т.п., а также их комбинации. Во многих случаях будет предпочтительным включить в композицию изотонические агенты, например, сахара, многоатомные спирты, такие как маннит, сорбит или хлорид натрия. Фармацевтически приемлемые носители могут далее содержать минорные количества добавочных субстанций, таких как увлажняющие агенты или эмульгаторы, консерванты или буферы, которые увеличивают время хранения или эффективность антитела или фрагмента антитела.
Композиции, соответствующие данному изобретению, могут находиться в различных формах. Они включают, например, жидкие, полутвердые и твердые дозированные формы, такие как жидкие растворы (например, растворы для инъекций и вливаний), дисперсии или суспензии, таблетки, пилюли, порошки, липосомы и суппозитории. Предпочтительная форма зависит от предполагаемого способа введения и терапевтического применения. Типичные предпочтительные композиции представлены формой растворов для инъекций или вливаний, таких как композиции, близкие к композициями, применяемым для пассивной иммунизации человека другими антителами. Предпочтительным способом введения является парэнтеральный (например, внутривенный, подкожный, внутрибрюшинный, внутримышечный). В предпочтительном варианте осуществления антитело вводят внутривенным вливанием или инъекцией. В другом предпочтительном варианте осуществления антитело вводят внутримышечной или подкожной инъекцией.
Терапевтические композиции в типичном случае должны быть стерильными и стабильными в условиях производства и хранения. Композиции могут быть сделаны в виде растворов, микроэмульсий, дисперсий, липосом или других назначенных структур, подходящих для высокой концентрации лекарственного вещества. Стерильные инъекционные растворы могут быть приготовлены введением активного соединения (т.е. антитела или фрагмента антитела) в необходимом количестве в соответствующий растворитель с одним или комбинацией перечисленных выше ингредиентов, как это требуется, с последующей стерилизацией путем фильтрации. В основном дисперсии готовят введением активного соединения в стерильный носитель, который содержит базовую дисперсионную среду и требующиеся иные ингредиенты из перечисленных выше. В случае стерильных порошков для приготовления стерильных инъекционных растворов предпочтительными способами приготовления являются вакуумная сушка и лиофилизация, с помощью которых получают порошок активного ингредиента с любым дополнительным желательным ингредиентом из его предварительного стерилизованного фильтрацией раствора. Надлежащая текучесть раствора может поддерживаться, например, использованием покрытия, такого как лецитиновое, поддержанием требующегося размера частиц в случае дисперсии и использованием поверхностно-активных веществ. Пролонгированное всасывание инъекционных композиций может быть достигнуто включением в композицию агента, который задерживает поглощение, например, солей моностеарата и желатина.
Антитела и фрагменты антител, соответствующие изобретению, могут вводиться различными способами, известными в уровне техники, хотя из многих способов терапевтического применения предпочтительным способом/путем введения является внутривенная инъекция или вливание. Как будет оценено специалистом, способ и/или путь введения будет изменяться в зависимости от желаемых результатов. В некоторых вариантах осуществления активное соединение может быть приготовлено с носителем, который будет защищать соединение от быстрого выхода так, как в препарате с контролируемым выходом, включая имплантаты, чрескожные пластыри и микроинкапсулированные системы доставки. Могут быть использованы биодеградируемые биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Многие способы приготовления таких препаратов запатентованы и в основном известны специалистам. См., например, Sustained and Controlled Release Drug Delivery Systems. J.R.Robinson ed. Marcel Dekker, Inc., New York, 1978.
В некоторых вариантах осуществления антитело или фрагмент антитела, соответствующие изобретению, могут быть введены перорально, например, с инертным разбавителем или усваиваемым съедобным носителем. Соединение (и другие ингредиенты, если желательно) могут быть также заключены в желатиновую капсулу с твердой или мягкой оболочкой, спрессованы в таблетки или введены непосредственно в пищу субъекта. Для перорального терапевтического введения соединения могут быть введены с наполнителями и использованы в форме таблеток для приема внутрь, защечных таблеток, лепешек, капсул, эликсиров, суспензий, сиропов, облаток и т.п. Для введения соединения, соответствующего изобретению, способом, отличным от парэнтерального, может быть необходимо покрытие соединения оболочкой из материала, препятствующего его инактивации, или совместное введение соединения с таким материалом.
Дополнительные активные соединения могут быть также введены в композиции. В некоторых вариантах осуществления антитело или фрагмент антитела, соответствующие изобретению, содержатся в препарате и/или вводятся совместно с одним или более из терапевтических агентов, которые используют для лечения нарушений, в которых активность hTNF является вредной. Например, антитело или фрагмент антитела против hTNF, соответствующие изобретению, могут входить в препарат и/или вводиться совместно с одним или более дополнительными антителами, которые связывают другие мишени (например, антитела, которые связывают другие цитокины или связывают поверхностные молекулы клетки), с одним или более цитокинами, с растворимым рецептором TNF (см., например, Публикацию РСТ No WO 94/06476) и/или с одним или более химическим агентом, которые ингибируют продукцию или активность hTNF (такими как производные циклогексанилидена, как описано в Публикации РСТ No WO 93/19751. Более того, одно или более антител, соответствующих изобретению, может быть использовано в комбинации с одним или более из вышеупомянутых терапевтических агентов. В таких способах комбинированной терапии могут удачно использоваться низкие дозы вводимых терапевтических агентов, избегая, таким образом, возможной токсичности или осложнений, ассоциированных с различными способами монотерапии.
Неограничивающие объем изобретения примеры терапевтических агентов от ревматоидного артрита, с которыми могут комбинироваться антитело или фрагмент антитела, соответствующие изобретению, включают следующие: нестероидное противовоспалительное лекарственное вещество(а) (NSAIDs); цитокин-супрессирующее противовоспалительное лекарственное вещество(а) (CSAIDs); CDP-571/BAY-10-3356 (гуманизированное антитело против hTNF; Celltech/Bayer); cA2 (химерное антитело против hTNF; Centocor); TNFR-IgG молекулярной массы 75 кД (слитый белок рецептор TNF-IgG мол. массы 75 кД; Immunex; см., например, Arthritis & Rheumatism (1994) Vol.37, 295; J. Invest. Med. (1996) Vol.44, 235A); TNFR-IgG молекулярной массы 55 кД (слитый белок рецептор TNF-IgG мол. массы 55 кД; Hoffmann-LaRoche); IDEC-CE9.1/SB 210396 (неистощающее приматизированное антитело против CD4; IDEC/SmithKline; см., например, Arthritis & Rheumatism (1995) Vol.38, 185; DAB 486-IL-2 и/или DAB 389-IL-2 (слитые белки IL-2; Seragen; см., например, Arthritis & Rheumatism (1993) Vol.36, 1223; Anti-Tac (гуманизированное антитело против IL-2Ror; Protein Design Labs/Roche); IL-4 (противовоспалительный цитокин; DNAX/Schering); IL-10 (SCH 52000; рекомбинантный IL-10, противовоспалительный цитокин; DNAX/Schering); IL-4; агонисты IL-10 и/или IL-4 (например, агонистические антитела); IL-1RA (антагонист рецептора IL-1; Synergen/Amgen); TNF-bp/s-TNFR (растворимый TNF-связывающий белок; см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 284; Amer. J.Physiol. – Heart and Circulatory Physiology (1995) Vol.268, pp.37-42); R973401 (ингибитор фосфодиэстеразы типа IV; см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 282); МК-966 (ингибитор СОХ-2 см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 81); илопрост (см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 82); метотрексат; талидомид см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 282) и близкие талидомиду лекарственные вещества (например, Celgen); лефлуномид (противовоспалительное и ингибитор цитокинов, см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 131; Inflammation Research (1996) Vol.45, pp.103-107); транексамовая кислота (ингибитор активации плазминогена; см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 284); Т-614 (ингибитор цитокина см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 282); простагландин Е1 (см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 282); тенидап (нестероидный противовоспалительный лекарственный препарат; см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 280); напроксен (нестероидный противовоспалительный лекарственный препарат; см., например, Neuro Report (1996) Vol.7, pp.1209-1213); мелоксикам (нестероидный противовоспалительныйлекарственный препарат); ибупрофен (нестероидный противовоспалительный лекарственный препарат); пироксикам (нестероидный противовоспалительный лекарственный препарат); диклофенак (нестероидный противовоспалительный лекарственный препарат); индометацин (нестероидный противовоспалительный лекарственный препарат); сульфасалазин (см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 281); азатиоприн (см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 281); ингибитор ICE (ингибитор фермента интерлейкина – 1-конвертирующего фермента); ингибитор zap-70 и/или lck (ингибитор тирозинкиназы zap-70 или lck); ингибитор VEGF и/или ингибитор VEGF-R (ингибиторы ростового фактора клеток сосудистого эндотелия или рецептора ростового фактора клеток сосудистого эндотелия; ингибиторы ангиогенеза); кортикостероидные противовоспалительные лекарственные вещества (например, SB203580); ингибиторы TNF-конвертазы; антитела против IL-12; интерлейкин-11 (см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 296); интерлейкин-13 (см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительны и), 308); ингибиторы интерлейкина-17 (см., например, Arthritis & Rheumatism (1996) Vol.39, No 9 (дополнительный), 120); золото, пеницилламин, хлорохин; гидроксихлорохин; хлорамбуцил; циклофосфамид; циклоспорин; общее лимфоидное облучение: глобулин против тимоцитов; антитела против CD4; токсины CD5; пептиды и коллаген для перорального применения; динатрий лобензарит; Цитокин-регулирующие агенты (CRA) HP228 и НР466 (Houghten Pharmaceuticals, Inc.); антисмысловые фосфоротиоатные олигодезоксинуклеотиды ICAM-1 (ISIS 2302; Isis Pharmaceuticals, Inc.); растворимый рецептор комплемента 1 (ТР10; Т Cell Sciences, Inc.); преднизон; орготеин; гликозаминогликана полисульфат; миноциклин; антитела против IL2R; морские и растительные липиды (жирные кислоты морских животных и растений; см., например DeLuca et al. (1995) Rheum. Dis. Clin. North Am. 21:759-777); ауранофин; фенилбутазон; меклофенамовая кислота; флуфенамовая кислота: иммунный глобулин для внутривенного введения; зилеутон; микофеноловая кислота (RS-61443); такролимус (FK-506); сиролимус (рапамицин); амиприлоза (терафектин); кладрибин (2-хлордезоксиаденозин) и азарибин.
Неограничивающие объем изобретения примеры терапевтических агентов от воспалительных заболеваний кишечника, с которыми можно комбинировать антитела или фрагменты антител, соответствующие изобретению, включают следующее: буденозид; эпидермальный ростовой фактор; кортикостероиды; циклоспорин; сульфасалазин; аминосалицилаты; 6-меркаптопурин; азатиоприн; метронидазол; ингибиторы липоксигеназы; месаламин; олсалазин; балсалазид; антиоксиданты; ингибиторы тромброксана; антагонисты рецептора IL-1; моноклональные антитела против IL-1; моноклональные антитела против IL-6; ростовые факторы; ингибиторы эластазы; соединения пиридинил-имидазола; CDP-571/BAY-10-3356 (гуманизированное антитело против hhTNF; Celltech/Bayer); cA2 (химерное антитело против hhTNF; Centocor); TNFR-IgG молекулярной массы 75 кД (слитый белок рецептор TNF-IgG мол. массы 75 кД; Immunex; см., например, Arthritis & Rheumatism (1994) Vol.37, 295; J. Invest. Med. (1996) Vol.44, 235A); TNFR-IgG молекулярной массы 55 кД (слитый белок рецептор TNF-IgG мол. массы 55 кД; Hoffmann-LaRoche); интерлейкин-10 (SCH 52000; Schering Plough); IL-4; агонисты IL-10 и/или IL-4 (например, агонистические антитела); интерлейкин-11; глюкуронид- или декстран-конъюгированные пролекарственные формы преднизолона, дексаметазона или будезонида; антисмысловые фосфоротиоатные олигодезоксинуклеотиды ICAM-1 (ISIS 2302; Isis Pharmaceuticals, Inc.); растворимый рецептор комплемента 1 (ТР10; Т Cell Sciences, Inc.); медленно высвобождающийся месалазин; метотрексат; антагонисты фактора активации тромбоцитов (PAF); ципрофлоксацин и лигнокаин.
Неограничивающие объем изобретения примеры терапевтических агентов от рассеянного склероза, с которыми можно комбинировать антитела или фрагменты антител, соответствующие изобретению, включают следующее: кортикостероиды; преднизолон; метилпреднизолон; азатиоприн; циклофосфамид; циклоспорин; метотрексат; 4-аминопиридин; тизанидин; интерферон-1а (Avonex; Biogen); интерферон-1a (Betaseron; Chiron/Berlex); Сополимер 1 (Сор-1; Copaxone; Teva Pharmaceutical Industries Inc.); гипербарический кислород; иммуноглобулин для внутривенного введения; клабридин; CDP-571/BAY-10-3356 (гуманизированное антитело против hTNF; Celltech/Bayer); cA2 (химерное антитело против hTNF; Centocor); TNFR-IgG молекулярной массы 75 кД (слитый белок рецептор TNF-IgG мол. массы 75 кД; Immunex; см., например, Arthritis & Rheumatism (1994) Vol.37, 295; J. Invest. Med. (1996) Vol.44, 235A); TNFR-IgG молекулярной массы 55 кД (слитый белок рецептор TNF-IgG мол. массы 55 кД; Hoffmann-LaRoche); IL-10; IL-4; агонисты IL-10 и/или IL-4 (например, агонистические антитела).
Неограничивающие примеры терапевтических агентов от сепсиса, с которыми можно комбинировать антитела или фрагменты антител, соответствующие изобретению, включают следующее: гипертонические солевые растворы; антибиотики; -глобулин; продолжительная гемофильтрация; карбапенемы (например, меропенем); антагонисты цитокинов, такие как TNF, IL-1, IL-6 и/или IL-8; CDP-571/BAY-10-3356 (гуманизированное антитело против hTNF; Celltech/Bayer); cA2 (химерное антитело против hTNF; Centocor); TNFR-IgG молекулярной массы 75 кД (слитый белок рецептор TNF-IgG мол. массы 75 кД; Immunex; см., например, Arthritis & Rheumatism (1994) Vol.37, 295; J. Invest. Med. (1996) Vol.44, 235A); TNFR-IgG молекулярной массы 55 кД (слитый белок рецептор TNF-IgG мол. массы 55 кД; Hoffmann-LaRoche); цитокин-регулирующие агенты (CRA) HP228 иНР466 (Houghten Pharmaceuticals, Inc.); SK & F 107647 (низкомолекулярный пептид; SmithKline Beecham); четырехвалентный гуанилгидразон CNI-1493 (Picower Institute); ингибитор пути биосинтеза тканевого фактора (TFPI; Chiron); PHP (химически модифицированный гемоглобин; APEX Bioscience); хелаторы и хелаты железа, включая комплекс диэтилентриаминпентауксусная кислота – железо (III) (DTPA iron (III); Molichem Medicines); лизофиллин (синтетическая маленькая молекула метилксантина; Cell Therapeutics, Inc.); PGG-глюкан (водорастворимый -1,3-глюкан; Alpha-Beta Technology); аполипопротеин А-1, реконструированный липидами; хиральные гидроксамовые кислоты (синтетические антибактериальные агенты, которые ингибируют биосинтез липида А); антитела против эндотоксина; Е5531 (синтетический антагонист липида A; Eisai America, Inc.); rBPI21 (рекомбинантный N-концевой фрагмент человеческого бактерицидного/повышающего проницаемость белка) и синтетические пептиды против эндотоксина (SAEP; BiosYnth Reserch Laboratories).
Неограничивающие примеры терапевтических агентов от респираторного дистресс-синдрома взрослых (ARDS), с которыми можно комбинировать антитела или фрагменты антител, соответствующие изобретению, включают следующее: антитела против IL-8; терапия замещением поверхностно-активных веществ; С DP-571 /BAY-10-3356 (гуманизированное антитело против hTNF; Celltech/Bayer); cA2 (химерное антитело против hTNF; Centocor); TNFR-IgG молекулярной массы 75 кД (слитый белок рецептор TNF-IgG мол. массы 75 кД; Immunex; см., например, Arthritis & Rheumatism (1994) Vol.37, 295; J. Invest. Med. (1996) Vol.44, 235A); TNFR-IgG мол. массы 55 кД (слитый белок рецептор TNF-IgG мол. массы 55 кД; Hoffmann-LaRoche).
Применение антител или фрагментов антител, соответствующих изобретению, в комбинации с другими терапевтическими агентами обсуждается ниже в подразделе IV.
Фармацевтические композиции, соответствующие изобретению, могут включать “терапевтически эффективное количество” или “профилактически эффективное количество” антитела или фрагмента антитела, соответствующего изобретению. “Терапевтически эффективное количество” означает количество, эффективное в дозах и в течение периодов времени, необходимых для достижения желаемого терапевтического результата. Терапевтически эффективное количество антитела или фрагмента антитела может изменяться в соответствии с такими факторами, как состояние болезни, возраст, пол и масса субъекта и способность антитела или фрагмента антитела вызывать желаемый ответ у субъекта. Терапевтически эффективное количество является также количеством, в котором какие-либо токсические или вредные эффекты антитела или фрагмента антитела перевешиваются терапевтически благоприятными эффектами. Профилактически эффективное количество означает количество, эффективное в дозах и в течение периодов времени, необходимых для достижения желаемого профилактического результата. В типичном случае, поскольку профилактическую дозу используют у субъектов до или на ранней стадии заболевания, то профилактически эффективное количество будет меньше, чем терапевтически эффективное количество.
Схемы дозировки могут быть доведены до получения оптимального желаемого ответа (например, терапевтического или профилактического ответа). Например, может быть введена одна ударная доза, может быть введено несколько отдельных доз через период времени или доза может пропорционально снижаться или повышаться, что определяется нуждами терапевтической ситуации. Особенным преимуществом является приготовление парэнтеральных композиций в стандартной дозированной форме для простоты применения и одинаковой дозировки. Стандартная дозированная форма, как используют здесь, означает физически дискретные единицы, пригодные в качестве отдельных доз для субъектов-млекопитающих, которых лечат; каждая единица содержит определенное количество активного соединения, рассчитанного для получения желаемого терапевтического эффекта, в сочетании с необходимым фармацевтическим носителем. Определение стандартных дозированных форм, соответствующих изобретению, диктуется и прямо зависит от (а) уникальных характеристик активного соединения и определенного терапевтического или профилактического эффекта, который предусматривают получить, и (b) ограничений, присущих способу получения такого активного соединения для лечения чувствительности у субъектов.
Примерный неограничивающий интервал для терапевтически или профилактически эффективного количества антитела или фрагмента антитела, соответствующего изобретению, составляет 0,1-20 мг/кг, более предпочтительно 1-10 мг/кг. Следует отметить, что значения дозы могут изменяться в зависимости от типа и остроты облегчаемого состояния. Кроме того, следует понять, что для любого определенного субъекта должны быть через период времени подобраны специфические схемы дозировки в соответствии с нуждами субъекта и профессионализмом специалиста, который вводит или руководит введением композиций, и что представленные здесь интервалы доз являются только примерными и не предназначены для ограничения объема или применения заявленной композиции.
IV. Применение антител, соответствующих изобретению
С учетом их способности связывать hTNF антитела против hTNF или их фрагменты, соответствующие изобретению, могут быть использованы для детекции hTNF (например, в биологическом образце, таком как сыворотка или плазма), с помощью известного метода иммуноанализа, такого как твердофазный иммуноферментный анализ (ELISA), радиоиммуноанализ (RIA) или иммуногистохимия ткани. Изобретение представляет способ детекции hTNF в биологическом образце, предусматривающий контактирование биологического образца с антителом или фрагментом антитела, соответствующим изобретению, и детекцию либо антитела (или фрагмента антитела), связанного с hTNF, либо несвязанного антитела (или фрагмента антитела) для выявления таким образом hTNF в биологическом образце. Антитело прямо или непрямо метят определяемой субстанцией для облегчения детекции связанного или несвязанного антитела. Подходящие определяемые субстанции включают различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы и радиоактивные материалы. Примеры подходящих ферментов включают пероксидазу хрена, щелочную фосфатазу, -галактозидазу или ацетилхолинэстеразу; примеры подходящих комплексов простетических групп включают стрептавидин/биотин и авидин/биотин; умбеллиферон, флуоресцеин, изотиоцианат флуоресцеина, родамин, дихлортриазиниламинфлуоресцеин, хлорид дансила или фикоэритрин; пример люминесцентного материала включает люминол и примеры подходящих радиоактивных материалов включают 125I, 131I, 35S или 3H
Альтернативно мечению антитела hTNF может быть определен в биологических образцах конкурентным иммуноанализом с использованием стандартов rhTNF, меченных определяемой субстанцией, и немеченых антител против hTNF. В данном анализе смешивают биологический образец, стандарты меченого rhTNF и антитело против hTNF и определяют количество стандарта меченого rhTNF, связанного с немеченым антителом. Количество hTNF в биологическом образце обратно пропорционально количеству стандарта меченого rhTNF, связанного с антителом против hTNF.
Антитело D2E7, соответствующее изобретению, также может быть использовано для детекции TNF из видов, отличных от человека, в частности hTNF приматов (например, шимпанзе, павиана, игрунки, циномолгуса и резуса), свиньи и мыши, поскольку D2E7 может связывать каждое их этих hTNF (обсуждается далее в Примере 4, подраздел Е).
Антитела или фрагменты антител, соответствующие изобретению, способны нейтрализовать активность hTNF in vitro и in vivo (см. Пример 4). Более того, по меньшей мере некоторые антитела, соответствующие изобретению, такие как D2E7, могут нейтрализовать активность hTNF, выделенного из другого вида. Соответственно антитела или фрагменты антител, соответствующие изобретению, могут быть использованы для ингибирования активности hTNF, например, в культуре клеток, содержащих hTNF, у человека или других млекопитающих, имеющих hTNF, с которыми антитела, соответствующие изобретению, перекрестно реактивны (например, шимпанзе, павиана, игрукки, циномолгуса и резуса, свиньи и мыши). В одном варианте осуществления изобретение представляет способ ингибирования активности hTNF, предусматривающий контактирование hTNF c антителом или фрагментом антитела, соответствующим изобретению, при этом активность hTNF ингибируется. Предпочтительно hTNF является человеческим hTNF. Например, антитело или фрагмент антитела, соответствующий изобретению, может быть добавлен в культуру клеток, содержащих или предположительно содержащих hTNF, с целью ингибирования активности hTNF в культуре.
В другом варианте осуществления изобретение представляет способ ингибирования активности hTNF у субъекта, страдающего нарушениями, при которых активность hTNF является вредной. hTNF участвовал в патофизиологии широкого ряда нарушений (см., например, Moeller A. et а1.(1990), Cytokine 2:162-169; Патент США No 5231024 Moeller et al.; European Patent Publication No 260610 B1 Moeller А.). Изобретение представляет способы, касающиеся активности hTNF у субъекта с такими нарушениями, данный способ предусматривает введение субъекту антитела или фрагмента антитела, соответствующего изобретению, при этом ингибируется активность hTNF у субъекта. Предпочтительно hTNF является человеческим hTNF и субъект является человеком. Альтернативно субъект может быть млекопитающим, экспрессирующим hTNF, с которым антитела, соответствующие изобретению, перекрестно реактивны. Далее, субъект может быть млекопитающим, которому был интродуцирован hTNF (например, посредством введения hTNF или экспрессии трансгена hTNF). Антитело, соответствующее изобретению, может быть введено человеку с терапевтическими целями (дальнейшее обсуждение ниже). Более того, антитело, соответствующее изобретению, может быть введено млекопитающему животному, экспрессирующему hTNF, с которым антитело перекрестно реактивно (например, примату, свинье или мыши) с ветеринарной целью или как модели на животном болезни человека. Относительно последнего, такие модели на животных могут быть использованы для оценки терапевтической эффективности антител, соответствующих изобретению (например, тестирования доз и времени курса применения).
Как используют здесь, термин “нарушение, при котором активность hTNF является вредной,” предусматривает включение заболеваний и других нарушений, при которых присутствие hTNF у субъекта, страдающего от нарушения, как было показано или предполагается, либо обусловливает патофизиологию нарушения, либо является фактором, который усугубляет нарушение. Соответственно нарушение, при котором активность hTNF является вредной, представляет собой нарушение, при котором ингибирование активности hTNF, как ожидается, уменьшает симптомы и/или развитие нарушения. Такие нарушения могут быть обнаружены, например, по повышению концентрации hTNF в биологической жидкости субъекта, страдающего от нарушения (например, повышение концентрации hTNF в сыворотке, плазме, синовиальной жидкости и т.п. субъекта), которая может быть определена, например, с помощью антитела против hTNF, как описано выше. Существует множество примеров нарушений, при которых активность hTNF является вредной. Применение антител и фрагментов антител, соответствующих нарушению, при лечении специфических нарушений далее обсуждается ниже.
А. Сепсис
Фактор некроза опухоли играет установленную роль в патофизиологии сепсиса с биологическими эффектами, включающими гипотензию, ослабление миокарда, синдром подтекания сосудов, некроз органов, стимуляцию выхода токсических вторичных медиаторов и активацию каскада свертывания крови (см., например, Moeller A.,et al. (1990), Cytokine 2:162-169; Патент США No 5231024 Moeller et al.; Европейский Патент No 260610 В1 Moeller A.; TraceyK.J. and Cerami A. (1994) Annu. Rev. Med. 45:491-503; Russel D. and Thompson R.C. (1993) Curr. Opin. Biotech. 4:714-721). Соответственно человеческие антитела и фрагменты антител, соответствующие изобретению, могут быть использованы для лечения сепсиса в любом из его клинических проявлений, включая септический шок, эндотоксический шок, сепсис, вызываемый грамотрицательными бактериями, и токсический шоковый синдром.
Более того, для лечения сепсиса антитела против hTNF и фрагменты антител, соответствующие изобретению, могут быть введены вместе с одним или более терапевтическим агентом, который может еще более ослабить сепсис, таким как ингибитор интерлейкина-1 (описанные в Публикациях РСТ No WO 92/16221 и WO 92/17583), цитокин интерлейкин-6 (см., например, Публикацию РСТ No WO 93/11793) или антагонист фактора активации тромбоцитов (см., например, Европейскую Патентную заявку No EP 374510). Другие способы комбинированной терапии для лечения сепсиса обсуждаются ниже в подразделе III.
Кроме того, в предпочтительном варианте осуществления антитело против hTNF и фрагменты антитела, соответствующие изобретению, вводят человеку из подгруппы пациентов с сепсисом, имеющих концентрацию IL-6 в сыворотке или плазме более 500 пг/мл и, более предпочтительно, 1000 пг/мл во время лечения (см. Публикацию РСТ NoWO 95/20978 Daum L. et al.).
В. Аутоиммунные заболевания
Фактор некроза опухоли, как было установлено, играет роль в патофизиологии ряда аутоиммунных заболеваний. Например, TNF участвовал в активации воспаления тканей и вызывал разрушение суставов при ревматоидном артрите (см., например, Moeller A. et al.(1990), Cytokine 2:162-169; Патент США No 5231024 Moeller et al.; Европейский Патент No 260610 В1 Moeller A.; Tracey K.J. and Cerami A., supra; Arend W.P. and Dayer J-M. (1995) Arth. Rheum. 38:151-160; Fava R.A. et al. (1993) Clin. Exp. Immunol. 94:261-266). TNF участвовал также в стимуляции гибели островков клеток и в образовании инсулин-резистентности при диабете (см., например, Tracey and Cerami, supra; Публикация РСТ NoWO 94/08609). TNF участвовал также в появлении цитотоксичности в отношении олигодендроцитов и индукции воспалительных бляшек при рассеянном склерозе (см., например, Tracey and Cerami, supra). Химерные и гуманизированные мышиные антитела против hTNF прошли клиническую проверку для лечения ревматоидного артрита (см., например, Elliot M.J. et al. (1994); Lancet 344:1125-1127; Elliot M.J. et al. (1994) Lancet 344: 1105-1110; Rankin E.C. etal. (1995) Br. J.Rheumatol. 34: 334-342).
Человеческие антитела и фрагменты антител, соответствующие изобретению, могут быть использованы для лечения аутоиммунных заболеваний, в частности, ассоциированных с воспалением, включая ревматоидный артрит, ревматоидный спондилит, остеоартрит и подагрический артрит, аллергию, рассеянный склероз, аутоиммунный диабет, аутоиммунный увеит и почечный синдром. В типичном случае антитело или фрагмент антитела применяются системно, хотя для некоторых нарушений может быть успешным местное применение антитела или фрагмента антитела в области воспаления (например, местное введение в суставы при ревматоидном артрите или наружное применение при диабетических язвах, отдельно или в сочетании с производным циклогексанилидена, как описано в Публикации РСТ No WO 93/19751). Антитело и фрагмент антитела, соответствующие изобретению, также могут применяться с одним или более дополнительными терапевтическими агентами, используемыми при лечении аутоиммунных заболеваний, как описано далее в подразделе III.
С. Инфекционные болезни
Фактор некроза опухоли участвовал в появлении биологических эффектов, наблюдаемых при ряде инфекционных болезней. Например, hTNF участвовал в развитии воспаления головного мозга и тромбозе капилляров и инфаркте при малярии. TNF также участвовал в развитии воспаления головного мозга, индукции разрушения гематоэнцефалического барьера, развитии синдрома септического шока и активации венозного инфаркта при менингите. TNF также участвовал в индукции кахексии, стимуляции пролиферации вирусов и образовании повреждений центральной нервной системы при синдроме приобретенного иммунодефицита (СПИД). Соответственно антитела и фрагменты антител, соответствующие изобретению, могут быть использованы для лечения инфекционных болезней, включая бактериальный менингит (см., например, Европейскую Патентную заявку No ЕР 585705), церебральную малярию, СПИД и СПИД-ассоциированный комплекс (ARC) (см., например, Европейскую Патентную заявку No ЕР 230574), а также вторичную инфекцию цитомегаловируса при трансплантации (см., например, Fietze E., et al. (1994) Transplantation 58: 675-680). Антитела и фрагменты антител, соответствующие изобретению, могут быть использованы для облегчения симптомов, ассоциированных с инфекционными болезнями, включая лихорадку и миалгии, обусловленные инфекцией (такой как грипп), и вторичную кахексию при инфекции (например, вторичную при СПИД или ARC).
D. Трансплантация
Фактор некроза опухоли рассматривали как ключевой медиатор в отторжении трансплантата и болезни трансплантат против хозяина (GVHD) и в образовании побочной реакции, которую отмечали, когда крысиное антитело ОКТЗ, направленное против комплекса Т-клеточный рецептор – CDR, использовали для ингибирования отторжения почечных трансплантатов (см., например, Eason J.D. et al. (1995) Transplantation 59:300-305; Suthanthiran M. and Strom T.B. (1994) New Engl. J.Med. 331: 365-375). Соответственно антитела и фрагменты антител, соответствующие изобретению, могут быть использованы для ингибирования отторжения трансплантата, включая отторжение аллотрансплантатов и ксенотрансплантатов, и для ингибирования GVHD. Хотя антитело или фрагмент антитела могут быть использованы отдельно, более предпочтительно их использование в комбинации с одним или более агентов, которые ингибируют иммунный ответ против аллотрансплантата или ингибируют GVHD. Например, в одном варианте осуществления антитело или фрагмент антитела, соответствующего изобретению, используют в комбинации с одним или более антител, направленных на другие мишени, участвующие в регуляции иммунных ответов, такие как поверхностные молекулы клетки CD25 (рецептор-интерлейкина-2), CD-11 a (LFA-1), CD54 (ICAM-1), CD4, CD45, CD28/CTLA4, CD80 (В7-1) и/или CD86 (В7-2). В еще одном варианте осуществления антитело или фрагментантитела, соответствующие изобретению, используют в комбинации с одним или более основных иммуносупрессирующих агентов, таких как циклоспорин А или FK506.
Е. Злокачественные образования
Фактор некроза опухоли участвовал в индукции кахексии, стимуляции роста опухоли, усилении метастатического потенциала и появлении цитотоксичности при злокачественных новообразованиях. Соответственно антитела или фрагменты антител, соответствующих изобретению, используют при лечении злокачественных новообразований для ингибирования роста опухоли или метастазов и/или для облегчения вторичной кахексии при злокачественных новообразованиях. Антитело или фрагмент антитела могут вводиться системно или местно в область опухоли.
F. Легочные нарушения
Фактор некроза опухоли участвовал в патофизиологии респираторного дистресс-синдрома взрослых (ARDS), включая стимуляцию активации эндотелиальных лейкоцитов, направленность цитотоксичности на пневмоциты и индукцию синдрома подтекания сосудов. Соответственно антитела или фрагменты антител, соответствующих изобретению, могут быть использованы при лечении различных легочных заболеваний, включая респираторный дистресс-синдром взрослых (см., например, Публикацию РСТ No WO 91/04054), легочный шок, хроническое легочное воспалительное заболевание, легочный саркоидоз, легочный фиброз и силикоз. Антитело или фрагмент антитела, соответствующие изобретению, могут вводиться с одним или более дополнительным терапевтическим агентом, используемым при лечении легочных нарушений, как описано ниже в подразделе III.
G. Кишечные нарушения
Фактор некроза опухоли участвовал в патофизиологии воспалительных кишечных нарушений (см., например, Tracy K.J., et al. (1986) Science 234:470-474; SunX-M., etal. (1988) J. Clin. Invest. 81:1328-1331; MacDonald T.T., et al. (1990) Clin. Exp. Immunol. 81: 301-305). Химерные мышиные антитела против hTNF прошли клиническую проверку для лечения болезни Крона (van Dullemen H.M. et al.(1995) Gastroenterology 109:129-135). Человеческое антитело или фрагмент антитела, соответствующие изобретению, могут быть также использованы для лечения кишечных нарушений, таких как идиопатическое воспалительное заболевание кишечника, которое включает два синдрома – болезнь Крона и язвенный колит. Антитело или фрагмент антитела, соответствующие изобретению, могут также вводится с одним или более дополнительным терапевтическим агентом, используемым при лечении кишечных нарушений, как описано ниже в подразделе III.
Н. Сердечные нарушения
Антитела или фрагменты антител, соответствующие изобретению, могут также быть использованы для лечения различных сердечных нарушений, включая ишемию сердца (см., например, Европейскую Патентную заявку No ЕР 453898) и сердечную недостаточность (слабость сердечной мышцы) (см., например, Публикацию РСТ No WO 94/20139).
I. Другие
Антитела или фрагменты антител, соответствующие изобретению, могут также быть использованы для лечения различных нарушений, при которых активность hTNF является вредной. Примеры других заболеваний и нарушений, при которых активность hTNF участвует в патофизиологии и которые, таким образом, могут лечиться с использованием антитела или фрагмента антитела, соответствующего изобретению, включают воспалительные костные нарушения и заболевания, связанные с резорбцией кости (см., например, Bertolini D.R., et al. (1986) Nature 319:516-518; Konig A., et al. (1988) J. Bone Miner. Res. 3:621-627; Lerner U.H. and Onlin A. (1993) J. Bone Miner. Res. 8:147-155 и Shankar G. and Stern P.H. (1993) Bone 14:871-876), гепатит, включая алкогольный гепатит (см., например, McClain C.J. and Cohen D.A. (1989) Hepatology 9:349-351; Felver M.E. et al. (1990) Alcohol Clin. Exp. Res. 14:255-259 и Hansen J. et al. (1994) Hepatology 20:461-474), вирусный гепатит (Sheron N. Et al. (1991) J. Hepatol. 12:241-245 и Hussain M.J. et al. (1994) J. Clin. Pathol. 47:1112-1115) и фульминантный гепатит; нарушения свертывания (см., например, van der Poll Т. et al. (1990) N. Engl. J.Med. 322:1622-1627; van der Poll T. et al. (1991) Prog. Clin. Biol. Res. 367:55-60; ожоги (см., например, Giroir B.P. et al. (1994) Am. J.Physiol. 267:H118-124; Liu X.S. et al. (1994) Burns 20:40-44), реперфузионные повреждения (см., например, Scales W.E. et al. (1994) Am. J.Physiol. 26:1122-1127; SerrickC. et al. (1994) Transplantation 58:1158-1162; Yao Y.M. et al. (1995) Resuscitation 29:157-168), формирование келоида (см., например, McCauleyR.L et al.(1992) J. Clin. Immunol. 12:300-308), формирование рубцовой ткани; гипертермию, заболевания периодонта, ожирение и токсичность облучения.
Данное изобретение далее иллюстрируется следующими примерами, которые не следует истолковывать как ограничивающие. Содержание всех ссылок, патентов и опубликованных патентных заявок, цитированных в данной заявке, введено здесь в виде ссылки.
Пример 1: Кинетический анализ связывания человеческих антител с hTNF
Происходящие в данный момент взаимодействия связывания лиганда (биотинилированного рекомбинантного человеческого hTNF (rhTNF), иммобилизованного на матриксе биосенсора) и аналита (антитела в растворе) измеряют с помощью поверхностного плазменного резонанса (SPR), используя систему BIAcore (Pharmacia Biosensor, Piscataway, NJ). В системе используют оптические свойства SPR для детекции изменений концентраций белка в декстрановом матриксе биосенсора. Белки ковалентно связаны с декстрановым матриксом при известных концентрациях. Антитела инжектируют в декстрановый метрике, и специфическое связывание инжектированных антител и иммобилизованных лигандов обусловливает повышенную концентрацию белков в матриксе и, как результат, изменение сигнала SPR. Эти изменения сигнала SPR регистрируют как единицы резонанса (RU) и выражают относительно времени по оси Y сенсограммы.
Для облегчения иммобилизации биотинилированного rhTNF на матриксе биосенсора стрептавидин ковалентно связывают посредством свободных аминогрупп с декстрановым матриксом с помощью карбоксильных групп на матриксе, предварительно активированных 100 мМ N-гидроксисукцинимида (NHS) и 400 мМ гидрохлорида N-этил-N’-(3-диэтиламинопропил)карбодиимида (EDC). Затем стрептавидин вводят в активированный матрикс. Тридцать пять микролитров стрептавидина (25 мкг/мл), разведенного в ацетате натрия, рН 4,5, вводят в активированный биосенсор и свободные амины на белке непосредственно связывают активированные карбоксильные группы. Непрореагировавшие EDC-сложные эфиры матрикса дезактивируют инъекцией 1 М этаноламина. Чипы связанного со стрептавидином биосенсора имеются также в продаже (Pharmacia BR-1000-16, Pharmacia Biosensor, Piscataway, NJ).
Для получения биотинилированного rhTNF сначала растворяют 5,0 мг биотина (N-гидроксисукцинимидный сложный эфир D-биотинил-с-аминокапроновой кислоты; Boehringer Mannheim, Каталог No 1008 960) и 500 мкл диметилсульфоксида для получения раствора 10 мг/мл. Десять микролитров биотина добавляют к 1 мл rhTNF (2,65 мг/мл) для получения молярного отношения биотина к rhTNF 2:1. Реакционную смесь осторожно перемешивают и инкубируют в течение двух часов при комнатной температуре в темноте. Колонку PD-10, SephadexG-25M (Pharmacia, каталожный номер 17-0851-01) уравновешивают 25 мл холодного PBS (забуференный фосфатом физиологический раствор) и наносят 2 мл rhTNF-биотина на колонку. Колонку элюируют 10×1 мл холодного PBS. Фракции собирают и определяют при OD280 (0,1 OD=1,25 мг/мл). Соответствующие фракции объединяют и хранят при -80°С до использования. Биотинилированный rhTNF также имеется в продаже (R&D Systems, Каталог No FTAOO, Minneapolis, MN).
Биотинилированный rhTNF для иммобилизации на матриксе с помощью стрептавидина разводят в буфере для PBS (Gibco Кат. No 14190-144, Gibco BRL, Grand Island, NY) с добавлением 0,05% (BIAcore) поверхностно-активного вещества Р20 (Pharmacia BR-1000-54, Pharmacia Biosensor, Piscataway, NJ). Для определения способности rhTNF-специфичных антител связывать иммобилизованный rhTNF анализ связывания выполняют следующим образом. Аликвоты биотинилированного rhTNF (25 нМ; аликвоты 10 мкл) инъецируют в стрептавидин-связанный декстрановый матрикс со скоростью потока 5 мкл/мин. Перед инъекцией белка и сразу после нее чистым PBS-буфером промывают каждую проточную камеру. Для того чтобы представить величину связывания данного образца (приблизительно 500 RU) берут разницу между базовым сигналом и сигналом, полученным через приблизительно 30 с после окончания инъекции антитела. Измеряют прямое связывание rhTNF-специфического антитела с иммобилизованным биотинилированным rhTNF. Антитела (20 мкг/мл) разводят в PBS-буфере и инъецируют аликвоты по 25 мкл в матриксы с иммобилизованным белком со скоростью потока 5 мкл/мин. Перед инъекцией белка и сразу после нее чистым PBS-буфером промывают каждую проточную камеру. Для того чтобы представить величину связывания данного образца берут разницу между базовым сигналом и сигналом, полученным после окончания инъекции антитела. Матриксы биосенсора регенерируют с использованием 100 мМ HCl перед инъекцией следующего образца. Для определения констант скорости (Кoff), скорости связывания (Kon) скорости ассоциации (Ka) и скорости диссоциации (Кd) используют программное обеспечение для оценки кинетики BIAcore (версия 2.1).
Полученнные результаты связывания D2E7 (антитела lgG4 полной длины) с биотинилированным rhTNF по сравнению с мышиным mAb МАК 195 (фрагмент F(ab’)2) представлены ниже в Таблице 1.
| Таблица 1 |
| Связывание lgG4 D2E7 или МАК 195 с биотинилированным rhTNF |
| Антитело |
[Ab], нМ |
rhTNF, связанное RUs |
Ab, связанное RUs |
rhTNF/Ab |
Кoff, сек-1, (Avg) |
| |
|
|
|
|
|
| D2E7 |
267 |
373 |
1215 |
1,14 |
8,45×10-5 |
| |
133 |
420 |
1569 |
1,30 |
5,42×10-5 |
| |
67 |
434 |
1633 |
1,31 |
4,75×10-5 |
| |
33 |
450 |
1532 |
1,19 |
4,46×10-5 |
| |
17 |
460 |
1296 |
0,98 |
3,47×10-5 |
| |
8 |
486 |
936 |
0,67 |
2,63×10-5 |
| |
4 |
489 |
536 |
0,38 |
2,17×10-5 |
| |
2 |
470 |
244 |
0,18 |
3,68×10-5 |
| |
|
|
|
|
(4,38×10-5) |
| Антитело |
[Ab], нМ |
rhTNF, связанное RUs |
Ab, связанное RUs |
rhTNF/Ab |
Koff, сек-1, (Avg) |
| |
|
|
|
|
|
| МАК 195 |
400 |
375 |
881 |
1,20 |
5,38×10-5 |
| |
200 |
400 |
1080 |
1,38 |
4,54×10-5 |
| |
100 |
419 |
1141 |
1,39 |
3,54×10-5 |
| |
50 |
427 |
1106 |
1,32 |
3,67×10-5 |
| |
25 |
446 |
957 |
1,09 |
4,41×10-5 |
| |
13 |
464 |
708 |
0,78 |
3,66×10-5 |
| |
6 |
474 |
433 |
0,47 |
7,37×10-5 |
| |
3 |
451 |
231 |
0,26 |
6,95×10-5 |
| |
|
|
|
|
(4,94×10-5) |
Во второй серии экспериментов молекулярно-кинетические взаимодействия между lgG1 формы D2E7 полной длины и биотинилированного rhTNF анализируют количественно, используя способ BIAcore, как описано выше, и полученные константы кинетики скорости представлены ниже в Таблицах 2, 3 и 4.
| Таблица 2 |
| Экспериментально полученные константы скорости диссоциации взаимодействия D2E7 с биотинилированным rhTNF |
| Эксперимент |
Kd (сек-1) |
| 1 |
9,58×10-5 |
| 2 |
9,26×10-5 |
| 3 |
7,60×10-5 |
| Среднее |
8,81±1,06×10-5 |
| Таблица 3 |
| Экспериментально полученные константы скорости ассоциации взаимодействия D2E7 с биотинилированным rhTNF |
| Эксперимент |
Ka (М-1 сек1) |
| 1 |
1,33×105 |
| 2 |
1,05×105 |
| 3 |
3,36×105 |
| Среднее |
1,91±1,26×105 |
| Таблица 4 |
| Экспериментально полученные константы кинетики скорости и аффинности D2E7 и биотинилированного rhTNF |
| Эксперимент |
Ka (М-1 сек-1) |
Kd (сек-1) |
Kd(M) |
| 1 |
1,33×105 |
9,58×10-5 |
7,20×10-10 |
| 2 |
1,05×105 |
9,26×10-5 |
8,82×10-10 |
| 3 |
3,36×105 |
7,60×10-5 |
2,26×10-10 |
| Среднее |
1,91±1,26×105 |
8,81±1,06×10-5 |
6,09±3,42×10-10 |
Константы скорости диссоциации и ассоциации вычисляют, анализируя участки диссоциации и ассоциации сенсограмм с помощью программного обеспечения анализа BIA. Для взаимодействия D2E7 и молекулы биотинилированного rhTNF принимают условные значения кинетики химических реакций: нулевой порядок кинетики диссоциации и первый порядок кинетики ассоциации. При выборе молекулярных моделей для анализа кинетических данных анализу подвергают взаимодействие только одного плеча бивалентного антитела D2E7 и одной единицы трехмерного биотинилированного rhTNF. Проводят три независимых эксперимента и анализируют результаты отдельно. Средняя полученная константа скорости диссоциации (kd) взаимодействия D2E7 и биотинилированного rhTNF составляет 8,81±1,06×10-5 сек-1 и средняя полученная константа скорости ассоциации (ka) составляет 1,91±1,26×105 М-1 сек-1. Полученную константу внутренней диссоциации (Кd) затем рассчитывают по формуле: Кd=kd/ka. Таким образом, значение Кd антитела D2E7 для rhTNF, выведенное из кинетических параметров, составляет 6,09±3,42×10-10 М. Незначительные различия в кинетических значениях для формы lgG1 антитела D2E7 (представлены в Таблицах 2, 3 и 4) и формы lgG4 D2E7 (представлены в Таблице 1 и Примерах 2 и 3) не могут считаться достоверными различиями, обусловленными присутствием константных участков либо lgG1, либо lgG4, а рассматриваются как связанные с более точными измерениями концентраций антител, используемыми при кинетическом анализе lgG1. Соответственно представленные здесь кинетические значения для формы lgG1 D2E7, как полагают, являются наиболее точными кинетическими параметрами для антитела D2E7.
Пример 2. Сканирующий мутагенез аланина в доменах CDR3 D2E7.
Серию мутаций по одному остатку аланина интродуцируют стандартными способами на протяжении домена CDR3 участков VL D2E7 и VH D2E7. Мутации легкой цепи иллюстрируют Фигурой 1 (LD2E7*.A1, LD2E7*.A3, LD2E7*.A4, LD2E7*.A5, LD2E7*.A7 и LD2E7*.A8, имеющие аланиновую мутацию в положении 1, 3, 4, 5, 7 или 8 соответственно, в домене CDR3 VL D2E7). Мутации тяжелой цепи иллюстрируют Фигурой 4 (HD2E7*.A1, HD2E7*.A2, HD2E7*.A3, HD2E7*.A4, HD2E7*.A5, HD2E7*.A6, HD2E7*.A7, HD2E7*.A8 и HD2E7*.A9, имеющие аланиновую мутацию в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11 соответственно, в домене CDR3 VH D2E7). Кинетику взаимодействия rhTNF с антителом, содержащим VL и VH D2E7 дикого типа, сравнивают с таковой кинетикой антител, содержащих 1) VL D2E7 дикого типа, связанную с аланин-замещенной VH D2E7; 2) VH D2E7 дикого типа, связанную с аланин-замещенной VL D2E7, или 3) аланин-замещенную VL D2E7, связанную с аланин-замещенной VH D2E7. Все антитела тестируют как молекулы lgG4 полной длины.
Кинетику взаимодействия антител с rhTNF определяют с помощью поверхностного плазменного резонанса, как описано в Примере 1. Константы скорости Кoff для различных пар VH/VL приведены ниже в Таблице 5:
| Таблица 5 |
| Связывание аланин-сканмутантов D2E7 с биотинилированным rhTNF |
| VH |
VL |
Koff (сек-1) |
| D2E7 VH |
D2E7 VL |
9,65×10-5 |
| |
|
|
| HD2E7*.A1 |
D2E7 VL |
1,4×10-4 |
| HD2E7*.A2 |
D2E7 VL |
4,6×10-4 |
| HD2E7*.A3 |
D2E7 VL |
8,15×10-4 |
| HD2E7*.A4 |
D2E7 VL |
1,8×10-4 |
| HD2E7*.A5 |
D2E7 VL |
2,35×10-4 |
| HD2E7*.A6 |
D2E7 VL |
2,9×10-4 |
| HD2E7*.A7 |
D2E7 VL |
1,0×10-4 |
| HD2E7*.A8 |
D2E7 VL |
3,1×10-4 |
| HD2E7*.A9 |
D2E7 VL |
8,1×10-4 |
| |
|
|
| D2E7 VH |
LD2E7*.A1 |
6,6×10-5 |
| VH |
VL |
Koff (сек-1) |
| D2E7 VH |
LD2E7*.A3 |
не определяется |
| D2E7 VH |
LD2E7*.A4 |
1,75×10-4 |
| D2E7 VH |
LD2E7*.A5 |
1,8×10-4 |
| D2E7 VH |
LD2E7*.A7 |
1,4×10-4 |
| D2E7 VH |
LD2E7*.A8 |
3,65×10-4 |
| |
|
|
| HD2E7*.A9 |
LD2E7*.A1 |
1,05×10-4 |
Данные результаты демонстрируют, что большинство положений доменов CDR3 участка VL и участка VH D2E7 пригодны для замены одним остатком аланина. Замена одним аланином в положении 1, 4, 5 или 7 в домене CDR3 VL D2E7 или в положении 2, 5, 6, 8, 9 или 10 в домене CDR3 VH D2E7 не оказывает существенного влияния на скорость диссоциации связывания hTNF по сравнению с исходным антителом D2E7 дикого типа. Замена аланином в положении 8 CDR3 VL D2E7 или в положении 3 CDR3 VH D2E7 приводит к повышению Кoff в 4 раза, а замена аланином в положении 4 или 11 CDR3 VH D2E7 приводит к повышению Кoff в 8 раз, указывая на то, что данные положения являются более критическими для связывания hTNF. Однако замена одним аланином в положении 1, 4, 5, 7 или 8 в домене CDR3 VL D2E7 или в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11 в домене CDR3 VH D2E7 приводит к тому, что антитело против hTNF имеет Кoff 1×10-3 сек-1, или менее.
Пример 3: Анализ связывания антител, близких D2E7
Серию антител с последовательностью, близкой D2E7, анализируют на связывание с rhTNF по сравнению с D2E7 с помощью поверхностного плазменного резонанса, как описано в Примере 1. Аминокислотные последовательности тестированных участков VL приводят на Фигурах 1 и 2. Аминокислотные последовательности тестированных участков VH приводят на Фигурах 3 и 4. Константы скорости Кoff для различных пар VH/VL (в указанном формате, либо в виде антител IgG1 или lgG4 полной длины, либо в виде scFv) представлены ниже в Таблице 6:
| Таблица 6 |
| Связывание близких D2E7 антител с биотинилированным rhTNF |
| VH |
VL |
Формат |
Koff (сек-1) |
| D2E7 VH |
D2E7 VL |
lgG1/lgG4 |
9,65×10-5 |
| VH1-D2 |
LOE7 |
lgG1/lgG4 |
7,7×10-5 |
| VH1-D2 |
LOE7 |
scFv |
4,6×10-4 |
| VH1-D2.N |
LOE7.T |
lgG4 |
2,1×10-5 |
| VH1-D2.Y |
LOE7.A |
lgG4 |
2,7×10-5 |
| VH1-D2.N |
LOE7.A |
lgG4 |
3,2×10-5 |
| VH1-D2 |
ЕР В12 |
scFv |
8,0×10-4 |
| VH1-D2 |
2SD4 VL |
scFv |
1,94×10-3 |
| 3С-Н2 |
LOE7 |
scFv |
1,5×10-3 |
| 2SD4 VH |
LOE7 |
scFv |
6,07×10-3 |
| 2SD4 VH |
2SD4 VL |
scFv |
1,37×10-2 |
| VH1A11 |
2SD4 VL |
scFv |
1,34×10-2 |
| VH1B12 |
2SD4 VL |
scFv |
1,01×10-2 |
| VH |
VL |
Формат |
Koff (сек-1) |
| VH1B11 |
2SD4 VL |
scFv |
9,8×10-3 |
| VH1E4 |
2SD4 VL |
scFv |
1,59×10-2 |
| VH1F6 |
2SD4 VL |
scFv |
2,29×10-2 |
| VH1D8 |
2SD4 VL |
scFv |
9,5×10-3 |
| VH1G1 |
2SD4 VL |
scFv |
2,14×10-2 |
| 2SD4 VH |
EP B12 |
scFv |
6,7×10-3 |
| 2SD4 VH |
VL10E4 |
scFv |
9,6×10-3 |
| 2SD4 VH |
VL100A9 |
scFv |
1,33×10-2 |
| 2SD4 VH |
VL100D2 |
scFv |
1,41×10-2 |
| 2SD4 VH |
VL10F4 |
scFv |
1,11×10-2 |
| 2SD4 VH |
VLLOE5 |
scFv |
1,16×10-2 |
| 2SD4 VH |
VLLOF9 |
scFv |
6,09×10-3 |
| 2SD4 VH |
VLLOF10 |
scFv |
1,34×10-2 |
| 2SD4 VH |
VLLOG7 |
scFv |
1,56×10-2 |
| 2SD4 VH |
VLLOG9 |
scFv |
1,46×10-2 |
| 2SD4 VH |
VLLOH1 |
scFv |
1,17×10-2 |
| 2SD4 VH |
VLLOH10 |
scFv |
1,12×10-2 |
| 2SD4 VH |
VL1B7 |
scFv |
1,3×10-2 |
| 2SD4 VH |
VL1C1 |
scFv |
1,36×10-2 |
| 2SD4 VH |
VL1C7 |
scFv |
2,0×10-2 |
| 2SD4 VH |
VL0.1F4 |
scFv |
1,76×10-2 |
| 2SD4 VH |
VL0.1H8 |
scFv |
1,14×10-2 |
Низкие скорости распада (т.е. Кoff (1×10-4 с-1) для антител полной длины (например, формы IgG), имеющих VL, выбранную из D2E7, LOE7, LOE7.T и LOE7.A, которые содержат либо треонин, либо аланин в положении 9, указывают, что положение 9 CDR3 VL D2E7 может быть занято любым из этих двух остатков без существенного влияния на Кoff. Соответственно консенсусный участок CDR3 VL D2E7 имеет аминокислотную последовательность Q-R-Y-N-R-A-P-Y-(T/A) (SEQ ID No 3). Более того, низкие скорости диссоциации (т.е. Кoff1×10-4 с-1) для антител, имеющих VH, выбранную из D2E7,VH1-D2.N и VH1-D2.Y, которые имеют либо тирозин, либо аспарагин в положении 12, указывают, что положение 12 CDR3 VH D2E7 может быть занято любым из этих двух остатков без существенного влияния на Кoff. Соответственно консенсусный участок для CDR3 VH D2E7 имеет аминокислотную последовательность V-S-Y-L-S-T-A-S-S-L-D-(Y/N) (SEQ ID No 4).
Результаты, приведенные в Таблице 6, демонстрируют, что в формате scFv антитела, содержащие участок CDR3 VL или VH 2SD4, имеют более высокую Кoff (т.е. Кoff (1×10-3 с-1) по сравнению с антителами, содержащими участок CDR3 VL или VH D2E7. По CDR3 VL 2SD4 отличается от D2E7 в положениях 2, 5 и 9. Однако, как указывалось выше, положение 9 может быть занято Ala (как в 2SD4) или Thr (как в D2E7) без существенного влияния на Кoff. Таким образом, при сравнении 2SD4 и D2E7 положения 2 и 5 CDR3 VL D2E7 (оба аргинины) могут быть идентифицированы как критические для ассоциации антитела с hhTNF. Эти остатки могут непосредственно вовлекаться в процесс связывания антитела, как контактирующие остатки, или могут играть критическую роль в поддержании структуры складывания молекулы антитела в данном участке. В отношении важности положения 2 отмечают, что замена Arg (в LOE7, которое имеет такой же CDR3 VL, как D2E7) на Lys (в ЕР В12) увеличивает скорость диссоциации на коэффициент два. В отношении важности положения 5 отмечают, что замена Arg (в D2E7) на Ala (в LD2E7*.A5), как описано в Примере 5, также увеличивает скорость диссоциации в два раза. Более того, без любого из Arg в положениях 2 и 5 (в 2SD4) скорость диссоциации повышается в пять раз. Однако следует отметить, что хотя положение 5 является важным для улучшенного связывания hTNF, изменение в этом положении может быть негативным в отношении изменений в других положениях, как видно у VLLOE4, VLLOH1 или VLO.1H8.
Среди CDR3 VH 2SD4 отличается от D2E7 в положениях 1, 7 и 12. Однако, как обсуждалось выше, положение 12 может быть занято Asn (как в 2SD4) или Tyr (как в D2E7) без существенного влияния на константу скорости Koff. Таким образом, при сравнении 2SD4 и D2E7 положения 1 и 7 CDR3 VH D2E7 могут быть идентифицированы как критические для связывания с hTNF. Какописано выше, данные остатки могут непосредственно находиться в сайте связывания антитела, как контактирующие остатки, или могут играть критическую роль в поддержании структуры складывания молекулы антитела в данном участке. Оба положения являются важными для связывания hTNF, поскольку, когда используют CDR3 VH 3C-H2 (который содержит замену валина аланином в положении 1 относительно CDR3 VH D2E7), то scFv имеет скорость диссоциации, которая в три раза выше, чем при использовании CDR3 VH D2E7, но эта скорость все же в четыре раза меньше, чем при использовании CDR3 VH D2E7 (который имеет изменений в обоих положениях 1 и 7 относительно CDR3 VH D2E7).
Пример 4: Функциональная активность D2E7
Для исследования функциональной активности D2E7 это антитело используют в ряде анализов, в которых определяют способность антитела ингибировать активность hTNF либо in vitro, либо in vivo.
А. Нейтрализация hTNF-индуцированной цитотоксичности в клетках L929
Человеческий рекомбинантный hTNF (rhTNF) вызывает клеточную цитотоксичность мышиных клеток L929 после инкубации в течение 18-24 час. Человеческие антитела против hTNF оценивают в анализах с использованием L929 при совместном инкубировании антител с rhTNF и клетками следующим образом. В 96-луночном планшете для микротитрования, содержащем 100 мкл Ab против hTNF, делают серийные дублированные разведения на 1/3 глубины планшета, используя среду RPMI, содержащую 10% эмбриональной телячьей сыворотки (FBS). Добавляют 50 микролитров rhTNF до конечной концентрации 500 пг/мл в каждую лунку с образцом. Затем планшеты инкубируют в течение 30 мин при комнатной температуре. Далее добавляют 50 мкл TNF-чувствительных мышиных фибробластов L929 до конечной концентрации 5×104 клеток на лунку, включая 1 мкг/мл актиномицина D. Контроль содержит среду с клетками и rhTNF с клетками. Эти контрольные опыты и стандартную кривую hTNF в интервале от 2 нг/мл до 8,2 пг/мл используют для определения качества анализа и получения “окна” нейтрализации. Затем лунки инкубируют в течение ночи (18-24 часа) при 37°С в 5% CO2.
Из каждой лунки удаляют сто микролитров среды и добавляют 50 мкл бромида 3,(4,4-диметилтиазол-2-ил)2,5-дифенил-татразолия (ММТ; имеющийся в продаже у Sigma Chemical Co., St. Louis, МО) в PBS с концентрацией 5 мг/мл. Затем планшеты инкубируют в течение 4 часов при 37°С. Затем в каждую лунку добавляют по 50 мкл 20%-ного раствора додецилсульфата натрия (SDS) и планшет инкубируют при 37°С в течение ночи. Измеряют оптическую плотность при 570/630 нм, строят кривые для каждого образца и определяют IC50 стандартными способами.
Характерные результаты для человеческих антител, имеющих различные пары VL и VH по сравнению с мышиным mAb МАК 195, представлены ниже на Фигуре 5 и в Таблице 7.
| Таблица 7 |
| Нейтрализация hTNF-индуцированной цитотоксичности L929 |
| VH |
VL |
Структура |
IC50, M |
| D2E7 |
D2E7 |
scFv |
1,1×10-10 |
| D2E7 |
D2E7 |
lgG4 |
4,7×10-11 |
| 2SD4 |
2SD4 |
scFv/lgG1/lgG4 |
3,0×10-7 |
| 2SD4 |
LOE7 |
scFv |
4,3×10-8 |
| VH1-D2 |
2SD4 |
scFv |
1,0×10-8 |
| VH1-D2 |
LOE7 |
scFv/lgG1/lgG4 |
3,4×10-10 |
| VH1.D2.Y |
LOE7.T |
lgG4 |
8,1×10-11 |
| VH1-D2.N |
LOE7.T |
lgG4 |
1,3×10-10 |
| VH1-D2.Y |
LOE7.A |
lgG4 |
2,8×10-11 |
| VH1-D2.N |
LOE7.A |
lgG4 |
6,2×10-11 |
| МАК 195 |
МАК 195 |
scFv |
1,9×10-8 |
| МАК 195 |
МАК 195 |
F(ab’)2 |
6,2×10-11 |
Результаты, представленные на Фигуре 5 и в Таблице 7, демонстрируют, что человеческие антитела D2E7 против hTNF и различные близкие D2E7 антитела нейтрализуют TNF-индуцированную цитотоксичность L929 с активностью, приблизительно равной активности мышиного mAb МАК 195 против hTNF.
В другой серии экспериментов исследуют способность формы lgG1 D2E7 нейтрализовать hTNF-индуцированную цитотоксичность L929, как описано выше. Результаты трех независимых экспериментов и их среднее значение представлены ниже в Таблице 8:
| Таблица 8 |
| Нейтрализация TNF-индуцированной цитотоксичности L929 посредством lgG1 D2E7 |
| Эксперимент |
IC50 [M] |
| 1 |
1,26×10-10 |
| 2 |
1,33×10-10 |
| 3 |
1,15×10-10 |
| Среднее |
1,25±0,01×10-10 |
Эта серия экспериментов подтверждает, что D2E7 в форме lgG1 полной длины нейтрализует hTNF-индуцированную цитотоксичность L929 со средним значением IC50 [М] 1,25±0,01×10-10.
В. Ингибирование связывания hTNF с рецепторами hTNF на клетках U-937
Способность человеческих антител против hTNF ингибировать связывание hTNF с рецепторами hTNF на поверхности клеток исследуют, используя клеточную линию U-937 (ATCC No CRL 1593), линию гистиоцитов человека, которая экспрессирует рецепторы hTNF. Клетки U-937 выращивают в среде RPMI с добавлением 10% эмбриональной телячьей сыворотки (Hyclone A-1111, Hyclone Laboratories, Logan, UT), L-глутамина (4 нМ), буферного раствора HEPES (10 мМ), пенициллина (100 мкг/мл) и стрептомицина (100 мкг/мл). Для исследования активности антител IgG полной длины клетки U-937 предварительно инкубируют с PBS с добавлением 1 мг/мл человеческого IgG (Sigma 1-4506, Sigma Chemical Co., St. Louis, МО) в течение 45 мин на льду и затем клетки три раза промывают связывающим буфером. Для анализа связывания рецептора клетки U-937 (5×106 клеток/лунку) инкубируют в связывающем буфере (PBS с добавлением 0,2% бычьего сывороточного альбумина) в 96-луночных планшетах для микротитрования (Costar Corp., Cambridge, MA) вместе с меченным 125I rhTNF (3×1-1-10 М; 25 мк Кюри/мл, получаемым от NEN Research Products, Wilmington, DE) в присутствии или без антител против hTNF в общем объеме 0,2 мл. Планшеты инкубируют на льду в течение 1,5 час. Затем 75 мкл каждого образца переносят в тест-пробирки емкостью 1,0 мл (Sarstedt 72.700, Sarstedt Corp., Princeton, NJ), содержащие дибутилфталат (Sigma-D-2270, Sigma Chemical Co., St. Louis, МО) и динонилфталат (ICN 210733, ICN, Irvine, CA). Тест-пробирки содержат 300 мкл смеси дибутилфталата и динонилфталата в объемном соотношении 2:1 соответственно. Свободный (т.е., несвязанный) меченный 125I rhTNF удаляют с помощью микроцентрифугирования в течение пяти минут. Затем конец каждой тест-пробирки, содержащий осадок клеток, отрезают ножницами для микропробирок (Bel-Art 210180001, Be I-Art Products, Pequannock, NJ). Осадок клеток содержит меченный 125I rhTNF, связанный с рецептором hTNF p60 или р80, тогда как водная фаза над масляной смесью содержит избыток свободного меченного 125I rhTNF. Все осадки клеток собирают в пробирке для определения (Falcon 2052, Becton Dickinson Labware, Lincoln Park, NJ) и проводят подсчет в сцинтиляционном счетчике.
Показательные результаты представлены на Фигуре 6. Величина IC50 для ингибирования с помощью D2E7 связывания hTNF с рецепторами hTNF на клетках U-937 составляет в данных экспериментах приблизительно 3×10-10 М. Эти результаты показывают, что человеческое антитело D2E7 против hTNF ингибирует связывание rhTNF с рецепторами hTNF на клетках U-937 в концентрациях, приблизительно равных концентрациям mAb МАК 195 против hTNF.
В другой серии экспериментов способность формы lgG1 D2E7 ингибировать связывание rhTNF с рецепторами hTNF на клетках U-937 исследуют, как описано выше. Результаты трех независимых экспериментов и их среднее значение приведены в Таблице 9.
| Таблица 9 |
| Ингибирование связывания рецептора hTNF на клетках U-937 посредством lgG1 D2E7 |
| Эксперимент |
IC50 [M] |
| 1 |
1,70×10-10 |
| 2 |
1,49×10-10 |
| 3 |
1,50×10-10 |
| Среднее |
1,56±0,12×10-10 |
Данная серия экспериментов подтверждает, что D2E7 в форме lgG1 полной длины ингибирует связывание рецептора TNF на клетках U-937 со средним IC50 [М] 1,56±0,12×10-10.
Для исследования ингибирующей активности D2E7 при связывании 125I-rhTNF, связанного с отдельными рецепторами р55 и р75, проводят твердофазный радиоиммуноанализ. Для определения значений IC50 D2E7 в отношении выделенных рецепторов TNF различные концентрации антитела инкубируют с концентрацией 3×10-10 125I-rhTNF. Затем смесь тестируют на отдельных чашках, содержащих рецепторы TNF р55, либо р75 доза-зависимым способом. Результаты представлены ниже в Таблице 10:
| Таблица 10 |
| Ингибирование связывания рецептора hTNF с р55 и р75 TNF посредством lgG1 D2E7 |
| |
IC50 [M] |
| Реагент |
р 55 TNFR |
р 75 TNFR |
| D2E7 |
1,47×10-9 |
1,26×10-9 |
| RhTNF |
2,31×10-9 |
2,70×10-9 |
Ингибирование связывания 125I-rhTNF с рецепторами TNF р55 и р75 на клетках U-937 с помощью D2E7 сопровождается простой сигмоидной кривой, показывающей близкие величины IC50 для каждого рецептора. В экспериментах с использованием твердофазного радиоиммуноанализа (RIA) с рекомбинантными рецепторами TNF значения IC50 для ингибирования связывания 125I-rhhTNF с рецепторами TNF р55 и р75 с помощью D2E7 вычисляют как 1,47×10-9 и 1,26×10-9 M соответственно. Снижение величин IC50 в твердой фазе, возможно, обусловлено высокой плотностью рецепторов в форме RIA, поскольку немеченый rhTNF также ингибируется с близкими значениями IC50. Значения IC50 для ингибирования связывания 125I-rhTNF с рецепторами р55 и р75 немеченым rhTNF составляют 2,31×10-9 и 2,70×10-9 М соответственно.
С. Ингибирование экспрессии ELAM-1 на HUVEC
Можно индуцировать экспрессию человеческими эндотелиальными клетками пупочной вены (HUVEC) молекулы адгезии 1 эндотелиальных лейкоцитов (ELAM-1) на поверхности клетки обработкой rhTNF, что может быть определено реакцией rhTNF-обработанных HUVEC с мышиным антителом против ELAM-1 человека. Способность человеческих антител против hTNF ингибировать данную hTNF-индуцированную экспрессию ELAM-1 на HUVEC исследуют следующим образом: HUVEC (ATCC NO CRL 1730) помещают в 96-луночные планшеты (5×104 клеток/лунку) и инкубируют в течение ночи при 37°С. На следующий день готовят серийные разведения человеческого антитела против hTNF (1:10) в микролитровой чашке, начиная с 20-100 мкг/мл антитела. Маточный раствор rhTNF готовят с концентрацией 4,5 нг/мл, аликвоты rhTNF добавляют в каждую содержащую антитело лунку и хорошо перемешивают содержимое. Контрольные опыты включают только среду, среду плюс антитело против hTNF и среду плюс rhTNF. Прекращают инкубирование планшетов с HUVEC при 37°С и среду осторожно удаляют отсасыванием из каждой лунки. Двести микролитров смеси антитело-rhTNF переносят в каждую лунку планшетов с HUVEC. Затем планшеты с HUVEC далее инкубируют при 37°С в течение 4 часов. Потом маточный раствор мышиного антитела против ELAM-1 разводят в соотношении 1:1000 в RPMI. Среду из каждой лунки планшета с HUVEC осторожно отсасывают, добавляют 50 мкл на лунку раствора антитела против ELAM-1 и планшеты с HUVEC инкубируют 60 минут при комнатной температуре. Раствор 125I-меченного антитела Ig против мыши готовят в RPMI (приблизительно 50000 импульсов/мин в 50 мкл). Среду из каждой лунки планшетов с HUVEC осторожно удаляют отсасыванием, лунки дважды промывают RPMI и в каждую лунку добавляют 50 мкл раствора 125I-меченного Ig против мыши. Планшеты инкубируют в течение одного часа при комнатной температуре, а затем каждую лунку три раза промывают RPMI. Сто восемьдесят микролитров 5% SDS (додецилсульфата натрия) добавляют в каждую лунку для лизирования клеток. Клеточный лизат из каждой лунки затем переносят в пробирку и подсчитывают в сцинтилляторе.
Полученные результаты представлены на Фигуре 7. Значение IC50 для ингибирования с помощью D2E7 hTNF-индуцированной экспрессии ELAM-1 на HUVEC составляет в данных экспериментах приблизительно 6×10-11 М. Данные результаты демонстрируют, что человеческое антитело против hTNF D2E7 ингибирует hTNF-индуцированную экспрессию ELAM-1 на HUVEC в концентрациях, приблизительно равных концентрациям мышиного mAb МАК 195 против hTNF.
В другой серии экспериментов исследуют способность формы lgG1 D2E7 ингибировать hTNF-индуцированную экспрессию ELAM-1 на HUVEC, как описано выше. Результаты трех независимых экспериментов и среднее значение из них представлены ниже в Таблице 11:
| Таблица 11 |
| Ингибирование TN F-индуцированной экспрессии ELAM-1 рецептором lgG1 D2E7 |
| Эксперимент |
IC50 [M] |
| 1 |
1,95×10-10 |
| 2 |
1,69×10-10 |
| 3 |
1,90×10-10 |
| Среднее |
1,85±0,14×10-10 |
Данная серия экспериментов подтверждает, что D2E7 в форме lgG1 полной длины ингибирует hTNF-индуцированную экспрессию ELAM-1 на HUVEC со средней IC50 [M] 1,85±0,14×10-10.
Нейтрализующую активность lgG1 D2E7 исследуют также в отношении rhTNF-индуцированной экспрессии двух других молекул адгезии ICAM-1 и VCAM-1. Поскольку кривая титрования rhTNF для экспрессии ICAM-1 через 16 часов очень близка к кривой экспрессии ELAM-1, используют такую же концентрацию rhTNF в экспериментах по нейтрализации с помощью антител. HUVEC инкубируют с rhTNF в присутствии различных концентраций D2E7 при 37°С в инкубаторе с СО2 в течение 16 часов и измеряют экспрессию ICAM-1 с помощью мышиного антитела против ICAM-1 и затем 125I-меченного овечьего антитела против мыши. Проводят два независимых эксперимента и вычисляют значения IC50. Неродственное человеческое антитело lgG1 не ингибирует экспрессию ICAM-1.
Экспериментальный способ тестирования ингибирования экспрессии VCAM-1 является таким же, как способ экспрессии ELAM-1 за исключением того, что вместо Mab против ELAM-1 используют Mab против VCAM-1. Проводят три независимых эксперимента и вычисляют значения IC50. Неродственное человеческое антитело lgG1 не ингибируетэкспрессию VCAM-1. Результаты суммируют ниже в Таблице 12:
| Таблица 12 |
| Ингибирование экспрессии ICAM-1 и VCAM-1 посредством lgG1 D2E7 |
| Ингибирование ICAM-1 |
IC50 [M] |
| Эксперимент |
IC50 [M] |
Эксперимент |
IC50 [M] |
| 1 |
1,84×10-10 |
1 |
1,03×10-10 |
| 2 |
2,49×10-10 |
2 |
9,26×10-11 |
| |
|
3 |
1,06×10-10 |
| Среднее |
2,17±0,46×10-10 |
Среднее |
1,01±0,01×10-10 |
Данные эксперименты демонстрируют, что обработка первичных эндотелиальных клеток пупочной вены человека rhTNF приводит к оптимальной экспрессии молекул адгезии: ELAM-1 и VCAM-1 через 4 часа и к максимуму регулируемой экспрессии ICAM-1 через 16 час. D2E7 способно ингибировать экспрессию трех молекул адгезии доза-зависимым образом.
Величины IC50 для ингибирования ELAM-1, ICAM-1 и VCAM-1 составляют 1,85×10-10, 2,17×10-10 и 1,01×10-10 М соответственно. Данные значения очень близки, что указывает на близкие требования к дозе сигнала активации rhTNF, необходимой для индукции экспрессии ELAM-1, ICAM-1 и VCAM-1. Интересно, что D2E7 имеет близкую эффективность при анализе более продолжительного ингибирования экспрессии ICAM-1. Анализ ингибирования ICAM-1 требует 16-часового совместного инкубирования rhTNF и D2E7 с HUVEC в противоположность 4 часам, требующимся для анализов ингибирования ELAM-1 и VCAM-1. Поскольку D2E7 имеет низкую скорость распада в отношении rhTNF, то понятно, что во время 16-часового периода совместного инкубирования не происходит значительной конкуренции за рецепторы TNF на HUVEC.
D. Нейтрализация rhTNF in vivo
Три различные системы in vivo используют для демонстрации того, что D2E7 является эффективным при ингибировании активности rhTNF in vivo.
I. Ингибирование TNF-индуцированной гибели у D-галактозамин-сенсибилизированных мышей
Инъекция рекомбинантного человеческого hTNF (rhTNF) D-галактозамин-сенсибилизированным мышам вызывает гибель в течение 24-часового периода времени. TNF-нейтрализующие агенты, как показывают, предотвращают гибель на этой модели. Для исследования способности человеческих антител против hTNF нейтрализовать rhTNF in vivo на этой модели мышам С57В1/6 внутрибрюшинно (i.p.) инъецируют различные концентрации D2T7-lgG1 или контрольного белка в PBS. Через 30 минут мышам вводят 1 мкг rhTNF и 20 мг D-галактозамина в PBS i.p. и проводят наблюдения через 24 часа. Данное количество rhTNF и D-галактозамина, как ранее устанавливают, вызывает 80-90% летальность у этих мышей. Полученные результаты, представленные как график в виде столбиков выживаемости в % по отношению к концентрации антител, показаны на Фигуре 8. Черные столбики означают D2E7, тогда как заштрихованные столбики означают МАК 196. Инъекция 2,5-25 мкг антитела D2E7 каждой мыши защищает животных от TNF-индуцированной гибели. Значение ED50 составляет приблизительно 1-2,5 мкг на мышь. Антитела положительного контроля МАК 195 близки по своей защищающей способности. Инъекция D2E7 в отсутствие rhTNF не вызывает никакого вредного эффекта на мышей. Инъекция неспецифического человеческого антитела lgG1 не дает никакой защиты от hTNF-индуцированной гибели.
Во втором эксперименте сорок девять мышей делят на 7 равных групп. Каждая группа получает различные дозы D2E7 за тридцать минут до получения дозы LD50 смеси rhTNF/D-галактозамин (10 мкг rhTNF и 20 мг D-галактозамина на мышь). Контрольная группа 7 получает нормальное человеческое каппа-антитело lgG1 в дозе 25 мкг на мышь. Мышей исследуют через 24 часа. Выживаемость для каждой группы представлена ниже в Таблице 13.
| Таблица 13 |
| Выживаемость через 24 час. после обработки D2E7 |
| Группа |
Выживаемость (живые/общее кол-во) |
Выживаемость, % |
| 1 (без антител) |
0/7 |
0 |
| 2 (1 мкг) |
1/7 |
14 |
| 3 (2,6 мкг) |
5/7 |
71 |
| 4 (5,2 мкг) |
6/7 |
86 |
| 5 (26 мкг) |
6/7 |
86 |
| 6 (26 мкг; без rhTNF) |
7/7 |
100 |
| 7 (25 мкг Hu lgG1) |
1/7 |
14 |
II. Ингибирование TNF-индуцированной гипертермии у кроликов
Исследуют эффективность D2E7 в ингибировании rhTNF-индуцированной гипертермии (лихорадки) у кроликов. Группам из трех самок кроликов NZW с массой приблизительно 2,5 кг каждая внутривенно инъецируют D2E7, rhTNF и иммунные комплексы D2E7 и rhTNF. Ректальную температуру измеряют температурными зондами на термографе Кауе каждую минуту в течение приблизительно 4 часов. Рекомбинантный человеческий TNF в солевом растворе, инъецированный в дозе 5 мкг/кг, вызывает повышение температуры более чем на 0,4°С приблизительно через 45 минут после инъекции. Сам препарат антител в солевом растворе в дозе 138 мкг/кг не вызывает повышения температуры у кроликов даже через 140 минут после введения. Во всех дальнейших экспериментах D2E7 или контрольные реагенты (человеческий IgGq или солевой раствор-носитель) инъецируют кроликам внутривенно через 15 минут после внутривенной инъекции rhTNF в солевом растворе в дозе 5 мкг/кг. Репрезентативные результаты некоторых экспериментов представлены ниже в Таблице 14:
| Таблица 14 |
| Ингибирование rhTNF-индуцированной гипертермии у кроликов с помощью D2E7 |
| |
Повышение температуры*, °C |
|
Молярное соотношение |
Пик температуры |
| Доза D2E7 (мкг/кг) |
rhTNF |
rhTNF+D2E7 |
% ингибирования** |
D2E7: rhTNF |
минуты после rhTNF |
| 14 |
0,53 |
0,25 |
53 |
1 |
60 |
| 24 |
0,43 |
0,13 |
70 |
1,6 |
40 |
| 48 |
0,53 |
0,03 |
94 |
3,3 |
50 |
| 137 |
0,53 |
0,00 |
100 |
9,5 |
60 |
| 792 |
0,80 |
0,00 |
100 |
55 |
60 |
* – пик температуры
** – % ингибирования – (1-{повышение температуры при rhTNF & D2E7/повышение температуры при одном rhTNF})×100. |
Внутривенное предварительное введение D2E7 в дозе 14 мкг/кг частично ингибирует пирогенную реакцию по сравнению с кроликами, которым предварительно вводят только солевой раствор. D2E7, введенный в дозе 137 мкг/кг, полностью супрессирует пирогенную реакцию rhTNF в данном эксперименте. Во втором эксперименте D2E7, введенный в дозе 24 мкг/кг, также частично супрессирует пирогенную реакцию по сравнению с кроликами, которым предварительно вводят только солевой раствор. Молярное отношение D2E7 к rhTNF составляет в данном эксперименте 1/6:1. В третьем эксперименте D2E7, инъецируемый внутривенно в дозе 48 мкг/кг (молярное соотношение D2E7:rhTNF=3.3:1), полностью супрессирует пирогенную реакцию по сравнению с кроликами, которым предварительно вводят человеческий lgG1 (контроль) в солевом растворе в дозе 30 мкг/кг. В последнем эксперименте у кроликов, которым предварительно вводят D2E7 (792 мкг/кг) при очень высоком молярном соотношении с rhTNF (55:1), не появляется никакого повышения температуры в течение 4 часов наблюдения. Лечение кроликов иммунными комплексами, полученными из смеси D2E7 и rhTNF, которую инкубируют при 37°С в течение 1 часа при молярном соотношении 55:1, без последующего введения rhhTNF также не приводит к какому-либо повышению температуры в данном эксперименте.
III.Профилактика полиартрита у трансгенных мышей Тg197
Эффект D2E7 на развитие заболевания исследуют на модели артрита на трансгенных мышах. Получают трансгенных мышей Тg197, которые экспрессируют человеческий TNF дикого типа (модифицированный в 3′-участке после кодирующей последовательности), и у данных трансгенных мышей развивается хронический полиартрит со 100% вероятностью в возрасте 4-7 недель (см. EMBO J. (1991) 10:4025-4031 для дальнейшего описания модели полиартрита на Тg197).
Трансгенных мышей идентифицируют с помощью ПЦР в возрасте трех дней. Потомство трансгенных мышей делят на шесть групп.Трансгенных мышей проверяют гибридизационным анализом типа слотблоттинг в возрасте 15 дней. Представляют следующие протоколы лечения для шести групп: Группа 1=без лечения; Группа 2=солевой раствор (носитель); Группа 3=D2E7, 1,5 мкг/г; Группа 4=D2E7, 15 мкг/г; Группа 5=D2E7, 30 мкг/г и Группа 6=контроль изотипа lgG1, 30 мкг/г. Потомство с нетрансгенными мышами также включают в исследования в качестве контроля (Группа 7 – нетрансгенные, без лечения). Каждая группа получает три внутрибрюшинные инъекции в неделю в соответствии с назначенным лечением. Инъекции продолжают 10 недель. Каждую неделю регистрируют макроскопические изменения морфологии суставов для каждого животного. На 10 неделе всех животных умерщвляют и ткани мышей хранят в формалине. Проводят микроскопическое исследование ткани.
Массу животных в граммах определяют для каждой мыши в начале каждой недели. В это же время определяют размер сустава (в мм) как показателя остроты заболевания. Размер сустава определяют как среднее из трех измерений задней правой лодыжки с использованием микрометра. Показатели артрита регистрируют еженедельно следующим образом: 0=отсутствие артрита (нормальный внешний вид и флексия); +=слабый артрит (деформация сустава); ++=средний артрит (распухание, деформация сустава) и +++=тяжелый артрит (анкилоз, определяемый по флексии и сильно затрудненному движению). Гистопатологические показатели на базе окрашивания срезов суставов гемотоксилином/эозином обозначают следующим образом: 0=отсутствие определяемого заболевания; 1=пролиферация синовиальной оболочки; 2=значительное синовиальное уплотнение; 3=деструкция хряща и эрозия кости.
Эффект лечения с помощью D2E7 на средний размер сустава трансгенных мышей с артритом Тg197 представлен на графике Фигуры 11. Гистопатологические показатели и показатели артрита трансгенных мышей Тg197 в возрасте 11 недель суммированы ниже в Таблице 15:
| Таблица 15 |
| Эффект D2E7 на гистопатологию и показатель артрита у мышей Тg197 |
| Группа |
Лечение |
Гистопатологический показатель |
Показатель артрита |
| 1 |
без лечения |
3 (7/7) |
+++ (7/7) |
| 2 |
солевой раствор |
3(8/8) |
+++ (8/8) |
| 6 |
контроль IgG1 |
3(9/9) |
+++ (7/9) |
| 3 |
D2E7, 1,5 мкг/г |
0(6/8) |
0 (8/8) |
| 4 |
D2E7, 15 мкг/г |
0(7/8) |
0 (8/8) |
| 5 |
D2E7, 30 мкг/г |
0(8/8) |
0 (8/8) |
Данный эксперимент демонстрирует, что антитело D2E7 имеет определенный благоприятный эффект в отношении трансгенных мышей, экспрессирующих человеческий TNF (Тg197) дикого типа, выражающийся в отсутствии артрита после периода лечения.
Е. Нейтрализация rhTNF других видов животных с помощью D2E7
Специфичность связывания D2E7 исследуют посредством определения их способности нейтрализовать факторы некроза опухоли других видов приматов и мыши, используя анализ цитотоксичности L929 (как описано выше в Примере 4, подраздел А). Результаты представлены ниже в Таблице 16:
| Таблица 16 |
| Способность D2E7 к нейтрализации TNF различных видов в анализе с L929 |
| hTNF* |
Источник |
IC50 для нейтрализации D2E7 (М)** |
| Человек |
Рекомбинантный |
7,8×10-11 |
| Шимпанзе |
LPS-стимулированный РВМС |
5,5×10-11 |
| Павиан |
Рекомбинантный |
6,0×10-11 |
| Игрунка |
LPS-стимулированный РВМС |
4,0×10-10 |
| Циномолгус |
LPS-стимулированный РВМС |
8,0×10-11 |
| Резус |
LPS-стимулированный РВМС |
3,0×10-11 |
| Собака |
LPS-стимулированный WBC |
2,2×10-10 |
| Свинья |
Рекомбинантный |
1,0×10-7 |
| Мышь |
Рекомбинантный |
>1,0×10-7 |
Результаты в Таблице 16 показывают, что D2E7 может нейтрализовать активность hTNF пяти приматов, приблизительно эквивалентно человеческому hTNF и, более того, может нейтрализовать активность собачьего hTNF (приблизительно в 10 раз слабее, чем человеческого hTNF), а свиного и мышиного hTNF (приблизительно в 1000 раз слабее, чем человеческого hTNF). Более того, связывание D2E7 с растворимой фазой rhTNF не ингибируется другими цитокинами, такими как лимфокин (TNF), IL-1, IL-1, IL-2, IL-4, IL-6, IL-8, IFN и TGF, указывая на то, что D2E7 является очень специфичным в отношении своего лиганда hTNF.
F. Отсутствие выхода цитокина в цельной крови человека, инкубируемой с D2E7
В данном примере исследуют способность D2E7 индуцировать секрецию цитокинов или выделять поверхностные клеточные молекулы у нормальных клеток крови человека. D2E7 инкубируют с разведенной цельной кровью от трех различных нормальных доноров в различных концентрациях в течение 24 час. LPS-положительный контроль проводят в это же время с использованием предварительно определенных концентраций, стимулирующих секрецию цитокинов у иммунокомпетентных клеток крови. Супернатанты собирают и тестируют на панели наборов для ELISA из десяти растворимых цитокинов, рецепторов и молекул адгезии: IL-1, IL-1, антагонист рецептора IL-1, IL-6, IL-8, TNF, растворимый рецептор I TNF, растворимый рецептор II TNF, растворимый ICAM-1 и растворимый Е-селектин. В результате совместного инкубирования антитела D2E7 при концентрациях до 343 мкг/мл не было определено каких-либо значительных количеств цитокинов или выделяемых поверхностных клеточных молекул. Контрольные культуры без добавления антитела также не образуют цитокины в определяемых количествах, тогда как контроль с совместной культурой LPS дает повышенные значения в интервале от многих пикограмм до нескольких нанограмм. Данные результаты указывают, что D2E7 не индуцирует секрецию цитокинов или выделение поверхностных клеточных белков сверх нормальных уровней в культурах ex vivo.
Составной частью данного описания является прилагаемый Список Последовательностей, содержание которого представлено в таблице ниже:
| SEQ ID No: |
Цепь антитела |
Участок |
Тип последовательности |
| 1 |
D2E7 |
VL |
аминокислота |
| 2 |
D2E7 |
VH |
аминокислота |
| 3 |
D2E7 |
VL CDR3 |
аминокислота |
| 4 |
D2E7 |
VH CDR3 |
аминокислота |
| 5 |
D2E7 |
VLCDR2 |
аминокислота |
| 6 |
D2E7 |
VH CDR2 |
аминокислота |
| 7 |
D2E7 |
VLCDR1 |
аминокислота |
| 8 |
D2E7 |
VH CDR1 |
аминокислота |
| 9 |
2SD4 |
VL |
аминокислота |
| 10 |
2SD4 |
VH |
аминокислота |
| 11 |
2SD4 |
VL CDR3 |
аминокислота |
| 12 |
ЕР В12 |
VLCDR3 |
аминокислота |
| 13 |
VL10E4 |
VL CDR3 |
аминокислота |
| SEQ ID No: |
Цепь антитела |
Участок |
Тип последовательности |
| 14 |
VL100A9 |
VL CDR3 |
аминокислота |
| 15 |
VLL100D2 |
VL CDR3 |
аминокислота |
| 16 |
VLLOF4 |
VL CDR3 |
аминокислота |
| 17 |
LOE5 |
VL CDR3 |
аминокислота |
| 18 |
VLLOG7 |
VL CDR3 |
аминокислота |
| 19 |
VLLOG9 |
VL CDR3 |
аминокислота |
| 20 |
VLLOH1 |
VL CDR3 |
аминокислота |
| 21 |
VLLOH10 |
VL CDR3 |
аминокислота |
| 22 |
VL1B7 |
VL CDR3 |
аминокислота |
| 23 |
VL1C1 |
VL CDR3 |
аминокислота |
| 24 |
VLO.1F4 |
VL CDR3 |
аминокислота |
| 25 |
VLO.1H8 |
VL CDR3 |
аминокислота |
| 26 |
LOE7.A |
VL CDR3 |
аминокислота |
| 27 |
2SD4 |
VH CDR3 |
аминокислота |
| 28 |
VH1B11 |
VH CDR3 |
аминокислота |
| 29 |
VH1D8 |
VH CDR3 |
аминокислота |
| 30 |
VH1A11 |
VH CDR3 |
аминокислота |
| 31 |
VH1B12 |
VH CDR3 |
аминокислота |
| 32 |
VH1E4 |
VH CDR3 |
аминокислота |
| 33 |
VH1F6 |
VH CDR3 |
аминокислота |
| SEQ ID No: |
Цепь антитела |
Участок |
Тип |
| |
|
|
последовательности |
| 34 |
3С-Н2 |
VH CDR3 |
аминокислота |
| 35 |
VH1-D2.N |
VH CDR3 |
аминокислота |
| 36 |
D2E7 |
VL |
нуклеиновая кислота |
| 37 |
D2E7 |
VH |
нуклеиновая кислота |
















Формула изобретения
1. Способ ингибирования активности человеческого TNF, отличающийся тем, что осуществляют контактирование человеческого TNF с выделенным антителом человека или его антигенсвязывающим фрагментом, диссоциирующим из человеческого TNF с Kd, равной 1×10-8 М или менее, и Кoff, равной 1×10-3с-1 или менее, определенных с помощью поверхностного плазменного резонанса, и которое нейтрализует цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50, равной 1×10-7 М или менее, и при этом ингибируется активность человеческого TNF.
2. Способ ингибирования активности человеческого TNF, отличающийся тем, что осуществляют контактирование человеческого TNF с выделенным антителом человека или его антигенсвязывающим фрагментом со следующими характеристиками:
диссоциация из человеческого TNF с константой скорости Кoff, равной 1×10-3 с-1 или менее, определенной с помощью поверхностного плазменного резонанса;
имеет легкую цепь по меньшей мере с доменом CDR3, содержащую аминокислотную последовательность SEQ ID No 3 или SEQ ID No 3, модифицированную заменой одним остатком аланина в положении 1, 4, 5, 7 или 8 или заменой от одной до пяти консервативных аминокислот в положениях 1, 3, 4, 6, 7, 8 и/или 9;
имеет тяжелую цепь по меньшей мере с доменом CDR3, содержащую аминокислотную последовательность SEQ ID No 4 или SEQ ID No 4, модифицированную заменой одним остатком аланина в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11 или заменой от одной до пяти консервативных аминокислот в положениях 2, 3, 4, 5, 6, 8, 9, 10, 11 и/или 12, и при этом ингибируется активность человеческого TNF.
3. Способ ингибирования активности человеческого TNF, отличающийся тем, что осуществляют контактирование человеческого TNF с выделенным антителом человека или его антигенсвязывающим фрагментом, имеющим вариабельный участок легкой цепи (LCVR), содержащий аминокислотную последовательность SEQ ID No 1, и вариабельный участок тяжелой цепи (HCVR), содержащий аминокислотную последовательность SEQ ID No 2, и при этом ингибируется активность человеческого TNF.
4. Применение выделенного антитела человека или его антигенсвязывающего фрагмента, диссоциирующего из человеческого TNF с Кd, равной 1×10-8 М или менее, и Koff, равной 1×10-3 с-1 или менее, определенных с помощью поверхностного плазменного резонанса, и нейтрализуещего цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50, равной 1×10-7 М или менее, в качестве компонента для производства средства для лечения нарушений, при которых активность TNF является вредной, посредством введения антитела субъекту.
5. Применение выделенного антитела человека или его антигенсвязывающего фрагмента со следующими характеристиками:
диссоциация из человеческого TNF с константой скорости Кoff, равной 1×10-3 с-1 или менее, определенной с помощью поверхностного плазменного резонанса;
имеет легкую цепь по меньшей мере с доменом CDR3, содержащую аминокислотную последовательность SEQ ID No 3 или SEQ ID No 3, модифицированную заменой одним остатком аланина в положении 1, 4, 5, 7 или 8 или заменой от одной до пяти консервативных аминокислот в положениях 1, 3, 4, 6, 7, 8 и/или 9;
имеет тяжелую цепь по меньшей мере с доменом CDR3, содержащую аминокислотную последовательность SEQ ID No 4 или SEQ ID No 4, модифицированную заменой одним остатком аланина в положении 2, 3, 4, 5, 6, 8, 9, 10 или 11 или заменой от одной до пяти консервативных аминокислот в положениях 2, 3, 4, 5, 6, 8, 9, 10, 11 и/или 12, в качестве компонента для производства средства для лечения нарушений, при которых активность TNF является вредной, посредством введения антитела субъекту.
6. Применение выделенного антитела человека или его антигенсвязывающего фрагмента, имеющего вариабельный участок легкой цепи (LCVR), содержащий аминокислотную последовательность SEQ ID No 1, и вариабельный участок тяжелой цепи (HCVR), содержащий аминокислотную последовательность SEQ ID No 2, для производства средства для лечения нарушений, при которых активность TNF является вредной, посредством введения антитела субъекту.
7. Применение антитела D2E7 в качестве компонента для производства средства для лечения нарушений, при которых активность TNF является вредной, посредством введения антитела субъекту.
8. Выделенное человеческое антитело или его антигенсвязывающий фрагмент, которое диссоциирует из человеческого TNF с Kd 1×10-8 М или менее и константой скорости Koff 1×10-3 с-1 или менее, определенных с помощью поверхностного плазменного резонанса, и нейтрализует цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50 1×10-7 М или менее.
9. Выделенное человеческое антитело или его антигенсвязывающий фрагмент по п.8, которое диссоциирует из человеческого TNF с константой скорости Koff 5×10-4 с-1 или менее.
10. Выделенное человеческое антитело или его антигенсвязывающий фрагмент по п.8, которое диссоциирует из человеческого TNF с константой скорости Koff 1×10-4 с-1 или менее.
11. Выделенное человеческое антитело или его антигенсвязывающий фрагмент по п.8, которое нейтрализует цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50 1×10-8 М или менее.
12. Выделенное человеческое антитело или его антигенсвязывающий фрагмент по п.8, которое нейтрализует цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50 1×10-9 М или менее.
13. Выделенное человеческое антитело или его антигенсвязывающий фрагмент по п.8, которое нейтрализует цитотоксичность человеческого TNF в стандартном анализе с использованием L929 in vitro с IC50 1×10-10 М или менее.
14. Выделенное человеческое антитело или его антигенсвязывающий фрагмент по п.8, которое является рекомбинантным антителом или его антигенсвязывающим фрагментом.
15. Выделенное человеческое антитело или его антигенсвязывающий фрагмент по п.8, которое ингибирует индуцированную человеческим TNF экспрессию ELAM-1 на клетках эндотелия пупочной вены человека.
РИСУНКИ
|
|