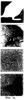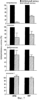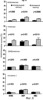Патент на изобретение №2255765
|
||||||||||||||||||||||||||||||||||||||||
(54) СПОСОБ ИНГИБИРОВАНИЯ РОСТА ОПУХОЛЕЙ ГОЛОВНОГО МОЗГА С ПОМОЩЬЮ ОТОБРАННЫХ АНТАГОНИСТОВ ИНТЕГРИНОВ
(57) Реферат:
Изобретение относится к медицине, онкологии, а именно к ингибированию роста опухолей головного мозга хозяина. Способ включает введение хозяину терапевтически эффективного количества антагониста интегрина
(56) (продолжение):
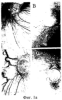
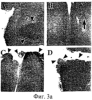
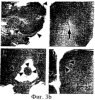
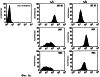
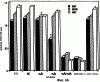
Предпосылки изобретения Область техники Настоящее изобретение, в целом, относится к ингибированию роста опухолей, конкретно к ингибированию роста опухолей головного мозга. Уровень техники В описании настоящей заявки различные ссылки приводятся в круглых скобках. Описания этих публикаций целиком включены в настоящую заявку в качестве ссылок для более полного описания состояния области техники, к которой относится настоящее изобретение. Полные библиографические сведения по этим ссылкам можно найти в конце настоящей заявки, перед формулой изобретения. Опухоли головного мозга, как и прочие солидные опухоли, требуют постоянного увеличения кровоснабжения для поддержания непрерывного роста свыше 1-2 мм3 (1,2). Это осуществляется с помощью ангиогенеза, процесса, который наблюдается в ответ на эндотелиальные факторы роста, высвобождаемые опухолевыми клетками. Ангиогенез включает индукцию пролиферации эндотелиальных клеток из микрососудистого русла, находящегося в состоянии покоя, миграцию неоэндотелия по направлению к опухолевому ложу и, наконец, созревание в новую капиллярную сеть (3). Опухоли головного мозга являются наиболее ангиогенными из всех новообразований у человека. Главными ангиогенными факторами, которые демонстрируются как гибридизацией in situ, так и специфичными антителами в срезах тканей пациентов с глиобластомой и медуллобластомой, наиболее распространенными злокачественными опухолями головного мозга, являются сосудистый эндотелиальный фактор роста (VEGF) и основной фактор роста фибробластов (bFGF) (4-7). Фактически, экспрессия VEGF и плотность микрососудов в глиальных опухолях прямо коррелирует со степенью злокачественных характеристик и общим результатом (8-9). Недавно полученные данные наводят на мысль о том, что ангиогенез регулируется путем активации интегринов эндотелиальных клеток, семейства трансмембранных рецепторов, которые управляют адгезией клеток к белкам внеклеточного матрикса (ЕСМ), посредством связывания с аминокислотной последовательностью Arg-Gly-Asp (RGD) (10). В ответ на bFGF и VEGF эндотелиальные клетки увеличивают экспрессию интегринов Исследования с использованием анти- Ранее проводившиеся исследования показали, что связывание клеток карциномы молочной железы, меланомы и аденокарциномы толстого кишечника HT29-D4 с витронектином зависит от Опухоли головного мозга, в силу их высокоинвазивной природы и высокой степени ангиогенеза, представляют собой отличную модель для дальнейшего изучения роли интегринов в прогрессировании опухолей. Многочисленные исследования показали, что плотность микрососудов коррелирует с исходами и степенью злокачественности астроцитом (57-59). Ингибиторы ангиогенеза, такие как TNP-470, тромбоспондин-1 и тромбоцитарный фактор 4, внедряли в экспериментальные опухоли головного мозга и показали ингибирование опухолевого роста (60-62). Однако до настоящего времени ни в одном исследовании не изучалось воздействие антагонизма в отношении интегринов на инвазию и ангиогенез опухолей головного мозга. Следовательно, существует необходимость изучения этого эффекта и, таким образом, создания нового способа лечения опухолей головного мозга. Краткое описание изобретения Настоящее изобретение основано на неожиданном открытии того факта, что направленный антагонизм в отношении интегринов, в частности, Соответственно, в одном аспекте настоящего изобретения представляется способ ингибирования роста опухоли в головном мозге хозяина. Указанный способ включает введение хозяину, который нуждается в указанном ингибировании, терапевтически эффективного количества антагониста интегрина. В одном варианте осуществления настоящего изобретения интегрин представляет собой Другим аспектом настоящего изобретения является представление способа ингибирования ангиогенеза в опухолевой ткани, расположенной в головном мозге хозяина. Указанный способ включает введение хозяину композиции, содержащей ингибирующее ангиогенез количество антагониста интегрина В одном варианте осуществления настоящего изобретения интегрин представляет собой Другим аспектом настоящего изобретения является представление способа ингибирования ЕСМ-зависимой клеточной адгезии клеток опухоли головного мозга, растущих в головном мозге хозяина. Указанный способ включает введение хозяину терапевтически эффективного количества антагониста интегрина Еще одним аспектом настоящего изобретения является представление способа ингибирования витронектин-зависимой клеточной миграции в клетках опухоли головного мозга, растущих в головном мозге хозяина. Указанный способ включает введение хозяину терапевтически эффективного количества антагониста В одном варианте осуществления настоящего изобретения антагонист представляет собой полипептидный антагонист Еще одним аспектом настоящего изобретения является представление способа индуцирования апоптоза опухолевых клеток, растущих в головном мозге хозяина. Указанный способ включает введение хозяину терапевтически эффективного количества антагониста интегрина. В одном варианте осуществления настоящего изобретения интегрин может представлять собой Способы по настоящему изобретению хорошо подходят для применения при лечении опухолей головного мозга in vivo. Настоящее изобретение создает новый терапевтический подход к лечению опухолей головного мозга. Описание фигур Упомянутые выше и другие признаки настоящего изобретения, а также способ их достижения будут более ясными и доступными для понимания при использовании следующего описания в сочетании с прилагаемыми чертежами. Эти чертежи изображают только типичный вариант осуществления настоящего изобретения и не ограничивают, таким образом, его объем. Они служат целям достижения большей конкретности и детальности. На этих чертежах: фиг.1а и 1b показывают ингибирование ангиогенеза (а) и роста опухоли на САМ (b) с помощью антагониста Фиг.2 показывает размер опухоли (А) и выживание мышей (В) после интрацеребральной инъекции клеток DAOY и U87MG; фиг.3а и 3b показывают гистопатологию ортотопически инъецированных клеток опухоли головного мозга DAOY (а) и U87MG (b), у животных, ежедневно получавших неактивный (А) или активный пептид (B-D). Большие интрацеребральные опухоли (показаны стрелками) видны у контрольных животных (А), в то время как у животных, получавших фиг.4 показывает выживание мышей после ортотопической имплантации опухоли головного мозга, получавших активный или неактивный пептид; фиг.5 показывает воздействие антагониста фиг.6a-6d показывают профиль интегринов (а) и воздействие антагониста фиг.7 показывает воздействие циклического пентапептида на адгезию опухолевых клеток к белкам ЕСМ; фиг.8 показывает эффект ингибирования адгезии, приводящий к гибели клеток (апоптозу), для клеток опухоли головного мозга и клеток капилляров головного мозга. Этот эффект ограничивается витронектином и тенасцином ЕСМ; фиг.9 показывает иммуногистохимию опухолей головного мозга U87, ксенотрансплантированных в передний мозг бестимусных мышей, которых лечили активным (анти- Подробное описание изобретения Одним аспектом настоящего изобретения является представление способа ингибирования опухолевого роста в головном мозге хозяина, включающего введение хозяину, который нуждается в указанном ингибировании, терапевтически эффективного количества антагониста интегрина. Способ по настоящему изобретению можно использовать для лечения любых опухолей, которые растут в головном мозге, до тех пор, пока рост опухолей в головном мозге требует взаимодействия интегрина с его лигандом. Примеры такой опухоли включают, но не ограничивают, глиобластому, медуллобластому (астроцитому, другие примитивные злокачественные опухоли, происходящие из нейроэктодермы и стволовых клеток глии головного мозга). Для цели настоящего изобретения, предпочтительно, рост опухоли наблюдается интрацеребрально в головном мозге хозяина. Хозяином может быть любое млекопитающее, включая, без ограничения, крысу и человека. Рост опухоли ингибируется, если рост замедляется лечением. Для цели настоящего изобретения интегрин представляет собой любой член специфического семейства гомологичных гетеродимерных трансмембранных рецепторов. Указанные рецепторы управляют адгезией клеток, посредством связывания с аминокислотной последовательностью Arg-Gly-Asp (RGD). Указанные рецепторы могут экспрессироваться как опухолевыми, так и нормальными клетками. Свойства интегринов хорошо известны и хорошо охарактеризованы и подробно описаны в цитируемых ссылках (1, 2), содержание которых, имеющее отношение к рассматриваемому вопросу, включено в настоящий документ в качестве ссылки. Примеры интегринов включают, но не ограничивают, семейство Антагонист интегрина представляет собой молекулу, которая блокирует или ингибирует физиологическую или фармакологическую активность интегрина путем ингибирования связывающей активности интегрина, рецептора, с его лигандом, различными белками матрикса, включая, без ограничения, витронектин, тенасцин, фибронектин и коллаген I. В одном варианте осуществления настоящего изобретения антагонист интегрина может представлять собой антагонист интегрина Например, антагонистом Подобно этому, антагонистом В одном варианте осуществления настоящего изобретения антагонист представляет собой полипептидный антагонист В соответствии с другим вариантом осуществления настоящего изобретения, антагонист может представлять собой моноклональное антитело, иммуноспецифичное в отношении Для цели настоящего изобретения антагонисты можно использовать в отдельности или в комбинации друг с другом в способе по настоящему изобретению для ингибирования роста опухолей головного мозга. Терапевтически эффективное количество представляет собой количество антагониста, достаточное для обеспечения поддающегося измерению ингибирования роста опухоли в головном мозге хозяина, подвергаемого лечению. Ингибирование роста опухоли можно определять микроскопически после окрашивания, как описано в настоящем документе, MRI-сканированием головного мозга мыши или инкорпорацией 3Н-тимидина; все эти методики хорошо известны специалистам. Поскольку антагонист интегрина может принимать форму RGD-содержащего пептида, анти- Эффективность антагониста можно измерить с помощью ряда способов, включая, без ограничения, ингибирование ангиогенеза в анализе САМ, изучении опухоли in vitro и путем измерения ингибирования связывания натуральных лигандов с интегринами, такими как Пределы доз для введения антагониста зависят от формы антагониста и его эффективности в отношении конкретного интегрина. Специалист может подобрать подходящую дозу для конкретного антагониста с учетом описания настоящего изобретения без излишнего экспериментирования. Доза должна быть достаточно велика, чтобы оказывать желаемое действие, благодаря которому рост опухоли головного мозга ингибируется. Доза не должна быть настолько большой, чтобы вызывать появление побочных эффектов, таких как отек мозга или быстрое высвобождение цитокинов из опухолей мозга, индуцирующее кахексию, например, когда антагонист интегрина по настоящему изобретению вводится в форме полипептида. Доза на кг массы тела может варьировать от 1 до 20 мг, при введении один или более раз в день, в течение одного или нескольких дней, или в течение неопределенного периода времени. Когда антагонист интегрина по настоящему изобретению вводят в форме моноклонального антитела, доза может варьироваться от 1 до 20 мг/кг, при введении один-два раза в неделю в течение неопределенного периода времени. Полипептид или моноклональные антитела по настоящему изобретению можно вводить парентерально путем инъекции или постепенной инфузии в течение некоторого периода времени. Несмотря на то, что ткань, подлежащую лечению, наиболее часто лечат интраперитонеальным или подкожным (антитело) введением, антагонисты по настоящему изобретению можно вводить также интраокулярно, внутривенно, внутримышечно, внутриполостным введением, чрескожно, и использовать при этом перистальтические технические средства. Композиции вводят таким способом, который является совместимым с данной лекарственной формой и в терапевтически эффективном количестве. Количество для введения, а также время введения зависят от субъекта, подвергаемого лечению, способности организма субъекта утилизировать активный ингредиент и степени желательного терапевтического эффекта. Точные количества активного ингредиента, которые требуются для введения, зависят от оценки практикующего врача и являются индивидуальными для каждого субъекта. Однако подходящие пределы доз для системного введения описываются в настоящем документе и зависят от пути введения. Подходящие схемы введения также разнообразны, однако типизируются первоначальным введением, с последующими повторными дозами с интервалами один или более часов инъекцией или другим способом введения. В соответствии с одним вариантом осуществления настоящего изобретения, настоящее изобретение относится также к фармацевтической композиции, пригодной для осуществления на практике терапевтических способов, описанных в настоящем документе. Композиции содержат антагонист по настоящему изобретению в комбинации с фармацевтически приемлемым носителем. Используемые в настоящем документе термины “фармацевтически приемлемый” “физиологически переносимый” и их грамматические варианты, которые относятся к композициям, носителям, разбавителям и реагентам, используются взаимозаменяемым образом и означают, что указанные материалы можно вводить млекопитающему без нежелательных физиологических эффектов. Препараты для парентерального введения пептида или антитела по настоящему изобретению включают стерильные водные или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и пригодные для инъекций органические сложные эфиры, такие как этилолеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая солевые и буферные среды. Парентеральные носители включают раствор хлорида натрия, декстрозу Рингера, декстрозу и хлорид натрия, раствор Рингера с лактатом или жирные масла. Внутривенные носители включают жидкостные и питательные восполнители, электролитные восполнители (такие как восполнители на основе декстрозы Рингера) и т.п. Консерванты и другие добавки также могут иметь место, такие как, например, антимикробные, антиоксидантные, хелатирующие агенты, а также инертные газы и т.п. Другой особенностью настоящего изобретения является способ ингибирования ангиогенеза в опухолевой ткани, расположенной в головном мозге хозяина. Указанный способ включает введение хозяину композиции, включающей ингибирующее ангиогенез количество антагониста интегрина. Как обсуждалось в разделе “Предпосылки к созданию изобретения” ангиогенез представляет собой образование неоваскулярной сети из уже существующих сосудов хозяина и требуется для роста опухоли свыше 1-2 мм3. Для цели настоящего изобретения ангиогенез подавлен, если ангиогенез и симптомы заболевания, опосредованного ангиогенезом, уменьшаются. В одном варианте осуществления настоящего изобретения опухолевая ткань находится интрацеребрально, в головном мозге хозяина. Хозяином может быть любое млекопитающее. Примеры хозяина включают, без ограничения, мышь, крысу и человека. Диапазоны доз для введения антагониста зависят от формы антагониста и его эффективности в отношении конкретного интегрина. Специалист может подобрать подходящую дозу для конкретного антагониста с учетом описания настоящего изобретения без излишнего экспериментирования. Доза должна быть достаточно велика, чтобы оказывать желаемое действие, благодаря которому ангиогенез и симптомы заболевания, опосредованного ангиогенезом, уменьшаются. Доза не должна быть настолько большой, чтобы вызывать появление побочных эффектов, таких как отек мозга из-за быстрого лизиса опухоли с высвобождением цитокинов или кровотечением. В одном варианте осуществления настоящего изобретения антагонист интегрина может представлять собой антагонист интегрина В соответствии с другим вариантом осуществления настоящего изобретения, антагонистом может быть антитело против Терапевтически эффективное количество представляет собой количество антагониста, достаточное для обеспечения поддающегося измерению ингибирования ангиогенеза в ткани, подвергаемой лечению, т.е. количество, ингибирующее ангиогенез. Ингибирование ангиогенеза можно измерить in situ методами иммуногистохимии, как описано в настоящем документе, или с помощью других методик, известных специалистам. В соответствии с одним вариантом осуществления настоящего изобретения, настоящее изобретение относится также к фармацевтической композиции, пригодной для осуществления на практике терапевтических способов, описанных в настоящем документе. Композиции содержат антагонист по настоящему изобретению в комбинации с фармацевтически приемлемым носителем. Еще одной особенностью настоящего изобретения является способ ингибирования ЕСМ-зависимой клеточной адгезии клеток опухоли головного мозга, которые растут в головном мозге хозяина, который включает введение хозяину терапевтически эффективного количества антагониста интегринов Для цели настоящего изобретения ЕСМ-зависимая клеточная адгезия включает любую клеточную адгезию, которая зависит от ЕСМ и которая опосредована Фраза “терапевтически эффективное количество”, используемая в настоящем документе, означает количество антагониста, которое достаточно для уменьшения ЕСМ-опосредованной адгезии опухолевых клеток. Клеточную адгезию можно измерить с помощью способов, описанных в настоящем документе, а также способов, хорошо известных специалистам. В соответствии с одним вариантом осуществления настоящего изобретения антагонист может представлять собой полипептидный антагонист Другой особенностью настоящего изобретения является способ ингибирования витронектин-зависимой клеточной миграции клеток опухоли головного мозга, растущей в головном мозге хозяина, который включает введение хозяину терапевтически эффективного количества антагониста Для цели настоящего изобретения ингибирование достигается, если миграция опухолевых клеток уменьшается. Фраза “терапевтически эффективное количество”, используемая в настоящем документе, означает количество антагониста, которое достаточно для уменьшения витронектин-зависимой миграции опухолевых клеток. Миграцию клеток можно измерить с помощью способов, описанных в настоящем документе, а также способов, хорошо известных специалистам. В соответствии с одним вариантом осуществления настоящего изобретения антагонист может представлять собой полипептидный антагонист Настоящее изобретение относится также к способу индуцирования апоптоза клеток опухоли головного мозга, растущей в головном мозге хозяина. Указанный способ включает введение хозяину терапевтически эффективного количества антагониста интегрина. Для цели настоящего изобретения апоптоз клеток опухоли головного мозга индуцируется, если после введения антагониста наблюдается увеличение количества апоптоза опухолевых клеток головного мозга. Терапевтически эффективное количество представляет собой количество антагониста, достаточное для получения поддающегося измерению апоптоза опухолевых клеток в головном мозге хозяина, подвергаемого лечению. Апоптоз опухолевых клеток в головном мозге можно измерить с помощью способов, описанных в настоящем документе, а также способов, хорошо известных специалистам. В соответствии с одним вариантом осуществления настоящего изобретения, интегрин может представлять собой ПРИМЕРЫ Материалы и методы Материалы: Активный циклический RGD пентапептид EMD 121974, цикло(Arg-Gly-Asp-D-Phe-[N-Me]-Val) и неактивный контрольный пептид cRAD (EMD 135981) были предоставлены А. Jonczyk, Ph.D., Merck KgaA, Darmstadt, Germany. Моноклональные антитела LM-609 и P1-F6 описаны (13). Клеточные линии опухоли головного мозга DAOY и U87MG были приобретены в АТСС, Rockville, MD. Первичные культуры эндотелиальных клеток капилляров головного мозга человека были предоставлены M.Stins, Ph.D., Children’s Hospital, Los Angeles (49). Человеческий витронектин был приобретен у компании Promega, Madison, WI. Анализ FACS: использовался цитометр FACS (Becton-Dickinson, San Jose). Условия описаны ранее (43). Первичные антитела представляли собой LM-609 и P1-F6 при 1:100, а вторичные Аb представляли собой меченные FITC антимышиные IgG козы при 1:250. Определение апоптоза производилось согласно инструкциям производителя (Clontech, Palo Alto, CA, набор для определения апоптоза Аро Alert Annexin V-FITC). Адгезия: Условия анализа были описаны ранее (15). Лунки, обработанные нетканевой культурой, инкубировали в течение ночи при 4° С с витронектином (1-10 мкг/мл PBS), промывали и блокировали в течение 30 мин денатурированным нагреванием 1% BSA в PBS, после чего промывали PBS. Контрольные и тест-клетки, предварительно инкубированные при 37° С с испытуемым веществом (20 мкг/мл) в адгезионном буфере в течение 30 мин, засевали в лунки (5× 104 клеток на лунку) и инкубировали в течение 1 ч при 37° С. После осторожного промывания лунок адгезионным буфером прикрепившиеся клетки фиксировали и окрашивали кристаллическим фиолетовым, краской, растворенной в метаноле и определяли оптическую плотность (ОП) при 600° А. Миграция: Межлуночные поликарбонатные фильтры (размер пор 8 мкм) инкубировали в течение 30 мин при 37° С с витронектином в PBS (1 мкг/мл на верхней стороне и 10 мкг на стороне дна), блокировали в течение 30 мин денатурированным нагреванием 1% BSA в PBS и промывали PBS. Тест-клетки (5× 104 клеток на лунку) добавляли в адгезионный буфер, содержавший указанные испытуемые вещества (20 мкг/мл), и инкубировали в течение 4 ч при 37° С; при этом нижняя камера содержала адгезионный буфер. Клетки из верхней камеры удаляли марлевой салфеткой, а клетки из донной части фильтра фиксировали и окрашивали кристаллическим фиолетовым, а количество мигрировавших клеток определяли путем подсчета. Апоптоз: Условия адгезии описаны выше, за исключением того, что 12-ячеечные чашки покрывали витронектином, а 5× 105 клеток на лунку засевали в адгезионный буфер. После 30 мин инкубации при 37° С адгезионный буфер заменяли буфером, содержавшим 20 мкг/мл активного cRGD или неактивного cRAD пептида, и инкубировали в течение еще 4 ч. Прикрепившиеся клетки трипсинизировали и объединяли с открепившимися клетками в надосадочной жидкости, а затем исследовали на предмет апоптоза клеток с использованием набора для определения апоптоза Аро Alert Annexin V-FITC (Clontech). Анализ САМ: Получение и подготовка яиц к эксперименту описаны ранее (23). Опухолевые клетки засевали в 50 мкл PBS в количестве 4× 106 клеток на яйцо для DAOY и в количестве 3,5× 106 клеток на яйцо для U87MG. Клетки выращивали до опухолей в течение 7 дней, затем собирали в стерильных условиях, обрезали до одинаковых размеров и повторяли процедуру на САМ 10-дневных эмбрионов. На следующий день в вену САМ инъецировали активный или неактивный пептид (100 мкг на яйцо). Опухоли фотографировали in situ через 7 дней роста, затем собирали, взвешивали и фиксировали в 4% формалине с буфером и заливали парафином. После получения серийных срезов, препараты окрашивали гематоксилином и эозином. Модель опухоли головного мозга: Детально модель описана заявителями (31). Опухолевые клетки (106/10 мкл PBS) инъецировали интрацеребрально в указанных участках. Интраперитонеальное лечение активным cRGD или неактивным cRAD пептидом (100 мкг/ 50 мкл/мышь) начинали на 3 день после имплантации для клеток U87MG и на 10 день для клеток DAOY и повторяли ежедневно до тех пор, пока не появлялось состояние кахескии и/или агонии. Животных затем умерщвляли с использованием анестезии СO2, головной мозг удаляли и быстро замораживали или фиксировали в формалине с буфером, заливали в парафин и полученные срезы окрашивали гематоксилин-эозином. Подкожный рост опухоли: Для подкожного роста опухоли клетки (106 на мышь) инъецировали п/к под правую переднюю лапу немедленно после интрацеребральной инъекции. Исследования на животных: Исследования на животных проводили согласно руководству NIH и с разрешения местного комитета защиты животных. Эксперименты Ангиогенез на САМ Для оценки влияния антагонизма в отношении Рост опухолей также ингибировался антагонистом Фиг.1а и 1b показывают ингибирование ангиогенеза (а) и опухолевого роста на САМ (b) антагонистом Ортотопическая модель опухоли Клетки DAOY и U87MG (106 клеток на мышь) стереотаксически инъецировали в правую часть коры лобной доли бестимусных мышей с целью создания системы, которая повторяет микроокружение головного мозга. Этот способ позволял получить модель онкогенеза в головном мозге человека с хорошей воспроизводимостью и делал возможным измерение роста опухоли и выживания мышей в течение времени, предшествовавшего интродукции антагонизма в отношении С целью изучения эффекта пептида cRGD на этой модели опухоли сначала выращивали в течение 7 дней (DAOY) или 3 дней (U87MG), затем производили ежедневное и/п введение антагониста У леченных мышей сохранялся прежний вес 21 г; однако у контрольных мышей с DAYO и U87MG вес снизился до 16 г и 15 г, соответственно, что составило потерю веса по сравнению с исходным в 5-6 г (см. таблицу 1). Среднее время появления неврологических симптомов в контрольной группе составляло 2,5 недели для мышей с U87MG и 4,5 недели для мышей с DAYO, в то время как у леченных животных неврологических симптомов не было в течение всего времени наблюдения.
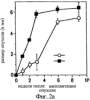
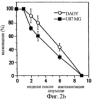
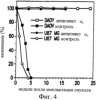
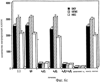
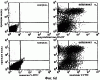
Дополнительное количество животных изучали на выживание. Выживание определяли на момент времени, в который животное необходимо было умертвить в связи с агональным состоянием. Всех контрольных животных умерщвляли до 6-недельного периода роста опухоли, и большинство животных (>50%) в обеих контрольных группах умерщвляли на 3-4 неделе (фиг.4). Фиг.4 показывает выживание мышей после ортотопической имплантации опухоли головного мозга, которые получали активный или неактивный пептид; и/п лечение (100 мкг на мышь в день) было начато на 3 день в случае клеток U87MG и на 7 день в случае клеток DAYO. Контрольные группы для DAYO и U87MG составляли по 16 животных, 32 мышей для леченых DAYO и 24 – для леченых U87MG. Фиг.4 показывает, что ни для одного из леченых животных не потребовалось умерщвление, и в настоящее время леченые мыши с DAYO выживают до 24 недель, а мыши с U87MG – 15 недель с момента инъекции опухолевых клеток. Ни у одного из леченых животных не появились неврологические симптомы, и все они продолжают нормально расти. Гетеротопическая модель опухоли Для определения влияния микроокружения на рост опухоли, подвергаемой антагонизму в отношении Для дальнейшего изучения этого феномена заявители инъецировали клетки DAYO и U87MG интракраниально и подкожно, перед началом лечения пептидами, как описано для случая ортотопической модели. Фиг.5 показывает влияние антагониста Экспрессия интегринов Витронектин представляет собой белок матрикса, который может влиять на различные биологические реакции, имеющие место в головном мозге и подкожных структурах. Он в изобилии содержится в экстраневральных областях, но в норме вырабатывается глиальной и нейрональной тканью (15, 20). Однако клетки злокачественной глиомы синтезируют витронектин, который, в свою очередь, может играть ключевую роль в их прикреплении и распространении (15). Альтернативно, витронектин может способствовать адгезии эндотелиальных клеток и их миграции по направлению к ложу опухоли, и, таким образом, способствовать ангиогенезу. Поскольку прикрепление клеток к витронектину опосредуется интегринами Фиг.6a-6d показывают профиль интегринов (а), воздействие антагониста Фиг.6a-6d показывают, что клетки U87MG экспрессируют высокие уровни Адгезия к витронектину Для того, чтобы определить, регулируют ли Миграция на витронектин Для того, чтобы установить, модулируют ли Апоптоз Астроцитомы продвинутых стадий вырабатывают витронектин на ведущем инвазивном крае, что наводит на мысль о том, что этот белок играет важную роль в прикреплении клеток опухоли головного мозга, их миграции и, возможно, выживании (15). Заявители ранее показали, что предварительная инкубация опухолевых или капиллярных клеток головного мозга с пептидом cRGD или с комбинацией анти- Влияние пентапептида на адгезию клеток опухоли головного мозга к различным субстратам ЕСМ Фиг.7 показывает воздействие циклического пентапептида на адгезию опухолевых клеток к белкам ЕСМ. Нетканевые культуральные планшеты инкубировали в течение 1 часа при 37°С с витронектином, тенасцином, фибронектином или коллагеном I (10 мкг/мл), затем промывали PBS. После промывания засевали 5× 105 клеток на лунку и инкубировали в течение 16 часов при 37°С. Культуры затем промывали и добавляли адгезионный буфер, содержавший 20 мкг/мл пентапептида или контрольного пептида, и инкубировали еще 2-24 часа. Культуры затем дважды промывали адгезионным буфером и окрашивали кристаллическим фиолетовым и определяли ОП 600. Данные представляют количество прикрепившихся клеток после 8 часов инкубации. U87 = глиобластома и DAOY = медуллобластома. Как показано на фиг.7, опухолевые клетки откреплялись от витронектина и тенасцина, прикрепление к которым было опосредовано интегринами Влияние прикрепления клеток на выживание клеток Фиг.8 показывает апоптоз клеток опухоли головного мозга и клеток капилляров головного мозга, выросших на ЕСМ и испытавших воздействие пентапептида. Условия эксперимента были такими же, как на фиг.7, за исключением того, что открепившиеся клетки и прикрепившиеся клетки объединяли после трипсинизации. Клетки затем промывали, суспендировали в 50 мкл PBS и центрифугировали на предметных стеклах (цитоспин). После фиксации 4% параформальдегидом клетки окрашивали для определения апоптоза с использованием набора Boehringer Apoptosis Kit. Результаты выражали как процент апоптотических клеток от общего количества клеток. Эксперимент проводили через 24 часа инкубации с пептидами. Влияние прикрепления клеток на выживание клеток показано на фиг.8. Как можно видеть, клетки, на которые в течение 24 часов воздействовал активный пептид и которые выросли на витронектине или тенасцине, показали возросшее количество мертвых клеток по сравнению с контрольными клетками. Напротив, пентапептид не изменял выживание клеток, выросших на коллагене I или фибронектине. Понижение выживания клеток наблюдалось для обоих типов опухолевых клеток, а также для клеток капилляров головного мозга. Сходные данные были получены, когда клетки окрашивали на ранний маркер гибели клетки, экспрессируемый на поверхности клетки (Armexin-V, данные не показаны). Таким образом, активный пентапептид индуцирует открепление и гибель клеток, как в случае клеток опухоли, так и в случае клеток капилляров головного мозга, которые прикрепляются к витронектину и тенасцину. Выработка белков ЕСМ в опухоли головного мозга человека Как указывалось выше, опухоли головного мозга человека вырабатывают витронектин и тенасцин, и полагают, что эти субстраты улучшают выживание опухолевых клеток и усиливают их инвазию. Заявители изучили выработку этих белков на своей модели опухоли головного мозга. Фиг.9 показывает иммуногистохимию опухолей головного мозга U87, ксенотрансплантированных в передний мозг бестимусных мышей, которых лечили активным (анти- Как показано на фиг.9, опухоли головного мозга действительно вырабатывали витронектин и тенасцин, и их происхождение из опухолевых клеток было доказано с помощью антител, специфичных в отношении белков человека. Введение активного пентапептида не влияло на синтез этих белков. Помимо этого, заявителям удалось также показать, что клетки опухоли головного мозга вырабатывают эти белки в культуре ткани (данные не показаны). Таким образом, мышиная модель, которую использовали заявители, имитирует ситуацию при опухолях головного мозга человека. Антиангиогенный эффект пентапептида показан на верхней части фиг.9, где заявители окрашивали срезы ткани для выявления маркера, специфичного для клеток капилляров, а именно, CD 31. В то время как опухоли, леченные контрольным пептидом, имели многочисленные сосуды, которые окрашивались по маркеру положительно, в опухолях, леченных активным пентапептидом, было очень мало таких сосудов (р<0,005), что указывает на подавление роста капилляров. Нижняя часть фиг.9 показывает окрашивание на апоптотические (мертвые) клетки. Этот способ аналогичен способу, описанному для фиг.8. Опухоли, леченные активным пентапептидом, имели достоверно больше мертвых клеток, чем опухоли, леченные контрольным пептидом (р<0,02). Фотографии были сделаны для глиобластомы U87, однако сходные данные были получены и для медуллобластомы DAOY (не показаны). Отношение гибели клеток и апоптоза опухолевых клеток Для того чтобы показать, что это увеличение гибели клеток происходит благодаря непосредственному апоптозу опухолевых клеток и не является следствием подавления роста капилляров, заявители имплантировали в головной мозг мышей клетки меланомы, которые экспрессировали (М21 L Таблица 2 показывает выживание мышей с
Как показано в таблице 2, мыши, которым трансплантировали В заключение, обсужденные выше данные поддерживают гипотезу о том, что циклический пентапептид индуцирует непосредственную гибель опухолевых клеток. Обсуждение Опухоли головного мозга являются высоко ангиогенными, и их непрерывный рост зависит от неоваскуляризации. Антиангиогенез, таким образом, является потенциально важной терапевтической стратегией, направленной против этих злокачественных новообразований. Интегрины С целью установить, можно ли получить сходные реакции на модели, которая повторяет микроокружение головного мозга, клетки опухоли головного мозга стереотаксически инъецировали в передний мозг бестимусных мышей. Клетки глиомы U87MG и медуллобластомы DAOY были выбраны для этого исследования потому, что их чувствительность к пептиду в исследовании САМ и их распространенная инвазивность и ангиогенез были ранее показаны in vivo (31, 32). На этой ортотопической модели cRGD вновь достоверно ингибировал рост опухолей головного мозга U87MG и DAOY. Контрольные мыши погибли из-за прогрессирования опухолей, и у них были обнаружены высокоинвазивные опухоли среднего размера более 3 мм3. Мыши, леченные cRGD, выживали без признаков заболеваемости, и жизнеспособные опухоли не были выявлены, кроме скудного остаточного количества клеток в месте имплантации или в малых агрегатах вдоль поверхностей желудочков и твердой мозговой оболочки. Все остаточные опухолевые очаги в группе, получавшей лечение, были менее 1 мм3 и, следовательно, не требовали реакции ангиогенеза со стороны хозяина. Альтернативные ингибиторы ангиогенеза, такие как тромбоспондин-1, TNP-470 и тробоцитарный фактор 4, также, как было показано, ингибируют рост экспериментальных опухолей головного мозга, хотя большинство этих результатов было ограничено подкожными ксенотрансплантатами или требовали прямой доставки лекарственного средства в ложе опухоли путем стереотаксической инъекции (32-34). Пептидный антагонист Представляет интерес тот факт, что опухоли DAOY и U87MG, выросшие одновременно под кожей, не показали или показали незначительный эффект лечения cRGD в исследовании заявителей. Это подчеркивает важность внеклеточного окружения в регуляции ответов опухолевая клетка хозяина-интегрин. Ангиостатин, как было установлено, равным образом ингибирует рост п/к и и/к инъецированных клеток опухоли головного мозга, что указывает, что это соединение подавляет ангиогенез посредством механизма, отличающегося от механизма действия RGD пептида (36). Одно возможное объяснение различия в ингибировании роста cRGD заключается в том, что эндотелиальные клетки подкожных структур могут по своей природе отличаться от эндотелиальных клеток микрососудистого русла ЦНС, что делает ангиогенез менее чувствительным к антагонизму в отношении интегринов. Это подтверждается недавним открытием, что у мышей, у которых отсутствует Антионкогенные эффекты пептида в исследовании заявителей, обычно более выражены по сравнению с теми, которые наблюдаются при использовании других антиангиогенных агентов. Поскольку многие опухоли также экспрессируют интегрины С целью прояснения этого вопроса заявители изучили адгезию и миграцию клеток к витронектину в присутствии антагонистов Данные заявителей показывают также, что циклический пентапептид может ингибировать адгезию клеток опухоли головного мозга к различным субстратам ЕСМ, таким как витронектин или тенасцин. Эффект такого ингибирования адгезии приводил к гибели клеток (апоптозу) как клеток опухоли головного мозга, так и клеток капилляров головного мозга. Этот эффект ограничивается витронектином и тенасцином ЕСМ. Заявители показали также, что увеличение гибели клеток происходит вследствие непосредственного апоптоза опухолевых клеток и не является следствием подавления роста капилляров. Иными словами, открытием настоящего изобретения является тот факт, что циклический пентапептид индуцирует непосредственную гибель опухолевых клеток. В целом, настоящее исследование предоставило данные о том, что направленный антагонизм в отношении интегринов, конкретно, Описанные ранее сведения предназначены для иллюстрации без ограничения объема настоящего изобретения. Разумеется, специалист может легко увидеть и разработать дополнительные варианты осуществления настоящего изобретения на основе идей, представленных в настоящем документе, без излишнего экспериментирования. ЛИТЕРАТУРА 1. Folkman, J., The role of angiogenesis in tumor growth, Sem. Cancer Biol., 3:65-71,1992. 2. Plate, К.Н., Risau, W., Angiogenesis in malignant gliomas, Glia., 15:339-347, 1995. 3. Folkman, J., Watson, К., Ingber, D., Hanahan, D., Induction of angiogenesis during the transition from hyperplasia neoplasia, Nature, 339:58-61,1989. 4. Chan, A.S., Leunng, S.Y., Wong, M.P., Yuen, S.T., Cheung, N., Fan, Y.W., Chung, L.P., Expression of vascular endothelial growth factor and its receptors in the anaplastic progression of astrocytoma, oligodendroglioma, and ependymoma, Am. J. Surg. Pathol., 22:816-826,1998. 5. Takano, S., Yoshii, Y., Kondo, S., Suzuki, H., Maruno, Т., Shirai, S., Nose, Т., Concentration of vascular endothelial growth factor in the serum and tumor tissue of brain tumor patients. Cower Res., 56:2185-2190,1996. 6. Brem, S., Tsanaclis, A.M., Gately, S., Gross, J.L., Herblin, W.F., Immunolocalization of basic fibroblast growth factor to the microvasculature of human brain tumors. Cancer, 70:2673-2680,1992. 7. Li,V.W., Folkerth, R.D., Watanabe, H., Yu, C., Rupnick, M., Barnes, P., Scott, R.M., Black, P.M., Sallan, S., Folkman, J., Microvessel count and cerebrospinal fluid basic fibroblast growth factor in children with brain tumors, Lacet, 344:82-86,1994. 8. Abulrauf, S.L, Edvardsen, К., Но K.L., Yang, X.Y., Rock, J.P., Rosenblum. M.L., Vascular endothelial growth factor expression and vascular density as prognostic markers of survival in patients with low-grade astrocytoma, J. Neurosvrg., 88:513-520, 1998. 9. Leon, S.P., Folkerth, R.D., Black, P.M., Microvessel density is a prognostic indicator for patients with astroglial brain tumors. Cancer, 77:362-372, 1996. 10. Hynes, R.O., Integrins: a family of cell surface receptor. Cell, 48:455-459, 1987. 11. Rusnati, M., Tanghetti, E., Dell’Era, P., Gualandris, A., Presta, M., Alphavbeta3 integrin mediates the cell-adhesive capacity and biological activity of basic fibroblast growth factor (FGF-2) in cultured endothelial cells, Mol. Biol. Cell, 8:2449-2461, 1997. 12. Varner, J.A., The role of vascular cell integrins alpha v beta 3 and alpha v beta 5 in angiogennesis, EXS, 79:361-390, 1997. 13. Friedlander, M., Brooks, P.C., Shaffer, R.W., Kincaid, C.M., Varner, J.A., Cheresh, D.A., Definition of two angiogenic pathways by distinct alphav integrins. Science, 270:1500-1502, 1995. 14. Gladson, G.L., Expression of integrin alpha v beta 3 in small blood vessels of glioblastoma tumors, J. Neuropathol. Exp. Neurol., 55:1143-1149, 1996. 15. Gladson, C.L., Wilcox, J.N., Sanders, L., Gillespie, G.Y., Cheresh, D.A., Cerebral microenvironment influences expression of the vitronectin gene in astrocytic tumors, J. Cell Sci., 108:947-956,1995. 16. Longhurst, C.M., Jennings, L.K., lntegrin-mediated signal transduction. Cell Mol. Life Sci., 54:514-526, 1998. 17. Howe, A., Aplin, A.E., Alahari, S.K.. Juliano, R.L., Integrin signaling and cell growth control, Curr. Opin. Cell Biol., 10:220-231, 1998. 18. Malik, R.K., Regulation of apoptosis by integrin receptors, J. Pediatr. Hematol. Oncol 19:541-545, 1997. 19. Scatena, M.. Almeida, M., Chisson, M.L., Fausto, N., Nicosia, R.F., Giachelli, C.M., NF-kappaB mediates alphavbeta3 integrin-induced endothelial cell survival, J. Cell Biol., 141-1083-1093, 1998. 20. Gladson, C.L., Cheresh, D.A., Glioblastoma expression of vitronectin and the avb3 integrin. Adhesion mechanism for transformed glial cells, J. Clin. Invest., 88:1924-1932, 1991. 21. Brooks, P.C., Clark, R.A., Cheresh, D.A., Requirement of vascular integrin alpha v beta 3 for angiogenesis. Science, 264:569-571, 1994. 22. Christofidou-Solomidou, M., Bridges, M., Murphy, G.F., Abelda, S.M, DeLisser, HM., Expression and function of endothelial cell av integrin receptors in wound-induced human angiogenesis in human skin/SCID mice chimeras. Am. J.Pathol., 151:975-983, 1997. 23. Brooks, P.C., Montgomery, A.M., Rosenfeld, M., Reisfeld, R.A., Нu, Т.. Klier, G., Cheresh, D.A., Integrin avb3 antagonists promote tumor regression by inducing apoptosis of angiogenic blood vessels, Cell, 79:1157-1164, 1994. 24. Molema, G., Griffioen, A.W., Rocking the foundations of solid tumor growth by attacking the tumor’s blood supply, Immunol. Today, 19:392-394, 1998. 25. Kim, K.J., Li, В., Winer, J., Armanini, M., Gillett, N., Phillips, H.S., Ferrara, N., Inhibition of vascular endothelial growth factor-induced angiogenesis suppresses tumor growth in vivo, Nature, 362-841-844, 1993. 26. Re, F., Zanetti, A., Sironi, M., Polentarutti, N., Lanfrancone, L., Dejana, E., Colotta, F., Inhibition of anchorage-dependent cell spreading triggers apoptosis in cultured human endothelial cells, J. Cell Biol., 127:537-546, 1994. 27. Montgomery, А.М., Reisfffeld, R.A., Cheresh, D.A., Integrin avb3 rescues melanoma cells. from apoptosis in three-dimensional dermal collagen, Proc. Natl. Acad. Sci., 91:8856-8860, 1994. 28. Isik, F.F., Gibran, N.S., Jang. Y., Sandell, L., Schwartz, S.M., Vitronectin decreases microvascular endothelial cell apoptosis, J. Cell Physiol., 173:149-155, 1998. 29. Germer, M., Kanse, S.M., Kirkegaard, Т., Kioler, L., Feldinng-Habermann, В., Goodman, S., Preissner, Т., Kinetic analysis of integrin-dependent cell adhesion on vitronectin. The inhibitory potential of plasminogen activator inhibitor-1 and RGD peptides, Eur. J. Biochem., 253:669-674, 1998. 30. Ruoslahti, E., Reed, J.C., Anchorage dependence, integrins, and apoptosis. Cell, 77:477-478, 1994. 31. MacDonald, T.J., Tabrizi, P., Shimada, H., Zlokovic, B.V., Laug, W.E., Detection of brain tumor invasion and micrometastasis in vivo by expression of enhanced green fluorescent protein, Neurosurgery, 43:1437-1443, 1998. 32. Hsu, S.C., Volpert, O.V., Steck, P.A., Mikkelsen, Т., Polverini, P.J., Rao, S., Chou Bouck, N.P., Inhibition of angiogenesis in human glioblastomas by chromosome 10 induction of throbospondin-1, Cancer Res., 56:5684-5691, 1996. 33. Таki, Т., Ohnishi, Т., Arita, N., Hiraga, S., Saitoh, Y., Izumoto, S., Mori, K., Hayakawa, Т., Anti-proliferative effects of TNnP-470 on human malignant glioma in vivo: potent inhibition of tumor angiogenesis, J. Neurooncoi, 19:251-258, 1994. 34. Tanaka, Т., Manome, Y., Wen, P., Kufe, D.W., Fine, H.A., Viral vector-mediated transduction of a modified platelet factor 4 cDNA inhibits angiogenesis and tumor growth, Nat. Med., 3:437-442, 1997. 35. Carron, C.P., Meyer, D.M., Pegg, J.A., Engleman, V.W., Nickols, M.A., Settlel, S.L., Westlin, W.F., Ruminski, P.G., Nickols, G.A., A peptidomimetic antagonist of the integrin avb3 inhibits Leydig cell tumor growth and the development of hypercalcemia of malignancy. Cancer Res., 58:1930-1935, 1998. 36. Kirsch, M., Strasser, J., Allende, R., Bello, L., Zhang, J., Black, P.M., Angiostatin suppresses malignant glioma growth in vivo, Cancer Res., 58:4654-4659, 1998. 37. Bader, B.L., Rayburn, H., Crowley, D., Hynes, R.O., Extensive vasculogenesis, angiogenesis, and organogenesis precede lethality in mice lacking all av integrins. Cell, 95:507-519, 1998. 38. Knott, J.C., Mahesparan, R., Garcia-Cabrera, I., Bolge, T.B., Edvardsen, K., Ness, G.O., Mark, S., Lund-Johansen, M., Bjerkvig, R., Stimulation of extracellular matric components in the normal brain by invading glioma cells, Int. J. Cancer, 75:864-872, 1998. 39. Seger, D., Gechtman, Z., Shaltiel, S., Phosphorylation of vitronectin by casein kinase II. Identification of the sites and their promotion of cell adhesion and spreading, J. Biol. Chem., 273:24805-24813, 1998. 40. Woodard, A., Garcia-Cardera, G., Leong, M., Madri, J., Sessa, W., Languino, L., The synergistic activity of alphavb3 integrin and PDGF receptor increases cell migration, J. Cell Sci., 111:469-478, 1998. 41. Sanders, R.J., Mainiero, F., Giancotti, F.G., The role of integrins in tumorigenesis and metastasis. Cancer Invest., 16:329-344, 1998. 42. Treasurywala, S., Berens, M.E., Migration arrest in glioma cells is dependent on the alpha-v integrin subunit, Glia., 24:236-243, 1998. 43. Won, N.C., Mueller, B.M., Barbas, C.F., Ruminski, P., Quaranta, V., Lin, B.C., Smith, J.W., Alpha-v integrins mediate adhesion and migration of breast carcinoma cell lines, Clin. Exp. Metastasis, 16:50-61, 1998. 44. Van Leeuwen, R.L., Yoshinaga, I.G., Akasaka, Т., Dekker, S.K., Vermeer, BJ., Byers, Н.R., Attachment, spreading and migration of melanoma cells on vitronectin. The role of alpha v beta 3 and alpha v beta 5 integrins, Exp. Dermatol., 5:308-315, 1996. 45. Lehmann, M., El Battari, A., Abadie, В., Martin, J.M., Marvallldi, J., Role of alpha v beta 5 and alpha v beta 6 integrin glycosylation in the adhesion of a colonic adenocarcinoma cell line (HT29-D4), J. Cell Biochem., 61:266-277, 1996. 46. Kost, С., Benner, К., Stockmann, A., Under, D., Preissner, K.T., Limited plasmin proteolysis of vitronectin. Characterization of the adhesion protein as morpho-regulatory and angiostarin binding factor. Еur. J. Biochem., 236:682-688, 1996. 47. Maenpaa, A., Kovanen, P.E., Paetau, A., Jaaskelainen, J., Timonen, Т., Lymphocyte adhesion molecule ligands and extracellular matrix proteins in gliomas and normal brain: expression of VCAM-1 in gliomas, Acta. Neuropathol., 94:216-225, 1997. 48. Brooks, P.C., Stromblad, S., Klemke, R., Visscher, D., Sarkar, F.H., Cheresh, D.A., Anti-integrin alpha v beta 3 blocks human breast cancer growth and angiogenesis in human skin, J. Clin. Invest., 96:1815-1822, 1995. 49. Stins, M.F., Gillesl, F., Kim, K.S., Selective expression of adhesion molecules on human brain microvascular endothelial cells, J. Neuroimmunol., 76:81-90, 1997. 50. Bafetti, L.M., Young, T.N., Itoh, Y., Stack, M.S., Intact vitronectin induces matrix metalloproteinase-2 and tissue inhibitor of metalloproteinase-2 expression and enhanced cellular invasion by melanoma cells, J. Biol. Chem., 273:143-149, 1998. 51. Wong, N.C., Mueller, B.M., Barbas, C.F., Ruminski, P., Quaranta, V., Lin, E.C., Smith, J.W., Alphav integrins mediate adhesion and migration of breast carcinoma cell lines, Clin. Exp. Metastasis, 16:50-61, 1998. 52. Van Leeuwen, R.L., Yoshinaga, I.G., Akasaka, Т., Dekker, S.K., Vermeer, B.J., Byers, H.R., Attachment, spreading and migration of melanoma cells on vitronectin. The role of alpha v beta 3 and alpha v beta 5 integrins, Exp. Dermatol., 5:308-315, 1996. 53. Lehmann, M., El Battari, A., Abadie, В., Martin, J.M., Marvaldi, J., Role of alpha v beta 5 and alpha v beta 6 integrin glycosylation in the adhesion of a colonic adenocarcinoma cell line (HT29-D4), J. Cell Biochem., 61:266-277, 1996. 54. Gladson, C.L., Cheresh, D-A., Glioblastoma expression of vitronectin and the integrin, J. Clin. Invest., 88:1924-1932, 1991. 55. Jaskiewicz, К., Chasen, M.R., Robson, S.C., Differential expression of extracellular matrix proteins and integrins in hepatocellular carcinoma and chronic liver disease, Anticancer Res., 13:2229-2237, 1993. 56. Brooks, P.C., Stromblad, S., Klemke, R., Visseher, D., Sarkar, F.H., Cheresh, D.A., Anti-integrin alpha v beta 3 blocks human breast cancer growth and angiogenesis in human skin, J. Clin. Invest., 96:1815-1822, 1995. 57. Leon, S.P., Folkerth, RD., Black, P.M., Microvessel density is a prognostic indicator for patients with astroglial brain tumors, Cancer, 77:362-372, 1996. 58. Wesseling, P., Van Der Laak, J.A., Link, M., Teepen, H.L., Ruiter, D.J., Quantitative analysis ofmicrovascular changes in diffuse astrocytic neoplasms with increasing grade of malignancy. Hum. Pathol; 29:352-358, 1998. 59. Li, V.W., Folkerth, R.D., Watanabe, H., Yu, C., Rupnick, M., Bames, P., Scott, M., Black, P.M., Sallan, S., Folkman, J., Microvessel count and cerebrospinal fluid basic fibroblast growth factor in children with brain tumors. Lancet, 344:82-86, 1994. 60. Taki, Т., Ohnishi, Т., Arita, N.. Hiraga, S., Saitoh, Y., Izumoto, S., Mori, K., Hayakawa, Т., Anti-proliferative effects of TNP-470 on human malignant glioma in vivo: potent inhibition of tumor angiogenesis, J. Neurooncol, 19:251-258, 1994. 61. Hsu, S.C., Volpert, O.V., Steck, P.A., Mikkelsen, Т., Polverini, P.J., Rao, S., Chou Bouck, N.P., Inhibition of angiogenesis in human glioblastomas by chromosome 10 induction of thrombospondin-1. Cancer Res., 56:5684-5691, 1996. 62. Tanaka, Т., Manome, Y., Wen, P., Kufe, D.W., Fine, H.A., Viral vector-mediated transduction of a modified platelet factor 4 CDNA inhibits angiogenesis and tumor growth, Nat. Med., 3:437-442, 1997.
Формула изобретения
Способ индуцирования апоптоза клеток опухоли головного мозга, растущих в головном мозге хозяина, включающий введение хозяину терапевтически эффективного количества антагониста интегрина
РИСУНКИ
|
||||||||||||||||||||||||||||||||||||||||
 v, представляющего собой цикло(Arg-Gly-Asp-D-Phe-[N-Me]-Val). Способ позволяет значительно ингибировать онкогенез в головном мозге in vivo, в том числе независимо от антиангиогенеза, за счет индукции непосредственной гибели опухолевых клеток. 9 ил., 2 табл.
v, представляющего собой цикло(Arg-Gly-Asp-D-Phe-[N-Me]-Val). Способ позволяет значительно ингибировать онкогенез в головном мозге in vivo, в том числе независимо от антиангиогенеза, за счет индукции непосредственной гибели опухолевых клеток. 9 ил., 2 табл. 3 and av
3 and av