Патент на изобретение №2154372
|
||||||||||||||||||||||||||
(54) СПОСОБ ОЦЕНКИ ЖИЗНЕСПОСОБНОСТИ СОСНОВЫХ ДРЕВОСТОЕВ
(57) Реферат: Изобретение относится к лесному хозяйству. Биоиндикаторный способ оценки жизнеспособности основных древостоев основан на использовании природных способностей ксилофильных насекомых чутко реагировать на малейшие изменения состояния деревьев, заселяя и успешно развиваясь на определенном этапе снижения их сопротивляемости. Для каждого вида насекомого этой группы присуще при этом свое оптимальное физиологическое состояние дерева как кормового объекта и среды обитания. Чем больше видов ксилофильных насекомых поселилось на дереве в процессе его отмирания, тем выше была его исходная резистентность (сопротивляемость). Для количественной оценки состояния жизнеспособности древостоев использован ряд параметров структурной организации ксилофильных энтомокомплексов, вошедших в эмпирическую формулу, отображающую интегральный показатель, изменяющийся от 0 до 100% и названный индексом резистентности. Способ позволяет объективно и количественно оценить роль ксилофильных насекомых в процессе отпада деревьев и состояние жизнеспособности древостоев. 1 табл. Изобретение относится к лесному хозяйству, а именно к способам оценки жизнеспособности деревьев. Комплекс ксилофильных насекомых, представляющий собой особую форму эфемероидных паразитоценозов, является неотъемлемой частью лесных экосистем, обеспечивая внутренний гомеостаз и нормальное их функционирование в постоянно изменяющихся условиях среды. Идея об использовании ксилофильных насекомых в качестве биоиндикаторов зародилась давно, однако, несмотря на большое число проведенных исследований [1-10], точных количественных способов до сих пор не найдено. В основе идеи лежит способность насекомых-ксилофагов чутко реагировать на малейшее изменение состояния деревьев, последовательно заселяя их на определенном этапе физиологического ослабления и образуя в итоге так называемые экологические группировки видов [1, 4, 5, 7, 9]. Первоначально на деревья нападают наиболее активные виды, способные подавлять их резистентность и более или менее успешно развиваться на вполне жизнеспособных кормовых объектах. По мере падения устойчивости деревьев получают возможность поселения и развития на них все более новые виды ксилофагов. Появление на кормовом объекте каждого нового вида насекомого свидетельствует о качественно новом этапе падения устойчивости дерева. К моменту завершения отмирания тканей в пределах всего ствола происходит окончательное формирование экологических группировок ксилофагов. Отсюда вытекает вывод: чем больше видов ксилофагов нашло себе приют на дереве, тем выше была его исходная жизнеспособность. В наиболее близком к заявляемому способу оценки жизнеспособности деревьев утверждается обратное, а именно, что число видов насекомых, заселивших дерево, прямо пропорционально степени его ослабленности [3, 5]. Заявляется биоиндикаторный способ оценки жизнеспособности сосновых древостоев, основанный на использовании закономерностей структурной организации ксилофильных энтомокомплексов, выраженных количественно в виде индекса резистентности (  ), вычисляемого по эмпирически найденной формуле = 100 ), вычисляемого по эмпирически найденной формуле = 100 {4BK2EV/[8,1(1-exp(-0,185D)]}2, (1) {4BK2EV/[8,1(1-exp(-0,185D)]}2, (1)где ВК – видовая концентрация энтомокомплекса (число видов насекомых, поселившихся в среднем на одном дереве); D – средний диаметр заселенных деревьев, см; E – индекс выравненности Пиелу, характеризующий степень однородности структуры комплекса ксилофильных насекомых, вычисляемый по формуле 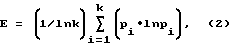 где k – общее число видов в энтомокомплексе; pi – встречаемость i-го вида на заселенных деревьях, доля единицы; V – относительная скорость заселения деревьев комплексом ксилофильных насекомых, показывающая эффективность “работы” по преодолению резистентности дерева, проделанной одним абстрактным видом, рассчитываемая по формуле V = 0,5 b/(c+1), (3)где b, c – параметры функции рангового распределения встречаемости ксилофагов в энтомокомплексе, имеющей следующий вид: pi = ВК [1-exp(-bRi)]c, Ri = 1, 2 … k, (4)где Ri – ранг вида в порядке снижения его встречаемости. Одним из факторов, определяющих величину индекса ВК и резистентности деревьев, является их размер – чем крупнее деревья, тем медленнее, как правило, идет процесс их отмирания и тем большее число видов ксилофагов находит себе на них приют. Однако, как показали исследования, на одном стволе дерева сосны обыкновенной поселяется в общей сложности не более восьми видов ксилофильных насекомых, участвующих в первичном разрушении отмирающих тканей флоэмы и ксилемы. Причем этот предел достигается довольно быстро, что аппроксимируется асимтотической функцией следующего вида: limВК = int[8,1 (1-exp(-0.185D)], (5)где limBK – предельное число видов ксилофагов, которое может быть встречено на стволе одного дерева; int – целая часть числа; D – диаметр дерева, см. математический конструкт (5) введен в формулу (1). Значение индекса изменяется от 0 до 100%, показывая величину исходной резистентности деревьев, отработанных насекомыми, и степень жизнеспособности древостоя. Это положение обосновано тем, что ксилофильные насекомые, как и все хищники, при выборе кормового объекта идут по пути наименьшего сопротивления, и их жертвами становятся, как правило, наиболее ослабленные особи. Следовательно, чем выше резистентность (сопротивляемость) жертв, тем выше жизнестойкость данной популяции. Изменение значений индекса резистентности и параметров математической модели рангового распределения ксилофильных насекомых в энтомокомплексах иллюстрирует ряд конкретных примеров, приведенных в таблице.
Установлено, что оптимум развития комплексов ксилофильных насекомых в различных очагах их размножения заключен в очень узких границах резистентности деревьев, составляющей 20-40% их физиологического предела. При величине резистентности деревьев, меньшей 20%, их ткани быстро отмирают и утрачивают пищевую ценность для личинок этих насекомых. При уровне резистентности более 40% значительная часть жизненной энергии популяций ксилофагов расходуется на преодоление защитного барьера, и смертность их потомства резко возрастает. При остром дефиците кормовых ресурсов в лесу некоторые виды ксилофагов могут успешно заселять вполне здоровые деревья, подавляя их резистентность, уровень которой приближается к 100%, массовостью своих “атак”, однако большая часть их потомства в этом случае погибает от недостатка пищи. Эти данные убедительно свидетельствуют о том, что очаги массового размножения ксилофильных насекомых возникают и затухают только в результате изменения жизнеспособности древостоев.
Заявляемый способ биоиндикации позволяет объективно и количественно оценить роль ксилофильных насекомых в процессе отпада деревьев и состояние жизнеспособности древостоев.
Список литературы1. Валента В.Т. Формирование экологических группировок стволовых вредителей сосен, зависимо от типа отмирания дерева/ Науч. тр. ЛитНИИЛХ. Каунас, 1960.- Т. 5.- С. 183-226. 3. Динамика численности лесных насекомых/Исаев А.С., Хлебопрос Р.Г., Недорезов Л.В. и др.- Новосибирск: Наука, 1984.- 224 с. 5. Исаев А.С., Гирс Г.И. Взаимодействие дерева и насекомых-ксилофагов.- Новосибирск: Наука, 1975.- 347 с. 7. Катаев О.А., Мозолевская Е.Г. Экология стволовых вредителей (очаги, их развитие, обоснование мер борьбы): Учебное пособие.- Л.: ЛТА, 1981.- 87 с. 8. Линдеман Г.В. Взаимоотношение насекомых-ксилофагов и лиственных деревьев в засушливых условиях.- М.: Наука, 1993.- 207 с. 9. Маслов А.Д., Кутеев Ф.С., Прибылова М.В. Стволовые вредители леса.- М.: Лесная пром-сть, 1973.- 144 с. 10. Мозолевская Е. Г., Катаев О.А., Соколова Э.С. Методы лесопатологического обследования очагов стволовых вредителей и болезней леса.- М.: Лесная пром-сть, 1984.- 152 с. Формула изобретения
= 100{4BK2EV/[8,1(1-exp(-0,185D)]}2,где ВК – видовая концентрация энтомокомплекса (число видов насекомых, поселившихся в среднем на одном дереве); E – индекс выравненности Пиелу; V – относительная скорость заселения деревьев комплексом ксилофильных насекомых; D – средний диаметр заселенных деревьев, см, а жизнеспособность древостоев определяют в соответствии со значением индекса резистентности – от 0 до 100%, при этом при максимальном значении индекса оценивают жизнеспособность сосновых древостоев как высокую, а при минимальном – как низкую. РИСУНКИ
MM4A Досрочное прекращение действия патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе
Дата прекращения действия патента: 30.07.2000
Номер и год публикации бюллетеня: 6-2002
Извещение опубликовано: 27.02.2002
|
||||||||||||||||||||||||||