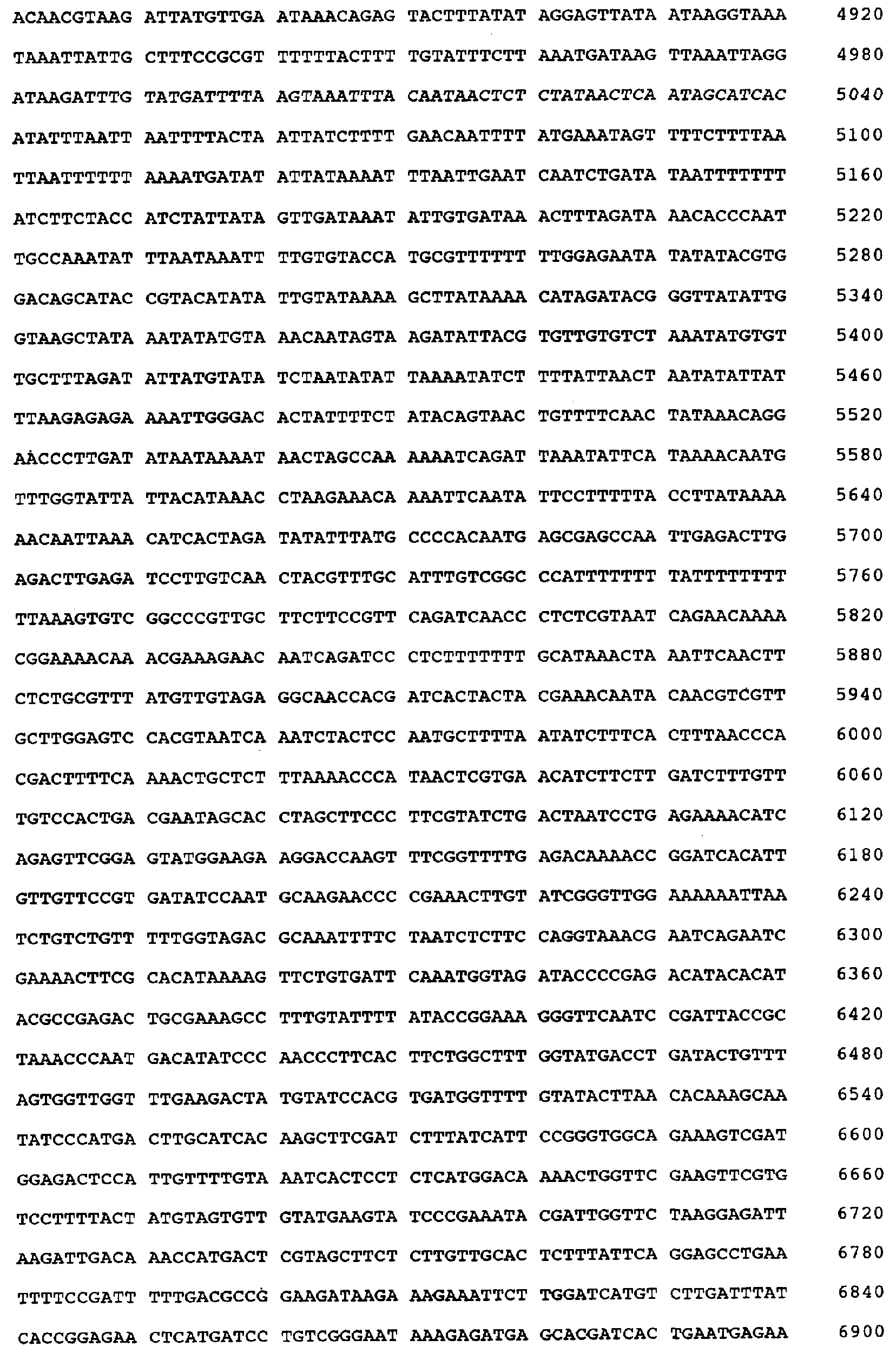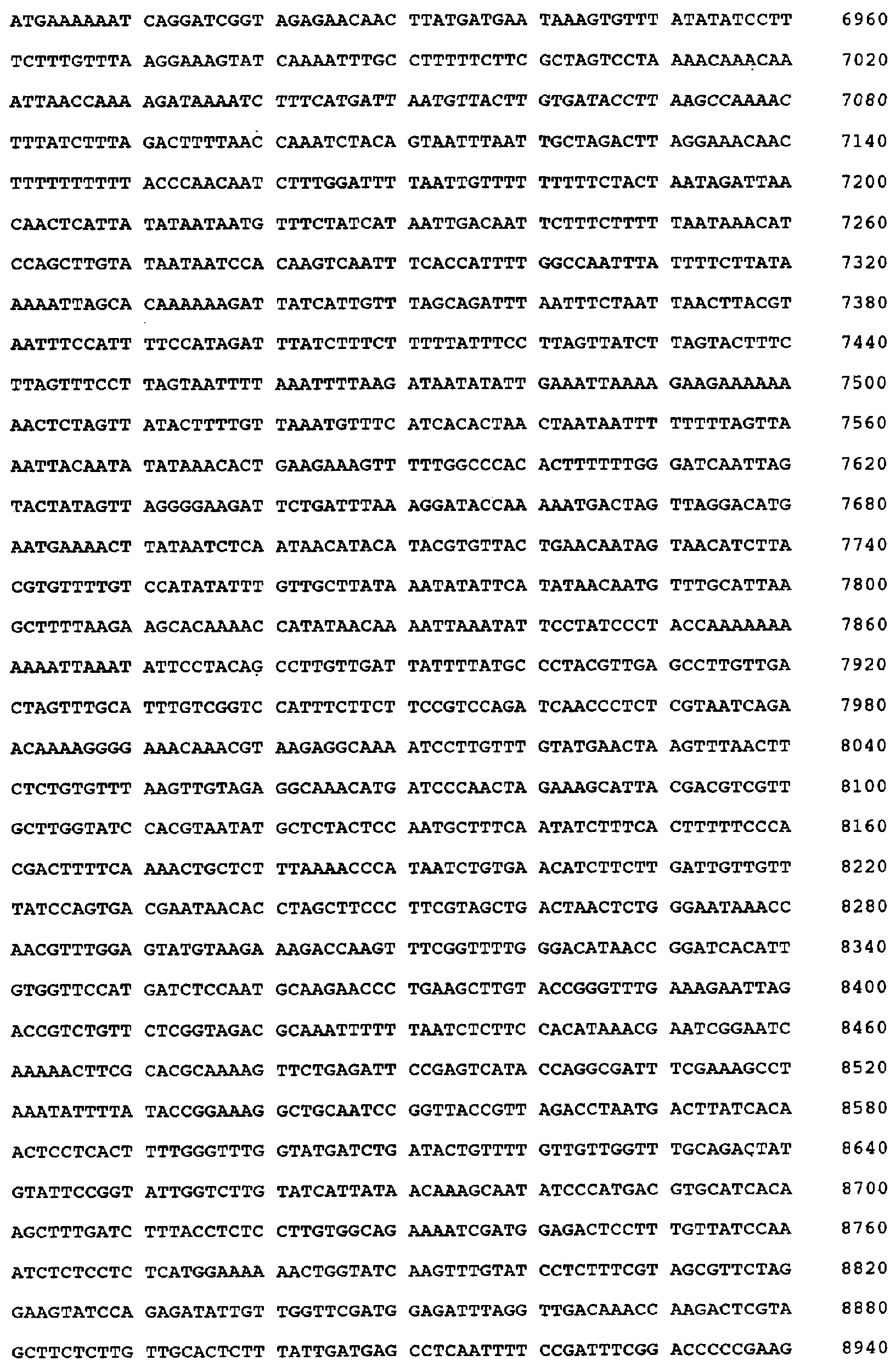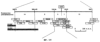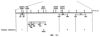Патент на изобретение №2252960
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
(54) ГЕН, ОБУСЛОВЛИВАЮЩИЙ УСТОЙЧИВОСТЬ РАСТЕНИЙ К БОЛЕЗНЯМ, И ЕГО ПРИМЕНЕНИЕ
(57) Реферат:
Изобретение относится к биотехнологии, в частности к выделенной молекуле ДНК, обеспечивающей устойчивость растений к болезням, а также к способу придания растениям устойчивости к болезням. Выделяют молекулу ДНК, которая включает ген NIM1. Конструируют рекомбинантный вектор, включающий активный в растении промотор, функционально связанный с вышеуказанной молекулой ДНК. Проводят трансформацию растения данным вектором. Изобретение позволяет снизить производственные затраты и повысить сбор сельскохозяйственной продукции. 3 с. и 6 з.п. ф-лы, 19 ил., 18 табл.
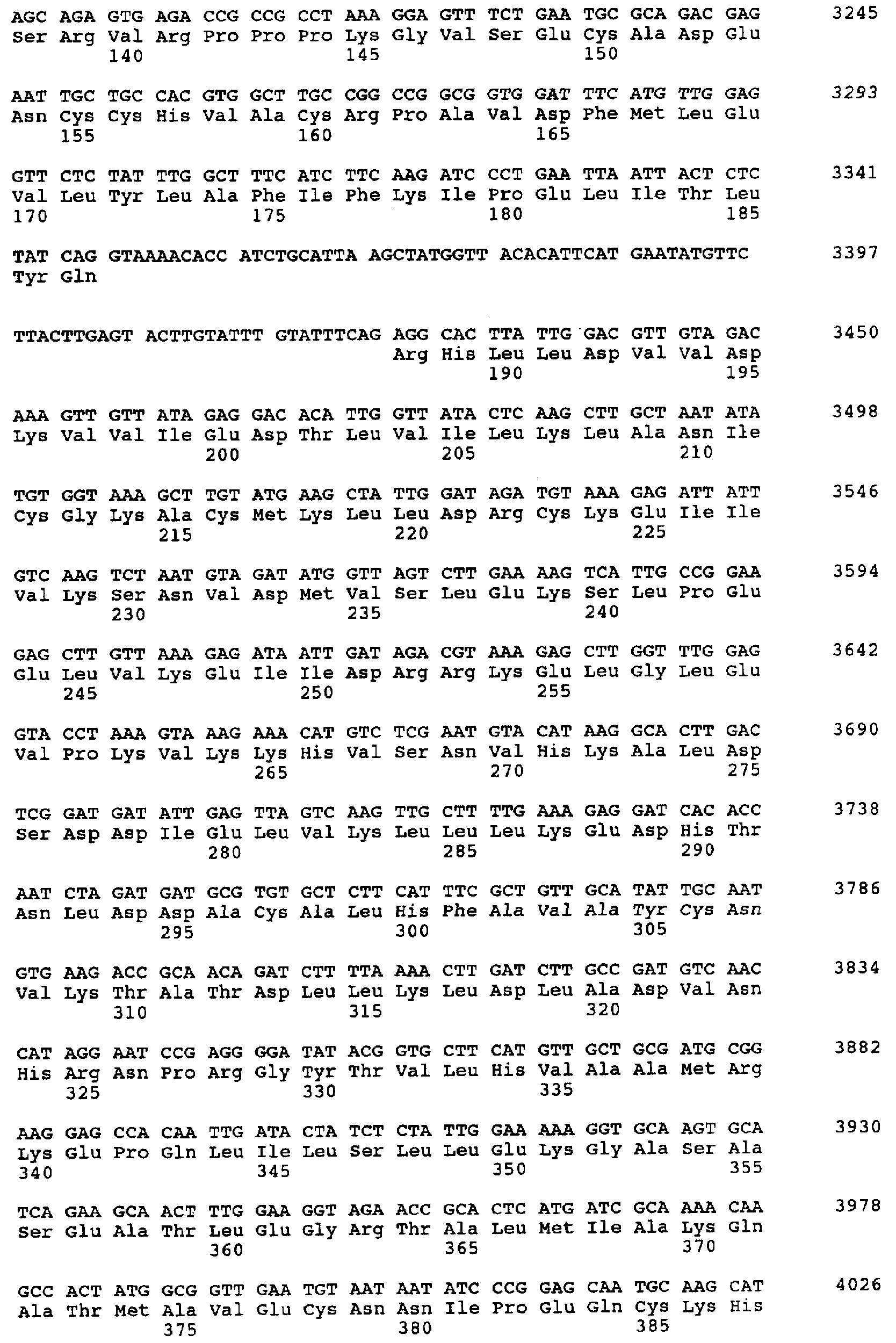
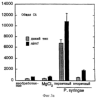
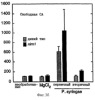
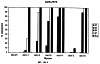
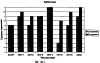
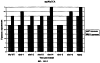
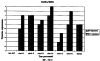
Настоящее изобретение относится к устойчивости растений к болезням и к выявлению и селекции растений, устойчивых к болезням. В частности, настоящее изобретение относится к выявлению, выделению и характеристике гена, обусловливающего, устойчивость растений к широкому спектру болезней. Растения постоянно подвергаются воздействию широкого спектра патогенных организмов, к которым относятся вирусы, бактерии, грибы и нематоды. Культурные растения являются особенно уязвимыми, поскольку их обычно выращивают в виде генетически однородных монокультур, при этом потери при поражении болезнью могут быть серьезными. Однако большинство растений имеют собственные врожденные механизмы защиты от патогенных организмов. Естественная изменчивость по признаку устойчивости к фитопатогенам была выявлелена селекционерами растений и фитопатологами и использована для селекции многих культурных растений. Эти естественные гены устойчивости к болезням часто обусловливают высокие уровени устойчивости или иммунитета по отношению к патогенам. У многих видов растений первоначальная инокуляция патогеном, вызывающим омертвение, может иммунизировать растения в отношении последующего заражения. Эта приобретенная устойчивость к болезням впервые описана в 1901 году и, вероятно, играет важную роль в сохранении растений в природе. Особенно полно описанными примерами иммунитета растений являются феномен системной приобретенной устойчивости (SAR от англ. systemic acquired resistance) и индуцированная устойчивость у таких растений, как табак, Arabidopsis и огурец. В этих системах инокуляция патогеном, вызывающим омертвение, приводит к системной защите от последующих заражений этим патогеном, а также целым рядом других важных с сельскохозяйственной точки зрения бактерий, грибов и вирусных патогенов. Системная приобретенная устойчивость также может вызываться химическим иммунизирующими соединениями, определенными химическими веществами, которые вызывают иммунный ответ у растений. Такие соединения могут быть естественного происхождения, например, салициловая кислота (CK), или могут представлять собой синтезированные химические вещества, такие, как 2,6-дихлоризоникотиновая кислота (ИНК) и S-метиловый эфир бензо(1,2,3)тиадиазол-7-карботионовой кислоты (БТК). Обработка патогеном или иммунизирующим соединением индуцирует экспрессию по крайней мере девяти наборов генов у табака, наиболее хорошо охарактеризованного вида растений. В других растениях могут экспрессироваться различные количества и типы генов. Уровень индукции связанных с SAR генов, вызванный иммунизирующими соединениями, более чем в 10000 раз превышает основной уровень. В частности, SAR характеризуется экспрессией SAR-генов, в том числе генов, связанных с патогенезом (PR от англ. pathogenesis-related). SAR-гены индицируются после заражения патогеном. Некоторые из этих генов играют определенную роль в придании растениям системной приобретенной устойчивости. Эти растительные протеины индуцируются в значительных количествах в ответ на заражение различными патогенами, включая вирусы, бактерии и грибы. PR-протеины первоначально были открыты в растениях табака (Nicotiana tabacum), отличающихся гиперчувствительностью к заражению вирусом мозаики табака (TMV). Затем PR-протеины были обнаружены у многих видов растений (см. Redolfi и др. (1983), Neth J. Plant Pathol 89: 245-254; Van Loon (1985), Plant Mol. Biol. 4: 111-116; Uknes и др., (1992) Plant Cell 4: 645-656). Такие протеины, вероятно, являются общей защитной системной реакцией растений на заражение патогенами. Связанные с патогенезом протеины включают протеины SAR8.2a и SAR8.2b, кислые и основные формы протеинов табака PR-1a, PR-1b и PR-1c, основные протеины PR-1’, PR-2, PR-2’, PR-2’’, PR-N, PR-O, PR-O’, PR-4, PR-P, PR-Q, PR-S и PR-R, пероксидазы огурца, основную пероксидазу огурца, хитиназу, которая является основной копией PR-P или PR-Q, бета-1,3-глюканазу (глюкан-эндо-1,3-бета-глюкозидаза, КФ 3.2.1.39), которая является основной копией PR-2, PR-N или PR-O, и индуцируемую патогеном хитиназу огурца. Такие PR-протеины описаны, например, у Uknes и др. (1992) в The Plant Cell 4: 645-656 и в указанных в этой статье ссылках. SAR- или SAR-подобные гены экспрессируются во всех видах растений, обладающих системной приобретенной устойчивостью. Экспрессия таких генов может быть выявлена зондированием с использованием известных последовательностей ДНК гена SAR, что описано, например, у Lawton и др. (1992), Proceedings of the Second European Federation of Plant Pathology (1983), В: Mechanisms of Defence Responses in Plants, под ред. В.Fritig и М.Legrand, Kluwer Academic Publishers, Dordrecht, стр.410-420; у Uknes и др. (1992), The Plant Cell 4: 645-656; у Ward и др., (1991) The Plant Cell 3: 1085-1094. Методы гибридизации и клонирования хорошо известны в данной области и описаны, например, в Molecular Cloning, A Laboratory Manual, 2-е изд., тома 1-3, под ред. Sambrook и др., Cold Spring Harbor Laboratory Press (1989), и в указанных в данной работе ссылках. Альтернативно этому SAR- или SAR-подобные гены могут быть обнаружны другими методами, такими, как скрининг протеинов, ±-скрининг (дифференциальный скрининг) и т.д, как описано, например, у Liang и Pardee (1992), Science 257: 967-971; у St. John и Davis (1979), Cell 16: 443. Несмотря на многочисленные исследования и применение сложных и обширных мероприятий, направленных на защиту урожая, включая генетическую трансформацию растений, потери от болезней ежегодно продолжают составлять миллионы долларов. Гены устойчивости к болезням клонировали ранее, однако трансгенные растения, трансформированные такими генами, как правило, обладали устойчивостью только к некоторым штаммам определенных видов патогенов. Несмотря на попытки клонировать гены, относящиеся к SAR, ген, контролирующий устойчивость к широкому спектру болезней, ранее не был выделен и охарактеризован. Некоторые независимые данные свидетельствует о том, что полученная эндогенно салициловая кислота (СК) принимает участие в пути трансдукции сигнала, объединяя восприятие заражения патогеном с началом развития системной приобретенной устойчивости. Мутанты, сохраняющие способность накапливать СК в ответ на заражение патогеном, но уже потерявшие способность индуцировать гены SAR или устойчивость после обработки СК или ИНК, были описаны у Delaney и др., Proc. Natl. Acad. Sci. 92: 6602-6606 (1995) и в заявке WO 94/16077, в полном объеме включенной в настоящее описание в качестве ссылки. Было обнаружено, что эти мутанты содержат мутантный ген, и этот ген в своей дикой форме контролирует экспрессию гена SAR и само явление SAR. Согласно изобретению было установлено, что мутантный ген придает мутантным растениям чувствительность к широкому спектру болезней и обусловливает отсутствие у них способности индуцироваться патогенами и химическими индукторами. Настоящее изобретение относится к идентификации, выделению и характеристике гена дикого типа (NIM1), т.е. гена, который придает растениям возможность активировать SAR и экспрессию гена SAR в ответ на биологические и химические индукторы. Мутантный ген был выявлен в подвергнутых мутагенезу растениях Arabidopsis. Было обнаружено, что эти растения имеют дефект по сравнению с их нормальной реакцией на заражение патогеном в том отношении, что они не экспрессируют гены, связанные с системной приобретенной устойчивостью (SAR) и не способны проявлять такую устойчивость. Эти мутанты содержат дефектный ген, который обозначен как nim1 (ген, отвечающий за неиндуцируемость иммунитета). Настоящее изобретение также относится к применению клонированного гена NIM1 и его вариантов для создания трансгенных растений, обладающих устойчивостью к широкому спектру болезней, и к полученным таким образом трансгенным растениям. Изобретение также относится к применению клонированного гена NIM1 и его вариантов для скрининга с целью выявления соединений, способных индуцировать у растений устойчивость к широкому спектру болезней. Краткое описание чертежей На фиг.1 показано воздействие химических индукторов на индукцию экспрессии гена PR в растениях дикого типа и в nim1-растениях. На фиг.2 показана экспрессия гена PR-1 в зараженных патогеном Ws-O-растениях и в nim1-растениях через 6 дней после начала заражения. На фиг.3 показаны уровни накопления СК в Ws-О-растениях и в nim1-растениях, зараженных Р.syringae. На фиг.4 приведена генетическая карта NIM1-области, полученная с помощью методов ПДАФ и ПДПП. На фиг.5 приведена физическая карта NIM1-области, полученная с помощью анализа клонов YAC. На фиг.6 приведена физическая карта удлиненного набора последовательностей фрагментов (contig) P1/ВАС. На фиг.7 приведена физическая карта, показывающая положения клонов Р1 и ВАС относительно фланкирующих ПДАФ-маркеров и YAC. На фиг.8 приведена физическая карта еще одного удлиненного набора последовательностей фрагментов Р1/ВАС, содержащего ген NIM1. На фиг.9 приведена объединенная генетическая и тонкая физическая карты NIM-области. На фиг.10 приведена карта NIM-области. На фиг.11 приведена объединенная карта NIM-области, включающая новые ПДАФ-маркеры. На фиг.12 представлено схематическое изображение рекомбинантов D169 и С105. На фиг.13 представлена общая карта хромосомной области вокруг NIM1 с выявленными рекомбинантами, включая ВАС, YAC и космиды в NIM1-области. На фиг.14 представлена последовательность области длиной 9,9 т.п.н. клона ВАС-04, содержащая ген NIM1. (На фиг.15 приведена нуклеотидная последовательность гена NIM1 и аминокислотная последовательность продукта гена NIМ1, включая изменения в различных аллелях.. На фиг.16 показана экспрессия NIM1, индуцируемая ИНК, БТК, СК и патогеном, в аллелях дикого типа и в мутантных аллелях nim1. На фиг.17 показана экспрессия PR-1 в растениях, имеющих мутацию nim1, и в растениях дикого типа. На фиг.18 показан уровень устойчивости к болезням у различных nim1-мутантов. На фиг.19 приведена аминокислотная последовательность, включающая экспрессируемую последовательность Tag-областей протеина NIM1 и кДНК протеиновых продуктов последовательностей 4-х генов риса (см. SEQ ID NO: 3). Определения
Ген NIM1 был клонирован методами картирования и “прогулки” вдоль хромосомы, которые показали, что ген входит в область длиной 105 т.п.н. (см. фиг.13 и таблицу 16). Эта область ограничена L84.6b-маркером слева и L84.T2-маркером справа. Только три перекрывающиеся космиды, полученные из ДНК дикого типа из области длиной 105 т.п.н., восстанавливают (комплементируют) мутантный фенотип; nim1 (фиг.13 и таблица 16). Эти три космиды перекрываются только в области длиной 9,9 т.п.н., обозначенной левым концом космидного клона D7 и правым концом космидного клона D5, как показано на фиг.13. Многие другие космиды, сконструированные на основе других участков области длиной 105 т.п.н., не восстанавливают фенотип nim1 (фиг.13 и таблица 16). Практически полноразмерный клон кДНК гена NIM1 позволяет выявить интрон-экзонные пограничные последовательности и определить аминокислотную последовательность генного продукта. Только NIM1-область гена внутри комплементарной области длиной 9,9 t.ii.il имеет изменения последовательности в различных мутантных аллелях nim1 (таблица 18). Обнаружено, что три других потенциальных области гена не имеют изменений последовательности, которые связаны с фенотипом nim1. Изменения последовательностей, обнаруженные в NIM1-области гена, согласуются с измененной функцией или с потерей функции генного продукта. Серьезность изменения в NIM1-области гена в определенном мутантном аллеле приблизительно коррелирует с наблюдаемой физиологической ролью этого nim1-аллеля. Только NIM1-область гена имела выявляемую РНК (транскрипция), и в этой РНК обнаружены многочисленные изменения, согласующиеся с физиологической ролью NIM1 в патогенезе (таблица 18 и фиг.16). Настоящее изобретение относится к выделенному генному фрагменту, гену NIM1, который представляет собой ключевой компонент пути метаболизма системной приобретенной устойчивости (SAR) в растениях. Ген NIM1 связан с активацией SAR химическими и биологическими индукторами и в сочетании с такими индукторами необходим для SAR и экспрессии SAR-генов. Локализацию гена NIM определяют с помощью анализа методами молекулярной биологии генома мутантных растений, для которых известно, что они несут мутантный ген nim1, который придает растениям-хозяевам очень высокую чувствительность к широкому диапазону патогенов и лишает их способности реагировать на патогены и химические индукторы SAR. nim1-Мутанты пригодны в качестве “универсальных чувствительных к болезням растений” (УЧБ)/благодаря их чувствительности ко многим штаммам и патотипам патогенов, растения-хозяина, а также к патогенам, которые в норме не заражают растение-хозяина, но которые заражают других хозяев. Они могут быть получены путем обработки семян или другого биологического материала мутагенными агентами с последующей селекцией потомства растений в отношении УЧБ-фенотипа путем обработки растений потомства известными химическими индукторами (например, ИНК) системной приобретенной реакции и последующего заражения растений известным патогеном. При таких обстоятельствах у мутантов с неиндуцируемой устойчивостью развиваются серьезные симптомы болезни, в то время как у немутантных растений химическое соединение индуцирует системную приобретенную устойчивость. nim1-Мутанты могут быть отобраны как из мутантных популяций, полученных в результате химического или радиоционного мутагенеза, так и из популяций, образованных с помощью инсерций (вставок) Т-ДНК и транспозониндуцируемого мутагенеза. Методы получения мутантных линий растений хорошо известны в данной области. min-Фенотип растений применяют в качестве инструмента для идентификации выделенного фрагмента гена, который придает растениям способность экспрессировать устойчивость к широкому спектру болезней. Настоящее изобретение относится к выделенной молекуле ДНК, включающей мутантный ген NIМ1, который представляет собой ген nim1. При использовании мутанта nim1 или растения для выделения гена NIM1 дикого типа, необходимого для конститутивной экспрессии генов SAR, признаки устойчивости в комбинации с другими характеристиками, важными для продуктивности и качества, могут быть включены в линии растений с помощью селекции. Подходы и методы селекции известны в данной области и описаны, например, в следующих публикациях: Welsh J.R., Fundamentals of Plant Genetics and Breeding, John Wiley & Sons, NY (1981); Crop Breeding, под ред. Wood D.R., American Society of Agronomy Madison, Wisconsin (1983); Mayo O., The Theory of Plant Breeding, 2-е изд., Clarendon Press, Oxford (1987); Singh, D.P., Breeding for Resistance to Diseases and Insect Pests, Springer-Verlag, NY (1986); Wricke и Weber, Quantitative Genetics and Selection Plant Breeding, Walter de Gruyter and Co., Berlin (1986). Еще одним объектом изобретения является химерный ген, включающий активный в растении промотор, функционально связанный с гетерологичной молекулой ДНК, кодирующей аминокислотную последовательность продукта гена NIM1 и его варианты по изобретению. Методологии конструирования растительных кассет экспрессии, а также интродукции чужеродной ДНК в растения в целом описаны в данной области техники. Как правило, для интродукции в растения чужеродной ДНК использовали Ti-плазмидные векторы для доставки чужеродной ДНК. Для такой доставки также применялось непосредственное введение ДНК, липосомы, электропорация, микроинъекция и микроснаряды. Такие методы описаны в данной области, например, в следующих публикациях: Bilang и др. (1991), Gene 100: 247-250; Scheid и др., (1991) Mol. Gen. Genet. 228: 104-112; Guerche и др., (1987) Plant Science 52: 111-116; Neuhause и др., (1987) Theor. Appl. Genet. 75: 30-36; Klein и др., (1987) Nature 327: 70-73; Howell и др., (1980) Science 208: 1265; Horsch и др., (1985) Science 227: 1229-1231; DeBlock и др., (1989) Plant Physiology 91: 694-701; Methods for Plant Molecular Biology (под ред. Weissbach и Weissbach) Academic Press, Inc. (1988); Methods in Plant Molecular Biology (под ред. Schuler и Zielinski) Academic Press, Inc. (1989), а также в заявке на патент США 08/438666, поданной 10 мая 1995, и в заявке WO 93/07278, причем обе эти публикации в полном объеме включены в настоящее описание в качестве ссылки. Следует отметить, что метод трансформации должен зависеть от растительной клетки, подлежащей трансформации. Также очевидно, что компоненты кассеты экспрессии могут быть модифицированы с целью усиления экспрессии. Например, могут применяться усеченные последовательности, нуклеотидные замещения или другие модификации. Трансформированные растительные клетки с такими модифицированными системами экспрессии далее должны проявлять сверхэкспрессию или конститутивную экспрессию генов SAR, что необходимо для активации SAR. Молекула ДНК или фрагмент гена, обусловливающие у растений устойчивость к болезням благодаря тому, что они обеспечивают экспрессию гена SAR,) могут быть включены в растительные или бактериальные клетки с использованием общепринятого метода рекомбинатной ДНК. Как правило, такая технология предусматривает встраивание (инсерцию) молекулы ДНК в систему экспрессии, для которой молекула ДНК является гетерологичной (т.е. в которой она в норме не присутствует). Гетерологичную молекулу ДНК встраивают в систему экспрессии или в вектор в соответствующей ориентации и в правильной рамке считывания. Вектор содержит необходимые элементы для транскрипции и трансляции встроенных последовательностей, кодирующих протеины. Может использоваться большое количество векторных систем, известных в данной области, таких, как плазмиды, вирусные бактериофаги и другие модифицированные вирусы. Пригодные векторы включают, но не ограничены ими, вирусные векторы, такие, как векторные системы лямбда Igti1, Igt10 и Charon 4, плазмидные векторы, такие, как pBI121, pBR322, pACYC177, pACYC184, серии pAR. pKK223-3, pUC8, pUC9, pUC18, pUC19, pLG339, pRK290, pKC37, pKC101, pCDNAII, и другие аналогичные системы. Последовательности ДНК могут быть клонированы в векторе с использованием стандартных в данной области методов клонирования, описанных у Maniatis и др. в Molecular Cloning: A Laboratory Manual, Cold Spring Laboratory, Cold Spring Harbor, New York (1982). Еще одним объектом изобретения является рекомбинантный вектор, включающий химерный ген по изобретению. С целью получения эффективной экспрессии гена или фрагмента гена по настоящему изобретению в векторе экспрессии должен присутствовать промотор. РНК-полимераза в норме связывается с промотором и инициирует транскрипцию гена. Промоторы варьируются по силе, т.е. по способности обеспечивать транскрипцию. В зависимости от используемой системы клетки-хозяина может применяться любой из многочисленных приемлемых промоторов. Приемлемые промоторы включают промотор убикитина, nos-промотор, промотор гена малой субъединицы рибулозобифосфаткарбоксилазы, промотор малой субъединицы связывающего хлорофилл А/В-полипептида, промотор 35S вируса мозаики цветной капусты и промоторы, выделенные из генов растений, При этом можно назвать, например, статью Vallejos и др., “Localization in the Tomato Genome of DNA Restriction Fragments Containing Sequences Homologous to the RRNA (45S), the major chlorophyll A/BBinding Polypeptide and the Ribulose Bisphosphate Carboxylase Genes,” Genetics 112: 93-105 (1986), в которой описан состав малых субъединиц. Промотор nos и промотор 35S вируса мозаики цветной капусты хорошо известны в данной области. Когда ген устойчивости к болезням по настоящему изобретению уже клонирован в системе экспрессии, им легко трансформировать растительную клетку. Ткани растения, пригодные для трансформации, включают ткани листа, ткани корня, меристемы и протопласты. Для трансформации растительных клеток могут использоваться бактерии рода Agrobacterium. Пригодные виды этой бактерии включают Agrobacterium tumefaciens и Agrobacterium rhizogens. Особенно целесообразно использовать вид Agrobacterium tumefaciens (например, штаммы LBA4404 или ЕНА105) вследствие его хорошо известной способности трансформировать растения. Другой подход к трансформации растительных клеток геном включает внесение инертных или биологически активных частиц в растительные ткани и клетки. Этот метод описан в US 4945050, US 5036006 и US 5100792, выданных на имя Sanford и др. В целом этот способ заключается в нанесении инертных или биологически активных частиц на клетку в условиях, обеспечивающих их прохождение через оболочку клетки и проникновение внутрь клетки. Когда используют инертные частицы, вектор может интродуцироваться в клетку путем покрытия частиц вектором, содержащим требуемый ген. В альтернативном варианте клетка-мишень может быть окружена вектором таким образом, чтобы вектор проникал в клетку вслед за частицей. Биологически активные частицы (например, высушенные дрожжевые клетки, высушенная бактерия или бактериофаг, каждый из которых содержит ДНК, которую предполагается интродуцировать) также могут быть внесены в ткань растительной клетки. Выделенный фрагмент гена по настоящему изобретению может использоваться для придания устойчивости к болезням широкому спектру растительных клеток, включая клетки голосемянных, однодольных и двудольных растений. Хотя ген может быть встроен в любую растительную клетку, входящую в эти широкие классы растений, особенно целесообразным является встраивание в клетки культурных растений, таких как рис, пшеница, ячмень, рожь, кукуруза, картофель, морковь, батат, сахарная свекла, фасоль, горох, цикорий, салат, капуста, цветная капуста, брокколи, турнепс, редис, шпинат, спаржа, лук, чеснок, баклажан, перец, сельдерей, тыква крупноплодная, тыква обыкновенная, цуккини, огурец, яблоня, груша, айва, дыня, слива, вишня, персик, нектарин, абрикос, земляника, виноград, малина, ежевика, ананас, авокадо, папайя, манго, банан, соя, табак, томат, сорго и сахарный тростник. В пригодных условиях система экспрессии по настоящему изобретению может использоваться для трансформации клетки практически любого культурного растения. Трансформированные клетки могут регенирироваться в целые растения, в результате чего ген придает устойчивость к болезням целым трансгенным растениям. Как указано выше, система экспрессии может быть модифицирована таким образом, что ген устойчивости к болезням экспрессируется постоянно или экспрессия является конститутивной. Трансформация Настоящая система может применяться в любом растении, которое может быть трансформировано и регенерировано. Такие методы трансформации и регенерации хорошо известны в данной области. Помимо указанных выше публикаций можно назвать также следующие: An G., Watson B.D. и Chiang С.С., Transformation of tobacco, tomato, potato, and Arabidopsis thaliana using a binary Ti vector system, Plant Physiol. 81: 301-305, 1986; Fry J., Barnason А. и Horsch R.B., Transformation of Brassica napus with Agrobacterium tumefaciens based vectors, PI. Cell Rep. 6: 321-325, 1987; Block M.D., Genotype independent leaf disc transformation of potato (Solanum tuberosum) using Agrobacterium tumefaciens, Theor. appl. genet. 76: 767-774, 1988; Deblock M., Brouwer D.D. и Tenning P., Transformation of Brassica napus and Brassica oleracea using Agrobacterium tumefaciens and the Expression of the bar and neo genes in the transgenic plants, Plant Physiol. 91: 694-701, 1989; Baribault T.J., Skene K.G.M., Cain P.A. и Scott N.S., Transgenic grapevines: regeneration of shoots expressing beta-glucuronidase, PI. Cell Rep. 41: 1045-1049, 1990; Hinchee M.A.W., Newell C.A., ConnorWard D.V., Armstrong T.A., Deaton W.R., Sato S.S. и Rozman R.J., Transformation and regeneration of non-solanaceous crop plants, Stadler. Genet. Symp. 203212.203-212, 1990; Barfield D.G. и Pua E.C., Gene transfer in plants of Brassica juncea using Agrobacterium tumefaciens-mediated transformation, PI. Cell Rep. 10: 308-314, 1991; Cousins Y.L., Lyon B.R. и Llewellyn D.J., Transformation of an Australian cotton cultivar: prospects for cotton improvement through genetic engineering, Aust. J. Plant Physiol. 18: 481-494, 1991; Chee P.P. и Slightom J.L., Transformation of Cucumber Tissues by Microprojectile Bombardment Identification of Plants Containing Functional and Nonfunctional Transferred Genes, GENE 118: 255-260, 1992; Christou P., Ford T.L. и Kofron M., The development of a variety-independent gene-transfer method for rice. Trends. Biotechnol. 10: 239-246, 1992; D’Halluin К., Bossut M., Bonne E., Mazur В., Leemans J. и Botterman J., Transformation of sugarbeet (Beta vulgaris L.) and evaluation of herbicide resistance in transgenic plants, Bio/Technol. 10: 309-314. 1992; Dhir S.K., Dhir S., Savka M.A., Belanger F., Kriz A.L., Farrand S.K. и Widholm J.M., Regeneration of Transgenic Soybean (Glycine Max) Plants from Electroporated Protoplasts, PLANT PHYSIOL 99: 81-88, 1992; На S.B., Wu F.S. и Thorne Т.К., Transgenic turf-type tall fescue (Festuca arundinacea Schreb.) plants regenerated from protoplasts, PI. Cell Rep. 11: 601-604, 1992; Blechi A.E., Genetic Transformation The New Tool for Wheat Improvement 78th Annual Meeting Keynote Address, CEREAL FOOD WORLD 38: 846-847, 1993; Casas A.M., Kononowicz A.K., Zehr U.B., Tomes D.T., Axtell J.D., Butler L.G., Bressan R.A. и Hasegawa P.M., Transgenic Sorghum Plants via Microprojectile Bombardment, PROC NAT ACAD SCI USA 90: 11212-11216, 1993; Christou P., Philosophy and Practice of Variety Independent Gene Transfer into Recalcitrant Crops, IN VITRO CELL DEV BIOL-PLANT 29P: 119-124, 1993; Damiani P., Nenz E., Paolocci F. и Arcioni S., Introduction of Hygromycin Resistance in Lotus spp Through Agrobacterium Rhizogenes Transformation, TRANSGENIC RES 2: 330-335, 1993; Davies D.R., Hamilton J. и Mullineaux P., Transformation of Peas, PI. Cell Rep. 12: 180-183, 1993; Dong J.Z. и Mchughen A., Transgenic Flax Plants from Agrobacterium Mediated Transformation Incidence of Chimeric Regenerants and Inheritance of Transgenic Plants, PLANT SCI 91: 139-148, 1993; Fitch M.M.M., Manshardt R.M., Gonsalves D. и Slightom J.L., Transgenic Papaya Plants from Agrobacterium Mediated Transformation of Somatic Embryos, PI. Cell Rep. 12: 245-249, 1993; Franklin C.I. и Trieu T.N., Transformation of the Forage Grass Caucasian Bluestem via Biolistic Bombardment Mediated DNA Transfer, PLANT PHYSIOL 102: 167, 1993; Golovkin M.V., Abraham M., Morocz S., Bottka S., Feher А. и Dudits D., Production of Transgenic Maize Plants by Direct DNA Uptake into Embryogenic Protoplasts, PLANT SCI 90: 41-52, 1993; Guo G.Q., Xu Z.H., Wei Z.M. и Chen H.M., Transgenic Plants Obtained from Wheat Protoplasts Transformed by Peg Mediated Direct Gene Transfer, CHIN SCI BULL 38: 2072-2078. 1993; Asano Y. и Ugaki M., Transgenic plants of Agrostis alba obtained by electroporationmediated direct gene transfer into protoplasts, PI. Cell Rep. 13, 1994; Ayres N.M. и Park W.D., Genetic Transformation of Rice, CRIT REV PLANT SCI 13: 219-239, 1994; Barcelo P., Hagel С., Becker D., Martin А. и Lorz H., Transgenic Cereal (Tritordeum) Plants Obtained at High Efficiency by Microprojectile Bombardment of Inflorescence Tissue, PLANT J 5: 583-592, 1994; Becker D., Brettschneider R. и Lorz H. Fertile Transgenic Wheat from Microprojectile Bombardment of Scutellar Tissue, PLANT J 5: 299-307, 1994; Biswas G.C.G., Iglesias V.A., Datta S.K. и Potrykus I., Transgenic Indica Rice (Oryza Sativa L) Plants Obtained by Direct Gene Transfer to Protoplasts, J BIOTECHNOL 32: 1-10, 1994; Borkowska M., Kleczkowski K., Klos В., Jakubiec J. и Wielgat В., Transformation of Diploid Potato with an Agrobacterium Tumefaciens Binary Vector System.1. Methodological Approach, ACTA PHYSIOL PLANT 16: 225-230, 1994; Brar G.S., Cohen B.A., Vick C.L. и Johnson G.W., Recovery of Transgenic Peanut (Arachis Hypogaea L) Plants from Elite Cultivars Utilizing Accell(R) Technology, PLANT J 5: 745-753, 1994; Christou P., Genetic Engineering of Crop Legumes and Cereals Current Status and Recent Advances, AGRO FOOD IND HI TECH 5: 17-27, 1994; Chupeau M.C., Pautot V. и Chupeau Y., Recovery of Transgenic Trees After Electroporation of Poplar Protoplasts, TRANSGENIC RES 3: 13-19, 1994; Eapen S. и George L., Agrobacterium Tumefaciens Mediated Gene Transfer in Peanut (Arachis Hypogaea L), PI. Cell Rep. 13: 582-586, 1994; Hartman C.L., Lee L., Day P.R. и Turner N.E., Herbicide Resistant Turfgrass (Agrostis Palustris Huds) by Biolistic Transformation, BID-TECHNOLOGY 12: 919-923, 1994; Howe G.T., Goldfarb В. и Strauss S.H., Agrobacterium Mediated Transformation of Hybrid Poplar Suspension Cultures and Regeneration of Transformed Plants, Plant Cell Tissue & Organ Culture 36: 59-71, 1994; Konwar B.K., Agrobacterium Tumefaciens Mediated Genetic Transformation of Sugar Beet (Beta Vulgaris L), J PLANTBIOCHEM BIOTECHNOL 3: 37-41, 1994; Ritala A., Aspegren К., Kurten U., Salmenkalliomarttila M., Mannonen L., Hannus R., Kauppinen V., Teeri Т.Н. и Enari T.M., Fertile Transgenic Barley by Particle Bombardment of Immature Embryos, PLANT MOL BIOL 24: 317-325, 1994; Scorza R., Cordts J.M., Ramming D.W. и Emershad R.L., Transformation of Grape (Vitis Vinifera L) Somatic Embryos and Regeneration of Transgenic Plants, J CELL BIOCHEM: 102, 1994; Shimamoto К., Gene Expression in Transgenic Monocots, CURR OPINBIOTECHNOL 5: 158-162, 1994; Spangenberg G., Wang Z.Y., Nagel J. и Potrykus I., Protoplast Culture and Generation of Transgenic Plants in Red Fescue (Festuca Rubra L), PLANT SCI 97: 83-94, 1994; Spangenberg G., Wang Z.Y., Nagel J. и Potrykus I., Gene Transfer and Regeneration of Transgenic Plants in Forage Grasses, J CELL BIOCHEM: 102, 1994; Wan Y.C. и Lemaux P.G., Generation of Large Numbers of Independently Transformed Fertile Barley Plants, PLANT PHYSIOL 104: 3748, 1994; Weeks J.T., Anderson O.D. и Blechi A.E., Stable Transformation of Wheat (Triticum Aestivum L) by Microprojectile Bombardment, J CELL BIOCHEM: 104, 1994; Ye X.J., Brown S.K., Scorza R., Cordts J. и Sanford J.C., Genetic Transformation of Peach Tissues by Particle Bombardment, JAMER SOCHORTSCI 119: 367-373, 1994; Spangenberg G., Wang Z.Y., Nagel J. и Potrykus I., PROTOPLAST CULTURE AND GENERATION OF TRANSGENIC PLANTS IN RED FESCUE (FESTUCA RUBRA L), Plant Science 1994 97: 83-94, 1995. Поскольку nim1-растения в качестве растений-хозяев могут также обладать чувствительностью к патогенам, в отличие от обычного круга хозяев, из которого они, как очевидно, выпадают, эти растения также имеют большое практическое значение для молекулярного, генетического и биологического изучения взаимосвязей растение-патоген. Кроме того, УЧБ-фенотип nim1-растений также позволяет использовать их для скрининга фунгицидов. nim1-Мутанты, отобранные для определенного хозяина, имеют большое практическое значение для скрининга фунгицидов с помощью этого хозяина и патогенов хозяина. Преимущество состоит в УЧБ-фенотипе мутанта, что позволяет решить проблемы, возникающие вследствие того, что хозяева обладают различной чувствительностью к различным патогенам и патотипам или даже устойчивостью к некоторым патогенам или патотипам. К патогенам по изобретению относятся, но не ограничены ими, вирусы и вироиды, например, вирус мозаики табака или огурца, вирус круговой пятнистости или вирус некроза, вирус курчавости листьев пеларгонии, вирус крапчатости красного клевера, вирус карликовости куста томата и подобные вирусы, грибы, например, Phythophthora parasitica и Peronospora tabacina, бактерии, например, Pseudomonas syringae и Pseudomonas tabaci, насекомые, такие, как, тли, например, Myzus persicae, и чешуекрылые, например, Heliothus spp., и нематоды, например, Meloidogyne incognita. Методы по изобретению пригодны в отношении многочисленных организмов, вызывающих болезни кукурузы, включая возбудителей ложной мучнистой росы, таких, как Scleropthora macrospora, Sclerophthora rayissiae, Sclerospora graminicola, Peronosclerospora sorghi, Peronosclerospora philippinensis, Peronosclerospora sacchari и Peronosclerospora maydis, ржавчины, таких, как Puccinia sorphi, Puccinia polysora и Physopella zeae, другие грибы, такие, как Cercospora zeae-maydis, Colletotrichum graminicola, Fusarium monoliforme, Gibberella zeae, Exserohilum twcicum, Kabatiellu zeae и Bipolaris maydis, и бактерии, такие, как Erwinia stewartii, но не ограничены ими. Описание перечисленных последовательностей SEQ ID NO: 1: геномная последовательность длиной 9919 пар оснований, приведенная на фиг.14. SEQ ID NO: 2: геномная последовательность длиной 5655 пар оснований, приведенная на фиг.15. SEQ ID NO: 3: аминокислотная (АК) последовательность NIM-протеина дикого типа, кодируемая/кодирующей последовательностью (cds) SEQ ID NO: 2. SEQ ID NO: 4: АК последовательность (33-155) риса Rice-1, приведенная на фиг.19. SEQ ID NO: 5: АК последовательность (215-328) риса Rice-1, приведенная на фиг.19. SEQ ID NO: 6: АК последовательность (33-155) риса Rice-2, приведенная на фиг.19. SEQ ID NO: 7: АК последовательность (208-288) риса Rice-2, приведенная на фиг.19. SEQ ID NO: 8: АК последовательность (33-155) риса Rice-3, приведенная на фиг.19. SEQ ID NO: 9: АК последовательность (208-288) риса Rice-3, приведенная на фиг.19. SEQ ID NO: 10: АК последовательность (33-155) риса Rice-4, приведенная на фиг.19. SEQ ID NO: 11: АК последовательность (215-271) риса Rice-4, приведенная на фиг.19. Депонирование Следующие векторные молекулы были депонированы в Американской коллекции типовых культур (American Type Culture Collection, 12301 Parklawn Drive Rockville, MD 20852, U.S.A.) в указанные ниже сроки: плазмида ВАС-04 была депонирована в АТСС 8 мая 1996 г. под номером АТСС 97543; плазмида Р1-18 была депонирована в АТСС 13 июня 1996 г. под номером АТСС 97606; космида D7 была депонирована в АТСС 25 сентября 1996 г. под номером АТС 97736. Примеры Пример 1: Выявление NIM1-клонов с помощью клонирования, основанного на картировании: генетическое картирование высокого разрешения и физическое картирование NIM1 в Arabidopsis 1. Растительный материал и выделение nim1-мутантов Мутанты nim1 выделяли из двух популяций растений Arabidopsis экотипа Ws-О согласно методу, описанному у Delaney и др. (1995), PNAS 92, 6602-6606. Одна мутантная популяция была в форме М 2-библиотеки, происходящей из семян, которые подвергали индуцированному мутатагенезу с помощью этилметансульфоната (ЭМС) (получены от фирмы Lehle, Round Rock, ТХ), а вторая была в форме популяции Т-ДНК, происходящей из семян, полученных из Ohio State University Arabidopsis Biological Resource Center (Columbus, OH). Основой для скрининга мутантов с неиндуцируемым иммунитетом (nim1) был поиск с использованием подвергнутых индуцированному мутагенезу популяциий растений, у которых устойчивость к вирулентному патогену не могла индуцироваться ИНК (2,6-дихлоризоникотиновая кислота; Metraux и др., 1991, в Advances in Molecular Genetics of Plant-Microbe Interactions, том 1, 432-439, под ред. Hennecke и Verma; Kessmann и др., 1993, в Mode of action of agrochemicals, под ред. Y Honma; Vernooij и др., 1995, Molec. PI. Microbe Interaction 8, 228-234). Растения из мутантных популяций выращивали с высокой плотностью в больших поддонах в поступающей в продажу смеси для выращивания. Когда растения достигали 2-недельного возраста, поддоны опрыскивали 0,25 мг/мл ИНК. Через 4 дня растения опрыскивали суспензией спор Peronospora parasitica, изолят EmWa (EmWa) с концентрацией от 5×104 до 1×105 спор/мл. Обычно этот гриб вирулентен в отношении экотипа Arabidopsis Ws-O, если устойчивость первоначально не индуцировалась у этих растений с помощью ИНК или аналогичного соединения. После инкубации в условиях с высокой влажностью выявляли растения с видимыми симптомами болезни, как правило, на 7-й день после заражения. Эти растения не проявляли устойчивости к грибам, несмотря на обработку индуцирующим устойчивость химическим соединением, и поэтому их рассматривали как потенциальные мутантные растения, не имеющие иммунитета, nim-мутанты (non-immunity). Из 360000 растений выявлено 75 потенциальных nim-мутантов. Эти потенциальные мутантные растения извлекали из ящика для рассады, помещали в условия с низкой влажностью и позволяли дать семена. Растения, полученные из этого семенного материала, подвергали скринингу аналогично тому, как проводили скрининг в отношении чувствительности к изоляту грибов EmWa, и в этом случае также после предварительной обработки ИНК. Потомство растений, которое проявляло симптомы заражения, обозначали как nim-мутанты. Таким образом было выявлено шесть nim-мутантов. Одну линию (nim1) выделяли из популяции Т-ДНК и пять из популяции, обработанной ЭМС. 2. Оценка реакций растений на ИНК и другие химические индукторы устойчивости к болезням I. Фенотипический анализ nim1 Салициловая кислота (СК) и S-метиловый эфир бензо(1,2,3)тиадиазол-7-карботионовой кислоты (БТК) представляют собой два химических соединения, которые аналогично ИНК индуцируют у растений дикого типа устойчивость к широкому спектру болезней, которую называют системной приобретенной устойчивостью (SAR). Поскольку ИНК не индуцирует устойчивость у nim1- растений, у этих растений также оценивали возникновение устойчивости к болезням в ответ на предварительную обработку СК и БТК как это, в частности, описано у Delaney и др., 1995, PNAS 92, 6602-6606). Растения опрыскивали 1, 5 или 15 мМ СК или 0,25 мг/мл БТК и через 5 дней заражали инокулятом EmWa (как описано выше в примере 1). Как СК, так и БТК не могли защитить nim1-растения от поражения грибом, что видно по наличию симптомов болезни и росту гриба на этих растениях. Таким образом, nim1-растения нечувствительны к любому из индуцирующих SAR химических соединений, позволяя предположить, что мутация расположена ниже по ходу транскрипции от точки (ек) вступления этих химических соединений в процесс индукции устойчивости. nim1-Растения также оценивали в отношении их чувствительности к болезням при заражении двумя несовместимыми инокулятами Р. parasitica Wela и Noco (т.е. эти штаммы гриба не вызывали заболевания у растений Ws-O дикого типа), nim1-Растения опрыскивали суспензией конидий, содержащей 5-10×104 спор/мл штамма Wela или Noco, и инкубировали при высокой влажности в течение 7 дней. В отличие от растений дикого типа у nim1-растений развились симптомы болезни в ответ на заражение и Wela и Noco. Симптомы состояли в некротической пятнистости и в пригибании к земле, при наличии определенной споруляции. После окрашивания лактофенолом голубым гифы гриба становились хорошо заметными на листьях nim1-растений. Таким образом, nim1-растения являются чувствительными к несовместимым в норме изолятам Р.parasitica. Эти результаты свидетельствуют о том, что nim1-растения не только дефектны в отношении индуцируемой устойчивости к болезням, но они также дефектны в отношении естественной устойчивости к микроорганизмам, которые в норме не являются для них патогенными. II. Биохимический анализ nim1 СК, ИНК и БТК индуцируют у Arabidopsis SAR и экспрессию генов SAR, которые включают связанные с патогенезом гены (Pathogenesis Related genes) PR-1, PR-2 и PR-5. Поскольку эти соединения не индуцируют устойчивость к болезням у nim1, с которыми в норме они несовместимы (как описано в примере 1.2, см. выше), эту мутантную линию анализировали в отношении экспрессии гена SAR после обработки СК, ИНК или БТК. После обработки nim1-растений СК, ИНК или БТК ткани растений собирали и анализировали на накопление РНК генов PR-1, PR-2 и PR-5. После этого общую РНК выделяли из обработанных тканей и подвергали электрофорезу на агарозном геле. Получали трехкратно повторенные блоты геля и каждый из них гибридизовали с зондом для одного из этих трех генов SAR согласно методу, описанному у Delaney и др., 1995, PNAS 92, 6602-6606. В отличие от того, что происходит в случае растений дикого типа, химические соединения не ндуцировали накопление РНК ни в одном из этих трех генов SAR в nim1-растениях, как показано на фиг.1. Совокупность полученных результатов свидетельствует о том, что химические соединения не индуцируют ни SAR, ни экспрессию генов SAR в nim1-растениях. Поскольку химические соединения не индуцировали ни SAR, ни экспрессию генов SAR в nim1-растениях, представляло интерес исследовать, может ли заражение патогеном индуцировать экспрессию генов SAR в этих растениях, как это происходит в растениях дикого типа. Ws-O- и nim1-растения опрыскивали спорами EmWa, как описано ранее, и через определенные промежутки времени брали ткани на анализ РНК. Заражение патогеном (EmWa) Ws-O-растений дикого типа индуцировало экспрессию гена PR-1 через 4 дня после заражения, как видно на фиг.2. Однако у nim1-растений экспрессия гена PR-1 не индуцировалась в течение 6 дней после заражения, а его уровень был ниже по сравнению с уровнем в растениях дикого типа в этот момент времени. Таким образом, после заражения патогеном экспрессия гена PR-1 в nim1-растениях замедляется и понижается по сравнению с растениями дикого типа. Заражение растений дикого типа патогенами, которые вызывают некротическую реакцию, приводит к накоплению СК в зараженных тканях. Было показано, что эта эндогенная СК необходима для сигнальной трансдукции в SAR-пути, т.е. разложение эндогенной СК приводит к снижению устойчивости к болезням. Это свидетельствует о том, что накопление СК является маркером для SAR-пути (Gaffney и др., 1993, Science 261, 754-756). Оценивали способность nim1-растений накапливать СК после заражения патогеном. Вирулентным для томатов штаммом Pseudomonas syringae DC 3000, несущим ген avrRpt2, заражали листья 4-недельных nim1-растений. Через 2 дня листья собирали для анализа СК согласно методу, описанному Delaney и др., 1995, PNAS 92, 6602-6606. Этот анализ показал, что nim1-растения накапливали высокие уровни СК в зараженных листьях, как видно на фиг.3. В незараженных листьях также происходило накопление СК, но не до таких уровней, как в зараженных листьях, а подобно тому, как это происходило в растениях Arabidopsis дикого типа. Это свидетельствует о том, что nim-мутация на генетической карте расположена ниже по ходу транскрипции относительно маркера СК в пути трансдукции сигнала. Это представлялось вероятным, поскольку для ИНК и БТК (неактивных в nim1-растениях) известна способность стимулировать компонент в SAR-пути, расположенный ниже по ходу транскрипции от СК (Vernooij и др., 1995, Molec. PI. Microbe Interaction 8, 228-234; Friedrich и др., 1996, The Plant Journal 9, в печати; Lawton и др., 1996, The Plant Journal 9, в печати). Кроме того, как описано в примере 1.2, экзогенно внесенная СК не защищает nim1-растения от заражения EmWa. 3. Генетический анализ nim1 Осуществляли обратное скрещивание nim1-растения с растениями дикого типа Ws-O и их потомство F1 после предварительной обработки ИНК тестировали в отношении устойчивости к EmWa согласно методу, описанному выше в примере 1.1. Ни у одного из F1-растений, предварительно обработанных ИНК, не обнаружено симптомов заражения, в то время как в контрольных nim1-растениях заражение обнаружено. Отсюда следует, что nim1-мутация является рецессивной. Популяцию второго поколения F2, полученного при скрещивании Ws-O × nim1, также после предварительной обработки ИНК анализировали в отношении устойчивости к болезням. В этой популяции приблизительно у 1/4 (32 из 130 растений) после заражения EmWa обнаружены симптомы болезни у растений, предварительно опрыснутых ИНК, и у 3/4 растений не выявлено заболевания. Эти результаты свидетельствуют о том, что nim-мутация расположена в отдельном генетическом локусе, и подтверждаются полученными с использованием F1 данными, свидетельствующими о рецессивной природе мутации. 4. Выявление маркеров и генетическое картирование локуса NIM Для общепринятого клонирования гена NIM на основе картирования должны быть выявлены маркеры, которые генетически тесно связаны с мутацией. Это исследование проводили в 2 стадии. Сначала nim1-растения скрещивали с другим генотипом Arabidopsis – Landsberg erecta (Ler) – и выявляли растения F2, полученные в результате этого скрещивания, которые имели фенотип nim1 (т.е. растения, которые являются гомозиготными nim/nim в локусе NIM). Далее с помощью молекулярного анализа среди этих растений выявляли растения, которые имели генотип Ler, расположенный непосредственно рядом с ДНК-маркером. В соответствии с критерием идентификации эти растения являются результатом рекомбинации между маркером и локусом NIM. Частота рекомбинантов определяет расстояние на генетической карте между маркером и локусом NIM. Вторым предварительным условием для клонирования на основе картирования является выявление маркеров, которые генетически очень близки к локусу NIM, т.е. маркеров, для которых обнаружено очень мало рекомбинантов. Если выявлены генетические маркеры, которые являются очень близкими, то они могут использоваться для выделения клонов геномной ДНК, которые близки к локусу NIM. Локус NIM затем может быть клонирован методом “прогулки вдоль хромосомы”, если он уже не присутствует на клонированной ДНК. “Прогулка вдоль хромосомы” может быть начата с обеих сторон гена. Этот метод основан на получении перекрывающихся клонов, которые последовательно расположены все ближе и ближе к представляющему интерес гену. Когда отдельный ДНК-маркер получают в результате “прогулки”, начатой, например, с Нозерн-конца, и выявляют отсутствие рекомбинантов между этим маркером и представляющим интерес геном, то этот маркер расположен очень близко к гену. Однако, если для маркера выявлен (ы) рекомбинант(ы) с Саузерн-концом, клон, из которого этот маркер был получен, должен образовывать перекресты с геном. Путем такого определения клонируют представляющий интерес ген. Он должен быть локализован между маркером и последним маркером, расположенным на Нозерн-конце, что определяют по наименьшему количеству рекомбинантов с Нозерн-концом. На первой стадии большое количество рекомбинантов получают при генетическом скрещивании. На второй стадии, рекомбинанты, которые близки к гену NIM, выделяют с помощью молекулярных маркеров. Многие маркеры описаны в литературе и существует несколько методов получения дополнительных маркеров. Примененный в данном изобретении подход основан на ряде маркерных систем, включающих ПДПП и ПДАФ (см. ниже). I. Генетические скрещивания С целью картирования хромосомного положения гена NIM относительно ПДПП- и ПДАФ-маркеров nim1 скрещивали с Ler, создавая картирующую популяцию. Выращивали растения F2, полученные в результате этого скрещивания, и листья собирали для дальнейших экстракций ДНК. Затем растения F2 оценивали в отношении nim1-фенотипа согласно методу, описанному выше в примере 1.1. Также выращивали популяции F3, полученные от отдельных растений F2, и оценивали в отношении nim1-фенотипа. ДНК экстрагировали из хранившейся ткани растений F2 и F3, имеющих фенотип nim1, методом СТаb, как описано у Rogers и Bendich, 1988, Plant Molecular Biology Manual, A6, 1-10. Эту ДНК использовали для картирования гена NIM, как описано ниже. II. Маркеры полиморфизма длин простых последовательностей Маркеры полиморфизма длин простых последовательностей (ПДПП) ATHGENEA и nga111 были описаны ранее (Bell и Ecker, 1994, Genomics 19, 137-144). Праймеры, примененные для выявления этих ПДПП-маркеров, приведены в таблице 1.
Генетическое картирование гена NIM относительно маркера ATHGENEA: При использовании праймеров ATHGENEA (1) для ПЦР-амплификации геномной ДНК Ler, ожидали получить полосу, соответствующую 205 парам оснований (п.о.), а при использовании геномной ДНК Ws-O ожидали получить полосу 211 п.о. (Bell и Ecker, 1994, Genomics 19, 137-144). Полученные продукты амплификации трудно разделить на обычных агарозных гелях. Поэтому были разработаны два различных метода разделения и обнаружения этих ПЦР-фрагментов. При использорвании первого метода набор праймеров ATHGENEA (1) (таблица 1) применяли для амплификации геномной ДНК в присутствии меченного с помощью 6-карбоксиродамина УТФ (дУТФ-R110, получен от фирмы ABI), получая меченные с помощью родамина ПЦР-фрагменты. ПЦР-реакции анализировали на секвенаторе ДНК, разрешающая способность которого в отношении фрагментов ДНК составляет один нуклеотид. Использовали следующие специфические реагенты: однократный ПЦР-буфер (1×ПЦР-буфер), 2 мМ MgCl2, по 200 мМ каждого дНТФ, 2 мМ дУТФ-R110, праймеры ATHGENEA (1) в концентрации 0,75 мМ, 10 нг ДНК и 0,75 ед. полимеразы Taq на 20 мл объема реакционной смеси. Условия амплификации были следующими: 3 мин при 94°С, затем 35 циклов по 15 с при 94°С, 15 с при 55°С и 30 с при 72°С. Эти образцы анализировали на секвенаторе ДНК типа ABI 377 DNA Sequencer, который способен обнаруживать флуоресцентномеченные фрагменты ДНК с разрешающей способностью один нуклеотид (нт.). С помощью этого метода получены следующие данные о генотипе растительных образцов: фрагмент ДНК длиной 205 нуклеотидов получали из ДНК Ler, а полосу, соответствующую 211 нуклеотидам, получали из ДНК Ws-O. Таким образом, могут быть легко разделены фрагменты ДНК, имеющие длину, отличающуюся на 6 нуклеотидов, что позволяет легко определить следующий генотип образцов в локусе ATHGENEA: гомозиготный Ws-O, гомозиготный Ler и гетерозиготный Ws-O/Ler. С целью увеличения производительности этой системы использовали мультиплексную схему. Некоторые образцы ДНК подвергали ПЦР-амплификации, как описано выше, с набором праймеров ATHGENEA (1), a другие образцы анализировали с помощью набора праймеров ATHGENEA (2) (указаны в таблице 2), в каждом случае в присутствии меченного с помощью 6-карбоксиродамина дУТФ. Набор праймеров ATHGENEA (2) создавали на основе опубликованной последовательности ATHGENEA (Simoens и др., 1988, Gene 67, 1-11). Этот набор праймеров обеспечивал амплификацию фрагмента ДНК длиной 139 пар оснований из ДНК Ler и полосу, соответствующую 145 парам оснований, из ДНК Ws-O. Условия реакции амплификации для набора праймеров ATHGENEA (2) были идентичны таковым, описанным выше для набора праймеров ATHGENEA (1). Отдельные реакционные смеси, полученные с помощью набора праймеров ATHGENEA (1), и отдельные реакционные смеси, полученные с помощью набора праймеров ATHGENEA (2), смешивали вместе перед электрофорезом в секвенаторе ДНК типа ABI 377 DNA Sequencer. Этот мультиплексный подход позволил проводить в секвенаторе определение генотипа 2-х образцов на одной дорожке секвенатора, одного в положениях 145/139 нт., а другого в положениях 211/205 нт. В другом методе ПЦР-фрагменты метили праймером, меченным с помощью флуоресцентного красителя РАМ-6 (6-карбоксифлуоресцеин) (фирмы Integrated DNA Technologies, Inc.). ATHGENEA-праймеры, обеспечивающие синтез по ходу цепи, в наборах праймеров ATHGENEA (1) и (2) идентичны по своей последовательности (таблица 1). Этот праймер метили с помощью FAM-6 и применяли для ПЦР-амплификации с использованием следующих реагентов (фирма Perkin Elmer): lxXL-буфер, 1 мМ MgCl2, по 200 мМ каждого дНТФ, по 0,50 мМ каждого праймера (праймер, обеспечивающий синтез по ходу цепи, меченный с помощью FAM-6), 10 нг геномной ДНК и 0,5 ед. полимеразы XL на 20 мл объема реакционной смеси. Условия циклов были следующие: 3 мин при 94°С, затем 35 циклов по 15 с при 94°С, 15 с при 59°С и 30 с при 72°С. И в этом случае отдельные реакционные смеси, полученные с использованием набора праймеров ATHGENEA (1), и отдельные реакционные смеси, полученные с использованием набора праймеров ATHGENEA (2), смешивали вместе перед электрофорезом в секвенаторе ДНК типа ABI 377 DNA Sequencer. Этот мультиплексный подход позволил проводить в секвенаторе определение генотипа 2-х образцов на одной дорожке секвенатора, одного в положениях 145/139 нт., а другого в положениях 211/205 нт. Все образцы из поколений F2 и F3 растений с фенотипом nim1 оценивали в отношении их генотипа в локусе ATHGENEA согласно описанному выше методу. Все образцы, которые были гетерозиготными по этому локусу, соответствовали растениям, имеющим рекомбинации между локусом NIM1 и локусом ATHGENEA. В популяции, состоящей из 1144 растений F2 с фенотипом nim1, и в популяциях F3 с nim1-фенотипом, которые оценивали таким образом, 98 оказались гетерозиготными по локусу ATHGENEA, позволяя установить, что генетическое расстояние между этим ПДПП-локусом и локусом NIM1 составляет 4,3 сМ (сантиморганид). Отсюда следует, что локус NIM1 находится на хромосоме 1 около маркера ATHGENEA. Генетическое картирование гена NIM1 относительно маркера nga111: Два набора праймеров для ПДПП-маркера nga111 (которые описаны у Bell и Ecker, 1994, Genomics 19, 137-144) применяли для амплификации геномной ДНК растений поколений F2 и F3 nim1-фенотипа и контрольных растений эпитопов Ws-O и Ler. Набор праймеров nga111 (1) (описан у Bell и Ecker, 1994, Genomics 19, 137-144 и представлен в таблице 1) применяли в следующих условиях: 1×ПЦР-буфер, 2 мМ MgCl2, по 200 мМ каждого дНТФ, по 0,75 мМ каждого праймера, 10 нг геномной ДНК и 0,75 ед. полимеразы Taq на 20 мл объема реакционной смеси. Набор праймеров nga111 (2) (представленный в таблице 1 и являющийся производным от набора праймеров ngalll (D) применяли в других условиях: 1×ПЦР-буфер, 1,5 мМ MgCl2, по 200 мМ каждого дНТФ, по 1 мМ каждого праймера, 10 нг ДНК и 1 ед. полимеразы Taq на 20 мл объема реакционной смеси. Обе реакционные смеси амплифицировали путем инкубации при 94°С в течение 1 мин, затем 40 циклов по 15 с при 94°С, 15 с при 55°С и 30 с при 72°С. Образцы анализировали на 3-5%-ных агарозных гелях. Полоса, полученная после амплификации ДНК Ws-O с любым набором праймеров, соответствовала 146 п.о., а амплификация ДНК Ler давала полосу 162 п.о. Образцы растений, гетерозиготных по локусу nga111, соответствовали растениям, которые имели рекомбинации между этим ПДПП-маркером и локусами NIM. В популяции, состоящей из 1144 растений F2 с фенотипом nim1, и в популяциях F3 с nim1-фенотипом, 239 растений оказались гетерозиготными по локусу nga111-маркера, позволяя установить, что генетическое расстояние между этим ПДПП-маркером и локусом NIM1 составляет 10,4 сМ. Отсюда следует, что локус NIM1 находится на хромосоме 1. Поскольку существует несколько растений с nim1-фенотипом, которые гетерозиготны и по ATHGENEA, и по nga111, было установлено, что локус NIM1 расположен между этими двумя маркерами, причем ATHGENEA локализован ближе к Нозерн-концу от гена NIM1, a nga111 локализован ближе к Саузерн-концу от гена NIM1. Это позволяет расположить ген NIM1 приблизительно на расстоянии 10 сМ ближе к Нозерн-концу от nga111, около положения 85 на хормосоме 1 (Lister и Dean, 1993, Plant J. 4, 745-750; Bell и Ecker, 1994, Genomics 19, 137-144). III. Маркеры полиморфизма длин амплифицированных фрагментов Для основанного на картировании клонирования гена NIM1 необходимо выявить молекулярные маркеры, которые расположены максимально близко к этому гену. Для этой цели получали маркеры полиморфизма длин амплифицированных фрагментов с помощью метода избирательной амплификации рестрикционных фрагментов методом, описанным у Zabeau и Vos (1993, ЕР 0534858) и Vos и др. (1995, Nucleic Acid Research 23, 4407-4414). Описание ПДАФ-технологии Применение ПДАФ-технологии для картирования основано на избирательной амплификации серии полос ДНК в двух генетически различных образцах. Обнаружение того факта, что любые полученные полосы различны у двух генотипов, позволяет выявить эти полосы в качестве маркеров для данного генотипа. Если для маркеров характерна высокая частота косегрегации с геном вставки (мутация), то этот маркер является близким к генетическому локусу. Селективную амплификацию небольшого набора фрагментов ДНК в комплексе с образцом ДНК получают с помощью двухстадийного процесса. Сначала получают фрагменты ДНК путем расщепления ДНК ферментами рестриктазами с последующим лигированием адаптеров с концами. На второй стадии используют праймеры, состоящие из последовательности, комплементарной адаптерам, плюс удлинение на 3’-конце (обычно на 0-3 нуклеотида), для амплификации только тех фрагментов ДНК, концы которых комплементарны этим праймерам. Если применяют удлинение в один нуклеотид, то теоретически каждый праймер будет “соответствовать” приблизительно 1/4 всех фрагментов, причем 1/16 всех фрагментов будет иметь праймеры, “соответствующие” обеим концам. Таким образом, с этими праймерами проводят амплификацию ограниченного набора фрагментов ДНК. Путем последующего мечения с помощью радиоактивного изотопа одного праймера могут быть получены еще более мелкие субнаборы видимых полос. ПДАФ-анализ Для ПДАФ-анализа образцов ДНК 50 нг ДНК расщепляли с помощью соответствующих ферментов (обычно EcoRI и Msel; см. ниже) и адаптеры (перечисленные в таблице 2, ниже) лигировали с фрагментами, полученными в результате обработки рестриктазами (обычно EcoRI и Msel). Последовательности праймеров и клоны YAC, P1 и ВАС подробно описаны ниже. Для реакций амплификации применяли матрицы (приблизительно по 0,5 нг ДНК на каждую реакцию) с использованием праймеров, комплементарных адаптерам, с короткими 3’-удлинениями (2 или 3 нуклеотида; последовательности праймеров приведены ниже). Поскольку один из праймеров является радиоактивномеченным (обычно EcoRI-праймер), для разделения полос используют только субнабор амплифицированных фрагментов, видимый при авторадиографическом анализе геля. Условия амплификации для клонированных ДНК (YAC, P1, космида) были следующие: 36 циклов по 30 с при 94°С (денатурация), отжиг 30 с и удлинение 1 мин при 72°С. Температура отжига в первом цикле составляла 65°С, и ее понижали на 0,7°С в каждом цикле в течение следующих 12 циклов и затем поддерживали на уровне 56°С. Для геномной ДНК растений Arabidopsis амплификацию проводили в 2 стадии: на первой стадии (предварительная амплификация) ДНК амплифицировали с праймерами, которые имеют удлинение на 1 нуклеотид (ни один из праймеров не метили). Условия реакции для этой реакции амплификации были следующими: 20 циклов по 30 с денатурация (94°С), отжиг 1 мин (56°С) и удлинение 1 мин (72°С). На второй стадии реакционную смесь для амплификации сначала разбавляли в 10 раз и далее проводили повторную амплификацию в течение 36 циклов с праймерами, содержащими полноразмерные удлинения (с использованием одного меченого праймера) в следующих условиях: 30 с при 94°С (денатурация), отжиг 30 с и удлинение 1 мин при 72°С. Температура отжига в первом цикле составляла 65°С, и ее понижали на 0,7°С в каждом цикле в течение следующих 12 циклов и затем поддерживали на уровне 56°С. Конечные продукты реакции разделяли на полиакриламидном геле и гель экспонировали на пленку, что позволяло визуализировать радиоактивномеченные ПЦР-полосы. Когда этот анализ применяли одновременно для ДНК двух генотипов, выявляли ПДАФ-полосы, которые были характерны в качестве диагностического признака для одного или для другого генотипа. Такие информативные полосы называют ПДАФ-полосами или ПДАФ-маркерами. В таблице 2 приведены адаптеры, которые использовали для ПДАФ-анализа.
Получение ПДАФ-маркеров и точное картирование локуса NIM1 Популяцию рекомбинантных инбредных линий, полученных при скрещивании таких экопитов Arabidopsis, как Landsberg erecta (Ler) и Columbia (Col) (Lister и Dean, 1993, Plant J. 4, 745-750), использовали для скрининга ПДАФ-маркеров. Для ПДАФ-скрининга использовали следующие праймеры: EcoRI-праймеры: 5’-GACTGCGTACCAATTCWN-3’ MseI-праймеры: 5’-GATGAGTCCTGAGTAAXWN-3’ Символ “N” в праймерах обозначает, что этот участок является вариабельным (А, С, G или Т), “W” обозначает А или Т и “X” обозначает С. Применяли все 8 возможных праймеров как для EcoRI-, так и для Msel-праймера. Это дает всего 64 (8×8) комбинации праймеров (КП), которые использовали для амплификации ДНК рекомбинантной инбредной линии и родительских генотипов Ler и Col, как описано выше. Реакции амплификации проводили в денатурирующем полиакриаламидном геле с целью разделения ПДАФ-фрагментов по размеру и гель экспонировали на пленку. Пленку анализировали с точки зрения всех полос, которые присутствуют только в одном генотипе, т.е. анализировали в отношении ПДАФ-маркеров. ПДАФ-маркеры, т.е. фрагменты ДНК, полиморфные для обоих родителей рекомбинантных инбредных линий, применяли для создания генетической карты популяции рекомбинантной инбредной линии. Ниже в примере 1.51 описано картирование гена NIM1 на хромосоме 1 Arabidopsis приблизительно в 85 положении. Те ПДАФ-маркеры, которые были картированы (с использованием рекомбинантной инбредной линии) между положением 81 и 88 хромосомы 1 Arabidopsis, были выбраны для анализа рекомбинантных растений на присутствие указанных ПДАФ-маркеров и, таким образом, для более точного картирования гена NIM1. Было установлено, что семь ПДАФ-маркеров из этой области являются информативными; они были полиморфными для обоих родителей при скрещивании nim1 x Ler. Шесть ПДАФ-маркеров оказались Ler-специфичными, т.е. эти ПДАФ-маркеры отсутствовали в Ws (а также в Col). Один ПДАФ-маркер был Ws-специфичным, т.е. Col-специфичным ПДАФ-маркером (отсутствует в Ler), который также присутствовал в Ws. Эти ПДАФ-маркеры представляют собой L81.1, L81.2, W83.1, L84, L85, L87 и L88 (L-маркер является специфичным для экотипа Ler, a W-маркер является специфичным для обоих экотипов Col и Ws; номера обозначают положение на карте). Эти ПДАФ-маркеры использовали для анализа рекомбинантных растений и скрещивания nim1 × Ler (см. ниже). Кроме того, ПДАФ-маркер С86 (специфичный для Col маркер, происходящий из рекомбинантной инбредной линии) применяли для выделения клонов ДНК (см. ниже). В таблице 3 приведены последовательности праймеров, которые использовали для получения этих ПДАФ-маркеров. В таблице 3 приведены комбинации праймеров ПДАФ-маркеров, полученные из популяции рекомбинантной инбредной линии. “EcoRI-” обозначает последовательность 5’-GACTGCGTACCAATTC-3’ и “MseI-” обозначает последовательность 5’-GATGAGTCCTGAGTAA-3’.
Подробную генетическую карту области создавали, используя описанные выше ПДАФ-маркеры, путем типирования рекомбинантов. Всего 337 рекомбинантных растений было получено из 1144 растений второго поколения F2 с фенотипом nim1. Эти рекомбинантные растения сначала подвергали скринингу с помощью фланкирующих Нозерн-конец ПДАФ-маркеров L81.2 и ATHGENEA и фланкирующих Саузерн-конец маркеров L88 и nga111. Сорок восемь растений оказались гомозиготными по nim1/nim1 и гетерозиготными по ATHGENEA и L81.2, а 21 растение оказалось гомозиготным по nim1/nim1 и гетерозиготным по nga111 и L88. Эти рекомбинантные растения затем анализировали с использованием девяти ПДАФ-маркеров в NIM-области, включая четыре ПДАФ-маркера, которые получены при картировании популяции рекомбинантной инбредной линии (W83.1, L84, L85 и L87), и пять ПДАФ-маркеров, полученных при анализе клонов YAC (W83.3/W84.1, W84.2, W85.1, W86.1 и L86, см. ниже). Основанная на использовании этого анализа генетическая карта NIM1 приведена на фиг.4. Видно, что 27 рекомбинантов были обнаружены между маркером W84.2 и NIM1, а 14 рекомбинантов обнаружено между W85.1 и NIM1. Маркер L85 расположен близко к NIM1, но этот маркер не мог быть картирован в клонах YAC, ВАС или Р1 (см. ниже) и, следовательно, не мог применяться для идентификации гена NIM1. 5. Физическое картирование NIM1-области I. Выделение клонов YAC с помощью ПДАФ-маркеров, расположенных близко к NIM1 Библиотеку CIC, библиотеку YAC экотипа Arabidopsis Columbia (Bouchez и др. 1995, 6th Int. Conf. on Arabidopsis Research (материалы 6-й Международной конференции по исследованию Arabidopsis), Мэдисон, шт. Висконсин) подвергали скринингу в отношении клонов YAC в NIM-области. Эта библиотека включает приблизительно 2,5 эквивалента ядерного генома и имеет средний размер вставки 450 т.п.н. Библиотеку YAC подвергали скринингу с помощью двух ПДАФ-маркеров: W83.1 и С86. W83.1 представляет собой наиболее близко расположенный к Нозерн-концу NIM1 ПДАФ-маркер, полученный из рекомбинантной инбредной линии, а С86 представляет собой ПДАФ-маркер, полученный из рекомбинантной инбредной линии, специфичный для Col (отсутствует в Ler и Ws). C86 картирован на Саузерн-конце гена NIM1 на карте популяции рекомбинантной инбредной линии. Этот специфичный для Col ПДАФ-маркер использовали вместо близко расположенных специфичных для Ler ПДАФ-маркеров (фиг.4), поскольку последние ПДАФ-маркеры выявляли только экотип Landsberg erecta и, следовательно, не могли быть использованы для скрининга библиотеки YAC экотипа Columbia. Библиотеку YAC подвергали скринингу в два этапа. Сначала клетки клонов YAC каждой пластины двенадцати 96-луночных планшетов для микротитрования объединяли в группу (пул пластин) и применяли для выделения ДНК согласно методу, описанному Ross и др. (1991, Nucleic Acids Res. 19, 6053). Пулы подвергали скринингу с обоими ПДАФ-маркерами. Затем из каждого позитивного пула пластин образцы ДНК из каждого ряда (пул из 8 клонов) и каждой колонки (пул из 12 клонов) подвергали скринингу с ПДАФ-маркером, для которого пул пластин был позитивным. С использованием этого метода могут быть выявлены отдельные позитивные клоны YAC. Скрининг привел к выделению всего четырех клонов YAC: YAC 12F04 и YAC 12H07 были выделены с использованием Нозерн-ПДАФ-маркера W83.1, a YAC 10G07 и YAC 7E03 – с использованием Саузерн-AFLP-маркера C86 (для номенклатуры клонов YAC используют принятую для CIC нумерацию). Были получены “фингерпринты” YAC с помощью ПДАФ, которые давали YAC-специфические ПДАФ-фрагменты. Фингерпринты YAC сравнивали и применяли для оценки перекрытий между YAC (см. также таблицы 5 и 6). На основе полученных с помощью ПДАФ фингерпринтов установлено, что клон 7Е03 в значительной степени перекрывается клоном 10G07 (см. также таблицу 5), а клон 12Н07 аналогичным образом в значительной степени перекрывается клоном 12F04 (см. также таблицу 6). III. Получение ПДАФ-маркеров из клонов YАС Поскольку описанные выше ПДАФ-маркеры были генетически относительно далеко расположены от гена NIM1 (см. фиг.3), дополнительные ПДАФ-маркеры были созданы с целью поиска маркеров, которые находились бы ближе к гену NIM. Скрининг дополнительных происходящих из YAC ПДАФ-маркеров проводили с использованием следующих образцов ДНК: ДНК из выделенных клонов YAC (как описано ранее, было выявлено 4 клона YAC), клона дрожжей без YAC и трех таких экотипов Arabidopsis, как Col, Ler и Ws. С помощью этого скрининга фрагменты, специфичные для клонов YAC (отсутствующие у штамма дрожжей и присутствующие в экотипе Col), могли быть оценены в отношении полиморфизма в растениях Ler и Ws (родителях рекомбинантных растений, выявленных в примере 1.5, см. ниже). Таким образом, все выявленные полиморфные фрагменты могут служить дополнительными ПДАФ-маркерами. При первом скрининге ПДАФ использовали комбинацию ферментов (КФ) EcoRI/MseI. При этом скрининге исследовали два клона YAC 10G07 и 7Е0З (обнаруженные с помощью ПДАФ-маркера С86, см. ниже), штамм дрожжей без YAC и три экотипа Arabidopsis Col, Ler и Ws. Примененные комбинации праймеров с селективными удлинениями могут быть подразделены на три группы, и они представлены в таблице 4. Всего подвергали скринингу 256 (64+96+96) комбинаций праймеров. Ниже в таблице 4 приведены последовательности праймеров, которые использовали при ПДАФ-скрининге двух клонов YAC, 10G07 и 7Е03, штамма дрожжей, несодержащего YAC, и трех экотипов Arabidopsis Col, Ler и Ws. Использовали три группы комбинаций праймеров. Символ “N” в праймерах свидетельствует о том, что эта часть была вариабельной (А, С, G или Т), “S” обозначает С или G, “W” обозначает А или Т и “Y” обозначает С или Т. Таблица 4 EcoRI – праймеры: 5’-GACTGCGTACCAATTCGW-3’ 5’- GACTGCGTACCAATTCTS-3’ MseI-праймеры: 5’- GATGAGTCCTGAGTAAAAS-3’ 5’- GATGAGTCCTGAGTAAASA-3’ 5’-GATGAGTCCTGAGTAAATN-3’ 5’-GATGAGTCCTGAGTAACAN-3’ 5’-GATGAGTCCTGAGTAACTN-3’ EcoRI-праймеры: 5’-GACTGCGTACCAATTCAN-3’ 5’-GACTGCGTACCAATTCCW-3’ 5’-GACTGCGTACCAATTCTW-3’ MseI-праймеры: 5’- GATGAGTCCTGAGTAAAAS-3’ 5’- GATGAGTCCTGAGTAAASA-3’ 5’-GATGAGTCCTGAGTAAGAY-3’ 5’-GATGAGTCCTGAGTAAGTW-3’ 5’-GATGAGTCCTGAGTAATCG-3’ 5’-GATGAGTCCTGAGTAATCT-3’ 5’-GATGAGTCCTGAGTAATGW-3’ EcoRI-праймеры: 5’-GACTGCGTACCAATTCGW-3’ 5’-GACTGCGTACCAATTCTN-3’ MseI-праймеры: 5’-GATGAGTCCTGAGTAAGAW–3’ 5’-GATGAGTCCTGAGTAAGCW–3’ 5’-GATGAGTCCTGAGTAAGTW-3’ 5’-GATGAGTCCTGAGTAATAN-3’ 5’-GATGAGTCCTGAGTAATCW-3’ 5’-GATGAGTCCTGAGTAATGW-3’ 5’-GATGAGTCCTGAGTAATTS-3’ В общей сложности было получено 83 Col-специфичных фрагмента, из которых 62 были общими для обоих клонов YAC. Три фрагмента представляли собой ПДАФ-маркеры полиморфизма между Ws и Ler, два из которых были ПДАФ-маркерами Ws (Col-фрагмент также присутствует в Ws и отсутствует в Ler), а один представлял собой ПДАФ-маркер Ler (Col-фрагмент также присутствует в Ler а отсутствует Ws). Эти результаты представлены ниже в таблице 5. В таблице 5 показано количество общих и уникальных ПДАФ-фрагментов, обнаруженных в клонах YAC 10G07 и 7Е03, и количество информативных ПДАФ-маркеров среди этих фрагментов в генотипах Ws и Ler.
Таким образом, ПДАФ-анализ позволил получить три новых ПДАФ-маркера (см. фиг.4 и далее по тексту). Их положения относительно друг друга и относительно ПДАФ-маркеров, полученных из рекомбинантной инбредной линии, определяли с помощью анализа рекомбинантов с этими ПДАФ-маркерами. Второй скрининг ПДАФ-маркеров проводили, анализируя все четыре выявленных клона YAC (см. ниже) и используя комбинацию ферментов PstI/MseI. Применяли следующие праймеры: PstI-праймеры: 5’-GACTGCGTACATGCAGAN-3’ 5’-GACTGCGTACATGCAGCW-3’ 5’-GACTGCGTACATGCAGGW-3’ 5’-GACTGCGTACATGCAGTN-3’ MseI-праймеры: 5’-GATGAGTCCTGAGTAAAN-3’ 5’-GATGAGTCCTGAGTAACW-3’ 5’-GATGAGTCCTGAGTAAGW-3’ 5’-GATGAGTCCTGAGTAATN-3’ Символ “N” в праймерах обозначает вариабельную часть последовательности (А, С, G или Т), и “W” в праймерах обозначает А или Т. Всего использовали 144 (12×12) комбинации праймеров для скрининга всех четырех выделенных клонов YAC: 12F04, 12Н07, 10G07 и 7Е03, штамма дрожжей, несодержащего YAC, и трех экотипов Arabidopsis Col, Ler и Ws. Всего было получено 219 ПДАФ-фрагментов, из которых 144 присутствовали в клонах YAC 12F04 и 12Н07 (72 оказались уникальными для клона 12F04 и 72 оказались общими для обоих клонов YAC) и из которых 75 присутствовали в клонах YAC 10G07 и 7Е03 (33 оказались уникальными для клона 10G07 и 42 оказались общими обоих клонов YAC). Три фрагмента, полученных из набора клонов YAC, были полиморфными (ПДАФ-маркеры Ws). Эти результаты представлены ниже в таблице 6. В таблице 6 показано количество общих и уникальных ПДАФ-фрагментов, выявленных в YAC, и из этих фрагментов количество информативных ПДАФ-маркеров для генотипов Ws и Ler.
Результаты свидетельствуют о том, что клон YAC 12H07 представляет собой часть более крупного клона YAC 12F04, а клон YAC 7E03 представляет собой часть более крупного клона YAC 10G07. Эти данные свидетельствуют о том, что более крупные клоны YAC 12F04 и 10G07 не перекрываются. Полученные данные не позволяют определить положение гена NIM1 на любом из указанных клонов YAC. Полный скрининг, включающий 400 комбинаций праймеров, который дал 302 ПДАФ-фрагмента в NIM-области, позволил получить пять пригодных ПДАФ-маркеров, четыре из которых оказались Ws-специфичными, а один – Ler-специфичным. Положение на карте этих 5 дополнительных ПДАФ-маркеров было определено с помощью анализа рекомбинантных растений (см. фиг.4 и далее по тексту), и они обозначены как W84.1 (a.k.a. W83.3), W84.2, W85.1, W86.1 и L86. В таблице 7 приведены последовательности праймеров, которые применяли для получения этих ПДАФ-маркеров. Эти 5 дополнительных ПДАФ-маркеров увеличили общее число ПДАФ-маркеров до 12 в области от L81.1 до L88 (см. фиг.4 и далее). В таблице 7 приведены комбинации праймеров ПДАФ-маркеров, полученных из клонов YAC. “EcoRI-” относится к последовательности 5’-GACTGCGTACCAATTC-3’, “MseI-” относится к последовательности 5’-GATGAGTCCTGAGTAA-3’ и “PstI-” относится к последовательности 5’-GACTGCGTACATGCAG-3’.
Эту информацию использовали для создания физической карты области, как показано на фиг.5, с приблизительными положениями клонов YAC на генетической карте. На карте показано, что область, содержащая локус NIM1 между маркерами W83.1 и W85.1, частично перекрыты тремя клонами YAC: 12F04 и 10G07/7E03. III. Конструирование набора последовательностей фрагментов Р1/ВАС, содержащего ген NIM1 В предыдущих разделах описано, каким образом выделяли ПДАФ-маркеры, связанные с NIM1-областью, и каким образом выявляли и картировали соответствующие этим маркерам клоны YAC. Результаты, полученные при локализации гена NIM1 на фрагменте хромосомы, не позволили определить специфический сегмент ДНК, содержащий ген NIM1: фланкирующие ПДАФ-маркеры картировали различные клоны YAC, которые не перекрывались. Поэтому оказалось невозможным определить точное физическое положение гена NIM1; он мог быть локализован либо на любом из двух YAC, либо в бреши между YAC. Для закрытия физической бреши между фланкирующими маркерами был выбран альтернативный подход: библиотеку Р1 и ВАС применяли для перекрытия бреши между фланкирующими ПДАФ-маркерами. IV. Идентификация клонов Р1, соответствующих фланкирующим маркерам Для скрининга групп клонов Р1 применяли фланкирующие маркеры Ws84.2 и Ws85.1, используя стратегию, описанную ранее для скрининга библиотеки YAC (см. пример 1.51). Отбирали клоны Р1, имеющие маркерные фрагменты, и выделяли “плазмидную” ДНК. Получали фингерпринты ДНК различных Р1-клонов, используя КФ EcoRI/MseI и HindIII/MseI и праймеры без селективных нуклеотидов. Путем сравнения ПДАФ-фингерпринтов создавали физическую карту, т.е. карту, на которой представлены размер и перекрытия клонов. Количество ПДАФ-фрагментов, которые являются уникальными, и количество ПДАФ-фрагментов, общих для клонов, характеризует степень перекрытий. Карта приведена на фиг.6. Получение ПДАФ-фингерпринтов показало, что было сконструировано два набора неперекрывающихся последовательностей Р1, каждый из которых содержал один из следующих фланкирующих маркеров: Р1-1 и Р1-2, включающие маркер Ws84.2, и Р1-3 и Р1-4, включающие маркер W85.1. Следовательно, брешь между фланкирующими маркерами не была закрыта (фиг.6). Положение набора последовательностей Р1 по отношению к набору последовательностей YAC определяли, получая ПДАФ-фингерпринты клонов YAC и Р1 с указанным выше количеством YAC-специфичных КП. Клоны Р1 Р1-1 и Р1-2, вероятно, полностью перекрывались клоном YAC CIC12F04, но только частично клоном YAC CIC12H07. Таким образом, последние клоны Р1 могут располагаться на наборе последовательностей YAC CIC12H07/12F04 (фиг.6). Клоны PI P1-3 и Р1-4 полностью перекрывали оба клона YAC CIC7E03 и CIC10G07 и, вероятно, что ПДАФ-маркер W86.1, а также W85.1 картирован в этом же наборе последовательностей Р1 (фиг.6). Далее маркер L85 применяли для идентификации соответствующих клонов Р1 и ВАС. L85 представляет собой специфичный для экотипа Landsberg маркер, и поэтому осуществляли гибридизацию колонии радиоактивномеченной ДНК L85 с Р1- и ВАС-фильтрами. Не был выявлен ни один отдельный клон Р1 или ВАС, гибридизующийся с L85. Это подтверждает полученные ранее данные о том, что последовательность L85 отсутствует в геноме экотипа Arabidopsis Columbia, что является наиболее вероятным объяснением того, что не были выявлены соответствующие клоны. V. Удлинение фланкирующих NIM1 наборов последовательностей Р1 Для удлинения фланкирующих наборов последовательностей Р1 использовали различные подходы. ПДАФ-фрагменты YAC, специфичные относительно Саузерн-конца клона YAC CIC12F04 (уникальные для CIC12F04, отсутствуют в CIC 12H07), применяли для идентификации клонов Р1 с помощью ПДАФ-скрининга групп библиотек. 1. ПДАФ-фрагменты YAC из клона YAC 10G07, которые перекрывались Р1-4, применяли для идентификации клонов Р1 с помощью ПДАФ-скрининга набора библиотек PL 2. EcoRI-рестрикционные фрагменты из клона Р1-6 (полученного в результате стадии 1 скрининга библиотеки Р1, основанного на использовании ПДАФ) использовали в качестве зондов для гибридизации с библиотекой ВАС на фильтрах. В результате этого скрининга были получены различные клоны Р1 и ВАС, и для них всех получали ПДАФ-фингерпринты, используя КФ EcoRI/MseI и HindIII/MseI и праймеры без селективных нуклеотидов. Создавали новую карту, как описано выше, которая приведена на фиг.7. В таблице 8 представлены различные ПДАФ-КП, имеющие ПДАФ-фрагменты, которые картированы на фланкирующих YAC и которые использовали для скрининга библиотеки Р1 в отношении соответствующих Р1-клонов. В таблице 8 представлены различные ПДАФ-КП, использованные для скрининга библиотеки Р1. В верхней части таблицы приведены КП, специфичные для Нозерн-концов YAC, а в нижней части таблицы приведены КП, специфичные для Саузерн-концов YAC. Также указаны клоны YAC и Р1, в которых обнаружены ПДАФ-фрагменты. Получали наборы последовательностей Р1/ВАС длиной приблизительно 250 т.п.н., покрывающие Саузерн-конец клона YAC CIC12F04 (который не выходил за пределы этого клона YAC) и который содержал маркер W84.2. Получали набор последовательностей Р1 длиной приблизительно 150 т.п.н., содержащий маркеры W85.1 и W86.1; этот набор последовательностей полностью находится внутри клона YAC CIC7E03. Анализ с помощью ПДАФ-маркеров конструкции, включающей набор последовательностей Р1/ВАС, покрывающий ген NIM1, на рекомбинантах с маркерами из Саузерн-конца Нозерн-набора последовательностей Р1/ВАС (WL84.4 и WL84.5, см. далее по тексту и в таблице 11) показал, что проведенные ранее стадии “прогулки по хромосоме” оказались неуспешными для конструирования набора последовательностей, содержащего ген NIM1 (см. следующий раздел). Поэтому существующий Нозерн-набор последовательностей Р1/ВАС удлиняли с Саузерн-конца с целью “прогулки” поперек гена NIM1, что должно было позволить выявить и выделить специфический сегмент ДНК, содержащий ген NIM1. Далее был использован подход с использованием гибридизации, в котором клоны Р1 или ВАС, локализованные на Саузерн-конце Нозерн-набора последовательностей РР1/ВАС, использовали для выявления клонов, расположенных более близко к NIM1 (Саузерн-пограничная последовательность). Новые клоны, полученные в результате стадий “прогулки”, картировали по отношению к существующим наборам последовательностей, используя полученные ПДАФ-фингерпринты с помощью КФ EcoRI/MseI и HindIII/MseI, как описано выше. По-видимому, для “пересечения” гена NIM1 необходимо всего 5 последовательных стадий “прогулки”. В таблице 9 приведены клоны, полученные в результате различных стадий “прогулки”. В таблице 9 для различных стадий “прогулки” приведены зонды гибридизации, которые применяли для скрининга библиотек Р1 и ВАС, и выбранные клоны, гибридизующиеся с зондами и удлиняющие последовательность в направлении Саузерн-конца.
Физическая карта различных клонов, полученных в результате этих стадий “прогулки”, приведена на фиг.8. В целом было проанализировано расстояние, составляющее приблизительно 600 т.п.н., начиная от начальной точки “прогулки” маркера W84.2. Саузерн-конец набора последовательностей, приведенного на фиг.8, вероятно, содержит ген NIM1 (см. следующий раздел). Набор последовательностей, простирающийся более чем на 300 т.п.н. к Саузерн-концу от клона YAC CIC12F04 и, вероятно, не перекрывающийся с клонами YAC CIC10G07 и CIC7E03, свидетельствует о том, что ген NIM1 расположен в бреши между фланкирующими YAC наборами последовательностей и что эта брешь имеет размер по крайней мере 300 т.п.н. VI. Создание обобщенной генетической и физической карты NIM1-области В предыдущих разделах было описано, каким образом выделяли ПДАФ-маркеры, связанные с NIM1-областью, каким образом выявляли клоны YAC, соответствующие фланкирующим маркерам, и каким образом конструировали набор последовательностей Р1/ВАС, простирающийся приблизительно на 550 т.п.н. к Саузерн-концу от ближайшего к Нозерн-концу фланкирующего ПДАФ-маркера W84.2. В данном разделе описано получение новых ПДАФ-маркеров из набора последовательностей Р1/ВАС, физическое картирование этих маркеров на этом наборе последовательностей и генетическое картирование этих маркеров с помощью доступных рекомбинантов. 1. Получение новых ПДАФ-маркеров из набора последовательностей Р1/ВАС Как описано в предыдущем разделе, набор последовательностей, полученных в результате удлинения клонов Р1 и ВАС, характеризовали с помощью полученных на основе ПДАФ фингерпринтов с использованием КФ EcoRI/MseI и HindIII/MseI. Этот метод позволяет достаточно точно определить протяженность перекрывающихся областей различных клонов Р1 и ВАС, а также получить ряд ПДАФ-фрагментов, специфичных для этих клонов. ПДАФ-праймеры без селективных нуклеотидов применяли для получения фингерпринтов очищенной плазмидной ДНК клонов Р1 или ВАС. Селективные нуклеотиды, однако, являются необходимыми для того, чтобы сделать возможным применение этих Р1- или ВАС-специфичных ПДАФ-фрагментов для выявления в Arabidopsis. Путем определения концевых последовательностей амплифицированных рестрикционных фрагментов могут быть сконструированы ПДАФ-праймеры, имеющие соответствующие селективные основания, для амплификации Р1- или ВАС-специфичного ПДАФ-фрагмента в Arabidopsis. Все ПДАФ-фрагменты происходят из экотипа Columbia (Col) и, следовательно, также могут быть выявлены, если ПДАФ-маркеры Columbia являются информативными в рекомбинантах NIM1, которые получают в результате скрещивания экотипов Landsberg erecta (Ler) и nim1-мутанта экотипа Wassilewskija (Ws-nim). В принципе существует 4 типа ПДАФ-фрагментов, два из которых являются пригодными маркерами, как показано ниже в таблице 10. В таблице 10 приведены типы обнаруженных ПДАФ-маркеров. (+) или (-) обозначает присутствие или отсутствие ПДАФ-фрагмента.
В целом получение фингерпринтов клонов Р1 и ВАС давало 30-40 EcoRI/MseI-ПДАФ-фрагментов и 60-80 HindIII/MseI-ПДАФ-фрагментов для каждого отдельного клона. Концевые последовательности отдельных фрагментов определяли стандартными методами секвенирования. Затем тестировали наборы специфичных ПДАФ-праймеров с селективными удлинениями, состоящими из трех нуклеотидов, для обоих EcoRI- или HindIII-праймеров и для Msel-праймера с помощью набора следующих ДНК: 1. ДНК клона Р1/ВАС, из которого происходил ПДАФ-маркер; 2а. ДНК дрожжей; 2б. ДНК клона YAC CIC12F04 (только для ПДАФ-фрагмента из Р1-7); 2в. ДНК клона YAC CIC10G07; 3а. ДНК Col, которая является источником библиотек Р1 и ВАС; 3б. ДНК Ler, родителя 1 рекомбинантов nim; 3в. ДНК Ws-nim, родителя 2 рекомбинантов nim. Было отобрано 6 клонов для анализа последовательности их EcoRI/MseI- и HindIII/MseI-ПДАФ-фрагментов: ВАС-01/Р1-7, Р1-17/Р1-18, ВАС-04/ВАС-06. Все ПДАФ-фрагменты из клона Р1-7 были обнаружены в YAC CIC12F04, свидетельствуя о том, что этот клон полностью содержится внутри этой YAC. Ни один из Р1/ВАС-специфичных ПДАФ-фрагментов не был обнаружен в клоне YAC CIC10G07, свидетельствуя о том, что набор последовательностей Р1/ВАС не является мостом для бреши между двумя фланкирующими YAC наборами последовательностей. ПДАФ-маркеры, выбранные для анализа рекомбинантов nim, приведены в таблице 11. В таблице 11 приведены выбранные ПДАФ-маркеры из ПДАФ-КП, специфичных для различных клонов Р1 и ВАС. “WL”-маркер представляет собой маркер, полученный из этой же КП и соответствует двум ПДАФ-маркерам, т.е. Ws- и Ler-маркеру, которые, вероятно, полностью связаны в фазе отталкивания при анализе рекомбинантов NIM.
2. Физическое картирование новых ПДАФ-маркеров Описанные выше ПДАФ-маркеры картировали на физической карте путем обнаружения их присутствия в различных клонах Р1 и ВАС. Результаты приведены на фиг.9-11. 3. Генетическое картирование новых ПДАФ-маркеров Все ПДАФ-маркеры проанализированы в отобранном наборе рекомбинантов. Полученные результаты обобщены в таблицах 12а, 12б и 12в. ПДАФ-маркеры Ler84.8, Ler84.9a, Ler84.9b и Ler84.9c видны на карте на Саузерн-стороне NIM1. Были обнаружены рекомбинанты, которые фенотипически проявлялись как nim1 (гомозиготы, генотип Ws-nim1/Ws-nim1) и гетерозиготные по этим ПДАФ-маркерам (был обнаружен Ler-специфичный ПДАФ-маркер, генотип Ws-nim1/Ler). ПДАФ-маркер Ler84.8 оказался ближайшим к NIM1: только один рекомбинант (С-105) был классифицирован как гетерозиготный Ws-nim1/Ler и гомозиготный Ws-nim1/Ws-nim1. ПДАФ-маркеры Ler84.7 и Ler84.6c оказались полностью косегрегированными с NIM1: все рекомбинанты имели идентичный генотип NIM1 и ПДАФ-маркер. Расположенный ближе к Нозерн-концу от NIM1 маркер L84.6b оказался ближайшим к NIM1: обнаружено три рекомбинантных растения с фенотипом nim1, C-074, D-169 и Е-103 (таблица 12в), которые являются гетерозиготными Ws-nim1/Ler по этому маркеру. С помощью космидного набора последовательностей, полученного из Р1-18, ВАС-04 и ВАС-06, AFLP-маркеры Ler84.6b и Ler84.8 были картированы в Р1-18 и ВАС-04 соответственно и обнаружено, что физическое расстояние между ними составляет приблизительно 110 т.п.н. Это позволяет установить, что nim1 должен быть локализован на сегменте ДНК, длина которого составляет 110 т.п.н. Из этого анализа установлено, что ген NIM1 содержится в клоне ВАС-04 или Р1-18. Клоны ВАС-04 и Р1-18 были депонированы в АТСС и им присвоены регистрационные номера АТСС 97543 и АТСС 97606 соответственно. VII. Генетическое и физическое тонкое картирование гена NIM1 В предыдущем разделе описано, каким образом сегмент ДНК, содержащий ген NIM, был определен с помощью физического картирования фланкирующих ПДАФ-маркеров (Ler84.6b и Ler84.8) на наборе последовательностей Р1/ВАС. Фланкирующие маркеры видны на карте на двух перекрывающихся клонах Р1-18 и ВАС-04. В данном разделе описан путь получения дополнительных ВАС-04-специфичных и Р 1-18-специфичных ПДАФ-маркеров с целью увеличения разрешающей способности генетического и физического картирования в области, содержащей ген NIM1. VIII. Получение дополнительных ПДАФ-маркеров из космидного набора последовательностей С целью получения дополнительных ПДАФ-маркеров для тонкого картирования NIM1 было отобрано 4 КФ: PstI/MseI, XbaI/MseI, BstYI/MseI и TaqI/MseI. PstI/MseI- и XbaI/MseI-ПДАФ-фрагменты получали из клонов Р1-18 и ВАС-04 и определяли селективные последовательности, необходимые для выявления у Arabidopsis. Аналогично этому ПДАФ-фрагменты и селективные последовательности определяли для BstYI/MseI и TaqI/MseI, но в этом случае процедуру проводили с использованием космидных ДНК: A11, C7, Е1 и Е8 для BstYI/MseI (полная NIM1-область) и D7, Е8 и Е6 для TaqI/MseI (Саузерн-сторона NIM1-области). Информативные ПДАФ-маркеры, отобранные для дальнейшего генетического и физического картирования, приведены в таблице 13. Дополнительные адаптеры, которые использовали в этой работе, приведены в таблице 14. В таблице 13 приведены ПДАФ-маркеры, которые использовали для генетического и физического тонкого картирования NIM1. “BstYI(T)” обозначает, что сайт рестрикции и соответствующий праймер представляли собой либо AGATCT, либо GGATCT.
В таблице 14 приведены эти же дополнительные адаптеры, которые использовали для идентификации новых ПДАФ-маркеров. IX. Физическое картирование новых ПДАФ-маркеров в космидных наборах последовательностей Осуществляли физическое картирование указанных выше маркеров в наборе космид, определяя их присутствие в различных космидных клонах (фиг.11). 1. Генетическое картирование новых ПДАФ-маркеров Для новых ПДАФ-маркеров проводили генетическое картирование с помощью ПДАФ-анализа ближайших Нозерн- и Саузерн-рекомбинантов. Картировали ближайшие к Нозерн-концу (рекомбинант D169) и к Саузерн-концу (рекомбинант С105) точки рекомбинации (см. таблицу 15). ПДАФ-анализ показал, что рекомбинант D169 имел Саузерн-рекомбинацию маркера L84.Y1, но Нозерн-рекомбинацию маркера W84.Y2. Точка рекомбинации в рекомбинанте С105 картирована между маркерами L84.T2 и L84.8. С помощью доступного набора рекомбинантов удалось дополнительно определить хромосомный интервал, содержащий NIM1; расстояние между фланкирующими точками рекомбинаций, по-видимому, составляет 60-90 т.п.н. (фиг.12). 2. Конструирование космидного набора последовательностей Для комплементации растительного фенотипа nim1 необходима трансформация nim1-растений геном NIM1 дикого типа. Это можно осуществить трансформацией этих растений космидой, содержащей этот ген. Для этой цели конструировали космидный набор последовательностей NIM1-области. Поскольку Arabidopsis трансформируют с помощью Agrobacterium, использованный космидный вектор представлял собой бинарный вектор. ДНК выделяли из ВАС-04, ВАС-06 и Р1-18 и осуществляли частичное расщепление с помощью рестриктазы SauSAI. Фрагменты длиной 20-25 т.п.н. выделяли в градиенте сахарозы, объединяли и заполняли с помощью дАТФ и дГТФ. Бинарный вектор (04541) расщепляли с помощью XhoI и заполняли дЦТФ и дТТФ. Затем фрагменты лигировали с вектором. Лигированную смесь упаковывали и трансдуцировали в E.coli. Осуществляли скрининг космидной библиотеки клонами ВАС-04, ВАС-06 и Р1-18 и выделяли позитивные клоны. У этих космид затем определяли ПДАФ-фингерпринты и из них формировали набор последовательностей перекрывающихся клонов, охватывающих NIM1-область. Определяли размеры космидных вставок и проводили ограниченное картирование с помощью рестриктаз. Результаты приведены на фиг.10. Пример 2: Выявление клона, содержащего ген NIM1 1. Комплементация с помощью стабильной трансформации Космиды, полученные из описанных выше клонов, которые охватывали NIM1-область (описанные выше) вносили в Agrobacterium путем трехродительского скрещивания. Эти космиды затем использовали для трансформации nim1-Arabidopsis с помощью вакуумной инфильтрации (Mindrinos и др., 1994, Cell 78, 1089-1099) или путем стандартной трансформации корня. Семена этих растений собирали и проращивали на агаровых пластинках с канамицином (или с другим пригодным антибиотиком) в качестве селектирующего агента. Только трансформированные космидной ДНК проростки могли детоксицировать селектирующий агент и выживать. Ростки, выжившие при такой селекции, переносили в почву и тестировали в отношении фенотипа nim или их потомство тестировали в отношении фенотипа nim. Трансформированные растения, которые более не обладали фенотипом nim, соответствовали космиде(ам), содержащей (им) функционально активный ген NIM1. 2. Комплементация во временной системе экспрессии Способность клонов ДНК восстанавливать (комплементировать) мутацию nim1 тестировали в двух временных системах экспрессии. В первой системе растения Arabidopsis с мутацией nim1, содержащие трансген PR1-люцифераза (PR1-lux), применяли в качестве реципиентного материала при бомбардировках. Эти растения получали путем трансформации растений экотипа Columbia с помощью конструкции PR1-lux путем вакуумной инфильтрации с последующей селекцией собранных семян на устойчивость к канамицину, как описано выше. Осуществляли самоопыление трансформированных растений, которые экспрессировали люциферазную активность после индукции ИНК, и получали гомозиготные растения. Их скрещивали с nim1-растениями. При экспресс-анализе потомство растений, полученное в результате этого скрещивания, которое являлось гомозиготным по генам nim1 и PR1-lux, использовали для идентификации клонов ДНК, которые могут восстанавливать фенотип nim1. В этом случае растения сначала обрабатывали ИНК, как описано выше в примере 1.1. Через 2 дня эти растения собирали, стерилизовали их поверхность и высаживали на агаровую среду GM. Затем листовую ткань подвергали бомбардировке космидой, клонами Р1 или ВАС (или подклонами) из NIM1-области и через один день измеряли люциферазную активность в листьях. Клоны, которые индуцируют люциферазную активность, содержат ген NIM1. Во второй системе растения с мутацией nim1 обрабатывали ИНК (как описано выше в примере 1.1) и через 2 дня подвергали бомбардировке клонированной ДНК (космидной, клонов Р1, ВАС и/или YAC или субклонов) из локуса NIM1-области и репортерной плазмидой. Репортерная плазмида содержала ген люциферазы, находящийся под контролем промотора PR1 Arabidopsis (PR1-lux). В nim1-растениях ИНК не активировала промотор PR1 (как описано выше в примере 1.2) и, таким образом, не могла индуцировать люциферазную активность репортерной плазмиды. Однако, когда совместно трансформированный клон ДНК содержал восстанавливаемый ген NIM1, ИНК не индуцировали промотор PR1, что видно по индукции люциферазной активности. Через один день после совместной бомбардировки измеряли люциферазную активность в целом растении. Клоны ДНК (космиды, клоны Р1 или ВАС либо субклоны), которые индуцировали люциферазную активность, существенно превышающую основные уровни, содержат ген NIM1. 3. Изменения транскриптов в линиях с фенотипом nim1 Поскольку растения с фенотипом nim1 имеют мутации в гене NIM1, представляется вероятным, что в некоторых линиях ген изменен таким образом, что в нем не транскрибируется мРНК или продуцируется отклоняющаяся от нормы (по размеру) мРНК. Для оценки этого факта в линиях nim1 проводили блоттинг-анализ РНК. РНК выделяли из растений экотипов Ws и Ler этих линий (после обработки водой, ИНК или БТК) и применяли для получения Нозерн-блотов. Эти блоты гибридизовали с фрагментами ДНК, выделенными из клонов наборов последовательностей ДНК локуса NIM1. Фрагменты ДНК, которые характерны для линий nim1 с отклоняющейся от нормы экспрессией РНК (отклоняющейся от нормы по размеру или по концентрации), вероятно, идентифицируют ген NIM1 (или часть его). Фрагмент ДНК и окружающуюся область ДНК секвенировали и применяли для выделения кДНК (путем скрининга библиотеки или с помощью ПЦР с обратной транскрипцией), которую также секвенировали. Клон, из которого выделяли фрагмент, или выделенную кДНК применяли для подтверждения комплементации фенотипа nim1 в стабильных и временных системах экспрессии. Пример 3: Определение последовательности ДНК гена NIM1 1. Геномное секвенирование Геномные клоны, которые могут содержать ген NIM1, секвенировали с помощью методов, известных в данной области. Они включают ВАС-04, Р1-18 и космиды из NIM1-области. Например, космиды расщепляли с помощью ферментов рестриктаз и фрагменты, полученные из вставки, клонировали в векторе общего назначения, таком, как pUC18 или Bluescript. Наиболее крупные клоны Р1 и ВАС случайным образом отщепляли и фрагменты клонировали в векторе общего назначения. Фрагменты в этих векторах секвенировали общепринятыми методами (например, с помощью “праймерной прогулки” или с помощью создания делеций вставок). Полученные последовательности объединяли в наборы последовательностей. Последовательность вставки комплементируемого клона содержит ген NIM1. Приблизительные начало и конец гена NIM1 выводили дедукцией, основываясь на обощенных данных о последовательности ДНК, последовательности мотивов, таких, как ТАТА-боксы, открытых рамок считывания, присутствующих в последовательности, обычных кодонов, данных о космидной комплементации, об относительном положении ПДАФ-маркеров и дополнительных доступных данных (см. пример 4, ниже). 2. Секвенирование кДНК Космиду(ы) или более крупные клоны, которые содержат ген NIM1 (как описано выше в примере 2), применяли для выделения кДНК. При этом использовали клоны (или фрагменты ДНК) в качестве зондов при скрининге библиотеки кДНК растений Arabidopsis дикого типа. Выделенные кДНК секвенировали, как описано для секвенирования космид, и использовали в тестах по комплементации. Для этого полноразмерные кДНК клонировали в приемлемом растительном векторе экспрессии под контролем конститутивного промотора. Эти конструкции использовали для экспресс-анализов, как описано ранее. В альтернативном варианте кДНК клонировали в бинарном векторе экспрессии, предназначенном для экспрессии в растительных тканях и для опосредованной Agrobacterium трансформации растений, как описано выше в примере 2. Секвенировали кДНК, содержащую ген NIM1 (что определено по комплементации, выделению с помощью близко связанного ПДАФ-маркера, выделению с помощью космидного фрагмента или с помощью другого дедуктивного анализа). Гены из растений Ws-O и nim1 выделяли и секвенировали. Гены получали из космид библиотеки кДНК с помощью фрагмента выделенного гена NIM1 в качестве зонда. В альтернативном варианте гены или кДНК выделяли с помощью ПЦР, используя в качестве матрицы специфичные для гена NIM1 праймеры и геномную ДНК или кДНК. Аналогичным образом выделяли и секвенировали аллели nim1 из других линий nim1 (см. пример 1.1, выше). Пример 4: Описание гена NIM1 и выведенной путем дедукции последовательности протеина Последовательность ДНК гена NIM1 или кДНК определяли по описанной выше в примере 3 методике. Эту последовательность анализировали с помощью программ анализа ДНК, например, программ, входящих в пакет программ Genetics Computer Group (GCG), в пакет программ Sequencer или Staden или в любой другой пакет программ для анализа ДНК. В частности, начало и конец гена определяли на основе анализа открытой рамки считывания, присутствия стоп-кодона и потенциального стартового кодона, присутствия мотивов потенциального промотора (таких, как ТАТА-бокс), присутствия сигналов полиаденилирования и т.п. Предсказанную аминокислотную последовательность также выводили методом дедукции на основе данных об открытой рамки считывания. Последовательность как ДНК, так и протеина применяли для исследования баз данных гомологичных последовательностей, таких, как факторы транскрипции, ферменты или мотивы этих генов или протеинов. Пример 5: Выделение гомологов гена NIM1 Ген NIM1 Arabidopsis, может применяться в качестве зонда при скрининге гибридизацией в условиях пониженной жесткости библиотеки геномной ДНК или кДНК с целью выделения гомологов NIM1 из других видов растений. В альтернативном варианте для этого применяли ПЦР-амплификацию с использованием праймеров, созданных на основе последовательности гена NIM1 Arabidopsis, и с использованием геномной ДНК или кДНК в качестве матрицы. Ген NIM1 может быть выделен из кукурузы, пшеницы, риса, ячменя, семян рапса, сахарной свеклы, картофеля, томата, бобов, огурцов, винограда, табака и других представляющих интерес культур, последовательности которых известны. Имея в распоряжении набор последовательностей гомологов гена NIM1, с помощью ПЦР-амплификации могут быть созданы новые праймеры из консервативных областей гена с целью выделения гомологов NIM1 из более отдаленных с точки зрения родства видов растений. Пример 6: Комплементация гена nim1-1 с помощью геномных Фрагментов 1. Конструирование космидного набора последовательностей Космидный набор последовательностей NIM1-области конструировали с помощью CsCl-очищенной ДНК из ВАС04, ВАС06 и Р1-18. ДНК.трех клонов смешивали в эквимолярных количествах и частично расщепляли с помощью рестриктазы Sau3A. Фрагменты длиной 20-25 т.п.н. выделяли в градиенте сахарозы, объединяли и заполняли с помощью дАТФ и дГТФ. Плазмиду pCLD04541 использовали в качестве Т-ДНК космидного вектора. Эта плазмида содержит репликон с широким спектром хозяев, имеющий в основе pRK290, ген устойчивости к тетрациклину для селекции бактерий и ген nptII для селекции растений. Вектор расщепляли с помощью XhoI и заполняли дЦТФ и дТТФ. Затем полученные фрагменты лидировали с вектором. Лигированную смесь упаковывали и трансдуцировали в штамм Е. coil XL1-blue MR (фирма Stratagene). Осуществляли скрининг образовавшихся трансформантов путем гибридизации с клонами ВАС04, ВАС06 и Р1-18 и выделяли позитивные клоны. Космидную ДНК выделяли их этих клонов и получали ДНК-матрицу, используя КФ EcoRI/MseI и HindIII/MseI. Схему полученных ПДАФ-фингерпринтов анализировали с целью определения порядка космидных клонов. Отбирали набор из 15 полуперекрывающихся космид, охватывающих nim-область (фиг.13). Космидные ДНК также расщепляли с помощью EcoRI, PstI, BssHII и SgrAI. Это позволило оценить размеры космидных вставок и подтвердить наличие перекрывающихся участков между различным космидами, что установлено с помощью ПДАФ-фингерпринтов. 2. Идентификация клона, содержащего ген NIM Космиды, полученные из клонов, охватывающих NIM1-область, встраивали в Agrobacterium tumefaciens AGL-1 путем переноса конъюгантов методом трехродительского спаривания с помощью штамма-хелпера НВ101 (pRK2013). Затем эти космиды использовали для трансформации чувствительной к канамицину линии nim1-1 Arabidopsis с помощью вакуумной инфильтрации (Mindrinos и др., 1994, Cell 78, 1089-1099). Семена зараженных растений собирали и проращивали на агаровых GM-пластинках, содержащих 50 мг/мл канамицина в качестве селектирующего агента. Только проростки, трансформированные космидной ДНК, могли осуществлять детоксикацию селектирующего агента и выживать. Приблизительно через две недели после посева ростки, выжившие при такой селекции, переносили в почву и тестировали в отношении фенотипа nim согласно описанному выше методу. Трансформированные растения, которые более не обладали фенотипом nim, соответствовали космиде(ам), содержащей (им) функционально активный ген NIM1. 3. Анализ фенотипа nim1 трансформантов Перенесенные в почву растения выращивали в фитотроне в течение приблизительно одной недели после пересадки. 300 мкМ ИНК вносили в виде легкого тумана, полностью покрывающего растения, используя мелкокапельный туманообразователь типа “chromister”. Через 2 дня листья собирали для экстракции РНК и анализа экспрессии генов PR-1. Затем растения опрыскивали Peronospora parasitica (изолят EmWa) и выращивали в условиях высокой влажности в климатической камере при дневной температуре 19°С/ночной температуре 17° и при световом цикле 8 ч света/16 ч темноты. Через 8-10 дней после заражения грибом растения исследовали и на основе данных о росте гриба оценивали как позитивные или негативные. Растения линий-Ws, и nim1, обработанные таким же образом, служили в качестве контролей в каждом эксперименте. Общую РНК экстрагировали из собранной ткани, используя буфер для экстракции LiCl/фенол (Verwoerd и др., NAR 17: 2362). Образцы РНК разгоняли на формальдегид/агарозном геле и блоттировали на мембранах типа GeneScreen Plus (фирма DuPont). Блоты гибридизовали с зондом, который представлял собой меченную с помощью 32Р кДНК PR-1. Полученные блоты экспонировали на пленку с целью определения, какие трансформанты обладали способностью экспрессировать PR-1 после обработки ИНК. Результаты обобщены в таблице 16. В таблице 16 показана комплементация фенотипа nim1 космидными клонами.
Пример 7: Секвенирование, области гена NIM1 длиной 9.9 т.п.н. ДНК ВАС04 (25 мкг, полученную от фирмы Key Gene) использовали в качестве источника ДНК для анализа последовательности. Было показано, что этот клон ВАС представляет собой клон, который полностью охватывает область, комплементарную мутантам nim1. ДНК случайным образом расщепляли с помощью метода, разработанного в лаборатории Cold Spring Harbor. В целом этот метод состоит в том, что ДНК клона ВАС расщепляли в распылителе до средней молекулярной массы приблизительно 2 т.п.н. Концы отщепленных фрагментов репарировали с использованием двухстадийного протокола с помощью дНТФ, полимеразы фага Т4 и фрагмента Кленова (фирма Boehringer). ДНК с репарированными концами разгоняли на 1%-ном агарозном геле с низкой температурой плавления и вырезали из геля область от 1,3 т.п.н. до 2,0 т.п.н. ДНК выделяли из фрагмента геля с помощью метода замораживания-оттаивания. Затем ДНК смешивали с расщепленным с помощью EcoRV pBRKanF4 и лигировали в течение ночи при 4°С. Плазмида pBRKanF4 является производной pBRKanF1, полученной от Kolavi Bhat из Университета Вандербилта (Bhat K.S., Gene 134(1), 83-87 (1993)). Штамм Е. coli DH5a трансформировали лигированной смесью и трансформированную смесь высевали на пластинки, содержащие канамицин и 5-бром-4-хлориндолил- Плазмиды секвенировали с помощью метода Dye Terminator chemistry (фирма Applied BioSystems, набор PRISM Dye Terminator Cycle Sequencing Ready Reaction Kit, P/N 402078) и праймеров, сконструированных для секвенирования обоех цепей плазмиды. Данные получали на секвенаторах ДНК типа ABI 377. Приблизительно 75% этих реакций дали полезную информацию о последовательности. Последовательности обработывали и объединяли в наборы последовательностей фрагментов с помощью секвенатора Sequencher 3.0 (фирма Gene Codes Corporation), Staden gap4 (Roger Staden, адрес электронной почты: rs@mrc-lmb.cam.ac.uk) и PHRED (Phil Green, адрес электронной почты: phg@u-washington.edu). Наибольший набор последовательностей (приблизительно 76 т.п.н.) перекрывал комплементирующую область до средней глубины 7 независимых сигналов/основание. С помощью анализа комплементации выявляли характеризующуюся перекрыванием космид Е1 и D7 область размером приблизительно 9,9 т.п.н., которая содержит nim1-область. Создавали праймеры, фланкирующие инсерционный сайт вектора и специфичные для Каракаса космиды, с помощью программного обеспечения Oligo 5.0 Primer Analysis Software (фирма National Biosciences, Inc.). ДНК выделяли из космид D7 и E1 с помощью модифицированного метода с использованием ацетата аммония (Traynor P.L., 1990 BioTechniques 9(6): 676.) Эту ДНК непосредственно секвенировали с помощью указанного выше метода Dye Terminator chemistry. Полученная последовательность позволила определить концы комплементирующей области. Также конструировали усеченную версию BamHI-EcoRV-фрагмента, получая конструкцию, которая не содержит ни одной генной области 3 “Gene 3” (фиг.13). Следующий подход был необходим из-за присутствия сайтов HindIII в Bam-Spe-области ДНК. Конструкцию BamHI-EcoRV полностью расщепляли с помощью Spel, а затем для проведения двойного расщепления разделяли на две различные реакционные смеси. Одну аликвоту расщепляли с помощью BamHI, a другую с помощью HindIII. BamHI-SpeI-фрагмент длиной 2816 пар оснований и HindIII-SpeI-фрагмент длиной 1588 пар оснований выделяли из агарозных гелей (набор для экстракции типа QiaQuick Gel) и лидировали с расщепленной с помощью BamHI-HindIII плазмидой pSGCG01. DH5a трансформировали лигированной смесью. Образовавшиеся колонии подвергали скринингу в отношении правильности вставки путем расщепления HindIII с последующим получением ДНК с помощью Wizard Magic MiniPreps (фирма Promega). Клон, содержащий правильную конструкцию, встраивали с помощью электропорации в штамм Agrobacterium GV3101 с целью трансформации растений Arabidopsis. Пример 8: Идентификация области гена NIM1 аллельным секвенированием 1. Генетический анализ С целью определения доминантности различных мутантов, проявляющих фенотип nim1, пыльцу растений дикого типа переносили на рыльце мутантов nim1-1, -2, -3, -4, -5, -6. Если мутация является доминантной, то фенотип nim1 должен проявляться в образовавшихся растениях поколения F1. Если мутация является рецессивной, то образовавшиеся растения F1 должны проявлять фенотип дикого типа. Данные, представленные в таблице 17, свидетельствуют о том, что когда nim1-1, -2, -3, -4 и -6 скрещивали с диким типом, образовавшиеся растения F1 проявляли фенотип дикого типа. Таким образом, эти мутации являются рецессивными. В отличие от этого все потомство F1, полученное в результате скрещивания nim1-5 × дикий тип, проявляло фенотип nim1, что свидетельствует о доминантности этой мутации. После обработки ИНК никакой споруляции Р.parasitica не обнаружено на растениях дикого типа, в то время как на растениях F1 наблюдался рост и определенная споруляция Р.parasitica. Однако фенотип nim1 у этих растений F1 был менее строгим по сравнению с тем, который наблюдался, когда мутация nim1-5 была гомозиготной. С целью определения аллелизма пыльцу с устойчивых к канамицину nim1-1-мутантных растений переносили на рыльце nim1-2, -3, -4, -5, -6. Для подтверждения их гибридного происхождения семени; семена, полученные в результате этого скрещивания, высаживали в планшеты со средой Мурасига и Скуга В5, содержащей канамицин в концентрации 25 мкг/мл. Устойчивые к канамицину (F1) растения переносили в почву и исследовали в отношении фенотипа nim1. Поскольку потомство F1, полученное в результате скрещивания мутанта nim1-5 с диким типом Ws, проявляло фенотип nim1, был также проведен анализ F2, полученного от скрещивания nim1-5 × nim1-1. Как видно из таблицы 17, все образовавшиеся растения F1 проявляли фенотип nim1-1. Таким образом, мутации nim1-2, -3, -4, -5, -6 не полностью восстанавливались с помощью nim1-1; все эти растения входили в одну группу комплементации и, следовательно, были аллельными. Анализ потомства F2, полученного от скрещивания nim1-5 × nim1-1 показал, что оно также проявляет фенотип nim1, подтверждая, что nim1-5 представляет собой аллель nim1. 2. Анализ последовательности и субклонирование NIM1-области Область длиной 9,9 т.п.н., содержащую NIM1-область, анализировали на наличие открытых рамок считывания во всех шести рамках, используя Sequencher 3.0 и пакет программ GCG. В качестве возможных генов было выявлено 4 области, содержащие большие ОРС (генные области 1-4). Эти 4 области амплифицировали с помощью ПЦР из ДНК родителя дикого типа и шести различных nim1-аллельных вариантов. Праймеры для этих амплификации были отобраны с помощью программы Oligo 5.0 (фирма National Biosciences, Inc.) и синтезированы фирмой Integrated DNA Technologies, Inc. ПЦР-продукты разделяли на 1,0%-ных агарозных гелях и очищали с помощью набора для экстракции гелей типа QIAquick. Очищенные геномные ПЦР-продукты непосредственно секвенировали, используя праймеры, применяемые для начальной амплификации, с дополнительными праймерами, предназначенными для секвенирования любых областей, которые не охватывались праймерами начальной амплификации. Средний охват этих генных областей составлял приблизительно 3,5 показаний/основание. Последовательности обрабатывали и объединяли, используя Sequencher 3.0. Замены оснований, специфичные для различных аллелей nim1, были выявлены только в области, обозначенной как генная область 2. Положения, приведенные в таблице 18, соответствуют фиг.14 и соответствуют верхней цепи области длиной 9,9 т.п.н., изображенной на фиг.13. Секвенировали открытые рамки считывания генных областей, обозначенных на фиг.13 как 1, 2, 3 и 4, при этом замены в различных аллелях nim1 приведены в таблице. В соответствии с фиг.13 описанные замены находятся на верхней цепи в направлении 5’ Очевидно, что ген NIM1 был клонирован и что он лежит внутри генной области 2, т.к. обнаружены аминокислотные замены или изменения последовательности внутри открытой рамки считывания генной области 2 во всех шести аллелях nim1. В то же время по крайней мере для одного из аллелей nim1 не выявлено никаких изменений в открытых рамках считывания в генных областях 1, 3 и 4. Следовательно, только один ген внутри области длиной 9,9 т.п.н., которая может представлять собой NIM1, представляет собой генную область 2, ген NIM1. Раздел, посвященный в таблице 18 экотипу Ws, свидетельствует о заменах в экотипе Arabidopsis Ws относительно экотипа Arabidopsis Columbia. Фиг.13, 14, 15 и другие, на которых приведены последовательности, относятся к экотипу Arabidopsis Columbia, который в проведенных экспериментах содержал ген дикого типа. Приведенные замены представляют собой аминокислотные замены в генной области 2 или в NIM1-области, и они показаны как замены в парах оснований в других областях. На фиг.13 приведены 4 различные панели, описывающие клонирование гена NIM1 и полную область длиной 9,9 т.п.н. На фиг.14 приведена последовательность полной области длиной 9,9 т.п.н. в такой же ориентации, что и на фиг.13. На фиг.15 приведена последовательность области, специфичной для гена NIM1, что соответствует генной области 2 на фиг.13; последовательность на фиг.15 содержит ген NIM1. На фиг.15 в однобуквенном коде приведена аминокислотная последовательность и показана полноразмерная; кДНК и RACE-продукт, который обозначен заглавными буквами в последовательности ДНК. Некоторые из аллельных мутаций, которые были обнаружены, приведены над последовательностью ДНК и частично обозначен nim1-аллель, который имел эту замену. Анализ последовательности области и секвенирование различных nim1-аллелей (см. ниже) позволил идентифицировать космидную область, которая содержит ген nim1. Эту область делинеаризировали, получая BamH1-EcoRV-рестрикционный фрагмент длиной ~5,3 т.п.н. Космидную ДНК из D7 и плазамидную ДНК из pBlueScriptI(pBSII) расщепляли с помощью BamHI и EcoRV (NEB). Фрагмент длиной 5.3 т.п.н. из D7 выделяли из агарозных гелей и очищали с помощью набора для экстракции гелей типа QIAquick (№28796, фирма Qiagen). Фрагмент лигировали в течение ночи с расщепленной с помощью Bam-EcoRV плазмидой pBSII и лигированной смесью трансформировали штамм Е. coli DH5a. Отбирали колонии, содержащие вставку, выделяли ДНК и ее строение подтверждали расщеплением с помощью HindIII. Затем для трансформации Arabidopsis конструировали Bam-EcoRV-фрагмент в бинарном векторе (pSGCG01). 3. Анализ методом Нозерн-блоттинга четырех генных областей Идентичные Нозерн-блоты создавали из образцов РНК, выделенных из обработанных водой, СК, БТК и ИНК линий Ws и nim1, как описано ранее (Delaney и др., 1995, PNAS 92, 6602-6606). Эти блоты гибридизовали с ПЦР-продуктами, полученными из четырех генных областей, выявленных в области гена NIM1 длиной 9,9 т.п.н. Только для одной генной области, содержащей ген NIM1 (генная область 2), выявлена гибридизация с образцами РНК, свидетельствуя о том, что только NIM1-область содержит транскрибируемый ген, который можно обнаружить (фиг.16 и таблица 18). В таблице 18 показана вариабельность последовательности nim1-аллеля.
Положения, указанные в таблице, соответствуют фиг.14, на которой приведена последовательность длиной 9,9 т.п.н. Все аллели с nim1-1 по nim1-6 соответствтуют штамму WS. Columbia-0 обозначает дикий тип С помощью проведения дополнительных экспериментов по комплементации также было показано, что генная область 2 (фиг.13) содержит функционально активный ген NIM1. BamHI/HindIII-фрагмент геномной ДНК, содержащий генную область 2, выделяли из космиды D7 и клонировали в бинарном векторе pSGCG01, содержащем ген, определяющий устойчивость к канамицину (фиг.13; Steve Goff, персональное сообщение). Образовавшейся плазмидой трансформировали штамм Agrobacterium GV3101 и позитивные колонии отбирали на среде с канамицином. Для подтверждения того, что отобранная колония содержала плазмиду, использовали ПЦР. Чувствительные к канамицину растения nim1-1 заражали этими бактериями, как описано ранее. Образовавшиеся семена собирали и высевали на GM-arap, содержащий 50 мкг/мл канамицина. Отобранные выжившие растения переносили в почву и тестировали в отношении комплементации. Трансформированные растения и контрольные Ws- и nim1-растения опрыскивали 300 мкМ ИНК. Через 2 дня листья собирали для экстракции РНК и анализа экспрессии генов PR-1. Затем растения опрыскивали Peronospora parasitica (изолят EmWa) и выращивали, как описано ранее. Через 10 дней после заражения грибом растения исследовали и оценивали в зависимости от роста гриба как позитивные или негативные. Все 15 трансформированных растений, а также контрольные Ws-растения отказались негативными в отношении роста гриба после обработки ИНК, в то время как контрольные nim1-растения оказались позитивными в отношении роста гриба. РНК экстрагировали из этих трансформантов и из контрольных растений и анализировали, как описано ранее. У контрольных Ws-растений и у всех 15 трансформантов после обработки ИНК обнаружена индукция гена PR-1, а у контрольных nim1-растениий после обработки ИНК не выявлено индукции PR-1. 4. Выделение кДНК NIM1 Библиотеку кДНК Arabidopsis, созданную в векторе экспрессии 1YES (Elledge и др., 1991, PNAS 88, 1731-1735), высевали и полученные бляшки переносили на фильтры. Фильтры гибридизовали с меченным с помощью 32Р-ПЦР-продуктом, полученным из генной области, содержащей nim-1. В результате скрининга приблизительно 150000 бляшек было выявлено 14 позитивных клонов. Каждую бляшку очищали и выделяли плазмидную ДНК. кДНК-вставки отщепляли от вектора с помощью EcoRI, очищали на агарозном геле и секвенировали. Последовательность, полученная из наиболее длиной кДНК, приведена на фиг.15. Для подтверждения того, что был получен 5’-конец кДНК, использовали набор Gibco BRL 5’ RACE, следуя инструкциям производителя. Образовавшиеся RACE-продукты секвенировали и обнаружили, что они включают дополнительные основания, приведенные на фиг.15. Транскрибируемая область присутствует в обоих клонах кДНК и обнаружена в RACE-продукте, и она приведена заглавными буквами на фиг.15. Замены в аллелях указаны над цепью ДНК. Заглавные буквы обозначают последовательность, присутствующую в клоне кДНК или обнаруженную после ПЦР с RACE. Пример 9: Характеристика гена NIM1 Конструировали множественный ряд последовательностей, используя программное обеспечение Clustal V (Higgins Desmond G. и Paul M, Sharp (1989), Fast and sensitive multiple sequence alignments on a microcomputer, CABIOS 5: 151-153) as part of the DNA* (1228 South Park Street, Madison Wisconsin, 53715) Lasergene Biocomputing Software package for the Macintosh (1994). Было установлено, что аминокислотная последовательность определенных областей протеина NIM1 гомологична аминокислотной последовательности четырех различных протеиновых продуктов кДНК риса. Гомологи были идентифицированы с помощью последовательностей NIM1, обнаруженных в GenBank BLAST. Сравнение обладающих гомологией областей для продуктов NIM1 и кДНК риса приведено на фиг.19. Показаны фрагменты протеина NIM1, имеющие от 36 до 48% аминокислотных последовательностей, идентичных аминокислотным последовательностям четырех продуктов риса. Пример 10: Фенотипическая характеристика различных аллелей nim1 1. Анализ химической реактивности в аллелях nim1 Проанализированы различия между различными аллелями nim1 с точки зрения химической индукции экспрессии гена PR и устойчивости к Peronospora parasitica (см. фиг.17 и 18). Мутантные растения обрабатывали химическими индукторами и затем анализировали в отношении экспрессии гена PR и устойчивости к болезням. 2. Рост растений и химическая обработка Семена дикого типа и семена, полученные от растений, имеющих каждый из алелей nim1 (nim1-1, -2, -3, -4, -5, -6), высевали в среду для выращивания типа MetroMix 300, закрывали прозрачным пластиковым куполом и выдерживали в темноте при температуре 4°С в течение 3 дней. После выдерживания при 4°С в течение 3-х дней растения переносили на 2 недели в фитотрон. Приблизительно через 2 недели после посева проросшие ростки дали 4 настоящих листа. После этого растения обрабатывали Н2О, 5 мМ СК, 300 мкМ БТК или 300 мкМ ИНК. Химические соединения вносили в виде легкого тумана до полного покрытия листьев с помощью туманообразователя типа “chromister”. Обработанные водой контрольные растения возвращали в ростовую климатическую камеру, а обработанные химическими соединениями растения держали в отдельных, но идентичных климатических камерах. Через 3 дня растения разделяли на 2 группы. Из одной группы экстрагировали и анализировали РНК. Вторую группу заражали Р. parasitica. 3. Инокуляция Peronospora parasitica и анализ Изолят Р. parasitica “EmWa” представляет собой изолят Р.р., совместимый с экотипом Ws. Совместимые изоляты представляют собой изоляты, способные вызывать болезнь у определенного хозяина. Изолят Р. parasitica “NoCo” несовместим с экотипом Ws, но совместим с экотипом Columbia. Несовместимые патогены распознаются потенциальным хозяином, вызывая у хозяина ответ, который предупреждает развитие болезни. На 3-й день после обработки водой или обработки химическим соединениями растения заражали совместимым изолятом “EmWa”. Изолятом “NoCo” инокулировали только обработанные водой растения. После инокуляции растения закрывали прозрачным пластиковым колпаком для поддержания высокой влажности, необходимой для успешного заражения Р. parasitica, и помещали в ростовую камеру с дневной температурой 19°С/ночной температурой 17°С и при световом цикле 8 ч света/16 ч темноты. Через различные промежутки времени после инокуляции растения анализировали под микроскопом для оценки развития симптомов. Под увеличением споруляция грибов может быть обнаружена на очень ранних стадиях развития болезни. В каждом горшке определяли процент растений, у которых обнаружена споруляция на 5, 6, 7, 11 и 14 день после инокуляции, а также оценивали плотность споруляции. На фиг.18 приведена оценка развития болезни у растений с различными аллелями nim1 после инокуляции Р.parasitica. Наиболее характерными точками были 5-й и 6-й день после инокуляции. На 5-й день после инокуляции у растений nim1-4 после проведения всех обработок индуцирующими химическими веществами обнаружено ~80%-ное заражение, ясно указывая на то, что этот аллель/генотип является наиболее чувствительным к болезни. На 6-й день после инокуляции у растений nim1, -2, -3, -4 и -6 после всех обработок индуцирующими химическими веществами обнаружена значительная частота заболеваемости. Однако у растений nim1-5 на 6-й день после всех обработок обнаружена меньшая зараженность, чем у растений дикого типа Ws. Следовательно, nim1-5 представляет собой аллель, наиболее устойчивый к болезням по сравнению с различными аллелями nim1. Вероятно, nim1-2 является промежуточным в отношении чувствительности к болезням после обработки БТК, но не после обработок другими индуцирующими агентами. Экспрессия гена PR-1 свидетельствует о том, что nim1-4 является наименее чувствительным ко всем из изученных индуцирующих химических веществ (фиг.17), а nim1-5 проявляет повышенные уровни экспрессии PR-1 в отсутствии индукторов. Эти результаты по экспрессии гена PR-1 согласуются с оценкой заболеваемости, которую проводили с использованием Р.parasitica (фиг.18), и свидетельствуют о том, что аллели nim1 могут обусловливать устойчивость или чувствительность. Полученные выше образцы использовали для анализа экспрессии гена NIM1 (фиг.17). У растений дикого типа мРНК NIM1 присутствовала в необработанных контрольных образцах. После обработки СК, ИНК, БТК или заражения совместимым патогеном мРНК NIM1 накапливалась до высоких уровней. У растений с аллелями nim1 по сравнению с диким типом наблюдалось различие в количестве транскриптов NIM1 (мРНК). Количество копий мРНК NIM1 у необработанных мутантных растений оказалось ниже, чем обнаруженное в диком типе, за исключением nim1-2 и -5, где количество копий оказалось близким. Растения с мутациями nim1-1, -3 и -4 имели меньшее количество транскриптов NIM1, а у растений с мутаций nim1-6 обнаружено очень небольшое накопление мРНК NIM1. Увеличение количества копий мРНК NIM1 после обработки СК, ИНК или БТК наблюдали у растений nim1-1, -2, -3, но не наблюдали у растений nim1-5 или -6. Однако такое увеличение оказалось меньшим, чем обнаруженное в растениях дикого типа. После патогенного заражения дополнительные полосы гибридизации с кДНК-зондом NIM1 обнаружены как в растениях дикого типа, так и в мутантных растениях, а количество копий мРНК NIM1 оказалось увеличенным по сравнению с необработанными контролями, за исключением nim1-6. На фиг.18 приведены данные оценки устойчивости к болезням, полученные на основе определения заражения растений с различными аллелями nim1, a также NahG-растений через различные промежутки времени после инокуляции Peronospora parasitica. WsWT обозначает родительскую линию Ws дикого типа, у которой обнаружены аллели nim1. Различные аллели nim1 указаны в таблице, в которую также включены данные о NahG-растении. Данные о NahG-растении опубликованы ранее (Delaney и др., Science 266, стр. 1247-1250 (1994)). NahG-тип Arabidopsis также описан в WO 95/19443. Ген NahG представляет собой ген из Pseudomonas putida, который превращает салициловую кислоту в катехол, тем самым препятствуя накоплению салициловой кислоты, необходимого сигнального компонента трансдукции SAR в растениях. Так, растения Arabidopsis с геном NahG не обладают нормальной SAR. Кроме того, в целом у них обнаружена существенно более высокая чувствительность к патогенам. Следовательно NahG-растения представляют собой тип универсального чувствительного контроля. Кроме того, NahG-растения еще сохраняют способность реагировать на химические индукторы ИНК и БТК, что показано в двух нижних панелях на фиг.17. Как следует из фиг.18, аллели nim1-4 и nim1-6, вероятно, являются наиболее строгими, что можно легко обнаружить в более ранние периоды времени, как описано выше в разделе “Результаты” в данном описании, а также следует из результатов, полученных с использованием панели EmWa-БТК (самая нижняя панель на чертеже). Кроме того, аллель nim1-5 дает наиболее выраженный ответ и на ИНК и на БТК и, следовательно, представляет собой наиболее слабый аллель nim1. У NahG-растений обнаружена очень хорошая реакция и на ИНК и на БТК, и она очень сходна с реакцией аллеля nim1-5. Однако в более поздние периоды времени, день 11 на чертеже, устойчивость к болезням, индуцированная у NahG-растений, начинает исчезать, свидетельствуя о выраженном различии между ИНК и БТК в том плане, что индуцированная ИНК устойчивость исчезает существенно быстрее и более заметно в сравнении с устойчивостью, индуцированной у NahG-растений с помощью БТК. Также из этих экспериментов видно, что ИНК и БТК индуцируют очень хорошую устойчивость у растений wsk EmWa, a nim1-1, nim1-2 и другие аллели~ *р18 62Xnim1 фактически не реагируют на СК или ИНК в отношении устойчивости к болезням. На фиг.18 указано в процентах количество растений, у которых обнаружена споруляция после заражения штаммом Р. parasitica EmWa, а каждая из гистограмм обозначает количество дней после заражения, в течение которых определяли устойчивость к болезням. Нозерн-анализ экспрессии одного из генов SAR PR1 также проводили на этих же образцах, как показно на фиг.17. На фиг.17 показано, что растения дикого типа дают очень хорошую реакцию на салицилат, ИНК, БТК, а также на заражение патогеном, что ясно видно по увеличенной экспрессии гена PR1. С другой стороны, растения с аллелем nim1-1 проявляют только ограниченную реакцию на все химические индукторы, включая патоген. Индукция патогеном по крайней мере в несколько раз ниже для аллеля nim1-1, чем для дикого типа. Для аллелей nim1-2, nim1-3 и nim1-6 обнаружена реакция на различные обработки, аналогичная реакции аллеля nim1-1. Однако для аллеля nim1-4 обнаружено отсутствие экспрессии в ответ на любой из использованных индукторов. По существу вся обнаруженная экспрессия представляла собой фоновый уровень. Для аллеля nim1-5 обнаружен очень высокий фоновый уровень по сравнению с обработанными водой контролями, и этот фоновый уровень сохранялся при всех обработках; однако наблюдалась ограниченная индукция или отсутствие индукции под воздействием химических индукторов. NahG-растения служат достаточно наглядным контролем в связи с тем, что они не способны индуцировать PR-1 в присутствии СК; с другой стороны и ИНК и БТК индуцируют очень высокий уровень экспрессии PR-1. Заражение патогеном аналогично воздействию СК; в обработанных EmWa NahG-растениях не наблюдается экспрессии PR-1. Те же самые образцы РНК, которые были получены при исследовании индукции, также зондировали геном NIM1, используя полноразмерный клон кДНК в качестве зонда. На фиг.16 можно увидеть, что ИНК индуцирует ген NIM1 в растении Ws с аллелем дикого типа. Однако мутантный аллель nim1-1 проявляет более низкий основной уровень экспрессии гена NIM1 и не способен индицироваться ИНК. Это сходно с тем, что было получено для аллеля nim1-3 и аллеля nim1-6. Аллель nim1-2 проявляет приблизительно нормальные уровни в необработанном образце и проявляет индукцию, аналогичную индукции образца дикого типа, равно как и аллель nim1-4. Аллель nim1-5, по-видимому, проявляет более высокий основной уровень экспрессии гена NIM1 и более выраженную экспрессию при индукции химическими индукторами. Индукция NIM1 химическими индукторами устойчивости и другими индукторами согласуется с его ролью в защите от патогенов, а также представляет собой дополнительное доказательство того, что согласно изобретению обнаружен требуемый ген в области длиной 9,9 т.п.н.
Формула изобретения
1. Выделенная молекула ДНК, обеспечивающая устойчивость растений к болезням, с ДНК молекулой, включающей NIM1 ген, который кодирует аминокислотную последовательность, представленную в SEQ ID NO: 3. 2. Выделенная молекула ДНК по п.1, включающая нуклеотидную последовательность, представленную в SEQ ID NO: 2. 3. Выделенная молекула ДНК по п.1, длиной приблизительно 9,9 т.п.н., которая кодирует продукт гена NIM1. 4. Выделенная молекула ДНК по п.1, включающая нуклеотидную последовательность, представленную в SEQ ID NO: 1. 5. Выделенная молекула ДНК по п.1, кодирующая аминокислотную последовательность продукта гена NIM1, представленную в SEQ ID NO: 2. 6. Рекомбинантный вектор, включающий активный в растении промотор, функционально связанный с ДНК молекулой по пп.1-5 и обеспечивающий экспрессию протеина, который активирует системную приобретенную устойчивость у растений. 7. Рекомбинантный вектор по п.6, где указанный вектор способен стабильно трансформировать клетку-хозяина. 8. Рекомбинантный вектор по п.6, где указанный вектор способен стабильно трансформировать растение, семена растения, ткань растения или клетку растения. 9. Способ придания устойчивости к болезням растительным клеткам, растениям и их потомству, отличающийся тем, что растительные клетки, растения или их потомство трансформируют молекулой ДНК по любому из пп.1-5.
РИСУНКИ
MM4A – Досрочное прекращение действия патента СССР или патента Российской Федерации на изобретение из-за неуплаты в установленный срок пошлины за поддержание патента в силе
Дата прекращения действия патента: 11.03.2007
Извещение опубликовано: 20.07.2010 БИ: 20/2010
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
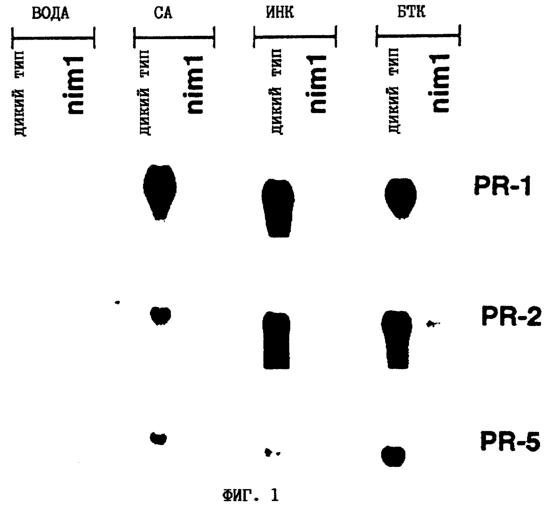
 3’)
3’)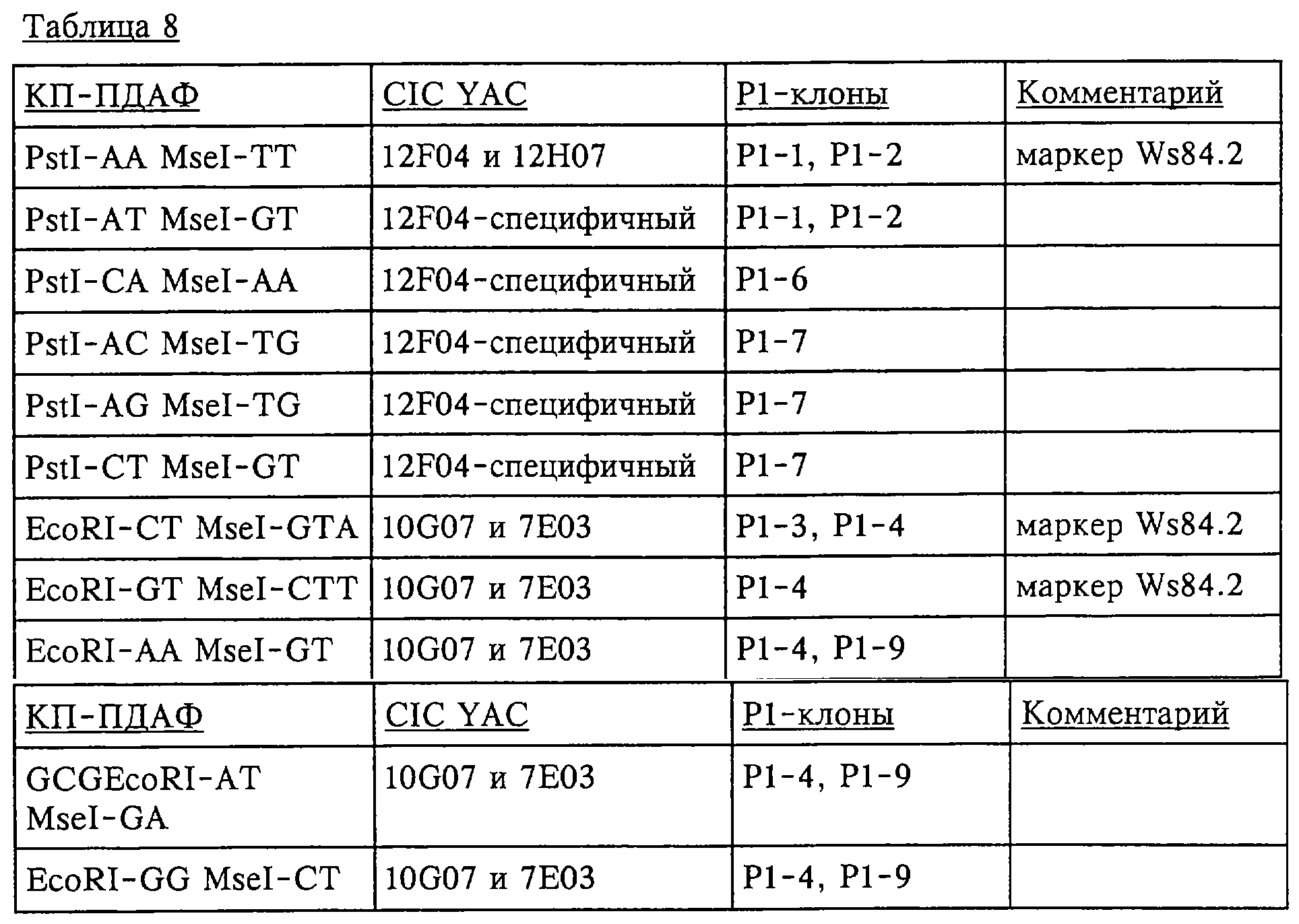
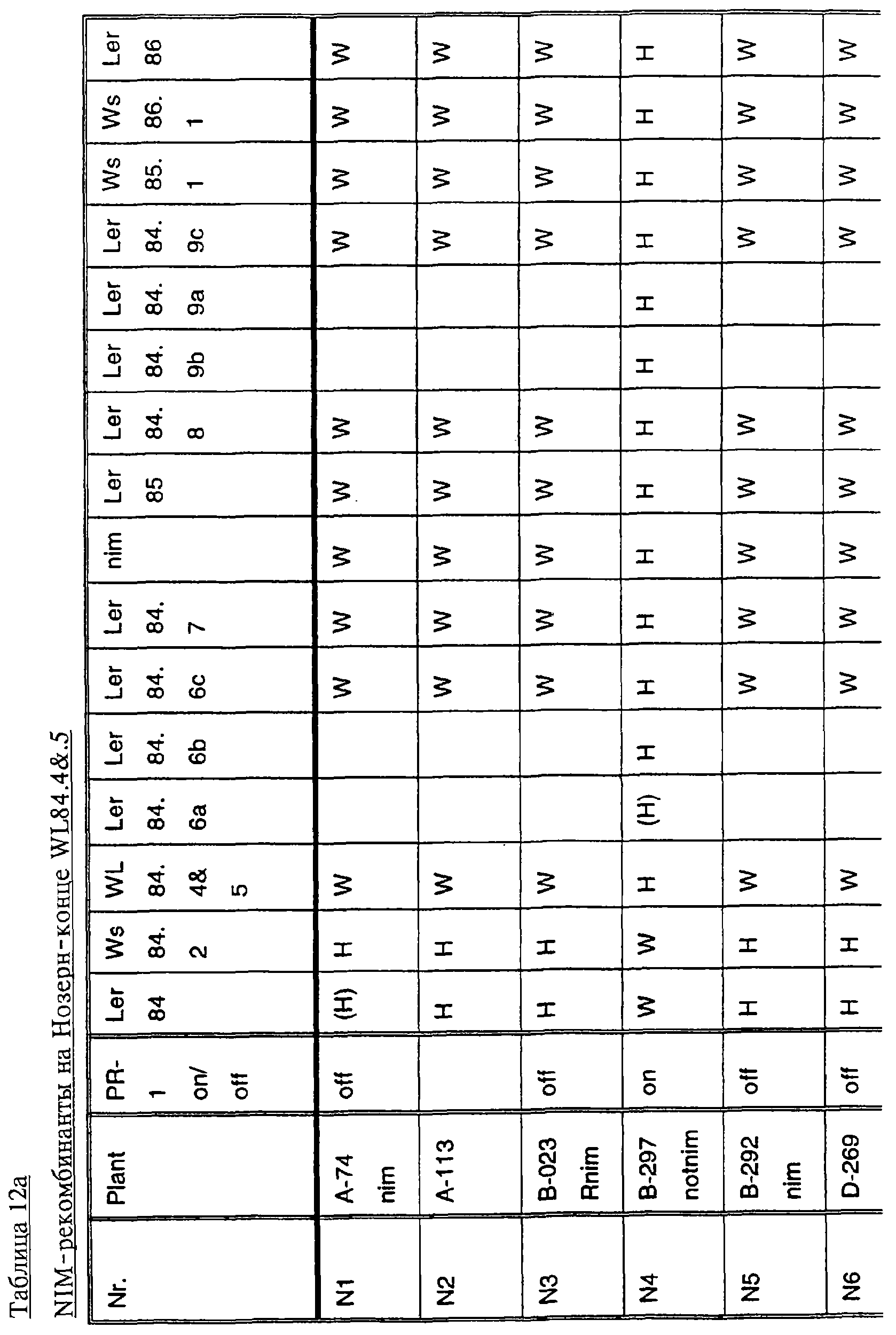
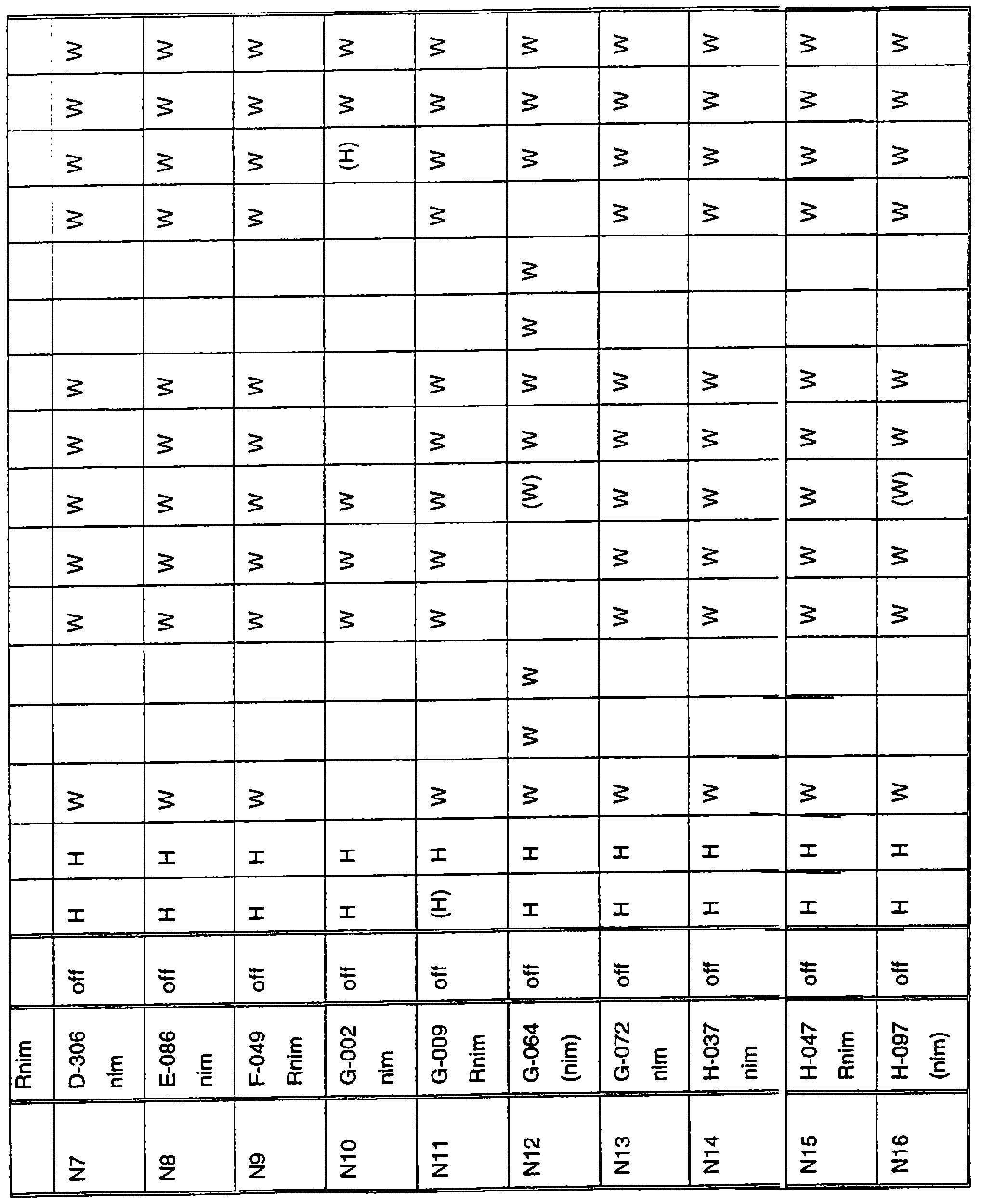
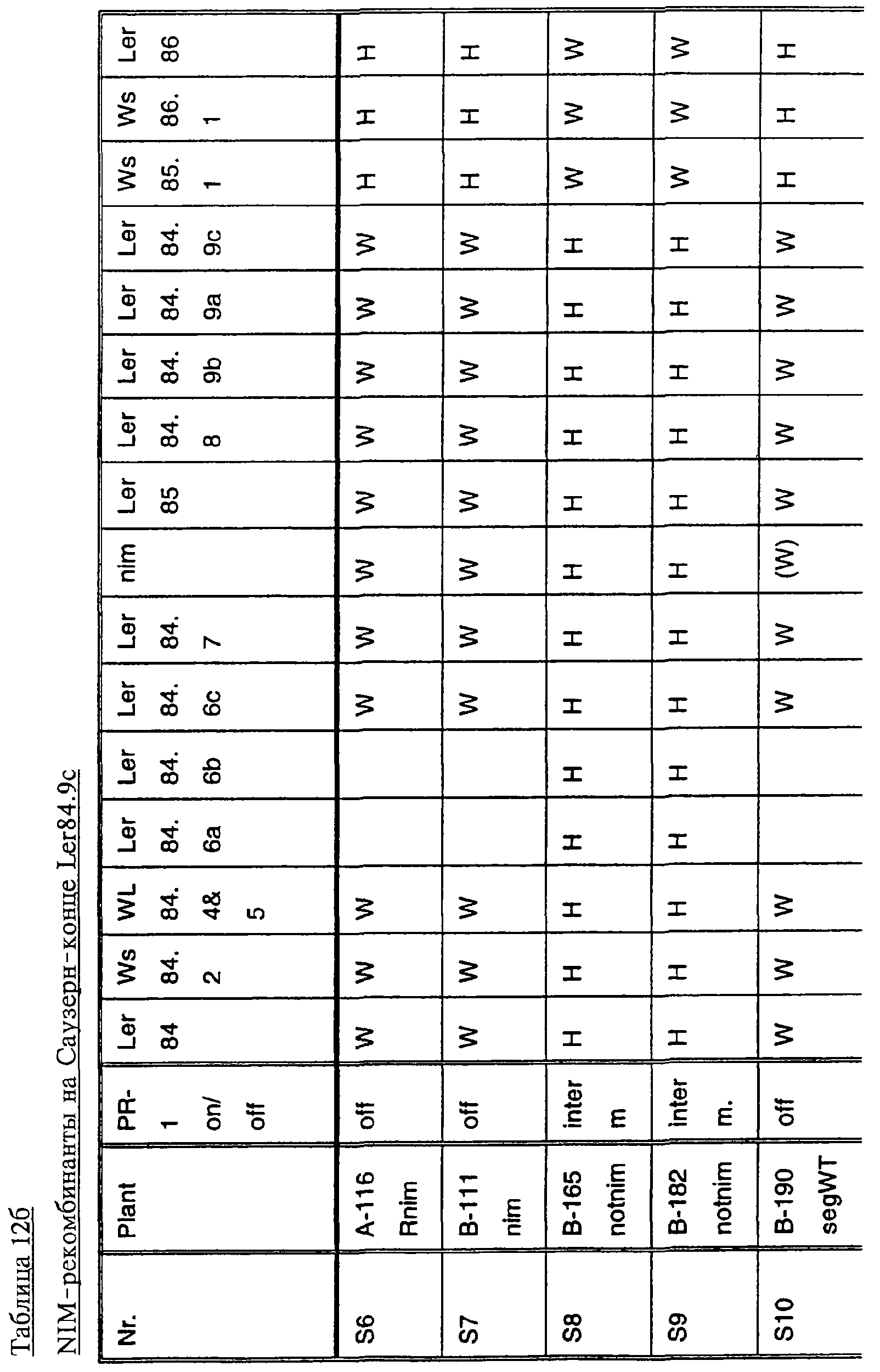
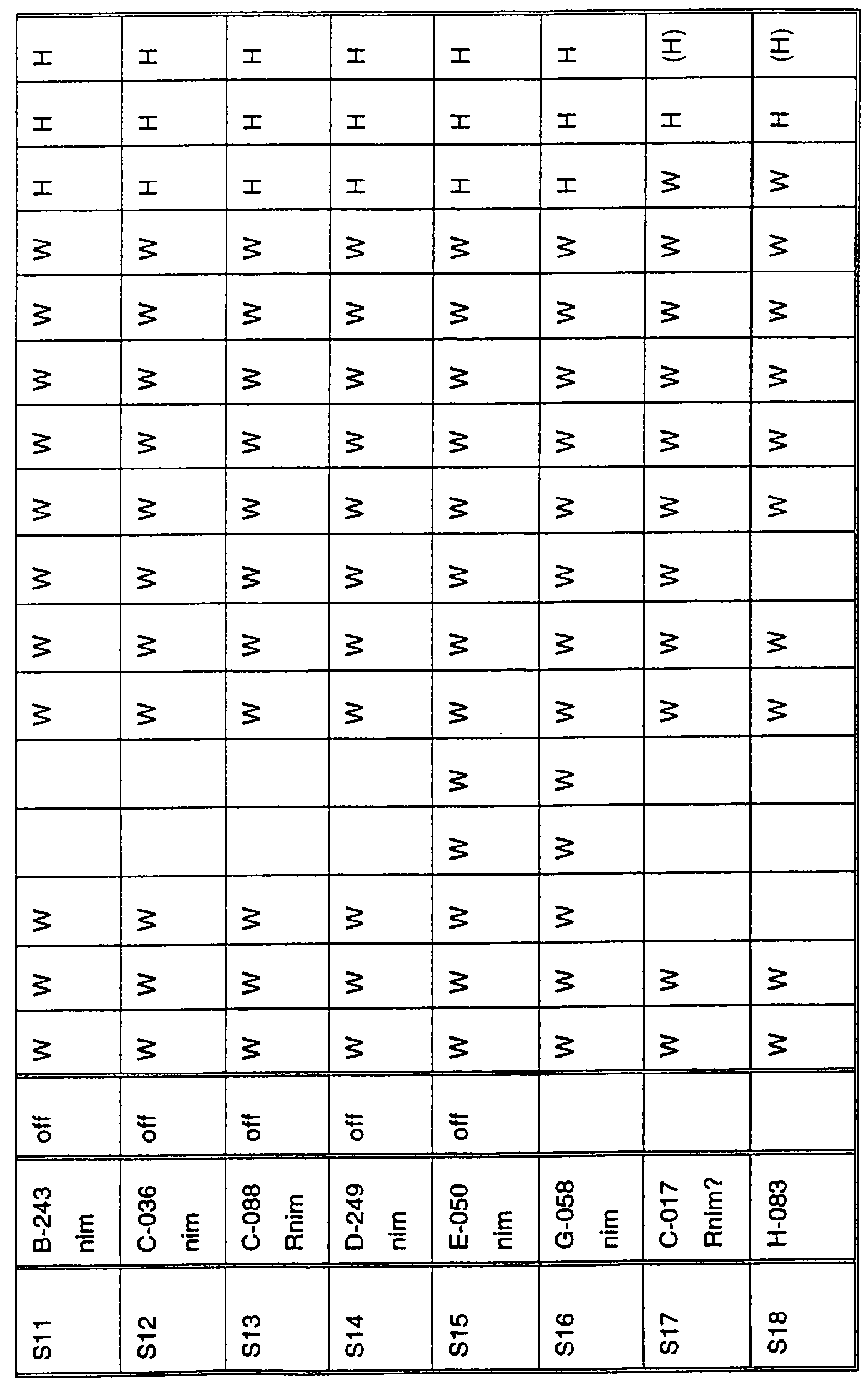
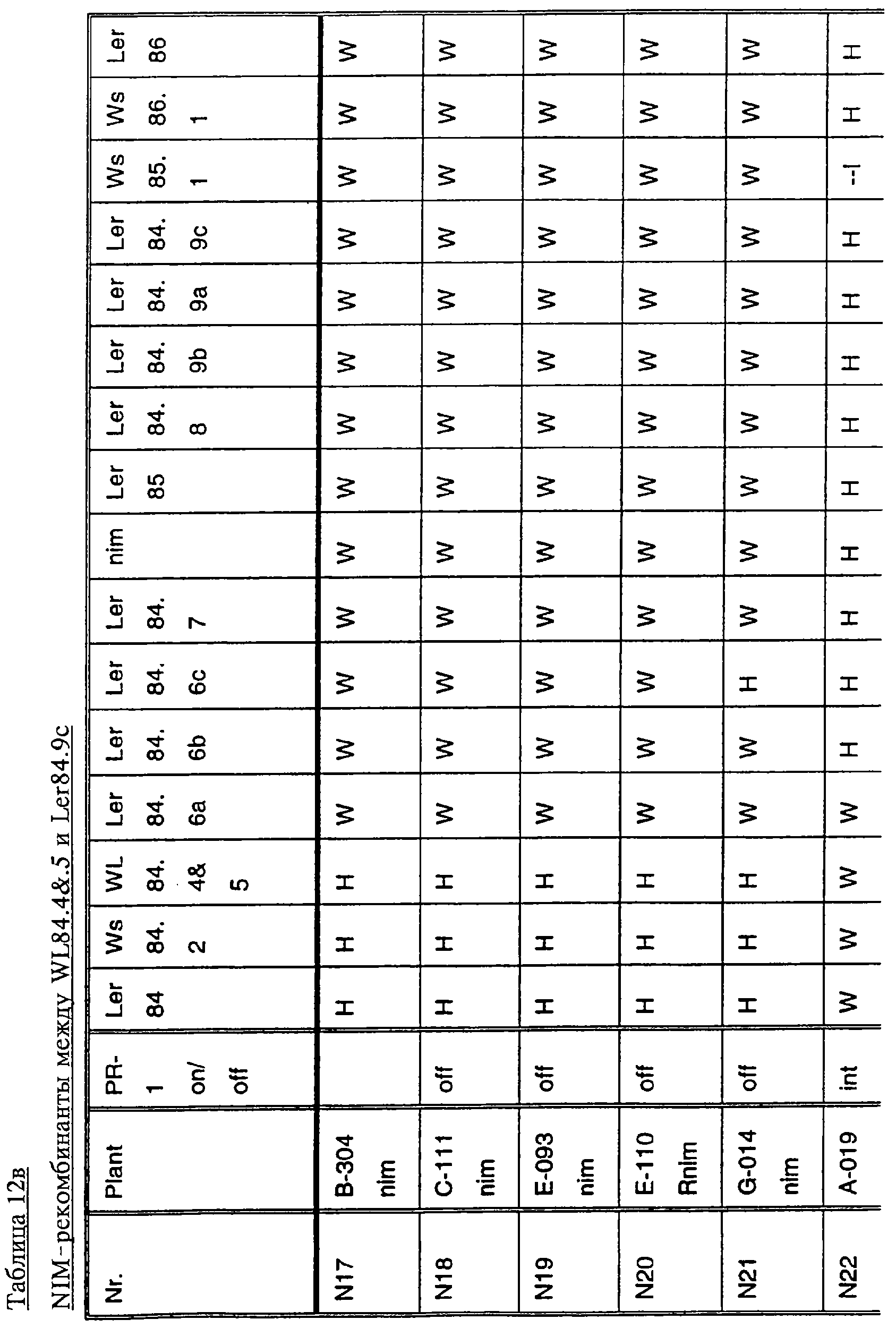

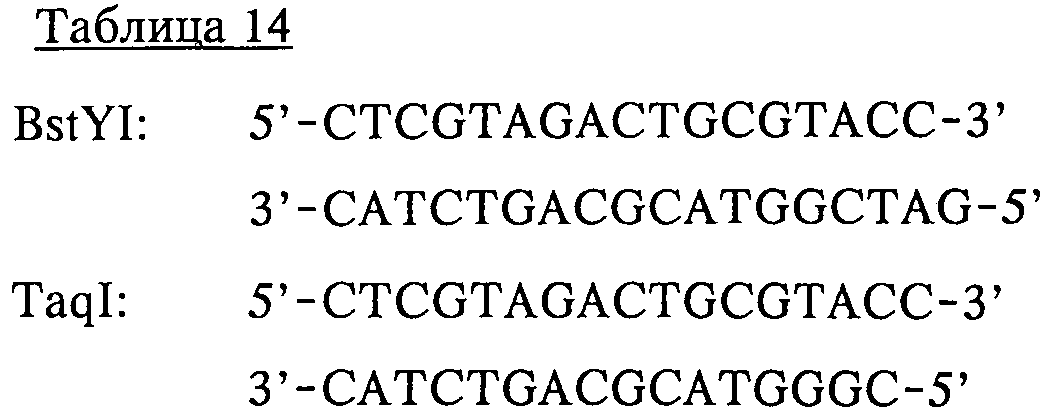
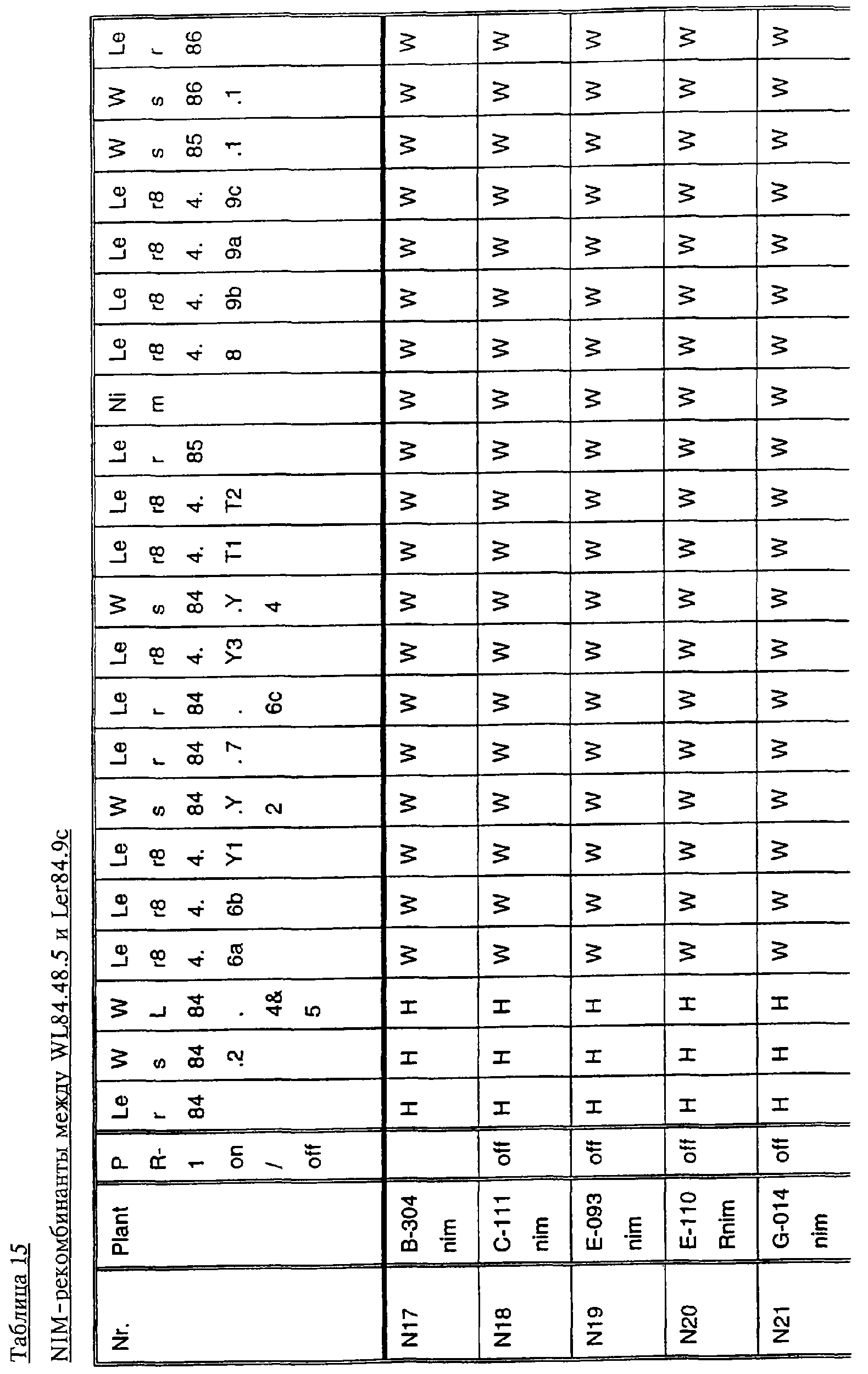

 -D-галактозидазу (X-gal). Для выделения плазмид отбирали 1600 белых или светло-голубых колоний KanR. Отдельные колонии вносили в 96-луночные планшеты с глубокими лунками (фирма Polyfiltronics, №U508), которые содержали 1,5 мл ТВ+Кан (канамицин) (50 мкг/мл). Планшеты покрывали и помещали на 16 ч в шейкер с вращающейся платформой при 37°С. Плазмидную ДНК выделяли, используя систему Wizard Plus 9600 Miniprep system (фирма Promega, №А7000) согласно рекомендациям производителя.
-D-галактозидазу (X-gal). Для выделения плазмид отбирали 1600 белых или светло-голубых колоний KanR. Отдельные колонии вносили в 96-луночные планшеты с глубокими лунками (фирма Polyfiltronics, №U508), которые содержали 1,5 мл ТВ+Кан (канамицин) (50 мкг/мл). Планшеты покрывали и помещали на 16 ч в шейкер с вращающейся платформой при 37°С. Плазмидную ДНК выделяли, используя систему Wizard Plus 9600 Miniprep system (фирма Promega, №А7000) согласно рекомендациям производителя.